

Вопросы ихтиологии, 2021, T. 61, № 5, стр. 545-552
Новые данные о распространении трёх филогенетических линий арктического гольца Salvelinus alpinus (Salmonidae) в областях их контакта на севере Восточной Сибири
Н. В. Гордеева 1, С. С. Алексеев 2, *, А. Ф. Кириллов 3, В. И. Романов 4, М. Ю. Пичугин 2, 5
1 Институт общей генетики РАН – ИОГен РАН
Москва, Россия
2 Институт биологии развития РАН – ИБР РАН
Москва, Россия
3 Якутский филиал Всероссийского научно-исследовательского института
рыбного хозяйства и океанографии − ЯкутскНИРО
Якутск, Россия
4 Томский государственный университет
Томск, Россия
5 Московский государственный университет
Москва, Россия
* E-mail: alekseyev@mail.ru
Поступила в редакцию 27.07.2020
После доработки 10.11.2020
Принята к публикации 11.11.2020
Аннотация
С целью изучить распространение выделенных по митохондриальной ДНК филогенетических линий арктического гольца Salvelinus alpinus в области их контакта на севере Восточной Сибири проанализированы нуклеотидные последовательности контрольной области мтДНК гольцов из десяти заполярных популяций на участке от Енисея до Лены. На плато Путорана отмечены гаплотипы атлантической и сибирской подгрупп евроазиатской филогенетической группы, в бассейне р. Хатанга – атлантической, в дельте Лены − сибирской подгруппы и берингийской группы. На Путоране и в дельте Лены найдены гаплотипы сибирской подгруппы, ранее отмечавшиеся в других районах Восточной Сибири. Новые данные вкупе с литературными позволяют уточнить ареалы трёх филогенетических линий и границы областей их перекрывания, а также свидетельствуют о широкой симпатрии гаплотипов атлантической и сибирской подгрупп в водоёмах Таймырского района и подтверждают близкое родство гольцов сибирской подгруппы из всех основных областей её распространения.
У арктического гольца Salvelinus alpinus и близких к нему видов/форм выделены пять филогенетических групп гаплотипов контрольной области митохондриальной ДНК (мтДНК) – атлантическая, сибирская, акадийская, арктическая и берингийская (Brunner et al., 2001). Первые две группы мы объединяем в евроазиатскую группу и рассматриваем в качестве её подгрупп (Гордеева и др., 2018). Гольцы с гаплотипами этих групп/подгрупп имеют викарирующие частично перекрывающиеся ареалы (Brunner et al., 2001; Alekseyev et al., 2009; Moore et al., 2015; Osinov et al., 2017; Гордеева и др., 2018; Salisbury et al., 2019). Области их перекрывания представляют особый интерес как зоны послеледниковых вторичных контактов и гибридизации этих плейстоценовых линий (Osinov et al., 2015, 2017). Были изучены зоны контакта берингийской и арктической групп на северо-западе Северной Америки (Moore et al., 2015) и на северо-востоке Евразии (Esin et al., 2017; Олейник и др., 2017; Осинов и др., 2018); арктической, атлантической и акадийской на Лабрадоре и Ньюфаундленде (Moore et al., 2015; Salisbury et al., 2019). На севере Восточной Сибири известны области вторичного контакта атлантической и сибирской, сибирской и берингийской групп/подгрупп (Радченко, 2003, 2004; Alekseyev et al., 2009; Osinov et al., 2015, 2017; Гордеева и др., 2018). Генетические исследования гольцов из этих труднодоступных областей основываются на небольшом числе особей и популяций и нуждаются в значительном расширении.
Согласно первоначальному описанию (Brunner et al., 2001), атлантическая группа распространена в Евразии на восток до Кольского п-ова, а сибирская − на запад до Финляндии и Шпицбергена. Столь западное положение границы между ареалами групп было обусловлено тем, что авторы включали во вторую несколько описанных ими гаплотипов, которые позже были переведены в первую группу (Гордеева и др., 2018). С учётом этого перевода и новых данных уточнённый ареал сибирской подгруппы ограничен континентальными водоёмами Восточной Сибири; его западной границей на сегодняшний день является плато Путорана (Brunner et al., 2001; Osinov et al., 2017), а восточной границей ареала атлантической − бассейн Хатангской губы (Brunner et al., 2001; Alekseyev et al., 2009). В промежутке между Хатангой и Кольским п-овом гольцы атлантической подгруппы найдены в бассейне Пясины, в том числе на Путоране (Alekseyev et al., 2009; Moore et al., 2015; Osinov et al., 2017; Гордеева и др., 2018), на Полярном Урале и на Новой Земле (Гордеева и др., 2018).
В трёх водоёмах Таймырского Долгано-Ненецкого района у гольцов обнаружены гаплотипы обеих филогенетических линий: в оз. Лама (по данным рестрикционного анализа участка АТФазы6/ND4L: Радченко, 2003; по данным анализа последовательностей гена цитохрома b: Радченко, 2004; Osinov et al., 2015; контрольной области мтДНК: Osinov et al., 2017), в оз. Кунгасалах и в р. Новая (Alekseyev et al., 2009). Всё это показывает, что в Таймырском регионе находится зона вторичного контакта атлантической и сибирской подгрупп (Osinov et al., 2015, 2017), но её точные границы пока не очерчены.
Это же относится к расположенной восточнее, в прибрежных районах от Анабара до Лены, зоне контакта арктических гольцов с гаплотипами сибирской подгруппы и арктических гольцов с интрогрессированным от северной мальмы S. malma malma гаплотипом берингийской группы (Alekseyev et al., 2009; Гордеева и др., 2018). В ней гаплотипы обеих линий встречаются совместно в одном озере в бассейне Оленёкской губы; в бассейне Анабара и в районе дельты Лены они отмечены в разных озёрах. Аналогичные области, в которых происходил захват арктическим гольцом или близким к нему гольцом Таранца мтДНК северной мальмы, обнаружены в Колымско-Охотском регионе (Радченко, 2003, 2004; Alekseyev et al., 2009; Osinov et al., 2017; Осинов и др., 2018), и на Камчатке (Esin et al., 2017).
Цель настоящей работы – привести новые данные о распространении арктических гольцов с мтДНК-гаплотипами трёх филогенетических линий в зонах их вторичных контактов на севере Восточной Сибири для уточнения границ этих зон и расширения сведений о филогеографии вида.
МАТЕРИАЛ И МЕТОДИКА
Материал (39 экз. арктического гольца) собран в десяти заполярных водоёмах Восточной Сибири (таблица; рис. 1а). Он включает сборы авторов в басссейне Хатанги, на плато Путорана (рис. 1б) и в дельте Лены (рис. 1в) в 1991 и 2018−2020 гг., а также пробы из последних двух районов, собранные экспедициями кафедры ихтиологии МГУ в 1975−1976 гг. и при участии второго и последнего авторов в 1979 г.
Рис. 1.
Распространение трёх филогенетических линий гаплотипов контрольной области мтДНК арктического
гольца Salvelinus alpinus на севере Сибири: а − карта-схема региона с ареалами групп и подгрупп гаплотипов
мтДНК; б, в − спутниковые снимки западной части плато Путорана (б) и участка Оленёкской
протоки дельты Лены (в). Местоположение популяций, в которых проводилось изучение
мтДНК: 1−10 – наши данные; B – Brunner et al., 2001; A – Alekseyev et al., 2009; M – Moore et
al., 2015; O – Оsinov et al., 2017, G – Гордеева и др., 2018); ( ) и (
) и ( ) − атлантическая подгруппа евроазиатской группы (включая гаплотипы SIB1-3,5,9); (⚪)
и (
) − атлантическая подгруппа евроазиатской группы (включая гаплотипы SIB1-3,5,9); (⚪)
и ( ) − сибирская подгруппа евроазиатской группы; (⚫) и (
) − сибирская подгруппа евроазиатской группы; (⚫) и ( ) – берингийская группа (арктические гольцы с гаплотипами, интрогрессированными от
северной мальмы); “?” – нет данных. Водоёмы: 1 – оз. Глубокое, 2 – оз. Собачье и р. Хоронен, 3 – оз. Кета, 4 – оз. Хантайское, 5 – оз. Аян, 6 – оз. Баяджяга, 7 – оз. Дальнее, 8 – оз. Переходное, 9 – оз. Булкурка-2, 10 – оз. Ладаннах-Кюеле (Форелевое, Аранастах). Масштаб: 20 км.
) – берингийская группа (арктические гольцы с гаплотипами, интрогрессированными от
северной мальмы); “?” – нет данных. Водоёмы: 1 – оз. Глубокое, 2 – оз. Собачье и р. Хоронен, 3 – оз. Кета, 4 – оз. Хантайское, 5 – оз. Аян, 6 – оз. Баяджяга, 7 – оз. Дальнее, 8 – оз. Переходное, 9 – оз. Булкурка-2, 10 – оз. Ладаннах-Кюеле (Форелевое, Аранастах). Масштаб: 20 км.

Изученные водоёмы значительно различаются по своим характеристикам (таблица). Три включённых в настоящую работу озера в дельте Лены (Булкурка-2, Дальнее и Переходное), а также ранее обследованные озёра Гусинка и Булкурка (Alekseyev et al., 2009) – небольшие низко расположенные безымянные (названия даны нами и членами экспедиции МГУ) термокарстовые водоёмы, находящиеся на ~50-километровом участке вдоль Оленёкской протоки. В этих озёрах обитают мономорфные популяции арктического гольца, представленные крупной формой. Озеро в отрогах Хараулахского хребта (бассейн зал. Неёлова), на топографических картах обозначаемое как Ладаннах-Кюеле, приводится в работе Савваитовой и Максимова (1980) под местным названием Форелевое. Приведённые в работе Борисова (1932) координаты и описание не оставляют сомнения в том, что это озеро Аранастах, гольца которого он на полвека раньше описал как отдельный вид − якутский голец S. jacuticus. В нём отмечены крупная и мелкая формы гольца (Савваитова, Максимов, 1980), но принадлежность изученного экземпляра к одной из них неизвестна.
Характеристика водоёмов, объём исследованного материала и обнаруженные гаплотипы контрольной области мтДНК у гольца Salvelinus alpinus
| № | Водоём | Район | Бассейн | Координаты | Длина*, км | Ширина*, км | Глубина*, м | Высота над уровнем моря, м | Год | Форма гольца | Гаплотип | n, экз. |
|
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| с.ш. | в.д. | ||||||||||||
| 1 | Оз. Глубокое | П | Глубокая → оз.Мелкое → → Норильская → → оз.Пясино → Пясина | 69°16′ | 90°10′ | 44.5 | 6.0 | 185 | 49 | 1975 | “Боганидская палия” | SIB10 | 1 |
| 2 | Оз. Собачье | П | Муксун → оз.Глубокое → → … → Пясина | 69°02′ | 91°15′ | 46.0 | 3.7 | 162 | 69 | 1991 | Голец Дрягина | ATL32 | 1 |
| Р. Хоронен (приток оз. Собачье) | 69°08′ | 91°55′ | – | – | – | – | 1991 | “Боганидская палия” | SIB20 | 2 | |||
| 3 | Оз. Кета | П | Рыбная → Норильская → → оз.Пясино → Пясина | 68°41′ | 90°26′ | 96.0 | 13.3 | 180 | 93 | 1976 | То же | ATL21(M) | 1 |
| 4 | Оз. Хантайское | П | Хантайка → Енисей | 68°17′ | 90°30′ | 110.0 | 18.0 | 420 | 73 | 2019 | Длиннотычиночный | ATL21(M) | 9 |
| “Тыптушка” | ATL21(M) | 2 | |||||||||||
| 5 | Оз. Аян | П | Аян → Хета → Хатанга | 69°10′ | 94°00′ | 55.0 | 3.2 | 256 | 470 | 2020 | “Гигант” | SIB4 | 2 |
| Озёрно-речной | SIB4 | 1 | |||||||||||
| “Путоранчик” | SIB4 | 7 | |||||||||||
| 6 | Оз. Баяджяга | Х | Попигай → Хатанга | 72°11′ | 110°51′ | 1.1 | 0.9 | – | – | 2018 | Крупная | ATL32 | 5 |
| 7 | Оз. Дальнее | ДЛ | Оленёкская протока | 72°20′45′′ | 125°40′ | 1.0 | 0.6 | ~3 | ~7 | 1979 | То же | BER10 | 1 |
| 8 | Оз. Переходное | ДЛ | То же | 72°20′30′′ | 125°50′ | 0.4 | 0.3 | ~15 | ~6 | 1979 | » | SIB10 | 1 |
| SIB35 | 1 | ||||||||||||
| 9 | Оз. Булкурка-2 | ДЛ | » | 72°18′ | 126°10′ | 1.2 | 1.1 | – | ~5 | 2018 | » | BER10 | 4 |
| 10 | Оз. Ладаннах-Кюеле (Форелевое, Аранастах) | ДЛ | зал. Неёлова | 71°48′ | 128°36′ | 1.2 | 0.7 | 4.5 | 68 | 1979 | Крупная или мелкая | BER24 | 1 |
| Всего | 39 | ||||||||||||
Примечание. Районы: П – плато Путорана, Х – район Хатанги и Хатангской губы, ДЛ – район дельты Лены; *приведены максимальные значения; название гаплотипа ATL21 дано по: Moore et al., 2015 (обозначено буквой M в скобках), он соответствует гаплотипу ATL20 по: Osinov et al., 2017; SIB4 соответствует SIB31 по: Moore et al., 2015 и SIB34 по: Алексеев и др., 2019; n − число особей.
Озёра плато Путорана – крупные глубокие горные водоёмы ледниково-тектонического происхождения, в сборах из них представлены разные формы гольца, совместно обитающие в озёрах региона: из оз. Хантайское − длиннотычиночный и “тыптушка”, из оз. Аян – “гигант”, озёрно-речной и “путоранчик”, из оз. Собачье – голец Дрягина и молодь “боганидской палии”. Последняя отнесена к этой форме потому, что поймана на предустьевом участке притока озера р. Хоронен, в котором нерестится только она (у гольца Дрягина и третьей формы из оз. Собачье – пучеглазки − нерест озёрный; их заходы в реку не отмечены). К этой же форме принадлежат экземпляры из озёр Глубокое и Кета. Баяджяга (бассейн Хатанги) – небольшое озеро, населённое крупной формой гольца.
Гольцов отлавливали жаберными сетями с ячеёй 10, 20, 25, 30 и 40 мм. ДНК выделяли из фиксированных 96%-ным этанолом жировых плавников, кусочков мышечной ткани или семенников с помощью коммерческого набора Diatom DNA Prep 100 (OOO “Лаборатория Изоген”). Секвенировали фрагмент 534−543 пар оснований (п.о.), включающий весь левый участок контрольной области мтДНК гольцов, используя праймеры и условия, описанные ранее (Alekseyev et al., 2009). Редактирование хроматограмм и выравнивание последовательностей проводили в программе Geneious (Kearse et al., 2012). Последовательности, изученные в настоящей, а также в нашей предыдущей работе (Гордеева и др., 2018) перекрываются с последовательностями, изученными в работах: Brunner et al. (2001), Alekseyev et al. (2009), Moore et al. (2015), Osinov et al. (2017) на 499−507 п.о. Последовательности, впервые выявленные в настоящем исследовании, помещены в базу GenBank под номерами MT586474 (BER24), MT586475 (ATL32) и MT586476 (SIB35). Поскольку в литературе встречаются одинаковые (в пределах перекрывающегося участка последовательности) гаплотипы с разными названиями и разные с одинаковыми, мы используем названия, первыми размещённые в базе GenBank/NCBI, и в случаях дублирующихся названий указываем в скобках первую букву фамилии первого автора. Для проверки принадлежности ранее не изучавшихся гольцов из озёр Баяджяга и Булкурка-2 к S. alpinus у них оценили диагностически важные меристические признаки – число жаберных тычинок (sp.br.) и пилорических придатков (pc). Морфоэкологические данные по гольцам из остальных изученных озёр приведены в ряде работ (Борисов, 1932; Савваитова, Максимов, 1980; Романов, 1983, 1996, 2003; Савваитова, 1989; Павлов и др., 1994; Павлов, 1997; Пичугин, 2009).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Выявлено 16 изменчивых сайтов, включая одну делецию, и восемь гаплотипов − три новых (ATL32, SIB35 и BER24) и пять опубликованных ранее: ATL21(M) (Moore et al., 2015), SIB4 (Brunner at al., 2001), SIB10, SIB20 и BER10 (Alekseyev et al., 2009). ATL21(M) соответствует гаплотипу ATL20(O) (Osinov et al., 2017), SIB4 – гаплотипам SIB31 (Moore et al., 2015) и SIB34 (Алексеев и др., 2019).
В районе дельты Лены в области контакта сибирской подгруппы и берингийской группы в оз. Переходное найдены сибирские гаплотипы SIB10 и новый SIB35; в озёрах Дальнее и Булкурка-2 − берингийский гаплотип BER10; в оз. Ладаннах-Кюеле – новый берингийский гаплотип BER24. Следует подчеркнуть, что ранее в северных прибрежных районах Восточной Сибири отмечался только один сибирский (SIB25) и один берингийский (BER10) гаплотипы (Alekseyev et al., 2009; Гордеева и др., 2018). Оз. Переходное находится в близком соседстве с озёрами, в которых отмечен гаплотип BER10, – в частности, всего в 6 км от оз. Дальнее. Фенотипически гольцы из озёр Дальнее, Булкурка-2 и Ладаннах-Кюеле являются арктическими гольцами, а не северной мальмой, поскольку в отличие от неё имеют в среднем не менее 23.5 жаберных тычинок и 34 пилорических придатков и укладываются в пределы варьирования этих диагностических признаков у арктического гольца (рис. 2).
Рис. 2.
Соотношение среднего числа жаберных тычинок (sp.br.) и среднего числа пилорических придатков (рс) в разных популяциях и у разных форм арктического гольца Salvelinus alpinus Европы и Сибири ((− −) – атлантическая, (−⚪−) − сибирская подгруппа евроазиатской группы гаплотипов; (⚫)
– берингийская группа гаплотипов, гольцы из озёр (см. на рис. 1) 7, 9 и 10 в районе дельны Лены) и северной мальмы S. malma malma Чукотки и Камчатки ((- -▲- -) − берингийская группа гаплотипов), по собственным данным
и данным литературы: Борисов, 1932; Есипов, 1952; Колюшев, 1967; Nilsson, Filipsson,
1971; Balon, Penczak, 1980; Савваитова, Максимов, 1980; Черешнев, 1982; Klemetsen,
1984; Черешнев и др.,1989; Первозванский, Шустов, 1999; Есин, Маркевич, 2017.
−) – атлантическая, (−⚪−) − сибирская подгруппа евроазиатской группы гаплотипов; (⚫)
– берингийская группа гаплотипов, гольцы из озёр (см. на рис. 1) 7, 9 и 10 в районе дельны Лены) и северной мальмы S. malma malma Чукотки и Камчатки ((- -▲- -) − берингийская группа гаплотипов), по собственным данным
и данным литературы: Борисов, 1932; Есипов, 1952; Колюшев, 1967; Nilsson, Filipsson,
1971; Balon, Penczak, 1980; Савваитова, Максимов, 1980; Черешнев, 1982; Klemetsen,
1984; Черешнев и др.,1989; Первозванский, Шустов, 1999; Есин, Маркевич, 2017.
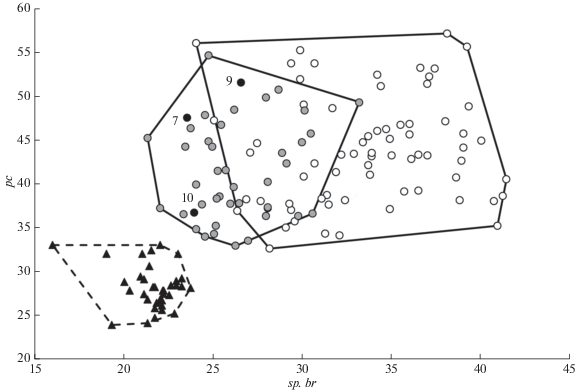
В области перекрывания ареалов атлантической и сибирской подгрупп в оз. Баяджяга (бассейн Хатанги) обнаружен атлантический гаплотип ATL32, в оз. Хантайское у двух форм гольца (длиннотычиночного и “тыптушки”) – гаплотип ATL21(M), отмеченный в оз. Лама (Moore et al., 2015; Osinov et al., 2017). Это соответственно самая восточная и самая южная точки распространения атлантической подгруппы в Восточной Сибири. Оз. Баяджяга находится вблизи границы ареала гольцов с гаплотипами берингийской группы, отмеченными в соседнем бассейне Анабара (Гордеева и др., 2018). Длиннотычиночные гольцы Хантайского озера имеют наибольшее число жаберных тычинок среди всех арктических гольцов атлантической подгруппы (sp.br. 30−38 (33.1): Романов, 2003) и сходны по этому признаку с гольцами из некоторых забайкальских популяций, принадлежащими к сибирской подгруппе, например, с гольцами мелкой формы из оз. Токко (sp.br. 31−37 (33.6): Алексеев и др., 2021). В свете полученных данных это сходство можно квалифицировать как случай параллелизма, хотя нельзя исключить возможность замещения мтДНК гольцов сибирской группы на мтДНК атлантической в результате интрогрессивной гибридизации.
В оз. Собачье у 1 экз. гольца Дрягина найден гаплотип ATL32, а у 2 экз. “боганидской палии” – сибирский SIB20, ранее отмечавшийся только в Забайкалье (Alekseyev et al., 2009; Гордеева и др., 2018). В оз. Глубокое у “боганидской палии” обнаружен гаплотип SIB10. Учитывая, что ранее (Гордеева и др., 2018) в этом озере отмечен атлантический гаплотип ATL22(M), список водоёмов с симпатрическими гаплотипами двух подгрупп пополняется ещё двумя озёрами. Важно отметить, что в этих трёх озёрах Норило-Пясинской системы (Лама, Собачье и Глубокое) и их притоках сибирские гаплотипы отмечены у “боганидской палии”, а атлантические (включая SIB9) – у других форм (голец Дрягина, пучеглазка, горный голец (Brunner et al., 2001; Osinov et al., 2017; Гордеева и др., 2018; настоящая работа)). Соответствие этих форм двум филогенетическим линиям, пока выявляемое у единичных особей, представляет большой интерес, но нуждается в проверке на более представительном материале. В оз. Кета такое соответствие не прослеживается, поскольку у изученного экземпляра “боганидской палии” обнаружен атлантический гаплотип ATL21(M). Впрочем, нельзя исключить ошибку в определении формы гольца, которую, к сожалению, невозможно проверить, поскольку экземпляр не сохранился.
В оз. Аян у всех форм нами отмечен сибирский гаплотип SIB4, ранее найденный у 2 экз. “путоранчика” из этого озера Бруннером с соавторами (Brunner et al., 2001) и Муром с соавторами (Moore et al., 2015). В последней работе он обозначен как SIB31, хотя полностью идентичен участку длиной 499 п.о. в последовательности, изученной Бруннером и его соавторами (550 п.о.). SIB4 соответствует гаплотипу SIB34 крупной формы гольца из оз. Малое Леприндо в Забайкалье (Алексеев и др., 2019). Таким образом, на сегодняшний день у гольцов Таймыра и Забайкалья известно три общих гаплотипа (SIB4, SIB10, SIB20).
Гаплотип SIB10 теперь обнаружен во всех основных районах распространения сибирской подгруппы: в Забайкалье, в бассейнах верхнего−среднего течения Яны и Индигирки, в дельте Лены и на Таймыре (Путоране). По-видимому, этот наиболее широко распространённый гаплотип сибирской подгруппы является для неё предковым (Гордеева и др., 2018). Полагаем, что сибирская подгруппа сформировалась в первой половине неоплейстоцена в гигантском Лено-Вилюйском палеоозере (оз. Лена, Якутское “море”), которое возникало на территории Восточной Сибири в периоды плейстоценовых оледенений в результате перекрывания северного стока Лены сползавшим с Верхоянского хребта ледником (Еникеев, 2009). Озеро имело сток в бассейн Енисея через Вилюй−Нижнюю Тунгуску и подходило близко к основным внутренним горным районам Восточной Сибири, в настоящее время населённым арктическим гольцом (Становое и Оймяконское нагорья, хребты Верхоянский, Сетте-Дабан, Сунтар-Хаята), а также к северным прибрежным районам. Связь его с Путораной осуществлялась через бассейн Нижней Тунгуски и низовья Енисея. Расселяясь по огромному озеру, гольцы имели возможность быстро и беспрепятственно достичь этих районов и обосноваться в них. Распространение сибирской подгруппы, очевидно, происходило практически исключительно за счёт пресноводных гольцов, и в этом состоит её отличие от атлантической, распространявшейся преимущественно за счёт проходных. Гаплотип SIB10, видимо, возник и распространился именно в Лено-Вилюйском озере, а в разных районах от него произошли уникальные для этих районов гаплотипы.
Таким образом, в северной части ареала арктического гольца на участке от Енисея до Лены выявлены новые митохондриальные гаплотипы трёх филогенетических линий – берингийской группы, атлантической и сибирской подгрупп евроазиатской группы, а также новые популяции гольцов с гаплотипами этих линий; продемонстрированы более широкие границы распространения атлантической подгруппы и зоны перекрывания её ареала с ареалом сибирской в бассейне Хатанги и в районе плато Путорана. Получены новые свидетельства того, что в Норило-Пясинских озёрах (возможно, за исключением оз. Кета) сибирская подгруппа представлена “боганидской палией”, а атлантическая – другими отмеченными в них формами. Обнаружение на севере трёх южных гаплотипов сибирской группы подтверждает ранее сделанный вывод (Alekseyev et al., 2009; Гордеева и др., 2018) об общем происхождении и близких филогенетических связях арктических гольцов сибирской подгруппы из разных областей Восточной Сибири.
Список литературы
Алексеев С.С., Пичугин М.Ю., Гордеева Н.В. и др. 2019. Репродуктивные стратегии и происхождение парапатрических и симпатрических форм арктического гольца Salvelinus alpinus (Salmonidae) в системе озёр Большое и Малое Леприндо (Северное Забайкалье) // Вопр. ихтиологии. Т. 59. № 4. С.430−447. https://doi.org/10.1134/S0042875219040015
Алексеев С.С., Самусенок В.П., Юрьев А.Л. и др. 2021. Морфологическая и экологическая дифференциация симпатрических форм арктического гольца Salvelinus alpinus (Salmonidae) в озере Токко (северное Забайкалье) // Там же. Т. 61. № 1. С. 65−87. https://doi.org/10.31857/S0042875221010021
Борисов П.Г. 1932. Новый вид гольца − Salvelinus jacuticus n. sp. // Материалы к ихтиофауне бассейна р. Лены. М.: Изд-во АН СССР. С. 1−42.
Гордеева Н.В., Алексеев С.С., Кириллов А.Ф. и др. 2018. Распространение, состав и родственные отношения филогенетических групп арктического гольца Salvelinus alpinus (L.) (Salmonidae) в европейской части России и Сибири по данным анализа нуклеотидных последовательностей митохондриальной ДНК // Вопр. ихтиологии. Т. 58. № 6. С. 659−669. https://doi.org/10.1134/S0042875218050107
Еникеев Ф.И. 2009. Плейстоценовые оледенения восточного Забайкалья и юго-востока Средней Сибири // Геоморфология. Т. 2. С. 33−49. https://doi.org/10.15356/0435-4281-2009-2-33-49
Есин Е.В., Маркевич Г.Н. 2017. Гольцы рода Salvelinus азиатской части северной Пацифики. Петропавловск-Камчатский: Камчатпресс, 188 с.
Есипов В.К. 1952. Рыбы Карского моря. Л.: Изд-во АН СССР, 147 с.
Колюшев А.И. 1967. К морфологии и систематическому положению палии озер Кольского п-ова // Тр. Карел. отд. ГОСНИОРХ. Т. 5. Вып. 1. С. 269−273.
Олейник А.Г., Скурихина Л.А., Кухлевский А.Д. 2017. Вторичный контакт между двумя дивергентными линиями гольцов рода Salvelinus в Северо-Западной Пацифике // Генетика. Т. 53. № 11. С. 1297−1310. https://doi.org/10.7868/S001667581711008X
Осинов А.Г., Павлов Д.А., Волков А.А. 2018. К вопросу о происхождении озёрных гольцов Salvelinus alpinus complex из бассейнов Колымы и Охотского моря // Вопр. ихтиологии. Т. 58. № 3. С. 313−330. https://doi.org/10.7868/S0042875218030086
Павлов С.Д. 1997. Симпатрические формы гольцов (род Salvelinus) из озера Аян (Таймырский полуостров) // Вопр. ихтиологии. Т. 37. № 4. С.465-474.
Павлов С.Д., Савваитова К.А., Максимов В.А. 1994. О взаимоотношениях симпатрических группировок арктических гольцов в озере Собачье (Норило-Пясинская водная система) // Систематика, биология и биотехника разведения лососевых рыб. СПб.: Изд-во ГосНИОРХ. С. 148−151.
Первозванский В.Я., Шустов Ю.А. 1999. Карликовая форма гольца Salvelinus alpinus (Salmonidae) в озере Верхний Нерис (Паанаярвский национальный парк, Карелия) // Вопр. ихтиологии. Т. 39. № 1. С. 131−132.
Пичугин М.Ю. 2009. Развитие искусственного гибрида и выявление элементов репродуктивной изоляции между симпатрическими формами гольца Дрягина и пучеглазки Salvelinus alpinus complex (Salmonidae) из горного озера Собачье (Таймыр) // Там же. Т. 49. № 2. С. 240−253.
Радченко О.А. 2003. Изменчивость митохондриальной ДНК в популяциях озерных гольцов рода Salvelinus Дальнего Востока и Сибири // Там же. Т. 43. № 4. С. 553−561.
Радченко О.А. 2004. Изменчивость нуклеотидных последовательностей гена цитохрома b митохондриальной ДНК гольцов рода Salvelinus // Генетика. Т. 40. № 3. С. 322−333.
Романов В.И. 1983. Экологическая структура гольцов (р. Salvelinus) Хантайского озера // Вопр. географии Сибири. Вып. 14. С. 73−88.
Романов В.И. 1996. К вопросу о популяционной структуре гольцов (р. Salvelinus) озера Кета // Матер. конф. “Задачи и проблемы развития рыбного хозяйства на внутренних водоемах Сибири”. Томск. С. 97−98.
Романов В.И. 2003. Морфологические особенности массовых форм гольцов (род Salvelinus) озера Хантайского // Вестн. ТГПУ. Сер. естеств. и точные науки. Вып. 4 (36). С. 66−72.
Савваитова К.А. 1989. Арктические гольцы (структура популяционных систем, перспективы хозяйственного использования). М.: Агропромиздат, 223 с.
Савваитова К.А., Максимов В.А. 1980. Формообразование у гольцов рода Salvelinus (Salmoniformes, Salmonidae) из озёр дельты Лены // Зоол. журн. Т. 59. № 12. С. 1820−1830.
Черешнев И.А. 1982. К вопросу о таксономическом статусе симпатрических проходных гольцов рода Salvelinus (Salmonidae) восточной Чукотки // Вопр. ихтиологии. Т. 22. Вып. 6. С. 922−936.
Черешнев И.А., Гудков П.К., Нейман М.Ю. 1989. Первые данные по биологии проходной мальмы бассейна р. Чегитунь (арктическое побережье восточной Чукотки) // Там же. Т. 29. Вып. 1. С. 68−83 .
Alekseyev S.S., Bajno R., Gordeeva N.V. et al. 2009. Phylogeography and sympatric differentiation of the Arctic charr Salvelinus alpinus (L.) complex in Siberia as revealed by mtDNA sequence analysis // J. Fish Biol. V. 75. № 2. P. 368–392. https://doi.org/10.1111/j.1095-8649.2009.02331.x
Balon E.K., Penczak T. 1980. The dwarfed charr of Dösener See, an alpine lake in Austria // Charrs: salmonid fishes of the genus Salvelinus. Perspectives in vertebrate science 1 / Ed. Balon E.K. The Hague: Dr. W. Junk Publ. P. 773−794.
Brunner P.C., Douglas M.R., Osinov A. et al. 2001. Holarctic phylogeography of Arctic charr (Salvelinus alpinus L.) inferred from mitochondrial DNA sequences // Evolution. V. 55. № 3. P. 573−586. https://doi.org/10.1554/0014-3820(2001)055[0573:hpoacs] 2.0.co;2
Esin E.V., Bocharova E., Mugue N.S., Markevich G.N. 2017. Occurrence of sympatric charr groups, Salvelinus, Salmonidae, in the lakes of Kamchatka: a legacy of the last glaciations // J. Fish Biol. V. 91. № 2. P. 628−644. https://doi.org/10.1111/jfb.13378
Kearse M., Moir R., Wilson A. et al. 2012. Geneious basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data // Bioinformatics. V. 28. № 12. P. 1647−1649. https://doi.org/10.1093/bioinformatics/bts199
Klemetsen A. 1984. The Arctic charr speciation problem as seen from northern Norway // Proc. Int. Symp. “Biology of the Arctic charr”. Winnipeg: Univ. Manitoba Press. P. 65−77.
Moore J.-S., Bajno R., Reist J.D., Taylor E.B. 2015. Post-glacial recolonization of the North American Arctic by Arctic char (Salvelinus alpinus): genetic evidence of multiple northern refugia and hybridization between glacial lineages // J. Biogeogr. V. 42. № 11. P. 2089−2100. https://doi.org/10.1111/jbi.12600
Nilsson N.-A., Filipsson O. 1971. Characteristics of two discrete populations of Arctic charr (Salvelinus alpinus L.) in a north Swedish lake // Rept. Inst. Freshwat. Res. Drott. V. 51. P. 90−108.
Osinov A.G., Senchukova A.L., Mugue N.S. et al. 2015. Speciation and genetic divergence of three species of charr from ancient Lake El’gygytgyn (Chukotka) and their phylogenetic relationships with other representatives of the genus Salvelinus // Biol. J. Linnean Soc. V. 116. P. 63−85. https://doi.org/10.1111/bij.12559
Osinov A.G., Volkov A.A., Alekseyev S.S. et al. 2017. On the origin and phylogenetic position of Arctic charr (Salvelinus alpinus complex, Salmonidae) from Lake Cherechen’ (middle Kolyma River basin): controversial genetic data // Polar Biol. V. 40. P. 777−786. https://doi.org/10.1007/s00300-016-2000-4
Salisbury S.J., McCracken G.R., Keefe D. et al. 2019. Extensive secondary contact among three glacial lineages of Arctic char (Salvelinus alpinus) in Labrador and Newfoundland // Ecol. Evol. V. 9. № 4. P. 2031−2045. https://doi.org/10.1002/ece3.4893
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии