

Вопросы ихтиологии, 2022, T. 62, № 2, стр. 148-159
О двух малоизученных видах угревидных ликодов (Lycenchelys, Zoarcidae)
1 Институт проблем экологии и эволюции РАН – ИПЭЭ РАН
Москва, Россия
2 Институт океанологии РАН – ИО РАН
Москва, Россия
* E-mail: prokartster@gmail.com
Поступила в редакцию 12.02.2021
После доработки 09.03.2021
Принята к публикации 10.03.2021
- EDN: XJZBIM
- DOI: 10.31857/S0042875222020199
Аннотация
Спустя 70 лет после поимки голотипа сообщается о нахождении второго экземпляра абиссального вида Lycenchelys uschakovi, по ряду признаков (число посторбитальных пор, степень развития чешуйного покрова) заметно отличающегося от голотипа. Это самая глубоководная находка лиценхелов в Северной Пацифике (4890–4984 м). С батиальных глубин (~800 м) у о-ва Парамушир описываются два экземпляра, соответствующие признакам номинального вида L. parini, также ранее известного только по голотипу. На основании анализа литературных данных по лиценхелам Северной Пацифики делается вывод о конспецифичности L. parini и L. melanostomias, последнее название имеет приоритет. Новая находка существенно расширяет известный ареал L. melanostomias к северу.
Угревидные ликоды рода Lycenchelys Gill, 1884 представлены более чем 60 видами, населяющими преимущественно батиальные и абиссальные глубины высоких широт всех океанов (Fricke et al., 2021). Виды данного рода характеризуются отсутствием подбородочных гребней, увеличенными порами сейсмосенсорной системы головы, 6–10 инфраорбитальными костями и таким же числом пор в подглазничном сенсорном канале; слабым развитием дыхательной перепонки и костей палато-птеригоидной серии; удлинённым, покрытым чешуёй телом; положением первого дорсального птеригиофора, несущего луч, над 2–21 позвонком и 0–16 свободными supraneuralia перед ним, отсутствием костных пластинок в основаниях дорсальных птеригиофоров; как правило, наличием брюшных плавников, ложножабры, зубов на сошнике и нёбных костях (но у отдельных видов отсутствуют) (Андрияшев, 1955; Anderson, 1994). В Беринговом море и водах северо-западной части Тихого океана отмечены 24 вида Lycenchelys, пять из них (L. albeola Andriashev, 1958, L. micropora Andriashev, 1955, L. plicifera Andriashev, 1955, L. volki Andriashev, 1955 и L. uschakovi Andriashev, 1958) известны с глубин, превышающих 3000 м (Андрияшев, 1955, 1958; Toyoshima, 1985; Anderson, 1995; Thiel et al., 2018; Kawarada et al., 2020). Все пять видов были описаны по сборам научно-исследовательского судна (НИС) “Витязь” в абиссали Берингова моря и Курило-Камчатской впадины в 1950-х гг., три из них (кроме L. micropora и L. plicifera) до сих пор известны только по голотипам (второй экземпляр L. plicifera первоначально был описан как голотип особого вида L. birsteini Andriashev, 1958).
На батиальных и верхнеабиссальных (до 2500 м) глубинах Курило-Камчатской впадины и охотоморского побережья Камчатки и Северных Курил могут быть встречены 11 видов (Андрияшев, 1955, 1958; Федоров, Андрияшев, 1993; Федоров, 1995; Anderson, 1995; Шейко, Федоров, 2000; Thiel et al., 2018; Kawarada et al., 2020): L. camchatica (Gilbert et Burke, 1912), L. fedorovi Anderson et Balanov, 2000, L. hippopotamus Schmidt, 1935, L. lenzeni Thiel, Knebelsberger et Eidus, 2018, L. makushok Fedorov et Andriashev, 1993, L. micropora, L. parini Fedorov, 1995, L. rassi Andriashev, 1955, L. ratmanovi Andriashev, 1955 (синоним – L. longirostris Toyoshima, 1985), L. squamosa Toyoshima, 1983 и L. vitiazi Andriashev, 1955. Большинство перечисленных видов в настоящее время известны по достаточному числу экземпляров, но L. parini и L. vitiazi до сих пор представлены в коллекциях лишь голотипами, и сведения об этих видах очень скудны.
Разбирая сборы рейсов 11А и 22 НИС “Академик М. Келдыш”, я обнаружил в них три ранее не исследованных экземпляра Lycenchelys, пойманных у восточного берега Камчатки и охотоморского побережья Северных Курил. Изучение этих рыб вскрыло явную неполноту сведений по таксономии и внутривидовой изменчивости лиценхелов данной акватории. Два молодых экземпляра с батиальных глубин (~800 м) к западу от о-ва Парамушир, пойманные в рейсе 11А, были довольно уверенно сопоставлены с L. parini, описанным Федоровым (1995) из того же района, но со стороны Курило-Камчатской впадины. Однако сравнение их с другими северотихоокеанскими видами позволило усомниться в валидности данного вида, оказавшегося неотличимым от японского L. melanostomias Toyoshima, 1983. Третий экземпляр, пойманный в 22-м рейсе в абиссали Курило-Камчатской впадины (4890–4984 м), является самой глубоководной находкой представителя рода в Северной Пацифике. Его определение вызвало существенные затруднения и первоначально предполагалось его описание в качестве нового для науки вида, однако анализ изменчивости другого абиссального вида – L. antarctica Regan, 1913 – в итоге позволил отнести рассматриваемый экземпляр к виду L. uschakovi. Такое решение предполагает значительную изменчивость этого вида по степени развития чешуйного покрова, превышающую ранее известную для других Lycodinae. Результаты изучения упомянутых экземпляров изложены в настоящей работе.
МАТЕРИАЛ И МЕТОДИКА
Описанные в работе рыбы хранились в растворе формалина до их обнаружения в 2020 г., когда они были переведены в 70%-ный этанол. Сохранность материала полная, за исключением вероятного частичного обесцвечивания вследствие длительного хранения. Изученный материал хранится в коллекции Института океанологии РАН, Москва (ИО РАН); для сравнения использованы материалы Зоологического института РАН, Санкт-Петербург (ЗИН РАН). Методика изучения и терминология соответствуют общепринятым (Андрияшев, 1955; Peden, Anderson, 1978; Anderson, 1982b, 1995; Toyoshima, 1985) со следующими дополнениями: вертикальный диаметр глаза измеряли через центр зрачка, высоту основания грудного плавника – от верхнего края основания самого верхнего до нижнего края основания самого нижнего луча этого плавника. В соответствии с рекомендацией Андерсона (Anderson, 1995) пропорции частей тела рассчитаны от стандартной длины. Общее число позвонков и число хвостовых позвонков включает уростиль. В работе использованы следующие сокращения: SL, TL – стандартная и абсолютная длина тела; lc, wc, hc – соответственно длина и максимальная ширина головы и её высота на уровне середины глаза; H, НА – максимальная высота тела и его высота на уровне начала анального плавника; lP, lV, lC – длина соответственно грудного, брюшного и хвостового плавников; hP – высота основания грудного плавника; aD, aA, aV – соответственно предорсальное, преанальное и превентральное расстояния; ao – длина рыла; oo, ho – горизонтальный и вертикальный диаметр глаза; io – минимальная ширина костного межглазничного промежутка; lmx – длина верхней челюсти; lbr, wbr – длина и ширина основания наибольшей жаберной тычинки; ibr – промежуток между основаниями четвёртой и пятой жаберных тычинок; lba – длина жаберной щели; D, A, P, V, С – соответственно спинной, анальный, грудные, брюшные и хвостовой плавники; r. br, sp. br и pbr – соответственно число жаберных лучей, тычинок наружного ряда первой жаберной дуги и элементов ложножабры; vert., vert. abd. – общее число позвонков (туловищных + хвостовых) и число туловищных позвонков; ст. – океанографическая станция. Показатели, различающиеся на разных сторонах тела одной и той же рыбы, разделены знаком “/”. В синонимии указаны только те работы, в которых приведены морфологические данные по обсуждаемым видам.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Lycenchelys uschakovi Andriashev, 1958
(рис. 1–3)

Рис. 2.
Lycenchelys uschakovi TL 188 мм, детали строения головы (а–г – вид соответственно сверху, сбоку, вентролатерально, снизу) и расположение пор сейсмосенсорной системы (д, е). N – трубочка ноздри, squ – участок чешуи на жаберной крышке; поры: io – инфраорбитальные, na – носовые, pm – преоперкуло-мандибулярные, po – заглазничная. Масштаб: а – 4.5, б–е – 8 мм (линейка общая).
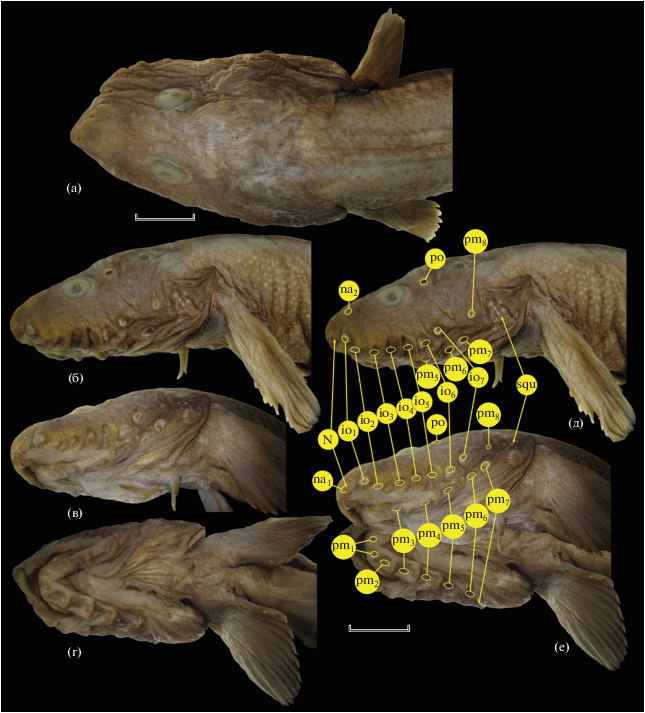
Рис. 3.
Lycenchelys uschakovi, чешуйный покров брюха: а – экз. TL 188 мм, б – голотип TL 259 мм. Масштаб: а – 10, б – 15 мм.

Lycenchelys uschakovi Андрияшев, 1958. С. 176. Рис. 3 (первоописание). Toyoshima, 1985. P. 155. Anderson, 1995. P. 58 (в ключе).
Материал. 1 экз. TL 188 мм (SL 184 мм), 53°05′40′′–53°07′00′′ с.ш., 161°55′20′′–161°56′12′′ в.д., НИС “Академик М. Келдыш”, рейс 22, ст. 2323, трал Сигсби, 4890–4984 м, 16:31–19:40, 10.08.1990 г. Экземпляр в полной сохранности, но рыло примято с правой стороны.
Сравнительный материал. Голотип L. uschakovi, ЗИН № 34670, TL 259 мм, восточнее о‑ва Парамушир, 3960–4070 м, НИС “Витязь”, ст. 2209, 24.06.1953 г. Lycenchelys antarctica: ЗИН № 44940 (голотип L. atacamensis Andriashev, 1980), TL 184 мм, Перуанско-Чилийский жёлоб, 05°52ʹ ю.ш., 81°48′ з.д., 5300–5320 м, НИС “Академик Курчатов”, ст. 301, 04.11.1968 г.; ЗИН № 48393, 2 экз. TL 141 и 242 мм, 14°45ʹ ю.ш., 76°49ʹ з.д., 4835–5000 м, НИС “Дмитрий Менделеев”, ст. 1656, 20.03.1978 г.
Диагноз. Вид рода Lycenchelys с 10–15 свободными supraneuralia, 28 туловищными позвонками, одной–двумя посторбитальными порами, без корональной и затылочных пор, с вентральной боковой линией и пигментированным телом. Сплошной чешуйный покров головы отсутствует, но отдельные участки с чешуёй появляются по достижении SL по меньшей мере 184 мм. D 101, A 94–96, V 2, vert. 119–122.
Описание. D 101, А 94, Р 17, V 2, C 11 (6 + 5); r. br 6, sp. br 14, pbr 1; vert. 28 + 91 = 119.
Тело сильно удлинённое, наиболее высокое у затылка (11.9 раза в SL), спереди почти круглое в сечении, к хвостовому концу сжатое с боков (рис. 1). Голова умеренной величины, 6.8 раза в SL, уплощённая, её дорсальный профиль плавно понижается от межглазничной области к круто обрывающейся вершине рыла. Глаз слабо эллиптический (его вертикальный диаметр в 1.14 раза меньше горизонтального), в профиль не достигает дорсального контура головы, его горизонтальный диаметр 1.2 раза укладывается в ширине костного межглазничного промежутка (рис. 2а, 2б). Трубочка ноздри, прижатая вперёд, не достигает края верхней губы. Рот нижний, верхняя челюсть нависает перед нижней, назад достигает вертикали переднего края глаза. Верхняя губа мясистая; нижнегубные лопасти крупные, с широко закруглённым свободным краем, начинаются от линии, проходящей через середину первой пары мандибулярных (подбородочных) пор, доходят назад до конца ротовой щели (рис. 2в, 2г). Зубы в челюстях тонкие, заострённые, расставленные, скрыты в массе соединительной ткани, но у симфизов выступающие наружу; озубление praemaxillaria однорядное; на нижней челюсти зубы в два ряда у симфиза, далее назад – однорядные. Зубы на vomer и palatina мелкие, конические, широко расставленные, сокрытые в толстой ткани мягкого нёба. Сошник с двумя зубами на переднем крае головки кости (один зуб в центре и один – с правой стороны, с левой стороны зубы отсутствуют); на palatina с обеих сторон по три зуба в ряд. Дыхательная перепонка слабо развита. Мягкие ткани крыши ротовой полости складчатые, складки образованы крупными папиллами. Жаберная щель доходит вниз до уровня нижнего края основания Р и вперёд – до уровня прикрепления V. С внутренней стороны жаберного отверстия у нижнего конца жаберной щели имеется маленький мясистый выступ. Жаберных тычинок на первой дуге 3 + 11, из них три самые нижние – в виде маленьких бугорков. Наиболее развитые тычинки в длину в 1.5 раза превышают свою ширину в основании, соизмеримы с длиной промежутка между четвёртой и пятой тычинками. Ложножабра представлена единственным коротким лепестком (изучена только с левой стороны). Оперкулярная лопасть хорошо выражена, сужающаяся к закруглённой вершине (рис. 2б).
Поры сейсмосенсорных каналов головы (рис. 2д, 2е) открываются крупными отверстиями, в особенности инфраорбитальные и преоперкуло-мандибулярные (“ноздревидные”). Две носовые (передние супраорбитальные) поры: передняя из них расположена кпереди от трубочки ноздри, а задняя – посередине расстояния между вершиной рыла и передним краем глаза. Два ряда свободных невромастов (2 + 3) на вершине рыла между передними носовыми порами соседних сторон. Инфраорбитальных пор семь, все расположены на горизонтальном отрезке костей окологлазничной серии; пятая пора располагается на вертикали заднего края глаза; последняя инфраорбитальная пора расположена на одном уровне с шестой преоперкуло-мандибулярной порой. Преоперкуло-мандибулярных пор восемь, из них четыре на dentale, одна на anguloarticulare и три на praeoperculum; восьмая пора заметно мельче предшествующих, открывается на середине промежутка между задним краем глаза и верхним краем основания Р на линии, проходящей выше седьмой инфраорбитальной поры на расстоянии половины диаметра последней. С каждой стороны головы присутствует единственная посторбитальная пора, расположенная на вертикали шестой инфраорбитальной поры. Корональная и затылочные поры отсутствуют. Туловищная боковая линия, представленная трудноразличимыми свободно сидящими невромастами, начинается от вершины оперкулярной лопасти, полого спускается вниз к вентральному краю тела (до уровня границы средней и нижней трети высоты основания Р) и пропадает на уровне начала А.
Начало D расположено позади конца прижатого к телу Р, примерно посередине между ним и вертикалью начала А; первый дорсальный птеригиофор, несущий луч, расположен между 16-м и 17-м невральными отростками; первый луч D очень мал, шесть передних лучей D скрыты в толще эпаксиальных мышц и соединительной ткани; имеется 10 свободных supraneuralia, из которых первое – вильчатое (вероятно, сформировано слиянием двух элементов), расположено между седьмым и восьмым невральными отростками; крошечный костный элемент (рудимент луча?) различим между 9-м и 10-м свободными supraneuralia; передние два анальных птеригиофора расположены впереди вершины первого гемального отростка. Концы лучей Р, прижатого вперёд, достигают заднего края глаза; дистальный край этого плавника закруглённый, его срединные лучи наибольшие. Длина V в 1.25 раза превышает горизонтальный диаметр глаза, эти плавники прикрепляются на вертикали середины промежутка между шестой инфраорбитальной порой и основанием Р. Анус расположен у начала А. Пилорические придатки не развиты.
Чешуя на голове отсутствует, за исключением 7/6 изолированных чешуй на жаберной крышке; совершенно нет чешуи на затылке и вдоль срединно-спинной линии в предорсальной области, а также на горле, груди и основании Р. Чешуя полностью отсутствует на брюхе между горизонталями, проходящими через нижние концы оснований Р правой и левой сторон назад вплоть до начала А (рис. 3а). Чешуя не заходит на вертикальные плавники, за исключением их самых задних отделов, где редкие, очень мелкие чешуйки присутствуют по ходу лучей D и А. Кожа головы подвижная и отчасти складчатая, на теле плотно прикреплённая. Вентральная складка отсутствует.
Окраска фиксированного экземпляра после 30 лет хранения в формалине очень светлая, бежевая, но заметно более тёмная, коричневатая на плавниках, в абдоминальной области и в особенности на боковой и вентральной поверхности головы (рис. 1). Ротовая полость светлая, жаберная – коричневатая; жаберные дуги светлые. Брюшина одноцветно-коричневая. Желудок коричневатый; кишка беловатая, с множественными меланофорами в виде чёрточек в заднем отделе.
Измерения. В % SL: lc 14.7, wc 7.6, hc 5.7, H 8.4, HA 7.6, lР 9.2, lV 2.7, hР 4.6, lС 2.5, aD 31.5, aA 35.3, aV 12.5, ao 4.4, oo 2.2, ho 1.9, io 2.6, lmx 4.4, lbr 0.4, wbr 0.3, ibr 0.4, lba 5.4. В % lc: wc 51.9, hc 38.9, H 57.4, HA 51.9, lР 63.0, lV 18.5, hР 31.5, lС 16.7, aD 214.8, aA 240.7, aV 85.2, ao 29.6, oo 14.8, ho 13.0, io 17.6, lmx 29.6, lbr 2.8, wbr 1.9, ibr 2.8, lba 37.0.
Сравнительные замечания. Только восемь видов лиценхелов описаны с глубин, превышающих 3000 м (пять из них отмечены в водах Берингова моря и Курило-Камчатской впадины). Все эти виды характеризуются редукцией передней части D, так что перед первым несущим луч птеригиофором всегда имеются свободные supraneuralia, а наиболее передние лучи D скрыты в мягких тканях и не видны снаружи. Различия между этими видами суммированы в таблице. У всех остальных видов рода редукция передней части D не отмечена, за исключением L. hureaui (Andriashev, 1979) из батиали Кергеленского района Южного океана, имеющего 0–1 свободных supraneuralia, однако этот вид может быть легко отличён по малому числу туловищных позвонков (22), наличию трёх посторбитальных пор и отсутствию V (Андрияшев, 1979).
Диагностические признаки абиссальных видов Lycenchelys, известных с глубин > 3000 м
| Признак | Вид, источник информации | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| L. alba | L. albeola | L. antarctica | L. monstrosa | L. pearcyi | L. plicifera | L. uschakovi | L. volki | ||
| Голотип | экз. TL 188 мм | ||||||||
| Geistdoerfer et al., 1970; Møller, 1999 | Андрияшев, 1958 | Anderson, 1988, 1995; наши данные | Anderson, 1982a, 1995 | Anderson, 1995 | Андрияшев, 1955; Anderson, 1995 | Андрияшев, 1958; наши данные | Наши данные | Андрияшев, 1955; наши данные | |
| D | 115–120 | 127 | 102–116 | 111–118 | 118–124 | 110–115 | 101 | 101 | 113 |
| A | 110–116 | 109 | 98–112 | 98–108 | 104–109 | 99–102 | 96 | 94 | 101 |
| vert. | 133–141 | 133 | 120–138 | 124–132 | 128–134 | 122–126 | 122 | 119 | 126 |
| vert. abd. | 25–29 | 27 | 26–29 | 26–30 | 27–30 | 28–30 | 28 | 28 | 30 |
| Число свободных supraneuralia | 10–12 | 2* | 6–16 | 1–7 | 1–3 | 5–6 | 15 | 10 | 7 |
| Корональная пора | – | – | – | – | + или – | – | – | – | +** |
| Число посторбитальных пор | 2 | 2 | 1–2 | 1*** | 1–2 | 1–2 | 2 | 1 | 2 |
| Чешуя на голове | – | – | – | – | – | – | Единичная на затылке, щеке и жаберной крышке | Единичная на жаберной крышке | – |
| Чешуя на брюхе | + | + | + | + | + | + | + | – | + |
| Складка кожи по средней линии брюха | – | – | – | – | + | + | – | – | – |
| Туловищная боковая линия | Двойная | Дорсальная | Вентральная | Двойная | Вентральная | Двойная | Вентральная | Вентральная | Вентральная |
| Пигментация тела | – | – | + | + | + | + | + | + | + |
| Географическое распространение | Северная Атлантика |
Курило-Камчатский жёлоб |
Антарктика и Перуанско-Чилийский жё-лоб | От Калифорнии до Панамы | От Орегона до Мексики | Алеутский и Курило-Камчатский жёлоб | Курило-Камчатский жёлоб | Берингово море | |
Примечание. “+” – есть, “–” – нет; * последующие пять птеригиофоров несут короткие слабые лучи, скрытые в тканях и не определимые снаружи; ** в первоописании эта пора указана в тексте, но не изображена на рисунке (Андрияшев, 1955. Рис. 14 ); *** у одного из 13 исследованных экземпляров с одной стороны посторбитальная пора двойная (Anderson, 1995).
Описанный экземпляр проявляет очевидное сходство с L. uschakovi, который до сих пор был известен только по голотипу, пойманному в Курило-Камчатской впадине на широте о-ва Парамушир при тралении на глубине 3960–4070 м. Однако некоторые признаки нового экземпляра не согласуются с таковыми у голотипа, в связи с чем его идентификация с этим видом нуждается в доказательствах. Новый экземпляр отличается от голотипа единственной (против двух) посторбитальной порой с каждой стороны головы, полным отсутствием чешуи на брюхе в промежутке между концами оснований Р и назад до начала А (тогда как у голотипа чешуя сплошь покрывает брюхо от линии, соединяющей нижние концы оснований Р, до начала А) (рис. 3), а также отсутствием чешуи в предорсальной области, её слабым развитием на вертикальных плавниках и меньшим числом свободных supraneuralia (10 против 15). Число посторбитальных пор является важным признаком в систематике рода: Федоров и Андрияшев (1993) даже предпринимали попытку разделить северотихоокеанские виды на группы по данному признаку. Однако внутривидовая изменчивость в числе этих пор не так уж и редка. Так, у абиссального L. antarctica отмечена как одна, так и две пары посторбитальных пор, что соответствует различиям, наблюдаемым между голотипом L. uschakovi и описываемым здесь экземпляром. Разница в одну–две поры в посторбитальной серии отмечена также для L. pearcyi Anderson, 1995, L. plicifera и для ряда батиальных видов (Anderson, 1995; Thiel et al., 2018; Kawarada et al., 2020). Кроме того, у голотипа L. uschakovi посторбитальные поры не одинаково развиты на разных сторонах головы – справа поры сближены гораздо сильнее, чем слева (Андрияшев, 1958. Рис. 3). Это позволяет предполагать, что у данного вида число и положение посторбитальных пор не строго детерминировано. По другим меристическим признакам различия между голотипом L. uschakovi и описываемым здесь экземпляром полностью укладываются в известные пределы изменчивости у L. antarctica. Так, число свободных supraneuralia у L. antarctica варьирует от 6 до 16 (Anderson, 1988, 1995), что значительно снижает диагностическую ценность данного признака (а также различий в положении начала видимой снаружи части D).
Однако различия в степени развития чешуйного покрова между голотипом L. uschakovi и новым экземпляром существенно превышают известные пределы возрастной и индивидуальной изменчивости по данному признаку среди Lycodinae. У нового экземпляра чешуя совершенно отсутствует на брюхе и по средней линии спины в предорсальной области; редкие, очень мелкие чешуйки присутствуют по ходу лишь самых задних лучей D и А, тогда как у голотипа брюхо и спина от уровня оснований Р покрыты чешуёй, на вертикальных плавниках чешуя имеется и в начальной их части (близ основания), а в задней сплошь покрывает их почти до края. Кроме того, у нового экземпляра абсолютные размеры отдельных чешуек меньше, а сам чешуйный покров гораздо более разрежен (промежутки между чешуйками больше их диаметра, тогда как у голотипа чешуйки почти соприкасаются). Поскольку новый экземпляр мельче голотипа (TL 188 против 259 мм), различия в степени развития чешуйного покрова можно было бы расценивать как возрастные, однако они слишком велики для столь консервативного в пределах группы признака. У абиссального вида L. antarctica, наиболее близкого к L. uschakovi по совокупности морфологических признаков (таблица) и столь же глубоководного, при сходных размерах различия в степени развития чешуи выявлены лишь в передней части брюха. У экземпляров из Перуанско-Чилийского жёлоба, имеющих TL 141 и 184 мм, брюхо лишено чешуи до уровня середины длины прижатых к телу Р, тогда как у более крупной рыбы TL 242 мм чешуя доходит вперёд до линии, соединяющей нижние концы оснований Р. На спине чешуйный покров правой и левой сторон соединяется перед началом D у всех рыб, на плавниках чешуя отсутствует, что, по-видимому, характерно для вида в целом. В то же время разреженность чешуи на теле у L. antarctica с ростом явственно уменьшается. Анализ литературных данных и экземпляров L. melanostomias и L. sarsii (Collett, 1871), имеющихся в коллекции ИО РАН, позволяет заключить, что характер изменчивости в развитии чешуи на теле, описанный выше для L. antarctica, характерен для рода в целом, а ситуация, наблюдаемая у L. uschakovi, если относить к этому виду новый экземпляр, оказывается уникальной. Насколько стабильна подобная задержка в степени очешуения предорсальной области и брюха в онтогенезе L. uschakovi, можно будет судить только после поимки новых экземпляров этого вида.
Поскольку за исключением степени развития чешуйного покрова не удалось найти других достоверных различий между голотипом L. uschakovi и описанным здесь экземпляром, я считаю последнего конспецифичным данному виду. Новая находка является одной из самых глубоководных для рода – с бóльших глубин известен лишь L. antarctica в Перуанско-Чилийском жёлобе (до 5320 м) (Anderson, 1995).
Lycenchelys melanostomias Toyoshima, 1983

Рис. 5.
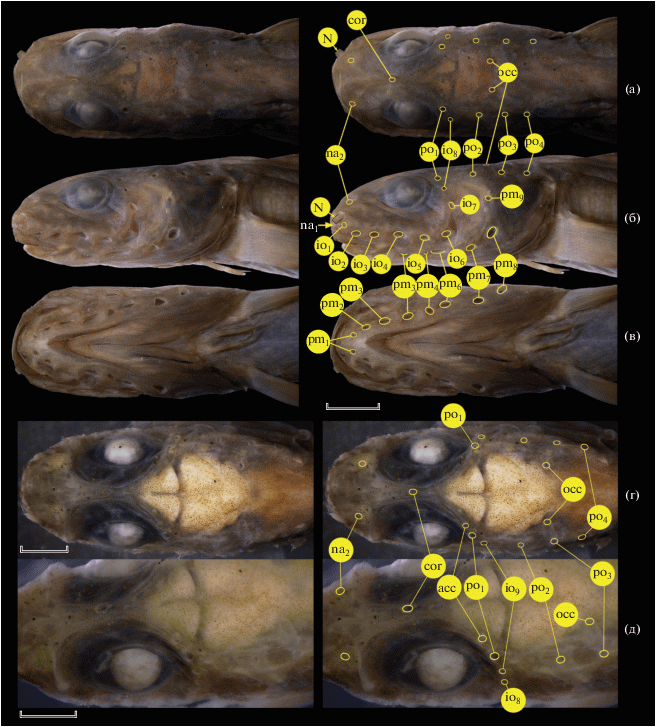
Lycenchelys melanostomias TL 103 (а–в) и 56 (г, д) мм; детали строения головы и расположение пор сейсмосенсорной системы, вид: а, г –сверху; б, в, д – соответственно сбоку, снизу, дорсолатерально. acc, cor, occ – соответственно дополнительная (задняя надглазничная), корональная и затылочные поры; ост. обозначения см. на рис. 2. Масштаб: а–в – 3 (линейка общая); г, д – 0.75 мм.
Lycenchelys melanostomias Toyoshima, 1983. P. 271. Pl. 157. Figs. 25–27 (первоописание). Toyoshima, 1985. P. 170. Figs. 26, 27, 31I . Hatooka, 2002. P. 1032 (в ключе). Imamura et al., 2004. P. 84. Figs. 1–3. Tabl. 1. Kawarada et al., 2020. P. 27. Figs. 21–24 . Tabl. 6.
Lycenchelys brevimaxillaris Toyoshima, 1985. P. 174. Figs. 29, 30, 31K (первоописание). Hatooka, 2002. P. 1033 (в ключе). Imamura et al., 2004. P. 84 (синонимия с L. melanostomias). Imamura et al., 2005. P. 2. Figs. 1–3. Tabl. 1.
Lycenchelys parini Федоров, 1995. С. 126. Рис. 1, 2 (первоописание). Kawarada et al., 2020. P. 32. Fig. 25 (сравнение с L. melanostomias, фотография голотипа плохого качества) (syn. nov.).
Материал. 2 экз. TL 56 и 103 мм (SL 54 и 100 мм), 50°31′ с.ш., 155°18′ в.д., НИС “Академик М. Келдыш”, рейс 11А, ст. 1413, проба № 5, трал Сигсби, 792–804 м, 23:00–23:35, 04.07.1986 г.
Диагноз. Вид рода Lycenchelys без свободных supraneuralia, с 22–25 туловищными позвонками, четырьмя–пятью посторбитальными порами, двумя (редко одной) порами в затылочной комиссуре, имеющий корональную пору, вентральную боковую линию (иногда различимую только в начальной части) и однотонно пигментированное тело. Чешуя на голове, на основании, перепонке и лучах Р отсутствует. Оперкулярная лопасть развита. D 112–120, A 98–107, V 3, vert. 117–125.
Описание. TL 103 мм. D 118, A 101, C 10, P 15, V 3; r. br 6, sp. br 12, pbr 3; vert. 22 + 99 = 121.
Тело тонкое, удлинённое, наиболее высокое у затылка (16.7 раза в SL), в туловищной части слабо, в хвостовой – сильно сжатое с боков (рис. 4). Голова 7.4 раза в SL, почти на всём протяжении равномерно высокая, лишь кпереди от орбит её дорсальный профиль слабо наклонный; вершина рыла круто обрезанная. Глаз эллиптический (его вертикальный диаметр в 1.4 раза меньше горизонтального), не достигает дорсального профиля головы, его горизонтальный диаметр в 1.4 раза превышает ширину костного межглазничного промежутка (рис. 5а, 5б). Трубочка ноздри короткая, немного не достигает верхнего края верхней губы. Рот умеренной величины, верхняя челюсть оканчивается на вертикали переднего края зрачка, задний конец ротовой щели немного не достигает вертикали переднего края глаза (примерно вдвое ближе к глазу, чем к заднему краю носовой трубочки). Губы мясистые, лопасти нижней губы доходят вперёд до уровня первой (подбородочной) пары мандибулярных пор (рис. 5в). Зубы на челюстях мелкие, конические: на praemaxillare однорядные, на dentale близ симфиза в два ряда, далее назад – в один; зубы на сошнике мелкие, конические, устроенные в поперечный ряд, числом восемь; на нёбных костях справа один, слева два ряда очень мелких трудноразличимых зубов. Мягкое нёбо с продольными рядами хорошо разобщённых, отчётливо выступающих мелких папилл. Жаберных тычинок на первой дуге 2 + 10, из них самая верхняя и самая нижняя слабо развиты; тычинки короткие и широкие, расстояние между наиболее развитыми тычинками соизмеримо с их длиной. Нижний конец жаберной щели слегка не достигает горизонтали нижнего конца основания Р. Оперкулярная лопасть вполне развита (рис. 5б).
В сейсмосенсорной системе головы (рис. 5а–5в) две передние надглазничные (носовые) поры, из которых первая расположена между ноздрёй и верхней губой, а вторая чуть позади основания трубочки ноздри, на одной вертикали с первой инфраорбитальной порой. Задняя надглазничная пора (по: Федоров, 1995. С. 127) или дополнительная пора (additional pore – по: Kawarada et al., 2020. P. 30) с обеих сторон отсутствует. Четыре заглазничные поры, из которых первая, расположенная у задневерхнего края глаза, отстоит от равнорасставленных последующих на промежуток в 1.5 раза больший, чем расстояние между второй и третьей заглазничной порами. Имеется непарная корональная пора, расположенная на уровне середины промежутка между вертикалями передних краёв глаза и зрачка. В затылочной комиссуре две поры. Инфраорбитальных пор восемь: в горизонтальной части подглазничного канала шесть очень крупных ноздревидных пор, в вертикальной (позади орбиты) – две более мелкие округлые поры. Преоперкуло-мандибулярных пор девять, большинство их тоже открывается очень крупными отверстиями (со второй по восьмую – ноздревидные), девятая (последняя) – самая мелкая, округлая. Восьмая пора преоперкуло-мандибулярного канала расположена на уровне шестой, девятая – чуть выше горизонтали верхнего края седьмой инфраорбитальной поры; пятая пора несколько развёрнута поперёк хода канала. Туловищная боковая линия (в виде поверхностного ряда невромастов) прослеживается только в самой начальной части туловища, довольно резко спускается от уровня верхнего края жаберного отверстия к середине длины прижатого к телу Р, далее назад неразличима.
Начало D немного отстоит от вертикали основания Р (примерно на величину интервентрального промежутка). Первый дорсальный птеригиофор расположен в промежутке между четвёртым и пятым невральными отростками, свободных supraneuralia нет, но около пяти передних лучей D укорочены и скрыты в коже. Плевральные рёбра развиты с третьего по 22-й позвонки; перед гемальным отростком 23-го позвонка три анальных птеригиофора, первый из них без луча (или его луч сильно редуцирован, на рентгенограмме не различим). Шестой–восьмой лучи Р наибольшие, вершины лучей с 9-го по 15-й утолщены и отчётливо выступают из межлучевой перепонки. Будучи прижатым вперёд, Р достигает своим концом вертикали середины глаза. V прикрепляются на середине промежутка между вертикалями нижнего конца жаберной щели и основания Р. Длина V в 1.1 раза превышает горизонтальный диаметр глаза. Чешуя на теле хорошо развита, отсутствует на голове, в предорсальной области, на истмусе, основании Р и вокруг V; на брюхе чешуя менее заметна и более разрежена, доходит вперёд до линии, соединяющей нижние концы основания Р. По среднебрюшной линии от уровня нижнего края основания Р до ануса проходит чёткая утолщённая светлая полоса, однако не образующая складки. Анус расположен у начала А. Два коротких супротивно расположенных пилорических придатка.
Измерения. В % SL: lc 13.5, wc 5.5, hc 5.5, H 6.0, HA 5.0, lР 9.0, lV 3.0, hР 3.2, lС 3.0, aD 15.0, aA 31.0, aV 10.0, ao 2.5, oo 2.8, ho 2.0, io 2.0, lmx 3.0, lbr 0.5, wbr 0.2, ibr 0.5, lba 4.5. В % lc: wc 40.7, hc 40.7, H 44.4, HA 37.0, lР 66.7, lV 22.2, hР 23.7, lС 22.2, aD 111.1, aA 229.6, aV 74.1, ao 18.5, oo 20.4, ho 14.8, io 14.8, lmx 22.2, lbr 3.7, wbr 1.5, ibr 3.7, lba 33.3.
Окраска фиксированного экземпляра светлая, кофейно-коричневая; нижняя поверхность головы, P, V и дистальные края непарных плавников тёмно-коричневые, выступающие утолщённые концы лучей нижней половины Р светлые (рис. 4); жаберно-ротовая полость и брюшина тёмные, верхняя сторона языка тёмная, нижняя – светлая; желудок и кишечник светлые с участками тёмно-коричневой пигментации.
Малёк TL 56 мм (рис. 5г, 5д). Число лучей в вертикальных плавниках и границу между туловищными и хвостовыми позвонками определить не удалось из-за недостаточной разрешающей способности рентгенограммы. Vert. 123. Тело более тонкое, его максимальная высота 21.6 раза укладывается в SL. Голова 7.2 раза в SL, дорсальный профиль рыла более пологий, чем у бóльшего экземпляра. Глаз очень большой, сильно вытянутый продольно (его горизонтальный диаметр в 1.8 раза превышает вертикальный), костный межглазничный промежуток узкий, в 5.5 раза меньше горизонтального диаметра глаза. Нёбные зубы однорядные. С правой стороны имеется хорошо развитая дополнительная (задняя надглазничная) пора (слева отсутствует). Корональная пора расположена на уровне переднего края зрачка. Инфраорбитальных пор девять (в горизонтальном отделе семь пор). Первый дорсальный птеригиофор расположен между третьим и четвёртым невральными отростками. Отогнутый вперёд Р немного заходит своим концом за передний край зрачка. Чешуя и боковая линия не различимы. Остальные признаки этого малька соответствуют описанным для экземпляра TL 103 мм.
Измерения. В % SL: lc 13.9, wc 4.6, hc 4.3, H 4.6, HA 3.2, lР 12.0, lV 3.7, hР 3.2, lС 3.7, aD 15.7, aA 29.6, aV 11.1, ao 2.8, oo 5.1, ho 2.8, io 0.9, lmx 4.6. В % lc: wc 33.3, hc 30.7, H 33.3, HA 23.3, lР 86.7, lV 26.7, hР 23.3, lС 26.7, aD 113.3, aA 213.3, aV 80.0, ao 20.0, oo 36.7, ho 20.0, io 6.7, lmx 33.3.
Сравнительные замечания. Идентификация описанных экземпляров требует обсуждения реальности различий между номинальными видами L. parini и L. melanostomias, так как они с равным основанием могут быть отнесены и к тому, и к другому виду. Федоров (1995) описал L. parini по единственному экземпляру TL 164 мм, пойманному у тихоокеанского побережья о-ва Итуруп (Курильские о-ва) на глубине 800 м. В первоописании в качестве уникальной особенности L. parini он указал наличие пары задних заглазничных пор, предположив, однако, что такое же строение сейсмосенсорной системы имеет L. melanostomias. Действительно, у одного из 42 исследованных экземпляров L. melanostomias также была обнаружена пара заглазничных пор (Kawarada et al., 2020. Fig. 22D ). Из исследованных мною рыб у более крупного экземпляра эти поры отсутствуют, а у меньшего задняя заглазничная пора присутствует лишь с правой стороны. Это позволяет предполагать, что наличие данных добавочных пор является редкой аберрацией и не имеет диагностического значения.
Следует также оговорить сходство между указанными номинальными видами по положению пятой и шестой преоперкуло-мандибулярных пор. Действительно, на рисунке голотипа L. melanostomias (Toyoshima, 1983. Figs. 25, 27 . Pl. 157; 1985. Fig. 27B ) продольные диаметры отверстий этих пор развёрнуты поперёк простирания самого канала, как и у голотипа L. parini, однако на рисунке L. brevimaxillaris (Toyoshima, 1985. Fig. 30 С) эти поры изображены в обычном положении. Конспецифичность L. brevimaxillaris и L. melanostomias была убедительно показана Имамурой с соавторами (Imamura et al., 2004, 2005). У изученных рыб только отверстие пятой поры несколько развёрнуто поперёк хода канала, подобно тому, как это изображено для L. melanostomias (Kawarada et al., 2020. Fig. 22A ). По-видимому, изменчивость этого признака существенно ограничивает его диагностическое значение.
Федоров (1995) привёл семь признаков, по которым можно отличить L. parini от L. melanostomias, однако после переописания последнего вида и сведения в его синонимы L. brevimaxillaris только первый из этих признаков (положение боковой линии) сохранил своё значение, тогда как остальные полностью уложились в пределы индивидуальной изменчивости японских популяций L. melanostomias (Imamura et al., 2004, 2005; Kawarada et al., 2020). Указанная для L. parini медиолатеральная боковая линия действительно резко отличает этот вид от L. melanostomias, имеющего вентральную боковую линию. Однако корректность оценки данного признака у голотипа L. parini вызывает сомнение. По моей просьбе М.В. Назаркин (ЗИН РАН) изучил боковую линию у голотипа L. parini и обнаружил следующее: “боковая линия практически неразличима – видны передние невромасты, идущие от задней посторбитальной поры вниз сразу позади основания Р, за ним различимы три–четыре невромаста в линии, направленной вниз и назад” (личное сообщение 23.12.2020 г.). Таким образом, положение боковой линии у голотипа L. parini ничем не отличается от наблюдаемого у исследованных мной экземпляров и от изображения, приведённого Тоёшимой (Toyoshima, 1985. Fig. 31K ) для L. brevimaxillaris, и нет никаких доказательств, что она действительно была медиолатеральной.
Наконец, комментариев требует ещё одна характеристика – пигментация желудка, по которой Тоёшима (Toyoshima, 1985) различал свои номинальные виды L. brevimaxillaris и L. melanostomias (светлая у первого и тёмная у второго). Имамура с соавторами (Imamura et al., 2004) указали, что и у L. melanostomias желудок светлый. В первоописании L. parini этот признак не охарактеризован, по наблюдениям М.В. Назаркина (личное сообщение 23.12.2020 г.), желудок, по-видимому, не пигментирован, очень слабые следы пигментации прослеживаются лишь в пилорической области. У исследованного экземпляра TL 103 мм (малёк TL 56 мм не вскрывался) желудок с отдельными участками коричневой пигментации, и можно ожидать значительную индивидуальную изменчивость в развитии таких участков.
Таким образом, не находится ни одного признака, по которому можно было бы уверенно разделить номинальные виды L. parini и L. melanostomias, и признаки голотипа первого вида и описанных здесь экземпляров от Северных Курил полностью укладываются в характеристику L. melanostomias из вод Японии в последних работах по этому виду (Imamura et al., 2004, 2005; Kawarada et al., 2020). На этом основании я считаю указанные номинальные виды конспецифичными.
Малёк TL 56 мм заметно отличается по ряду габитуальных особенностей от более крупной рыбы. Насколько мне известно, столь мелкие экземпляры лиценхелов в литературе никогда не описывались. Исследованный малёк характеризуется меньшей длиной головы, меньшей высотой головы и тела, более длинными парными плавниками и в особенности более крупным, продольно вытянутым глазом с очень узким межглазничным промежутком по сравнению с экземпляром TL 103 мм. Чешуя у него ещё не выражена. Вместе с тем по развитию озубления и пор сейсмосенсорной системы малёк TL 56 мм не имеет существенных отличий от более крупных рыб. Поры сейсмосенсорных каналов несколько мельче, а их просветы, наоборот, шире, чем у экземпляра TL 103 мм, и промежуток между затылочными порами заметно более широкий (рис. 5а, 5г), но в остальном отличий не наблюдается. Интересно отметить, что дополнительная (задняя надглазничная) пора появляется уже при столь малых размерах – это позволяет заключить, что даже непостоянные поры у лиценхелов формируются очень рано.
Ранее L. melanostomias указывался только от побережья Японии, где он известен для вод у охотоморского и тихоокеанского побережья о-ва Хоккайдо от префектуры Аомори до префектуры Ибараки на глубинах 425–1440 м (Kawarada et al., 2020). Голотип L. parini был пойман в Курило-Камчатской впадине близ о-ва Итуруп (44°39′ с.ш., 149°02′ в.д.) на глубине 800 м. Новая находка, сделанная у охотоморского побережья о-ва Парамушир, расширяет известный ареал вида далее к северу и позволяет предполагать, что он может населять воды как охотоморского, так и тихоокеанского побережья Курильской гряды на всем её протяжении.
Список литературы
Андрияшев А.П. 1955. Обзор угревидных ликодов [Lycenchelys Gill (Pisces, Zoarcidae) и близкие формы] морей СССР и сопредельных вод // Тр. ЗИН АН СССР. Т. 18. С. 349–384.
Андрияшев А.П. 1958. Добавление к обзору угревидных ликодов (Lycenchelys Gill) с описанием трех новых видов из Курило-Камчатской впадины // Вопр. ихтиологии. Вып. 11. С. 171–180.
Андрияшев А.П. 1979. О первом нахождении рыб семейств Zoarcidae и Liparidae у острова Кергелен // Биология моря. № 6. С. 28–34.
Федоров В.В. 1995. Lycenchelys parini sp. nova (Perciformes: Zoarcidae) с батиальных глубин Курило-Камчатской впадины // Вопр. ихтиологии. Т. 35. № 1. С.126–129.
Федоров В.В., Андрияшев А.П. 1993. Lycenchelys makushok sp. nova (Perciformes, Zoarcidae) с батиальных глубин Курило-Камчатской впадины // Там же. Т. 33. № 1. С. 133–136.
Шейко Б.А., Федоров В.В. 2000. Класс Cephalaspidomorphi – Миноги. Класс Chondrichthyes – Хрящевые рыбы. Класс Holocephali – Цельноголовые. Класс Osteichthyes – Костные рыбы // Каталог позвоночных Камчатки и сопредельных морских акваторий. Петропавловск-Камчатский: Камчат. печат. двор. С. 7–69.
Anderson M.E. 1982a. A new eelpout (Teleostei: Zoarcidae) from the eastern tropical Pacific Ocean // Bull. Mar. Sci. V. 32. № 1. C. 207–212.
Anderson M.E. 1982b. Revision of the fish genera Gymnelus Reinhardt and Gymnelopsis Soldatov (Zoarcidae), with two new species and comparative osteology of Gymnelus viridis // Natl. Mus. Can. Publ. Zool. № 17. P. 1–76.
Anderson M.E. 1988. Studies on the Zoarcidae (Teleostei: Perciformes) of the southern hemisphere: I. The Antarctic and subantarctic regions // Biology of the Antarctic Seas XIX. Antarctic Res. Ser. V. 47. P. 59–113. https://doi.org/10.1029/AR047p0059
Anderson M.E. 1994. Systematics and osteology of the Zoarcidae (Teleostei: Perciformes) // Ichthyol. Bull. J.L.B. Smith Inst. Ichthyol. № 60. 120 p.
Anderson M.E. 1995. The eelpout genera Lycenchelys Gill and Taranetzella Andriashev (Teleostei: Zoarcidae) in the eastern Pacific, with descriptions of nine new species // Proc. Calif. Acad. Sci. V. 49. № 2. P. 55–113.
Fricke R., Eschmeyer W.N., van der Laan R. (eds.) 2021. Eschmeyer’s catalog of fishes: genera, species, references (http://researcharchive.calacademy.org/research/ichthyology/catalog/ fishcatmain.asp. Version 03/2021).
Geistdoerfer P., Hureau J.C., Rannou M. 1970. Deux poissons abyssaux nouveaux capturés dans l’atlantique nord et est: Bathytyphlops azorensis n. sp. (Ipnopidae) et Lycenchelys labradorensis n. sp. (Zoarcidae) // Bull. Mus. Natl. Hist. Nat. Ser. 2. V. 42. № 3. P. 452–459.
Hatooka K. 2002. Zoarcidae // Fishes of Japan with pictorial keys to the species. Tokyo: Tokai Univ. Press. P. 1028–1044, 1581–1583.
Imamura H., Machida Y., Ohta S. 2004. Lycenchelys melanostomias Toyoshima, 1983, a senior synonym of L. brevimaxillaris Toyoshima, 1985 (Teleostei: Zoarcidae) // Ichthyol. Res. V. 51. № 1. P. 84–88. https://doi.org/10.1007/s10228-003-0194-2
Imamura H., Shinohara G., Shiogaki M. 2005. Rediscovery of the type specimens of Lycenchelys brevimaxillaris (Actinopterygii: Perciformes: Zoarcidae) with taxonomic notes on the species // Species Divers. V. 10. № 1. P. 1–6. https://doi.org/10.12782/specdiv.10.1
Kawarada S., Imamura H., Narimatsu Y., Shinohara G. 2020. Taxonomic revision of the genus Lycenchelys (Osteichthyes: Zoarcidae) in Japanese waters // Zootaxa. V. 4762. № 1. P. 1–66. https://doi.org/10.11646/zootaxa.4762.1.1
Møller P.R. 1999. Taxonomic status of the western North Atlantic eelpout, Lycenchelys paxillus (Goode & Bean, 1879) (Pisces, Zoarcidae) // Ichthyol. Res. V. 46. № 3. P. 323–328.
Peden A.E., Anderson M.E. 1978. A systematic review of the fish genus Lycodapus (Zoarcidae) with description of two new species // Can. J. Zool. V. 56. № 9. P. 1925–1961. https://doi.org/10.1139/z78-262
Thiel R., Knebelsberger T., Eidus I. 2018. Description and DNA barcoding of Lycenchelys lenzeni, a new species of eelpout (Perciformes: Zoarcidae) from the deep sea off the Kuril Archipelago // Zootaxa. V. 4370. № 1. P. 45–56. https://doi.org/10.11646/zootaxa.4370.1.3
Toyoshima M. 1983. Zoarcidae // Fishes from the North-Eastern seas of Japan and Okhotsk Sea off Hokkaido. The intensive research of unexploited fishery resources on continental slopes. Tokyo: JFRCA. P. 136–149, 208–210, 258–277, 329–335.
Toyoshima M. 1985. Taxonomy of the subfamily Lycodinae (family Zoarcidae) in Japan and adjacent waters // Mem. Fac. Fish. Hokkaido Univ. V. 32. № 2. P. 131–243.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии