

Вопросы ихтиологии, 2022, T. 62, № 3, стр. 296-302
Оценка видового разнообразия ихтиопланктона северной части Центрально-Восточной Атлантики
А. Г. Архипов 1, *, Ф. В. Коломейко 1, Р. А. Пак 1
1 Атлантический филиал Всероссийского научно-исследовательского института
рыбного хозяйства и океанографии − АтлантНИРО
Калининград, Россия
* E-mail: arkhipov@atlantniro.ru
Поступила в редакцию 20.04.2021
После доработки 03.05.2021
Принята к публикации 11.05.2021
- EDN: NKCOYY
- DOI: 10.31857/S0042875222020011
Аннотация
По материалам многолетних (2000–2019) исследований ихтиопланктона в смежных районах северной части Центрально-Восточной Атлантики (исключительные экономические зоны северной и южной частей Марокко и Мавритании) проанализировано видовое разнообразие ихтиопланктона с помощью индексов, широко применяемых в эколого-фаунистических исследованиях. Выявлена общая тенденция увеличения видового разнообразия от северной части Марокко к району Мавритании (средние значения индекса Маргалефа повышаются от 4.63 до 7.02). Ихтиопланктонные комплексы рассматриваемых районов характеризуются средним и высоким биотическим разнообразием (индекс Симпсона – 0.22–0.90); разная степень их монодоминантности наблюдается чаще (в 25 из 44 наблюдений индекс выравненности Пиелу ≤ 0.5), чем относительно равное обилие всех видов. В северной части Марокко динамика индексов видового разнообразия близка к таковой индексов численности европейской сардины Sardina pilchardus ранних стадий развития, в южной части Марокко и в Мавритании – с индексами численности икринок и личинок круглой сардинеллы Sardinella aurita. В ихтиопланктоне рассматриваемых районов доминирующим видом является S. pilchardus, субдоминантами – S. aurita, европейский анчоус Engraulis encrasicolus и спаровые (Sparidae).
Природные сообщества, в том числе и морские, значительно различаются по числу видов. Знания лишь качественного состава организмов не позволяют оценить многие экологические особенности сообщества. Поэтому, как правило, видовое разнообразие представляют в виде индексов, позволяющих оценить различия в биоценозах из разных районов, в разные сезоны или оценить изменения, происходящие в экосистеме под влиянием загрязнения или какого-либо другого фактора, приводящего к стрессовому состоянию сообщества (Cameron, 1972; Willson, 1974; Margalef, 1975; Odum, 1975; Серебряков, 1984; Архипов, 2006). Видовое разнообразие в сообществах также рассматривают как функцию, учитывающую не только число видов, но и выравненность распределения особей по видам (Песенко, 1982).
На выживание икринок и личинок, а соответственно, и на формирование численности будущих поколений влияют такие факторы, как температура, солёность, содержание в воде кислорода и биогенов, развитие кормовой базы и другие. Интегрированным показателем состояния любого сообщества может служить его видовое разнообразие (Jacobs, 1975; Песенко, 1982; Серебряков, 1984). Пианки (1980) отмечал, что на практике часто бывает трудно изучить видовое разнообразие всего сообщества, и поэтому внимание уделяют какой-либо его части, тем более что использование разнообразия видов имеет смысл только для систем со слабо взаимодействующими элементами. По мнению Свирежева (1977), для характеристики экологических сообществ видовое разнообразие более или менее подходит лишь при анализе ранних стадий развития, когда виды слабо конкурируют друг с другом. Икринки и личинки рыб, составляющие ихтиопланктонные комплексы, в наибольшей степени соответствуют этим условиям. Кроме того, видовой состав ихтиопланктона и численность его компонентов отражают в определённой степени условия воспроизводства и выживания рыб. Поэтому особый интерес представляет изучение видового разнообразия ихтиопланктонных комплексов. В ихтиопланктонных пробах, собранных на стандартных станциях в северной части Центрально-Восточной Атлантики (ЦВА), число видов рыб и численность их икринок и личинок от года к году существенно менялись. Видовое разнообразие ихтиопланктона было оценено с помощью разных индексов, широко применяемых в эколого-фаунистических исследованиях (Jacobs, 1975; Пианки, 1980; Федоров, Гильманов, 1980; Песенко, 1982; Серебряков, 1984; Одум, 1986; Бигон и др., 1989; Бурковский, 2006).
Цель предлагаемой работы – проанализировать видовое разнообразие ихтиопланктона с помощью разных индексов на основе материалов многолетних исследований ранних стадий развития рыб в смежных районах северной части ЦВА (с использованием ретроспективных данных и новых материалов, а также базы данных АтлантНИРО по ихтиопланктону океанических районов).
МАТЕРИАЛ И МЕТОДИКА
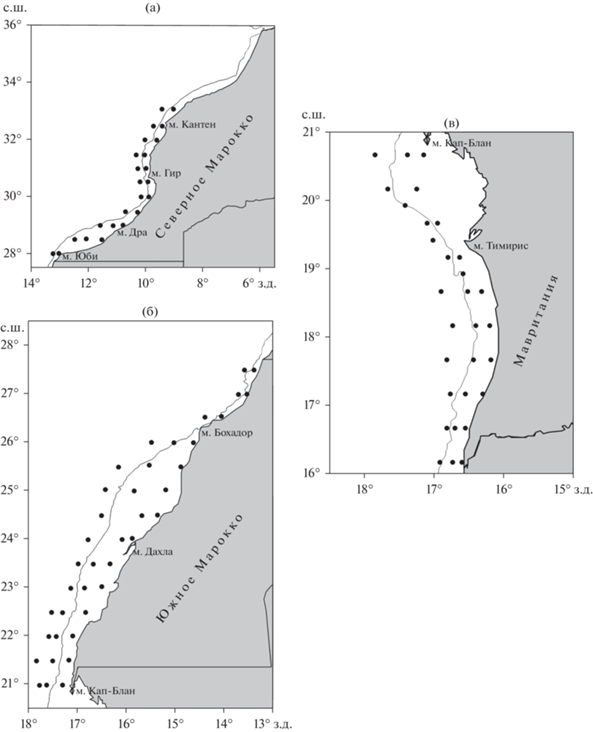
В работе проанализированы данные 25 ихтиопланктонных съёмок, выполненных в 2000−2019 гг. в водах исключительных экономических зон северной (32°–28° с.ш.) и южной (28°–21° с.ш.) частей Марокко (рис. 1а, 1б) и Мавритании (21°–16° с.ш.) (рис. 1в). Планктонные станции охватывали акваторию над глубинами от 20 до 1000 м. Ступенчато-косой лов планктоносборщиком “Бонго-20” (газ № 17–21) выполняли на горизонтах 100, 50, 35, 25, 10 и 0 м по 1.5–3.0 мин на каждом горизонте при скорости судна 2–3 узла (Smith, Richardson, 1977; Методические указания …, 1983). Собранные пробы обрабатывали в лабораторных условиях под бинокулярными микроскопами МБС-10 (увеличение 8 × 2, 8 × 4). В ходе камеральной обработки определяли количественный и качественный составы икринок и личинок рыб.
Рис. 1.
Сетки станций (⚫) в северной (а) и южной (б) частях Марокко и в Мавритании (в); (—) – изобата 200 м.
Видовое разнообразие ихтиопланктона оценено с помощью наиболее распространённого в экологии информационно-статистического императивного индекса Шеннона (Shannon, 1948; Алимов, 1989):
где ni – число особей i-того вида, экз.; N – общее число особей, экз., W – общее число видов.Хотя индекс Шеннона учитывает степень выравненности видов, очень часто для наглядной оценки степени доминирования в биоценозах разных видов отдельно приводят показатель этой выравненности (императивный индекс), предложенный Пиелу (Pielou, 1966) и представляющий собой отношение наблюдаемого разнообразия к максимальному: E = HS/log2 W, где W – общее число видов. Значения этого индекса варьируют в пределах от 0 до 1. Когда индекс стремится к 0, наблюдается ситуация преобладания одного–двух видов в сообществах с относительно упрощённой организацией, а когда он приближается к 1, отмечается равное обилие многих видов в сравнительно сложно организованных сообществах (Бурковский, 2006). Кроме того, были рассчитаны императивный индекс Симпсона (Simpson, 1949) − $D = 1 - \sum\nolimits_{i - 1}^w {\frac{{{{n}_{i}}({{n}_{i}} - 1)}}{{N(N - 1)}}} $, а также параметрический индекс Маргалефа (Margalef, 1951) − d = (W – 1)/ln N. Императивные индексы в основном заимствованы из теории информации и имеют то преимущество перед параметрическими, что применимы даже тогда, когда форма распределения видового разнообразия по станциям и/или по годам значительно меняется. Параметрические же индексы показывают наличие большего или меньшего числа видов в сравниваемых биоценозах вне зависимости от численности каждого вида.
Связи видового разнообразия с разными характеристиками экосистемы проанализированы по методике, предложенной Одумом (Odum, 1975). Он выделил группы с низким биотическим разнообразием (D в пределах 0.05–0.20), группы с высоким биотическим разнообразием (D > 0.70) и промежуточные группы со средним биотическим разнообразием (D – 0.21–0.70). В первую группу попадают экосистемы, находящиеся в состоянии стресса (например, в случае загрязнения), а также экосистемы, активно регулируемые человеком (например, агроценозы); во вторую − сравнительно устойчивые экосистемы, не получающие извне концентрированную энергию и биогенные материалы. Большинство же экосистем, как правило, находятся в промежуточной группе.
Для оценки вклада каждого вида в состав ихтиопланктонных комплексов применён индекс доминирования Палия−Ковнацки (Kownacki, 1971): Di = = 100mi/Mni/N, где Di – индекс доминирования для i-того вида, %; mi − число проб, в которых был найден вид i; M – общее число проб, ni – число особей i-того вида, экз.; N – общее число особей, экз.
Индексы рассчитывали с помощью соответствующего модуля географической информационной системы (ГИС) АтлантНИРО (Коломейко, Сердобинцев, 2019).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Исследователи применяют индексы видового разнообразия для самых разных целей: для изучения степени загрязненности водоёмов (Винберг, 1976; Снетков, Вавилин, 1977), для сравнения изменений видового разнообразия фито-, зоо-, ихтиопланктона и зообентоса (Гиляров, Горелов, 1974; Серебряков, 1984; Архипов, 2006; Лидванов, 2014; Дмитриева, 2017; Климова и др., 2017, 2019; Гусев, 2020), для анализа выравненности видовых обилий и т.д. (Виленкин, 1966; Pielou, 1966; Wilhm, 1967). Обобщая многочисленные данные литературы по исследованию индексов видового разнообразия, Джейкобс (Jacobs, 1975) выделил ряд факторов, которые непосредственно влияют на разнообразие экологических сообществ. Он утверждал, что одним из таких факторов являются стрессовые условия среды, которые обычно отрицательно коррелируют с видовым разнообразием.
Выбор того или иного индекса видового разнообразия обосновывается и теоретически, и эмпирически, однако, как отмечал Ллойд (Lloyd, 1968), тот индекс лучше, который, будучи применимым для практических целей, даёт наиболее достоверные предсказания экологических изменений и наилучшее их объяснение. Рассмотрим динамику индексов видового разнообразия ихтиопланктона северной части ЦВА по трём исследуемым районам в 2000–2019 гг.
В северной части Марокко все индексы изменялись синхронно, причём наибольшие колебания наблюдались у параметрического индекса Маргалефа – 2.79–5.90 (в среднем 4.63) (рис. 2а). Изменения императивных индексов были более сглаженными. Индекс Симпсона менялся в пределах 0.53–0.83, следовательно, ихтиопланктонный комплекс северной части Марокко имел среднее и высокое биотическое разнообразие. Показатель выравненности Пиелу варьировал в диапазоне 0.41–0.65, т.е. иногда наблюдалась ситуация определённой степени монодоминантности (в шести из 15 лет наблюдений Е ≤ 0.5), но чаще отмечалось равное обилие всех видов в сложно организованном ихтиопланктонном комплексе.
Рис. 2.
Изменения индексов видового разнообразия ихтиопланктонных комплексов в смежных районах северной части Центрально-Восточной Атлантики: а − северная часть Марокко, б – южная часть Марокко, в – Мавритания; (- ◼ -) – индекс Маргалефа, (- × -) – индекс Симпсона, (–▲–) – индекс Пиелу, (–⚫–) – индекс Шеннона.
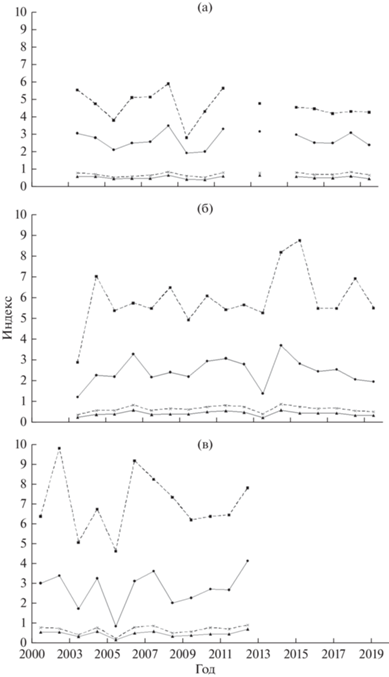
В рассматриваемом районе преобладает субтропическая ихтиофауна. Отмеченное в 2005–2008 и 2015–2019 гг. увеличение индексов видового разнообразия ихтиопланктона (рис. 2а) согласуется с повышением индексов численности особей ранних стадий развития массового субтропического вида – европейской сардины Sardina pilchardus (Архипов, Пак, 2019).
В южной части Марокко, как и в северной, все индексы изменялись синхронно; но значения параметрического индекса Маргалефа были выше, а диапазон варьирования шире – 2.88–8.76 (5.92) (рис. 2б). Изменения императивных индексов были менее выраженными. Индекс Симпсона менялся в пределах 0.35–0.87, т.е. ихтиопланктонный комплекс южной части Марокко имел среднее и высокое биотическое разнообразие. Судя по значениям показателя выравненности Пиелу (0.25–0.59), чаще наблюдалась ситуация в определённой степени монодоминантности (в 12 из 17 лет наблюдений), а в некоторые годы отмечалось равное обилие всех видов в относительно сложно организованном ихтиопланктонном комплексе.
В данном районе смешиваются субтропическая и тропическая ихтиофауны. Увеличение индексов видового разнообразия ихтиопланктона в 2008 и в 2014–2015 гг. (рис. 2б) совпадает с повышением индексов численности ранних стадий развития массового тропического вида – круглой сардинеллы Sardinella aurita (Архипов, Пак, 2019). Тогда как у субтропической европейской сардины тенденция сходных колебаний этих показателей явно не прослеживается.
В районе Мавритании все рассматриваемые индексы также изменялись синхронно, колебания императивных индексов были более сглаженными, чем параметрического (рис. 2в). По сравнению с северной и южной частями Марокко значения индекса Маргалефа были больше, а диапазон варьирования наиболее значительным − 4.64–9.81 (7.02). Индекс Симпсона изменялся в пределах 0.22–0.90, т.е. ихтиопланктонный комплекс в районе Мавритании имел среднее и высокое биотическое разнообразие. Исходя из показателя выравненности Пиелу (0.15–0.70), здесь, как и в южной части Марокко, в основном наблюдалась ситуация определённой монодоминантности (в семи из 12 лет наблюдений), в остальные годы отмечалось равное обилие всех видов в сравнительно сложно организованном ихтиопланктонном комплексе.
В районе Мавритании преобладает тропическая ихтиофауна. Наиболее значимое увеличение индексов видового разнообразия ихтиопланктона в 2001 и в 2010–2012 гг. (рис. 2в) согласуется с высокими индексами численности в эти годы икринок и личинок массового тропического вида – круглой сардинеллы (Архипов, 2015).
Анализируя изменения индексов видового разнообразия по трём рассматриваемым акваториям, можно отметить общую тенденцию увеличения видового разнообразия от северной части Марокко к району Мавритании (средние значения индекса Маргалефа увеличиваются от 4.63 до 7.02), что подтверждает выводы наших более ранних исследований (Архипов, 2015; Архипов, Пак, 2019). Ихтиопланктонные комплексы рассматриваемых районов, оцениваемые по колебаниям индекса Симпсона (0.22–0.90), согласно классификации Одума (Odum, 1975), имели среднее и высокое биотическое разнообразие. Показатель выравненности Пиелу свидетельствует, что чаще наблюдалась определённая степень монодоминантности (в 25 из 44 наблюдений Е ≤ 0.5), реже отмечалось равное обилие всех видов в относительно сложно организованных ихтиопланктонных комплексах. Отметим, что повышение и понижение значений рассматриваемых индексов в трёх исследуемых районах по годам происходило не синхронно (рис. 2).
В северной части Марокко наблюдались одинаковые тенденции динамики индексов разнообразия и индексов численности у европейской сардины, в южной части Марокко и в Мавритании – у круглой сардинеллы. Следовательно, можно заключить, что индексы видового разнообразия в определённой степени отражают условия воспроизводства и выживания массовых видов рыб на ранних стадиях развития.
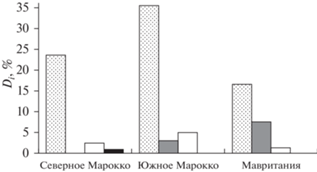
Согласно значениям индекса Палия−Ковнацки доминирующим видом в ихтиопланктоне в рассматриваемых районах является европейская сардина, субдоминантами – круглая сардинелла, европейский анчоус Engraulis encrasicolus и спаровые (Sparidae) (рис. 3). Причём в составе ихтиопланктона доля доминанта (европейская сардина, субтропический вид) уменьшается, а доля субдоминанта (круглая сардинелла, тропический вид) увеличивается от Марокко к Мавритании.
Рис. 3.
Индексы доминирования Палия–Ковнацки (Di) массовых видов рыб в ихтиопланктонных комплексах Северного и Южного Марокко и Мавритании:
( ) − европейская сардина Sardina pilchardus, (
) − европейская сардина Sardina pilchardus, ( ) – круглая сардинелла Sardinella aurita, (◻) − спаровые (Sparidae), (◼) − европейский анчоус Engraulis encrasicolus.
) – круглая сардинелла Sardinella aurita, (◻) − спаровые (Sparidae), (◼) − европейский анчоус Engraulis encrasicolus.
ВЫВОДЫ
1. Наблюдается общая тенденция увеличения видового разнообразия ихтиопланктонных комплексов от северной части Марокко к району Мавритании: средние значения индекса Маргалефа повышаются от 4.63 до 7.02.
2. В рассматриваемых районах ихтиопланктонные комплексы имеют среднее и высокое биотическое разнообразие (индекс Симпсона − 0.22–0.90).
3. Разная степень монодоминантности ихтиопланктонных комплексов (индекс выравненности Пиелоу ≤ 0.5) наблюдается чаще, чем относительно равное обилие всех видов.
4. В северной части Марокко динамика индексов видового разнообразия ихтиопланктонных комплексов близка к таковой индексов численности европейской сардины ранних стадий развития, в южной части Марокко и в Мавритании − к индексам численности икринок и личинок круглой сардинеллы.
5. В ихтиопланктоне рассматриваемых районов доминирующим видом является европейская сардина, субдоминантами – круглая сардинелла, европейский анчоус и спаровые.
Список литературы
Алимов А.Ф. 1989. Введение в продукционную гидробиологию. Л.: Гидрометеоиздат, 152 с.
Архипов А.Г. 2006. Динамика численности и особенности распределения ихтиопланктонных сообществ северной части центрально-восточной Атлантики и морей Средиземноморского бассейна. Калининград: Изд-во АтлантНИРО, 232 с.
Архипов А.Г. 2015. Динамика численности икринок и личинок массовых видов рыб северной части центрально-восточной Атлантики // Вопр. ихтиологии. Т. 55. № 2. С. 173–179. https://doi.org/10.7868/S0042875215020010
Архипов А.Г., Пак Р.А. 2019. Динамика численности массовых видов ихтиопланктона в водах Марокко // Там же. Т. 59. № 3. С. 278–285. https://doi.org/10.1134/S0042875219020012
Бигон М., Харнер Дж., Таунсенд К. 1989. Экология. Особи, популяции и сообщества. Т. 2. М.: Мир, 480 с.
Бурковский И.В. 2006. Морская биогеоценология. Организация сообществ и экосистем. М.: Т-во науч. изд. КМК, 285 с.
Виленкин Б.Я. 1966. Кибернетический и энергетический подход к проблемам биоценологии // Экология водных организмов. М.: Наука. С. 66–77.
Винберг Г.Г. 1976. Итоги исследований пресноводных сообществ всех трофических уровней // Ресурсы биосферы. Вып. 2. С. 145–157.
Гиляров А.М., Горелова Т.А. 1974. Корреляция между трофической структурой, видовым разнообразием и биомассой зоопланктона северных озёр // Зоол. журн. Т. 53. Вып. 1. С. 25–33.
Гусев А.А. 2020. Многолетняя динамика макрозообентоса в юго-восточной части Балтийского моря: Автореф. дис. … канд. биол. наук. Борок: ИБВВ РАН, 26 с.
Дмитриева О.А. 2017. Исследование закономерностей пространственно-временных изменений структурных и количественных показателей фитопланктона в различных районах Балтийского моря: Автореф. дис. … канд. биол. наук. М.: ВНИРО, 24 с.
Климова Т.Н., Вдодович И.В., Игнатьев С.М. и др. 2017. Состояние ихтиопланктона в устьевой части Севастопольской бухты (Черное море) // Журн. Сиб. федерал. ун-та. Сер. биол. Т. 10. № 1. С. 74–86.
Климова Т.Н., Субботин А.А., Мельников В.В. и др. 2019. Пространственное распределение ихтиопланктона у крымского полуострова в летний сезон 2013 г. // Мор. биол. журн. Т. 4. № 1. С. 63–80. https://doi.org/10.21072/mbj.2019.04.1.06
Коломейко Ф.В., Сердобинцев С.П. 2019. Системы поддержки принятия решений в исследованиях и промысле водных биоресурсов // Изв. КГТУ. № 54. С. 51–60.
Лидванов В.В. 2014. Мезозоопланктон в районе Канарского апвеллинга: Автореф. дис. … канд. биол. наук. СПб.: ЗИН РАН, 26 с.
Методические указания по сбору проб зоо- и ихтиопланктона планктоносборщиком “Бонго” и их обработке. 1983. Калининград: Изд-во АтлантНИРО, 36 с.
Одум Ю. 1986. Экология. Т. 2. М.: Мир, 376 с.
Песенко Ю.А. 1982. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука, 288 с.
Пианки Э. 1980. Эволюционная экология. М.: Мир, 400 с.
Свирежев Ю.М. 1977. Математические модели биологических сообществ // Число и мысль. Вып. 1. М.: Знание, 158 с. http://mathemlib.ru/books/item/f00/s00/z0000015/st005.shtml.
Серебряков В.П. 1984. Размножение и ранний онтогенез промысловых рыб Северной Атлантики: Автореф. дис. … докт. биол. наук. М.: ВНИРО, 48 с.
Снетков М.А., Вавилин В.А. 1977. Оценка загрязнения водоёмов по интегральным показателям качества воды // Научные основы контроля качества поверхностных вод по гидробиологическим показателям. Л.: Гидрометеоиздат. С. 65–78.
Федоров В.Д., Гильманов Т.Г. 1980. Экология. М.: Изд-во МГУ, 464 с.
Cameron G.N. 1972. Analysis of insect trophic diversity in two salt marsh communities // Ecology. V. 53. № 1. P. 58–73. https://doi.org/10.2307/1935710
Jacobs I. 1975. Diversity stability and maturity in ecosystems influenced by human activities // Unifying concepts in ecology / Eds. van Dobben W.H., Lowe-McConnell R.H. Dordrecht: Springer. P. 187–207. https://doi.org/10.1007/978-94-010-1954-5_15.
Kownacki A. 1971. Taxocens of Chironomidae in streams of the Polish Hight Tatra, Mts // Acta Hydrobiol. V. 13. № 2. P. 439−463.
Lloyd S.K. 1968. On the calculation of diversity // Amer. Midland Naturalist. V. 79. P. 257–272.
Margalef R. 1951. Diversidad de expecies en las comunidades naturales // Pablnes. Inst. Biol. Apl. Barcelona. V. 9. № 5. P. 5–28.
Margalef R. 1975. Diversity stability and maturity in natural ecosystems // Unifying concepts in ecology / Eds. van Dobben W.H., Lowe-McConnell R.H. Dordrecht: Springer. P. 151–160. https://doi.org/10.1007/978-94-010-1954-5_12.
Odum E.P. 1975. Diversity as a function of energy flow // Ibid. P. 11–14. https://doi.org/10.1007/978-94-010-1954-5_1
Pielou E.C. 1966. Species-diversity and pattern-diversity in the study of ecological succession // J. Theor. Biol. V. 10. № 2. P. 370–383. https://doi.org/10.1016/0022-5193(66)90133-0
Shannon C.E. 1948. A mathematical theory of communication // Bell. System Tech. J. V. 27. P. 623–656.
Simpson E.H. 1949. Measurement of diversity // Nature. V. 163. P. 688. https://doi.org/10.1038/163688a0
Smith P.E., Richardson S.L. 1977. Standard techniques for pelagic fish egg and larvae surveys // FAO Fish. Tech. Pap. № 175. 100 p.
Wilhm I.L. 1967. Comparison of some diversity indices applied to populations of benthic macro invertebrates in a stream receiving organic wastes // J. Water Pollut. Control Federation. V. 39. № 10. P. 1673–1683.
Willson M.F. 1974. Avian community organization and habitat structure // Ecology. V. 55. № 5. P. 1017–1030. https://doi.org/10.2307/1940352
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии