

Вопросы ихтиологии, 2022, T. 62, № 3, стр. 272-280
Сравнительный анализ структуры различий между некоторыми морфотипами крупных африканских усачей рода Barbus (Labeobarbus auctorum) из озера Тана, Эфиопия
М. В. Мина 1, *, А. Н. Мироновский 2
1 Институт биологии развития РАН − ИБР РАН
Москва, Россия
2 Институт проблем экологии и эволюции РАН − ИПЭЭ РАН
Москва, Россия
* E-mail: mvmina@bk.ru
Поступила в редакцию 26.04.2021
После доработки 10.06.2021
Принята к публикации 10.06.2021
- EDN: KQDQIG
- DOI: 10.31857/S0042875222030134
Аннотация
Показано, что филетические отношения морфотипов крупных африканских усачей Barbus (Labeobarbus auctorum) можно оценивать по результатам сравнительного анализа структуры морфологических различий между ними. При этом структура различий между двумя выборками характеризуется распределением векторных нагрузок признаков на ту главную компоненту, по которой выявляется их наиболее чёткое разобщение, а сходство структуры различий между парами оценивается посредством сопоставления структуры различий между выборками в первой и во второй парах. Подтверждается гипотеза, согласно которой фенетическая иерархия морфотипов крупных африканских усачей из оз. Тана возникла в результате независимых ответвлений онтогенетических каналов от канала генерализованной формы. Сходство структуры различий между морфотипами характеризуется сходством векторных нагрузок в попарных сравнениях и сходством расположения признаков, ранжированных по величине их векторных нагрузок.
Крупные африканские усачи рода Barbus (Labeobarbus auctorum), обитающие в оз. Тана (Эфиопия), характеризуются чрезвычайным фенетическим разнообразием. Взрослые особи, сходные по внешним признакам, образуют дискретные или не вполне дискретные группировки, которые Банистер (Banister, 1973) называл экофенотипами, а Нагелькерке с соавторами вначале морфотипами (Nagelkerke et al., 1994), а позже – видами (Nagelkerke, Sibbing, 2000). Можно ли считать все или некоторые морфотипы видами, вопрос спорный, обсуждение которого не входит в задачу настоящего исследования.
Опубликовано немало работ, посвящённых анализу фенетических отношений морфотипов крупных африканских усачей, оцениваемых по величине различий между ними как по внешним признакам, так и по признакам черепа (Nagelkerke et al., 1994; Mina et al., 1996а; Nagelkerke., Sibbing 2000; Мина и др., 2011). Однако различия одинаковой величины могут иметь разное происхождение, и фенетические отношения морфотипов могут не отражать филетические.
Цель работы − на основе многомерного анализа морфологических признаков крупных африканских усачей оценить филетические отношения морфотипов с использованием ранее не применявшегося сравнительного анализа структуры различий между морфотипами. При этом структура различий между двумя морфотипами (выборками) характеризуется распределением векторных нагрузок признаков на ту главную компоненту, по которой выявляется их наиболее чёткое разобщение, а сходство структуры различий между морфотипами одной пары и морфотипами другой пары оценивается посредством сопоставления структуры различий этих пар.
МАТЕРИАЛ И МЕТОДИКА
Материал, взятый нами из коллекции Совместной российско-эфиопской биологической экспедиции РАН, собран в 1992–2010 гг. Операциональными единицами служили морфотипы, т.е. группы взрослых особей, выделенные по признакам внешней морфологии (Nagelkerke et al., 1994). Ранее было показано, что эти группы различаются и по признакам черепа (Mina et al., 1996а, 1996b; Мина и др., 2011).
Усачи, имевшие морфологические признаки пищевой специализации (de Graaf et al., 2008), рассматривались как представители специализированных морфотипов, а не имевшие таких признаков – как представители генерализованной формы11 (далее GF). В нашем материале GF из бассейна оз. Тана представлена выборками из собственно озера – 119 особей (Tn) и из впадающей в него р. Гельда – 38 особей (Ge), а специализированные морфотипы − выборками рыбоядных форм: acute (ac) – 70 особей, bigmouth big-eye (be) – 51 особь, bigmouth small-eye (se) – 49 особей, dark (da) – 32 особи, troutlike (tr) – 32 особи, white hunch (wh) – 33 особи, а также выборкой бентофага barbel (ba) – 33 особи (рис. 1). Кроме того, была использована выборка (za), включавшая пять большеротых (рыбоядных) особей из р. Дидесса, впадающей в Голубой Нил ниже оз. Тана. Эта выборка представляет морфотип, описанный как вид Barbus zaphiri Boulenger, 1906.
Рис. 1.
Представители генерализованной формы (GF) (а) и специализированных морфотипов (б–з) усачей комплекса Barbus intermedius sensu Banister (1973) из оз. Тана: б – bigmouth big-eye (be), в – bigmouth small-eye (se), г – troutlike (tr), д – acute (ac), е – barbel (ba), ж – white hunch (wh), з – dark (da); названия специализированных морфотипов – по: Nagelkerke et al., 1994.

Всего были проанализированы 14 промеров костей черепа (рис. 2). В расчётах использованы индексы − отношения абсолютных значений промеров к базальной длине черепа (BL). Далее в обсуждении, упоминая тот или иной признак, мы имеем в виду его индекс, а не сам промер. Значения индексов преобразовывали в натуральные логарифмы для нормализации распределений.
Рис. 2.
Схема промеров черепа усачей комплекса Barbus intermedius: BL − базальная длина черепа; B1, B2, B3 – расстояние между внешними краями соответственно frontalia, pterotica и sphenotica; B4 – ширина черепа на уровне соединения frontale и pteroticum; HS1, HS2 – высота черепа на уровне соответственно изгиба parasphenoideum и заднего края parasphenoideum; Hm – высота hyomandibulare, Pop − длина praeoperculum, Op – высота передней части operculum; Pmx, Mx, De, Iop – длина соответственно praemaxillare, maxillare, dentale и interoperculum.

Статистическая обработка данных выполнена средствами пакетов NTSYS 2.02k (Rohlf, 1998) и Статистика 6. В анализе главных компонент (АГК) собственные векторы считали по корреляционной матрице; длину вектора принимали равной 1.
Алгоритм расчётов
1. Проводится АГК попарных сочетаний выборок, представляющих морфотипы М1, М2, М3 ….. Мn (сравнение двух морфотипов обозначается как Мi−Мj), оцениваются векторные нагрузки признаков. Поскольку морфотипы в попарных сравнениях наиболее чётко расходятся по ГК1, структура различий между ними характеризуется распределением векторных нагрузок признаков на эту компоненту.
2. Сравниваются векторные нагрузки признаков на ГК1 в разных попарных сравнениях путём построения двумерного распределения, где по абсциссе откладываются оценки нагрузок для одной пары морфотипов, а по ординате – для другой. При этом сравниваются пары, либо включающие разные выборки (Мi−Мj и Мg−Мh), либо одна и та же выборка входит в состав обеих пар (Мi−Мj и Мi−Мg). Оценена структура различий между выборками в 30 парах и проведены 134 сравнения распределений векторных нагрузок признаков между разными парами, из них восемь сравнений серии Мi−Tn и Tn−Ge, 21 − серии Мi−Tn и Мj−Tn и 105 − серии Мj−Мi и Мg−Мi
3. Сходство структуры различий между морфотипами, представленными выборками в разных парах, характеризуется по форме скаттера и по величине коэффициента корреляции рангов Спирмена (rS). Сходство тем больше, чем чётче в двумерном распределении прослеживается линейная связь переменных. Ранее коэффициент корреляции рангов Спирмена применяли при анализе структуры различий выборок алтайских османов рода Oreoleuciscus из озёр Монголии (Дгебуадзе и др., 2008). Подход, основанный на оценке взаимосоответствия векторных нагрузок признаков, использовали Андреев и Решетников (1978) для выяснения направлений внутривидовой дифференциаций сига Coregonus lavaretus. Эти авторы визуально оценивали расположение скаттера (двумерного распределения).
В качестве показателя обобщённого фенетического расстояния сходства между выборками специализированных морфотипов и GF мы использовали обобщённую таксономическую дистанцию D (Rohlf, 1998). Кластерный анализ матрицы дистанций проводили невзвешенным парно-групповым методом (UPGMA).
РЕЗУЛЬТАТЫ
Анализ полученных данных показал, что сходство структуры различий между морфотипами (форма скаттера и rS) сильно варьирует. В одних случаях структура различий между специализированным морфотипом и GF из Таны близка к наблюдаемой между выборками GF из Таны и р. Гельды (Мi−Tn и Tn−Ge) (рис. 3а–3г), тренд двумерного распределения описывается прямой; лишь в случае морфотипа ac тренд искривлён (рис. 3д), причём во всех случаях имеет место сильная корреляция рангов (rS > 0.80). Несколько неожиданно самое высокое сходство структуры различий наблюдается при сравнении пары, включающей рыбоядную форму za из бассейна р. Дидесса и Tn c парой выборок GF (Tn−Ge) из бассейна оз. Тана (рис. 3е) (rS = 0.97, тренд линейный).
Рис. 3.
Структура различий по индексам черепа между специализированными морфотипами и морфотипом Tn в сравнении с таковой между генерализованными морфотипами усачей комплекса Barbus intermedius из оз. Тана и р. Гельда (Tn−Ge): а – da–Tn, rS = 0.92 (здесь и далее р < 0.001); б – se−Tn, rS = 0.92; в – be–Tn, rS = 0.85; г – wh–Tn, rS = 0.82; д – ac–Tn, rS = 0.86; е – za−Tn, rS = 0.97. Нагрузки параметров: (⚫) − челюстной дуги: Pmx, Mx, De; (▲) – жаберной крышки: Pop и Op, (△) – Iop; (⚪) – нейрокраниума: B1, B2, B3, B4, HS1, HS2; (◼) – Hm. (za) – рыбоядная форма из бассейна р. Дидесса; ост. обозначения см. на рис. 1, 2.

Сходна структура различий и между парами, включающими разные специализированные морфотипы и Tn: Мi−Tn и Mj−Tn (рис. 4а–4г). Исключение составляют сравнения с парой ac–Tn (рис. 4д, 4е). В этих случаях единого тренда нет, но сильна корреляция рангов (rS ≥ 0.80). Структура различий между парами МJ–Мi и Мg–Мi, в которых все морфотипы специализированные (причём один и тот же морфотип входит в состав обеих пар), крайне разнообразна, и в большинстве случаев сходства структуры различий не наблюдается (рис. 5), но в сравнении se–ba и se–da (рис. 5а: линейный тренд и rS = 0.94) картина та же, что и в сравнениях Мi–Tn и Mj–Tn (рис. 4). Однако если вместо ba и da взяты другие морфотипы, то подобное сходство структуры различий отсутствует (рис. 5б–5е).
Рис. 4.
Сравнение структуры различий по индексам черепа усачей комплекса Barbus intermedius из оз. Тана между парами, включающими разные специализированные морфотипы и генерализованную форму (Tn): а – tr–Tn и da–Tn, rS = 0.92 (здесь и далее р < 0.001); б – se–Tn и da−Tn, rS = 0.87; в – be–Tn и wh–Tn, rS = 0.81; г – da–Tn и be–Tn, rS = = 0.80; д – ac–Tn и wh–Tn, rS = 0.83; е – ac–Tn и ba–Tn, rS = 0.83. Обозначения см. на рис. 1–3.
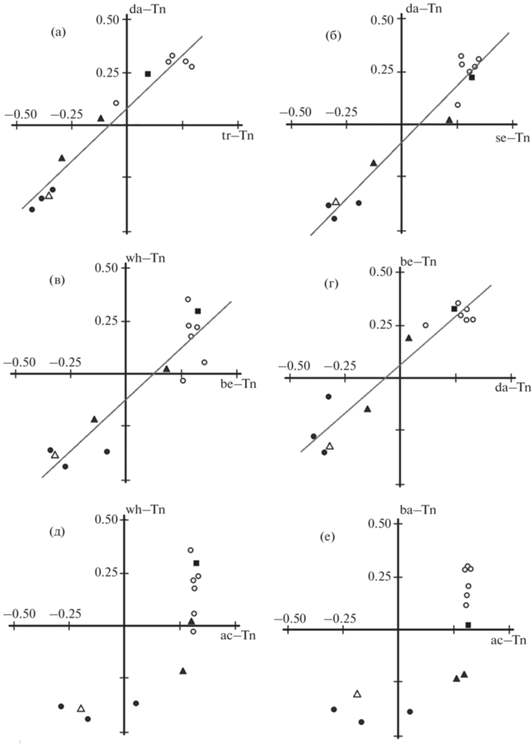
Рис. 5.
Сравнение структуры различий по индексам черепа усачей комплекса Barbus intermedius из оз. Тана между парами, в которых все морфотипы специализированные, причём один и тот же морфотип входит в состав обеих пар: а – se–da и se–ba, rS = 0.94, р < 0.001; б – be–tr и be–wh, rS = 0.77, р < 0.01; в – da–tr и da–wh, rS = 0.65, р < 0.05; г – se–be и se–wh, rS = 0.57, р < 0.05; д – tr–se и tr–wh, rS = –0.006, р > 0.05; е – wh–tr и wh–se, rS = –0.52, р > 0.05. Обозначения см. на рис. 1−3.

ОБСУЖДЕНИЕ
Нет оснований сомневаться, что специализированные морфотипы крупных усачей, существующие или существовавшие в реках и озёрах Африки, в том числе в оз. Тана, произошли от генерализованной формы, но пути и способы их происхождения могли быть различны.
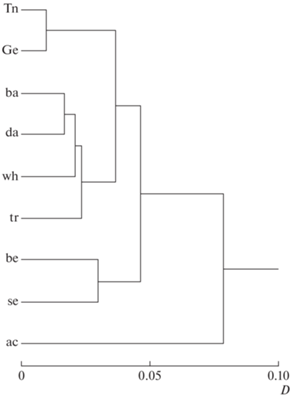
Фенетическое разнообразие танских усачей иерархически организовано (рис. 6). Если предположить, что приведённая дендрограмма отражает филетическую схему диверсификации, то надо сделать вывод о том, что вначале от общего генерализованного предка, сходного с представленными в нашем материале морфотипами Tn и Ge, отделился морфотип ac, потом – общий предок морфотипов be и se (или один из этих морфотипов, от которого затем произошёл другой), а затем от предковой GF дивергировал общий предок морфотипов wh, da, ba и tr.
Рис. 6.
Дендрограмма сходства (UPGMA) морфотипов усачей комплекса Barbus intermedius из оз. Тана и р. Гельда: Tn, Ge – генерализованная форма из озера и реки; названия остальных морфотипов см. на рис. 1.
Согласно альтернативной гипотезе, морфотипы обособлялись от генерализованных предков независимо друг от друга подобно тому, как это происходит в онтогенезе. Ранее при анализе морфологических изменений усачей оз. Тана было показано (Mina at al., 1996b), что в процессе онтогенеза специализированный морфотип формируется в терминальном участке онтогенетического канала, причём каналы разных специализированных морфотипов отходят от канала GF (морфотипа intermedius) при разных размерах рыб, и раньше прочих ответвляется канал морфотипа ac. Эту гипотезу подтверждают результаты настоящего исследования. Большое сходство структуры различий Tn−Ge с парами, включающими специализированный морфотип и морфотип Tn, позволяет предполагать, что дивергенция каждого специализированного морфотипа от GF была подобна разделению Tn и Ge, филетическая близость которых несомненна.
О независимости дивергенций специализированных морфотипов свидетельствует сходство структуры различий между каждым из них и морфотипом intermedius (Мj–Tn и Мg–Tn, где Мj и Мg – специализированные морфотипы) (рис. 4), при том что сходство структуры различий между парами специализированных морфотипов, когда один и тот же морфотип входит в состав обеих пар (Мi–Мj и Мi–Мg), не описывается единой схемой и сходство структуры различий в большинстве случаев отсутствует или невелико (рис. 5). Сходство структуры различий в сравнении se–ba и se–da мы склонны расценивать как случайное совпадение структуры различий в сравниваемых парах, т.е. как реализацию одного из многих возможных вариантов.
Заметим, что предковая GF могла быть не тождественной Tn или Ge. Об этом свидетельствует большое сходство структуры различий между za–Tn и Tn–Ge (рис. 3е). Поскольку za происходит из р. Дидесса, находящейся далеко от оз. Тана, хотя тоже в бассейне Голубого Нила, маловероятно, что генерализованный морфотип, от которого дивергировал za, мог быть идентичен одному из танских. Соответственно, мы не можем утверждать, что все танские морфотипы произошли от одного морфотипа GF, в частности, от Tn или Ge.
Следует отметить, что в период сбора материала особи всех исследованных морфотипов, кроме da, нерестились в одно и то же время, заходя в одни и те же реки, и между ними не было полной репродуктивной изоляции (Nagelkerke, Sibbing, 1996; Dgebuadze et al., 1999; Дзержинский и др., 2007). Если такая ситуация существовала и в период формирования морфотипов, то надо сделать вывод о том, что они возникли не от репродуктивно изолированных популяций GF. Однако весьма вероятно, что ситуация в прошлом отличалась от современной и, возможно, морфотипы возникли от разных существовавших в то время репродуктивно изолированных популяций и пришли в контакт, уже имея предрасположенность к скрещиванию с себе подобными.
Судя по нашим данным, в процессе дивергенции специализированного морфотипа от GF структура различий между ними изменяется. Структура различий между ac и Tn выглядит иначе, чем между другими специализированными морфотипами и Tn. Это можно объяснить более ранним обособлением ac от генерализованных предков, о чём свидетельствует и наибольшая фенетическая дистанция между ac и Tn (рис. 6). Примечательно, что в процессе дивергенции тесная ранговая корреляция структуры различий между специализированным и генерализованным морфотипами сохраняется, несмотря на изменения формы скаттера, т.е. порядок расположения признаков в ряду ранжированных по величине их векторных нагрузок изменяется мало.
Выяснить, в какой мере предлагаемый нами метод может быть полезен при изучении фенетического разнообразия рыб, − задача дальнейших исследований. Можно надеяться, что с его помощью удастся выявлять случаи, когда иерархия фенотипов возникла в результате независимых дивергенций от общего дерева.
Список литературы
Андреев В.Л., Решетников Ю.С. 1978. Анализ фенотипической изменчивости географически удалённых популяций одного вида // Математические методы в экологии и географии. Владивосток: Изд-во ДВНЦ АН СССР. С. 98−110.
Дгебуадзе Ю.Ю., Мина М.В., Мироновский А.Н. 2008. К оценке фенетических отношений алтайских османов (Oreoleuciscus, Cyprinidae) из трех озер Монголии по признакам черепа // Вопр. ихтиологии. Т. 48. № 3. С. 315−323.
Дзержинский К.Ф., Шкиль Ф.Н., Белай Абдисса и др. 2007. Нерест “крупных усачей” (Barbus intermedius complex) в реке Дукалит бассейна озера Тана (Эфиопия) и репродуктивные отношения между некоторыми формами, описанными как виды // Там же. Т. 47. № 5. С. 676–683.
Мина М.В., Мироновский А.Н., Капитанова Д.В. 2011. Фенетические отношения и вероятные пути морфологической диверсификации африканских усачей комплекса Barbus intermedius из озера Тана (Эфиопия) // Там же. Т. 51. № 2. С. 149−163.
Banister K.E. 1973. A revision of the large Barbus (Pisces, Cyprinidae) of East and Central Africa. Pt. II // Bull. Brit. Mus. Nat. Hist. Zool. V. 26. P. 1–148.
de Graaf M., Dejen E., Osse J.W.M., Sibbing F.A. 2008. Adaptive radiation of Lake Tana’s (Ethiopia) Labeobarbus species flock (Pisces, Cyprinidae) // Mar. Freshwat. Res. V. 59. № 5. P. 391–407. https://doi.org/10.1071/MF07123
Dgebuadze Yu.Yu., Mina M.V., Alekseev S.S., Golubtsov A.S. 1999. Observations on reproduction of the Lake Tana barbs // J. Fish. Biol. V. 54. P. 417–423.
Mina M.V., Mironovsky A.N., Dgebuadze Yu.Yu. 1996a. Lake Tana large barbs: phenetics, growth and diversification // Ibid. V. 48. № 3. P. 383−404. https://doi.org/10.1111/j.1095-8649.1996.tb01435.x
Mina M.V., Mironovsky A.N., Dgebuadze Yu.Yu. 1996b. Morphometry of barbel of lake Tana, Ethiopia: multivariate ontogenetic channels // Folia Zool. V. 45. Suppl. 1. P. 109–116.
Nagelkerke L.A.J., Sibbing F.A. 1996. Reproductive segregation among the Barbus intermedius complex of Lake Tana, Ethiopia. An example of intralacustrine speciation // J. Fish. Biol. V. 49. № 6. P. 1244–1266. https://doi.org/10.1111/j.1095-8649.1996.tb01793.x
Nagelkerke L.A.J., Sibbing F.A. 2000. The large barbs (Barbus spp., Cyprinidae, Teleostei) of Lake Tana (Ethiopia), with a description of a new species, Barbus osseensis // Netherl. J. Zool. V. 50. № 2. P. 179–214. https://doi.org/10.1163/156854200505946
Nagelkerke L.A.J., Sibbing F.A., van den Boogaart J.G.M. et al. 1994. The barbs (Barbus spp.) of Lake Tana: a forgotten species flock? // Environ. Biol. Fish. V. 39. № 1. P. 1–21. https://doi.org/10.1007/BF00004751
Rohlf F.J. 1998. NTSYS-pc: numerical taxonomy and multivariate analysis system. Version 2.02. N.Y.: Exeter Software Press, 31 p.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии