

Вопросы ихтиологии, 2022, T. 62, № 5, стр. 645-652
Звукогенерация у мраморного гурами Trichopodus trichopterus (Osphronemidae) во время питания
А. О. Касумян 1, *, Е. С. Михайлова 1
1 Московский государственный университет
Москва, Россия
* E-mail: alex_kasumyan@mail.ru
Поступила в редакцию 24.11.2021
После доработки 21.12.2021
Принята к публикации 22.12.2021
- EDN: KUCVSO
- DOI: 10.31857/S0042875222050101
Аннотация
Изучена генерация звуков у половозрелых мраморных гурами Trichopodus trichopterus при реагировании на агар-агаровые гранулы, содержащие классические вкусовые вещества и аминокислоты. Звуки (щелчки) возникают нерегулярно (лишь в 14.6% всех опытов), но зарегистрированы они со всеми типами гранул. Среднее число звуков варьирует от 1.5–4.0 для гранул с пролином, норвалином, треонином и хлоридом натрия до 8.0–9.5 для гранул с глутамином, аргинином, лимонной кислотой. Звуки следуют непрерывно или сериями с короткими интервалами, максимальное число звуков в опыте – 31. В ответ на одни и те же типы гранул звуки у мраморного гурами возникают реже, а максимальное число издаваемых звуков (щелчков) почти в два раза меньше, чем у исследованного ранее жемчужного гурами. Возникновение и число звуков не связано с вкусовой привлекательностью гранул, но тесно коррелирует с касаниями – быстрыми прикосновениями или короткими удержаниями гранулы губами, наблюдавшимися в 35.9% опытов. Среднее число касаний в опытах с разными тестируемыми веществами – 3.0–8.6, максимальное – >20. Спорадичность и тесная корреляция возникновения звуков с проявлением рыбами двигательных действий при реагировании на гранулы предполагает отсутствие у генерируемых звуков сигнальной функции.
Способность рыб производить звуки хорошо известна. У многих рыб звуки выполняют важные коммуникативные функции и входят в число обязательных элементов ритуального поведения, проявляемого при размножении, охране территории и внутригрупповых контактах, в том числе при конкуренции за пищу (Amorim, Hawkins, 2000; Amorim et al., 2004; Kasumyan, 2009). При питании рыбы производят и неспециализированные звуки, возникающие при схватывании и внутриротовой обработке кормовых объектов (Colson et al., 1998; Lagardère, Mallekh, 2000). Такие непреднамеренные звуки питающихся рыб также могут иметь сигнальное значение и привлекать партнёров по группе или стае в места, благоприятные для откорма (Касумян, Павлов, 2018).
К числу рыб, звукогенерация у которых давно известна и хорошо изучена, относятся различные гурами (Osphronemidae). Издаваемые ими звуки (croaking sounds) слышны невооружённым ухом и напоминают короткие щелчки. Возникают щелчки при быстрых движениях грудных плавников и генерируются благодаря особенностям морфологии скелета и мышц плавников. Звуки издаются в основном при конфликтах и сигнализируют, как полагают, о потенциальных возможностях соперников (Daugherty, Marshall, 1976; Ladich et al., 1992; Henglmüller, Ladich, 1999). Звуковая сигнализация выявлена у гурами и при нересте (Ladich, 2007). Звуки способны издавать особи обоих полов, но у самцов они более сильные и продолжительные и отличаются по частотным параметрам (Liesch, Ladich, 2020).
Недавно характерные щелчки были зарегистрированы у жемчужного Trichopodus leerii и мраморного T. trichopterus гурами при реагировании на искусственные пищевые объекты – агар-агаровые гранулы, использовавшиеся для оценки вкусовых предпочтений этих рыб (Виноградская и др., 2017; Михайлова, Касумян, 2021). Было обнаружено, что у жемчужного гурами генерация звуков коррелирует с числом схватываний гранул, но не связана со вкусовой привлекательностью содержащихся в них веществ (Виноградская и др., 2017). Последнее может быть обусловлено ограниченным числом использованных веществ. Цель настоящей статьи – оценить связь между генерацией звуков мраморным гурами и вкусовой привлекательностью гранул с использованием широкого спектра химических веществ, а также пищевым поведением, проявляемым рыбами при реагировании на гранулы разного вкусового качества.
МАТЕРИАЛ И МЕТОДИКА
Для анализа использованы новые сведения о поведении и звукогенерации, полученные ранее в ходе исследования вкусовых предпочтений мраморного гурами и не вошедшие в предыдущую публикацию (Михайлова, Касумян, 2021). Эксперименты проведены на 16 половозрелых особях TL 75–95 мм, рассаженных поодиночке в изолированные аквариумы (объём 10 л, температура воды 26°С). Рыб кормили личинками Chironomidae ежедневно до насыщения. В каждом опыте в аквариум вносили гранулу из 2%-ного агар-агарового геля (“Reanal”, Венгрия), содержащую одно из вкусовых веществ (таблица). Использовали также гранулы с водным экстрактом личинок Chironomidae (175 г/л) и контрольные гранулы, содержащие только краситель Ponceau 4R (5 мкM), присутствовавший также во всех других гранулах. Цилиндрические гранулы имели длину 4 мм и диаметр 2 мм. Детальное изложение методики приведено ранее (Михайлова, Касумян, 2021). В ходе каждого опыта регистрировали число схватываний гранулы, продолжительность её удержания во рту рыбой при первом схватывании и в течение всего опыта, потребление или отвергание гранулы к концу опыта. Дополнительно визуально подсчитывали число совершаемых рыбой касаний гранулы – быстрых прикосновений или коротких удержаний гранулы губами, обычно предшествующих первому схватыванию. При удержании губами гранула остаётся снаружи в отличие от схватываний, когда вся гранула оказывается внутри ротовой полости рыбы. Одновременно подсчитывали число производимых рыбой звуков (щелчков). Если рыба хотя бы один раз не схватывала поданную гранулу, то такие случаи опытом не считали. Общее число опытов – 2592. Статистический анализ выполнен с применением U-критерия Манна–Уитни и рангового коэффициента корреляции Спирмена (rs).
Вкусовые ответы (М ± m) мраморного гурами Trichopodus trichopterus на гранулы с тестируемыми веществами
| Раздражитель | Концентрация, М (%) |
Опыты с касаниями | Опыты с генерацией звуков | ||||
|---|---|---|---|---|---|---|---|
| все опыты | опыты, завершившиеся потреблением или отверганием гранулы | ||||||
| доля опытов, % | число касаний | доля опытов, % | число звуков | число звуков | число опытов | ||
| Классические вкусовые вещества | |||||||
| Лимонная кислота | 0.260 (5) | 63.5 ± 4.9 | 7.3 ± 0.7 | 28.1 ± 4.6 | 9.0 ± 1.2 | $\frac{{11.9 \pm 2.5}}{{7.2 \pm 1.0}}$ | $\frac{{10}}{{17}}$ |
| CaCl2 | 0.900 (10) | 50.0 ± 5.1 | 6.3 ± 0.5 | 32.2 ± 4.8 | 7.1 ± 0.8 | $\frac{{7.7 \pm 1.7}}{{6.8 \pm 0.8}}$ | $\frac{{10}}{{20}}$ |
| NaCl | 1.730 (10) | 36.5 ± 5.0 | 4.0 ± 0.4 | 25.0 ± 4.4 | 4.5 ± 0.6 | $\frac{{6.5 \pm 1.5}}{{4.3 \pm 0.6}}$ | $\frac{2}{{22}}$ |
| Сахароза | 0.290 (10) | 46.9 ± 5.1 | 5.2 ± 0.5 | 32.3 ± 4.8 | 5.5 ± 0.7 | $\frac{{15.0}}{{5.2 \pm 0.6}}$ | $\frac{1}{{30}}$ |
| Свободные L-аминокислоты | |||||||
| Пролин | 0.1 | 9.4 ± 3.0 | 3.0 ± 0.4 | 2.1 ± 1.5 | 1.5 ± 0.5 | $\frac{{1.5 \pm 0.5}}{ - }$ | $\frac{2}{0}$ |
| Серин | 0.1 | 33.3 ± 4.8 | 5.3 ± 0.7 | 14.6 ± 3.6 | 5.1 ± 0.8 | $\frac{{5.5 \pm 1.0}}{{4.0 \pm 1.0}}$ | $\frac{{11}}{3}$ |
| Треонин | 0.1 | 27.1 ± 4.6 | 7.0 ± 1.6 | 13.5 ± 3.5 | 6.1 ± 1.0 | $\frac{{6.0 \pm 1.2}}{{6.5 \pm 0.5}}$ | $\frac{{10}}{2}$ |
| Цистеин | 0.1 | 19.8 ± 4.1 | 3.9 ± 0.4 | 7.3 ± 2.7 | 5.3 ± 0.7 | $\frac{{5.2 \pm 0.8}}{{6.0}}$ | $\frac{6}{1}$ |
| Глицин | 0.1 | 36.5 ± 4.9 | 5.0 ± 0.6 | 16.7 ± 3.8 | 5.4 ± 0.8 | $\frac{{5.3 \pm 0.9}}{{5.7 \pm 1.9}}$ | $\frac{{13}}{3}$ |
| Аланин | 0.1 | 33.3 ± 4.8 | 6.2 ± 0.7 | 13.5 ± 3.5 | 6.6 ± 1.1 | $\frac{{7.8 \pm 1.3}}{{4.0 \pm 0}}$ | $\frac{{10}}{3}$ |
| Норвалин | 0.1 | 32.3 ± 4.8 | 4.0 ± 0.4 | 5.2 ± 2.3 | 4.8 ± 0.7 | $\frac{{4.5 \pm 0.9}}{{6.0}}$ | $\frac{4}{1}$ |
| Фенилаланин | 0.1 | 53.1 ± 5.1 | 5.6 ± 0.5 | 21.9 ± 4.2 | 5.7 ± 0.8 | $\frac{{6.1 \pm 1.0}}{{4.7 \pm 0.9}}$ | $\frac{{15}}{6}$ |
| Валин | 0.1 | 31.3 ± 4.8 | 5.1 ± 0.6 | 12.5 ± 3.4 | 5.3 ± 0.9 | $\frac{{5.0 \pm 1.1}}{{5.6 \pm 1.7}}$ | $\frac{7}{5}$ |
| Метионин | 0.1 | 44.8 ± 5.1 | 4.7 ± 0.4 | 17.7 ± 3.9 | 5.1 ± 0.8 | $\frac{{5.5 \pm 1.2}}{{4.3 \pm 0.5}}$ | $\frac{{11}}{6}$ |
| Гистидин | 0.1 | 53.1 ± 5.1 | 6.2 ± 0.6 | 18.8 ± 4.0 | 6.9 ± 1.1 | $\frac{{8.8 \pm 1.6}}{{4.6 \pm 0.9}}$ | $\frac{{10}}{8}$ |
| Глутамин | 0.1 | 46.9 ± 5.1 | 7.0 ± 0.7 | 14.6 ± 3.6 | 7.7 ± 1.4 | $\frac{{7.7 \pm 1.8}}{{7.8 \pm 2.4}}$ | $\frac{9}{5}$ |
| Аспарагин | 0.1 | 37.5 ± 5.0 | 5.6 ± 0.6 | 8.3 ± 2.8 | 7.5 ± 1.0 | $\frac{{7.3 \pm 1.7}}{{7.6 \pm 1.3}}$ | $\frac{3}{5}$ |
| Лизин | 0.1 | 26.0 ± 4.5 | 5.7 ± 0.7 | 6.3 ± 2.9 | 8.4 ± 2.1 | $\frac{{7.5 \pm 5.0}}{{9.0 \pm 2.0}}$ | $\frac{2}{3}$ |
| Аргинин | 0.1 | 43.8 ± 5.1 | 7.3 ± 0.8 | 22.9 ± 4.3 | 10.5 ± 1.1 | $\frac{{6.0 \pm 1.5}}{{11.2 \pm 1.2}}$ | $\frac{3}{{19}}$ |
| Аспарагиновая кислота | 0.01 | 20.8 ± 4.2 | 4.8 ± 0.5 | 9.4 ± 0.3 | 5.9 ± 0.7 | $\frac{{6.6 \pm 0.8}}{{5.0 \pm 1.1}}$ | $\frac{5}{4}$ |
| Изолейцин | 0.01 | 27.1 ± 4.6 | 3.8 ± 0.6 | 7.3 ± 2.7 | 5.1 ± 1.7 | $\frac{{5.5 \pm 3.5}}{{5.0 \pm 2.2}}$ | $\frac{2}{5}$ |
| Глутаминовая кислота | 0.01 | 16.7 ± 3.8 | 4.3 ± 0.6 | 3.1 ± 1.8 | 5.0 ± 2.1 | $\frac{{--~}}{{5.0 \pm 2.1}}$ | $\frac{0}{3}$ |
| Лейцин | 0.01 | 35.4 ± 4.9 | 5.7 ± 0.9 | 9.4 ± 3.0 | 9.3 ± 2.8 | $\frac{{29.0}}{{6.9 \pm 1.4}}$ | $\frac{1}{8}$ |
| Триптофан | 0.01 | 39.6 ± 5.0 | 6.0 ± 0.6 | 10.4 ± 3.1 | 6.0 ± 1.0 | $\frac{{4.0 \pm 0.0}}{{6.5 \pm 1.1}}$ | $\frac{2}{8}$ |
| Тирозин | 0.001 | 60.4 ± 5.0 | 6.5 ± 0.9 | 16.7 ± 3.8 | 6.4 ± 1.0 | $\frac{{4.0~}}{{6.6 \pm 1.1}}$ | $\frac{1}{{15}}$ |
| Экстракт Chironomidae | 175.0 | 24.0 ± 4.4 | 6.4 ± 1.3 | 12.5 ± 3.4 | 8.7 ± 2.2 | $\frac{{8.7 \pm 2.2}}{ - }$ | $\frac{{12}}{0}$ |
| Контроль | 13.5 ± 3.5 | 8.6 ± 1.0 | 11.5 ± 3.3 | 8.8 ± 1.2 | $\frac{{10.5 \pm 2.5}}{{8.4 \pm 1.4}}$ | $\frac{2}{9}$ | |
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Характерные щелчки, издаваемые мраморным гурами, хорошо слышны даже на удалении в несколько метров от аквариума с рыбой. Они возникают при питании рыб личинками Chironomidae и в опытах при реагировании на гранулы. В ответ на внесение гранулы звуки возникают нерегулярно – лишь в 14.6% всех опытов (рис. 1), но зарегистрированы они со всеми типами гранул. Доля опытов с генерацией звуков варьирует от 2–5% для гранул с глутаминовой кислотой, пролином, лизином, норвалином; до >20–30% для гранул с хлоридами натрия и кальция, сахарозой, лимонной кислотой, аргинином, фенилаланином (таблица). Возникновение звуков не связано со вкусовой привлекательностью гранул (рис. 2), что полностью подтверждает вывод, сделанный ранее для жемчужного гурами на примере сравнительно небольшого набора вкусовых веществ (Виноградская и др., 2017).
Рис. 1.
Доля опытов (%) с генерацией звуков без касаний ( ) и с касаниями (
) и с касаниями ( ) гранулы; и без генерации звуков без касаний (◻) и с касаниями (
) гранулы; и без генерации звуков без касаний (◻) и с касаниями ( ), выполненных на мраморном гурами Trichopodus trichopterus.
), выполненных на мраморном гурами Trichopodus trichopterus.
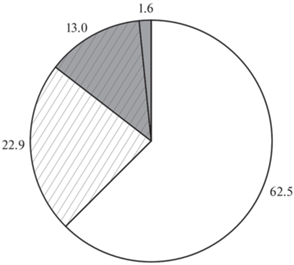
Рис. 2.
Коэффициент корреляции Спирмена между параметрами вкусового ответа на гранулы с тестируемыми веществами у мраморного гурами Trichopodus trichopterus: звуки – доля опытов с генерацией звуков, касания – доля опытов с касаниями гранулы, потребление – доля потреблённых гранул; отличия достоверны при * p < 0.05, *** p < 0.001.
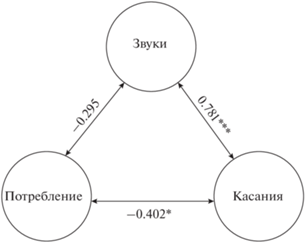
Касания. Число опытов с генерацией звуков тесно коррелирует с числом опытов, в которых наблюдались касания, совершаемые рыбами обычно после приближения и короткого броска к грануле. Касания могут происходить также за счёт приносящего потока воды, создаваемого рыбой при расширении ротовой полости. Касания наблюдаются в опытах со всеми типами гранул, но в среднем их тем меньше, чем выше процент потребления гранул (рис. 2).
Доля опытов с касаниями составляет 35.9% всех выполненных исследований, из них в 13% зарегистрированы звуки, а в 22.9% звуки отсутствовали (рис. 1). Рыбы в отдельных случаях совершали >20 касаний в опыте, но среднее их число варьирует от 3.0 до 8.6 для гранул с разными веществами (таблица) и не связано с потреблением (рис. 3). Таким образом, несмотря на отсутствие связи между вкусовой привлекательностью гранул и возникновением звуков, вкусовые свойства гранул могут влиять на звукогенерацию опосредованно, снижая вероятность касаний гранулы. Это влияние, по-видимому, слабое, поскольку в опытах с генерацией звуков, завершившихся потреблением или отверганием гранулы, число звуков не различается (таблица).
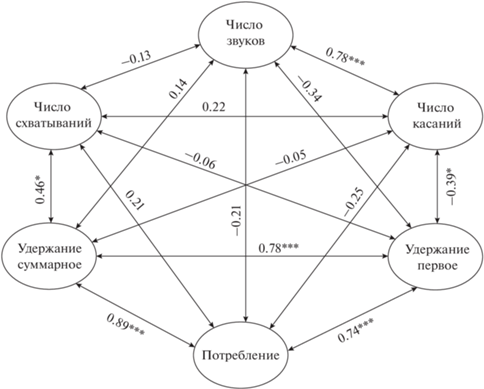
Рис. 3.
Коэффициент корреляции Спирмена между параметрами вкусового ответа на гранулы с тестируемыми веществами у мраморного гурами Trichopodus trichopterus: число звуков – среднее число звуков, число касаний – среднее число касаний гранулы, число схватываний – среднее число схватываний гранулы, удержание первое – средняя продолжительность удержания гранулы после первого схватывания, удержание суммарное – средняя продолжительность удержания гранулы в течение всего опыта; ост. обозначения см. на рис. 2.
Известно, что на губах у гурами имеются вкусовые почки (Scharrer et al., 1947). Прикосновения или кратковременное удержание гранулы позволяют гурами получать предварительную информацию о её вкусовых качествах. Такие действия ещё более характерны для рыб-бентофагов и рыб, питающихся в сумеречное и ночное время (Atema, 1980; Døving, Selset, 1980; Kasumyan, 1999; Касумян, Сидоров, 2010; Касумян и др., 2016). Наружные вкусовые почки у них могут быть не только на губах, но и на усах, голове, плавниках и других участках тела (Kapoor et al., 1975; Devitsina, 2005). Предварительная информация о сенсорных качествах пищи повышает эффективность питания рыб, у которых зрительная оценка объектов питания затруднена или невозможна. Гурами питаются преимущественно планктоном, схватывая своих жертв поштучно (Rainboth, 1996; Graham, 1997; Talde et al., 2004; Webb et al., 2007; Froese, Pauly, 2021). Несмотря на особенности предпочитаемых биотопов (водные заросли), выбор жертвы и ориентация на неё у гурами должны происходить под зрительным контролем. Возможно, поэтому касания или удержания гранул губами наблюдаются у них менее чем в половине опытов (35.9%). Получение предварительной информации о вкусовых качествах пищи для гурами имеет, по-видимому, второстепенное (факультативное) значение.
Звуки. Среднее число звуков (щелчков) в опытах, в которых они регистрируются, варьирует для разных типов гранул от 1.5–4.0 (пролин, норвалин, треонин, хлорид натрия) до 8.0–9.5 (глутамин, аргинин, лимонная кислота, контрольные гранулы). Звуки следуют непрерывно или сериями с короткими интервалами, максимальное число звуков в опыте – 31. Среднее число звуков зависит от числа совершаемых рыбой касаний столь же сильно, как и доля опытов с генерацией звуков от потребления гранул, но не зависит от вкусовой привлекательности гранул, числа схватываний гранул и продолжительности удержания гранулы после первого схватывания и суммарно в течение всего опыта (рис. 3). Если для корреляционного анализа учитывать не все опыты, а только те, в которых происходили касания и/или генерировались звуки, корреляция между средним числом звуков и средним числом касаний гранул с разными веществами ещё более тесная – rs = 0.858 (p < 0.001).
Связь возникновения звуков и их числа с числом касаний, но не с числом схватываний гранулы довольно неожиданная. Схватывания, в отличие от касаний, требуют более сильных машущих движений грудными плавниками – действий, приводящих к генерации щелчков у гурами. Схватывания (хотя бы одно) происходили во всех опытах, но звуки отмечены лишь в 14.6% случаев, тогда как из всех опытов с касаниями звуки возникали в 36% случаев. Касания не отмечены лишь в 1.6% всех опытов, или в 11% от всех опытов с генерацией звуков (рис. 1). В плечевом поясе гурами две из идущих к лучам грудного плавника мышц гипертрофированно развиты. В начале движения плавника вперёд крупные выступы в основании лучей препятствуют смещению сухожилий этих мышц. Но при продолжении движения натянутые сухожилия срываются с выступов, что и приводит к возникновению характерного щелчка, состоящего из одного или двух последовательных звуковых импульсов (Daugherty, Marshall, 1976; Henglmüller, Ladich, 1999). Возможно, что для совершения таких двигательных актов, как касания и схватывания, рыбы располагают грудные плавники по-разному или движение плавников имеет разный размах и разную траекторию. Нельзя исключать и того, что схватывания гурами осуществляют не только или не столько за счёт грудных плавников, но и благодаря хвостовому плавнику. Участие этих движителей в схватывании может варьировать от опыта к опыту в зависимости от расстояния от рыбы до гранулы и их взаимного положения. Все эти особенности хоть и усложняют понимание причин, но все же не влияют на главный вывод о связи между возникновением звуков и касаниями гранулы, характерными для поведения мраморного гурами.
В отличие от мраморного у жемчужного гурами генерация звуков коррелирует с числом схватываний гранул (Виноградская и др., 2017). Это может быть объяснено тем, что в опытах с жемчужным гурами схватывания и касания подсчитывали не раздельно, как в данной работе, а совместно. Касания у мраморного гурами хорошо заметны, возможно, из-за более крупных размеров опытных рыб, и раздельный подсчёт числа касаний и числа схватываний гранулы в опыте не вызывал каких-либо затруднений.
Гурами относятся к социальным рыбам. В период размножения им присуща территориальность и агрессивность, вне этого периода – внутригрупповая иерархия, обостряющаяся при появлении новых особей или при конкуренции за пищу (Forselius, 1957; Hollis, 1999; Webb et al., 2007). Звуковую сигнализацию гурами используют при формировании доминантно-соподчинённых отношений в группе и при нересте (Ladich et al., 1992; Ladich, 2007). Звуки, возникающие при реагировании гурами на гранулы, вряд ли можно считать сигнальными. Аргументов в пользу такого предположения может быть несколько. Во-первых, звукогенерация при скармливании гурами гранул или натуральной пищи (личинки Chironomidae) происходит редко – примерно в одном случае из семи (для гранул). Во-вторых, тесная корреляция между возникновением звуков и определённым типом двигательных действий (касаниями гранулы) также указывает на то, что звуки, скорее всего, сопутствуют пищевому поведению гурами и поэтому не могут быть отнесены к специализированным, т.е. имеющим сигнальное предназначение. Ещё одним доводом в пользу этого предположения может быть высокая степень связи между потреблением гранул во всех опытах и в опытах с генерацией звуков – 0.94 (p < 0.001) (рис. 4). Такая тесная связь свидетельствует, что звуки не могут быть сигналами о намерениях рыб потребить пищу или отказаться от неё. Однако следует учитывать, что в наших опытах рыбы находились в изолированных аквариумах и взаимодействия между опытными рыбами были невозможны. Вероятно, при наличии условий для зрительной или иной коммуникации звукогенерация происходила бы чаще. Окончательный ответ дадут эксперименты на рыбах, имеющих возможность сенсорных контактов друг с другом.
Рис. 4.
Потребление мраморным гурами Trichopodus trichopterus гранул с классическими вкусовыми веществами (1–4), L-аминокислотами (5–25) и водным экстрактом личинок Chironomidae (26) во всех опытах (◼) и в опытах, в которых зарегистрированы звуки ( ): 1 – лимонная кислота (0.26 М, 5%), 2 – хлористый кальций (0.9 М, 10%), 3 – хлористый натрий (1.73 М, 10%), 4 – сахароза (0.29 М, 10%); 5–19 – 0.1 М: 5 – пролин, 6 – серин, 7 – треонин, 8 – цистеин, 9 – глицин, 10 – аланин, 11 – норвалин, 12 – фенилаланин, 13 – валин, 14 – метионин, 15 – гистидин, 16 – глутамин, 17 – аспарагин, 18 – лизин, 19 – аргинин; 20–24 – 0.01 М: 20 – аспарагиновая кислота, 21 – глутаминовая кислота, 22 – изолейцин, 23 – лейцин, 24 – триптофан; 25 – тирозин (0.001 М), 26 – экстракт личинок Chironomidae (175.0 г/л), 27 – контроль. Даны средние значения, (
): 1 – лимонная кислота (0.26 М, 5%), 2 – хлористый кальций (0.9 М, 10%), 3 – хлористый натрий (1.73 М, 10%), 4 – сахароза (0.29 М, 10%); 5–19 – 0.1 М: 5 – пролин, 6 – серин, 7 – треонин, 8 – цистеин, 9 – глицин, 10 – аланин, 11 – норвалин, 12 – фенилаланин, 13 – валин, 14 – метионин, 15 – гистидин, 16 – глутамин, 17 – аспарагин, 18 – лизин, 19 – аргинин; 20–24 – 0.01 М: 20 – аспарагиновая кислота, 21 – глутаминовая кислота, 22 – изолейцин, 23 – лейцин, 24 – триптофан; 25 – тирозин (0.001 М), 26 – экстракт личинок Chironomidae (175.0 г/л), 27 – контроль. Даны средние значения, ( ) – ошибка средней.
) – ошибка средней.

Таким образом, наше исследование подтвердило, что звукогенерация, наблюдаемая у гурами при питании, не зависит от вкусовых качеств пищевых объектов. Она характерна для обоих исследованных видов – для мраморного и исследованного ранее жемчужного гурами (Виноградская и др., 2017). Спорадичность возникновения звуков и их корреляция с проявлением рыбами одного из элементов пищевого поведения предполагает отсутствие у этих звуков сигнальной функции. Несмотря на то что оба вида гурами близки по образу жизни, предпочитаемым биотопам, поведению и питанию (Forselius, 1957; Roberts, 1989; Graham, 1997; Talde et al., 2004; Zahid, 2008; Pranata et al., 2016), а эксперименты выполнены в идентичных условиях, звукогенерация у мраморного и жемчужного гурами различается. В ответ на одни и те же типы гранул звуки у жемчужного гурами возникают чаще, а максимальное число издаваемых звуков (щелчков) почти в два раза больше, чем у мраморного гурами. Известно, что у рыб с ростом изменяются частотно-амплитудные характеристики звуков (Kasumyan, 2008), поэтому интенсивность звукогенерации мраморного и жемчужного гурами скорее всего не обусловлена разными размерами опытных рыб, а отражает видовые различия между ними.
Список литературы
Виноградская М.И., Михайлова Е.С., Касумян А.О. 2017. Вкусовые предпочтения, оросенсорное тестирование и генерация звуков при питании у жемчужного гурами Trichopodus leerii (Osphronemidae) // Вопр. ихтиологии. Т. 57. № 3. С. 324–337. https://doi.org/10.7868/S004287521703016X
Михайлова Е.С., Касумян А.О. 2021. Вкусовые предпочтения и оросенсорное тестирование пищи у мраморного гурами Trichopodus trichopterus (Osphronemidae) // Там же. Т. 61. № 6. С. 697–712. https://doi.org/10.31857/S0042875221060126
Касумян А.О., Павлов Д.С. 2018. Стайное поведение рыб. М.: Т-во науч. изд. КМК, 273 с.
Касумян А.О., Сидоров С.С. 2010. Вкусовые предпочтения и поведение тестирования вкусовых качеств корма у усатого гольца Barbatula barbatula (Balitoridae, Cypriniformes) // Вопр. ихтиологии. Т. 50. № 5. С. 708–720.
Касумян А.О., Марусов Е.А., Сидоров С.С. 2016. Хеморецепция и пищевое поведение у обыкновенного ерша // Обыкновенный ёрш Gymnocеphalus cernuus (Linnaeus, 1758). Систематика, морфология, образ жизни и роль ерша в экосистемах. М.: Полиграф-Плюс. С. 77–91.
Amorim M.C.P., Hawkins A.D. 2000. Growling for food: acoustic emission during competitive feeding of the streaked gurnard // J. Fish Biol. V. 57. № 4. P. 895–907. https://doi.org/10.1111/j.1095-8649.2000.tb02200.x
Amorim M.C.P., Stratoudakis Y., Hawkins A.D. 2004. Sound production during competitive feeding in the grey gurnard // Ibid. V. 65. № 1. P. 182–194. https://doi.org/10.1111/j.0022-1112.2004.00443.x
Atema J. 1980. Chemical senses, chemical signals and feeding behavior in fish // Fish behavior and its use in the capture and culture of fishes. ICLARM Conference Proceedings. V. 5. Manila: ICLARM. P. 57–101.
Colson D.J., Patek S.N., Brainerd L., Lewis S.M. 1998. Sound production during feeding in Hyppocampus seahorses (Sygnathidae) // Environ. Biol. Fishes. V. 51. № 2. P. 221–229. https://doi.org/10.1023/A:1007434714122
Daugherty J., Marshall J.A. 1976. The sound producing mechanism of the croaking gurami, Trichopsis vittatus (Pisces, Belontiidae) // Physiol. Zool. V. 49. № 2. P. 227–244. https://doi.org/10.1086/physzool.49.2.30152543
Devitsina G.V. 2005. Comparative morphology of intraoral taste apparatus in fish // J. Ichthyol. V. 45. Suppl. 2. P. 286–306.
Døving K.B., Selset R. 1980. Behavior patterns in cod released by electrical stimulation of olfactory tract bundlets // Science. V. 207. № 4430. P. 559–560. https://doi.org/10.1126/science.7352272
Forselius S. 1957. Studies of anabantid fishes // Zool. Bidrag från Uppsala. V. 32. P. 93–597.
Froese R., Pauly D. (eds.). 2021. FishBase. World Wide Web electronic publication. www.fishbase.org. Version 08/2021
Graham J.B. 1997. Air-breathing fishes. Evolution, diversity and adaptation. San Diego: Acad. Press, 299 p. https://doi.org/10.1016/B978-0-12-294860-2.X5000-4
Henglmüller S.M., Ladich F. 1999. Development of agonistic behaviour and vocalization in croaking guramis // J. Fish Biol. V. 54. № 2. P. 380–395. https://doi.org/10.1111/j.1095-8649.1999.tb00837.x
Hollis K.L. 1999. The role of learning in the aggressive and reproductive behaviour of blue gouramis, Trichogaster trichopterus // Environ. Biol. Fish. V. 54. № 4. P. 355–369. https://doi.org/10.1023/A:1007529628117
Kapoor B.G., Evans H.E., Pevzner R.A. 1975. The gustatory system in fish // Adv. Mar. Biol. V. 13. P. 53–108. https://doi.org/10.1016/S0065-2881(08)60280-1
Kasumyan A.O. 1999. Olfaction and taste senses in sturgeon behaviour // J. Appl. Ichthyol. V. 15. № 4–5. P. 228–232. https://doi.org/10.1111/j.1439-0426.1999.tb00240.x
Kasumyan A.O. 2008. Sounds and sound production in fishes // J. Ichthyol. V. 48. № 11. P. 981–1030. https://doi.org/10.1134/S0032945208110039
Kasumyan A.O. 2009. Acoustic signaling in fish // Ibid. V. 49. № 11. P. 963–1020. https://doi.org/10.1134/S0032945209110010
Ladich F. 2007. Females whisper briefly during sex: context- and sex-specific differences in sounds made by croaking gouramis // Anim. Behav. V. 73. № 2. P. 379–387. https://doi.org/10.1016/j.anbehav.2006.04.014
Ladich F., Brittinger W., Kratochvil H. 1992. Significance of agonistic vocalization in the croaking gourami (Trichopsis vittatus, Teleostei) // Ethology. V. 90. № 4. P. 307–314. https://doi.org/10.1111/j.1439-0310.1992.tb00841.x
Lagardère J.P., Mallekh R. 2000. Feeding sounds of turbot (Scophthalmus maximus) and their potential use in the control of food supply in aquaculture. I. Spectrum analysis of the feeding sounds // Aquaculture. V. 189. № 3–4. P. 251–258. https://doi.org/10.1016/S0044-8486(00)00375-6
Liesch N., Ladich F. 2020. Both sexes produce sounds in vocal fish species: testing the hypothesis in the pygmy gourami (labyrinth fishes) // J. Exp. Biol. V. 223. № 10. Article jeb223750. https://doi.org/10.1242/jeb.223750
Pranata N.D., Purnama A.A., Yolanda R., Karno R. 2016. Iktiofauna Sungai Sangkir Kabupaten Rokan Hulu Provinsi Riau Ichthyofauna of Sangkir river, Rokan Hulu District Riau Province // Depik. V. 5. № 3. P. 100–106. https://doi.org/10.13170/depik.5.3.5574
Rainboth W.J. 1996. Fishes of the Cambodian Mekong. FAO species identification field guide for fishery purposes. Rome: FAO, 265 p.
Roberts T.R. 1989. The freshwater fishes of Western Borneo (Kalimantan Barat, Indonesia) // Mem. Calif. Acad. Sci. № 14. 210 p.
Scharrer E., Smith S.W., Palay S.L. 1947. Chemical sense and taste in the fishes, Prionotus and Trichogaster // J. Comp. Neurol. V. 86. № 2. P. 183–198. https://doi.org/10.1002/cne.900860204
Talde C.M., Mammaril Sr. A.C., Palomares M.L.D. 2004. The diet composition of some economically important fishes in the three floodplain lakes in Agusan Marsh wildlife sanctuary in the Philippines // Sri Lanka J. Aquat. Sci. V. 9. № 1. P. 45–56. https://doi.org/10.4038/sljas.v9i1.7465
Webb A., Maughan M., Knott M. 2007. Pest fish profiles: Trichogaster trichopterus − three spot gourami. Townsville: ACTFR, James Cook Univ., 5 p.
Zahid A. 2008. Ekologi trofik ikan-ikan dominan (Trichogaster leerii, T. trichopterus, dan Rasbora dusonensis) di hutan rawa gambut Desa Dadahup, Kalimantan Tengah. Bogor: Institut Pertanian Bogor. P. 75.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии