

Вопросы ихтиологии, 2022, T. 62, № 5, стр. 634-644
Реореакция Danio rerio (Cyprinidae): влияние скорости потока и доступности зоны без течения
Д. С. Павлов 1, В. Ю. Паршина 1, В. В. Костин 1, *
1 Институт проблем экологии и эволюции РАН ИПЭЭ РАН
Москва, Россия
* E-mail: povedenie@yandex.ru
Поступила в редакцию 05.05.2022
После доработки 12.05.2022
Принята к публикации 12.05.2022
- EDN: SZJOKA
- DOI: 10.31857/S0042875222050150
Аннотация
Представлены результаты экспериментального изучения влияния скорости потока на показатели реореакции Danio rerio – реопреферендум и соотношение частот проявления типов реореакции. При 0.30 от критической скорости течения для рыб их реопреферендум был положительным, при 0.80 – отрицательным. Соотношение типов реореакции на этих скоростях было характерным для резидентных рыб. При 0.45 от критической скорости течения реопреферендум не был выражен, а поведение рыб было характерным для особей, мигрирующих против течения. Снижение доступности лимнозоны (при уменьшении ширины входа в неё) в кольцевом лотке сокращает число возвращений рыб в неё и изменяет соотношение типов их реореакции. При максимальной ширине входа (19 см) в лимнозону соотношение типов реореакции данио отражает поведение, характерное для мигрирующих особей, при всех остальных вариантах (10, 3 и 1 см) – для резидентных особей. Показано, что реопреферендум является менее полной и надёжной оценкой реореакции, чем соотношение типов реореакции. Обсуждены области применения показателей реореакции.
Реореакция (отношение рыб к течению) – это врождённый поведенческий ответ рыб на течение. Она связана с жизнью рыб в подвижной среде и проявляется в выборе особями участка водоёма с оптимальными (для актуального поведения) скоростями течения и вектора собственной скорости в нём (Павлов и др., 2020а). Реореакция возможна только в определенном диапазоне скоростей течения – от пороговых до критических. При критических и более высоких скоростях течения рыб сносит потоком, а при подпороговых ориентация и вектор движения рыб не зависят от скорости течения (Павлов, 1979; Павлов, Скоробогатов, 2014).
Реореакция является одним из основных поведенческих механизмов распределения, кочёвок и миграций рыб (Pavlov et al., 2010; Павлов и др., 2010а, 2010б; Звездин, 2016). Простой оценкой отношения рыб к течению является такой показатель, как реопреферендум – предпочтение или избегание рыбами потока (Pavlov et al., 2001). Более полная оценка – это показатель соотношения частот проявления типов реореакции (далее соотношение типов реореакции): положительного (ПТР) – движение рыб против течения, отрицательного (ОТР) – движение по течению, компенсаторного (КТР) – сохранение положения рыб в потоке относительно неподвижных ориентиров (Maclean, Gee, 1971; Pavlov et al., 2010; Johnston et al., 2017; Zens et al., 2018; Павлов и др., 2020а). В последние годы был выделен факультативный тип реореакции (ФТР) – уход рыб из потока в зону со слабым (близким к пороговому для реореакции) течением или с его отсутствием (Павлов и др., 2020а). Наличие в экспериментальных установках такой лимнической зоны (лимнозоны) моделирует естественные условия в заливах и затишных зонах прибрежья рек.
Скорость течения является базовой характеристикой потока, и с ней связаны как возникновение реореакции, так и особенности её проявления. В текучих водоёмах имеется континуум скоростей, при этом рыбы отнюдь не всегда выбирают участки со скоростями течения в диапазоне от пороговых до критических. В зависимости от физиологического состояния и актуального поведения они периодически уходят с течения в зоны слабых и подпороговых скоростей – прибрежье, укрытия и заливы. Поэтому при экспериментальном исследовании реореакции целесообразно использовать установки, имеющие лимническую зону, позволяющую рыбам уходить из потока.
Основная цель работы – исследовать влияние скорости течения на реопреферендум и соотношение типов реореакции. Известно, что скорость течения влияет на соотношение проявляемых рыбами типов реореакции (Pavlov et al., 2010; Пономарева и др., 2017). Однако эти данные получены на установках без лимнозоны, т.е. в условиях искусственного ограничения возможности для рыб выхода из потока. Отсутствие лимнозоны искажает указанное соотношение, а в некоторых случаях может изменить даже преобладающий тип реореакции (Павлов и др., 2020а). Влияние может оказывать также и доступность лимнозоны для рыб. В связи с этим изучение влияния лимнозоны и её доступности на соотношение типов реореакции также стали задачами нашей работы. В качестве объекта исследования выбрали Danio rerio (Hamilton, 1822) (далее – данио). Этот вид широко используется во многих поведенческих, генетических, токсикологических, фармакологических и других исследованиях. При этом часть этих исследований основана на показателях поведения рыб в потоке воды (Lucon-Xiccato et al., 2021).
МАТЕРИАЛ И МЕТОДИКА
Исследования проводили в ноябре–декабре 2020 г. на данио стандартной длиной тела 21–32 (в среднем 26.4 ± ошибка среднего 0.5) мм. Рыб содержали в аэрируемых аквариумах при плотности посадки 1 экз/5 л воды. Температуру воды на уровне 22°C поддерживали терморегуляторами. Освещённость с 10 до 19 ч составляла 450–500 лк. В остальное время суток в аквариальном помещении искусственное освещение отсутствовало. Кормили рыб один раз в сутки после окончания опытов кормом для аквариумных рыб Tropical Fish Flakes (“Prodac”, Италия).
В экспериментах использовали две гидродинамические установки – двухканальный лоток для определения реопреферендума и кольцевой лоток с лимнозоной для оценки соотношения типов реореакции. При исследовании поведения рыб в потоке, в том числе их реореакции, скорости течения в установках должны быть нормированы в соответствии с плавательными возможностями исследуемых рыб. Для этого используется показатель “критическая скорость течения для рыб” (Vk) (Pavlov et al., 2010), которая перед началом выполнения работы была определена у 35 особей по стандартной методике (Павлов, 1979) в гидродинамической трубе длиной 1 м и диаметром 22 мм. В дальнейшем этих рыб в тестах не использовали. При плавном увеличении скорости потока находили такую скорость, при которой рыба переходила на бросковый режим плавания или её сразу сносило потоком. Время такого определения составляло ~30 с. Средняя критическая скорость для данио составила 0.35 ± 0.03 м/с. Скорости течения в обеих установках измеряли, используя гидрометрическую микровертушку (диаметр лопасти 8 мм) с контроллером для связи с компьютером. Температура воды и освещённость в экспериментальных установках были такими же, как в аквариумах.
Реопреферендум рыб определяли в двухканальной гидродинамической установке размерами 100 × 20 см с двумя каналами шириной по 10 см и длиной 35 см (рис. 1). Установку помещали в гидродинамический лоток, в котором создавали течение. Скорость течения в одном из каналов составляла 0.3 (0.11 м/с), 0.45 (0.16 ) или 0.8 Vk (0.28), в другом канале течение отсутствовало (Pavlov et al., 2001). В нижней по течению части установки располагался стартовый отсек, отгороженный по всей ширине лотка заградительной решёткой. Длина рабочей части лотка (от стартовой камеры до каналов) составляла 50 см.
Рис. 1.
Схема гидродинамической установки для определения реопреферендума рыб: 1 – канал с течением, 2 – канал без течения, 3 – съёмная заградительная решётка, 4 – стартовый отсек. Здесь и на рис. 2: (→) – направление течения, все линейные размеры приведены в см.
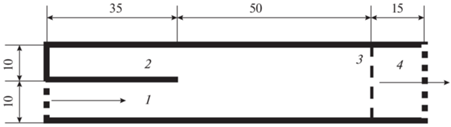
Одиночную рыбу помещали в стартовую камеру установки и через 3 мин акклимации снимали заградительную решётку (Pavlov et al., 2001). Эксперимент продолжали до первого захода рыбы в один из каналов, но не более 5 мин от момента подъёма заградительной решётки. Фиксировали один из возможных результатов опыта: выбор рыбой канала с течением (положительный реопреферендум), без течения (отрицательный реопреферендум) или отказ от захода в каналы.
Оценку соотношения типов реореакции проводили в кольцевом лотке с лимнозоной. Установка представляла собой кольцевой канал шириной 15 см и внешним радиусом 60 см (рис. 2). Длина канала по его центральной окружности радиусом 52.5 см–3.3 м. Высота стенок 15 см, внутренняя стенка выше 8 см сделана из капронового сита (ячея 0.9 мм). Через это сито вода стекала из рабочего канала в центральную ёмкость, откуда помпа Atman AT-107 (“Chuangxing Electrical Appliances”, Китай) возвращала её в рабочий канал установки. Высота слоя перелива составляла 2 см, а глубина воды в канале – 10 см. Через каждые 40 см на дно установки были нанесены линии шириной 1 см, обозначавшие границы секторов. Лимнозона диаметром 30 см располагалась с внутренней стороны канала. Влияние доступности лимнозоны на соотношение типов реореакции оценивали при разной ширине (19, 10, 3 и 1 см) входа/выхода из лимнозоны в канал, которую регулировали, используя задвижки.
Рис. 2.
Схема гидродинамической установки “кольцевой лоток с лимнозоной”: а – вид сверху,
б – разрез А–А, П – помпа, Л – лимнозона, 1–8 – секторы установки, (…) – границы секторов и лимнозоны, (‒ ‒ ‒) – часть стенки,
выполненная из капронового сита, ( ) – заполненная водой часть установки.
) – заполненная водой часть установки.
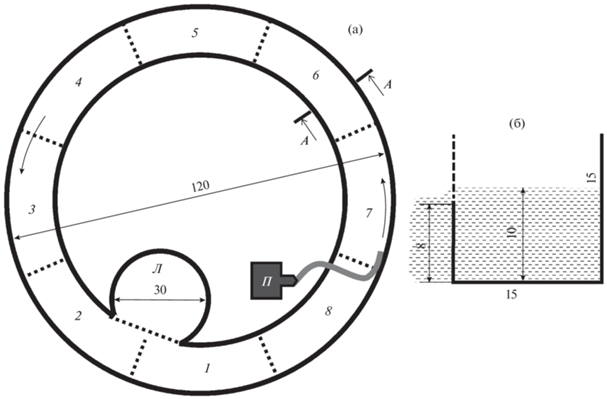
В канале установки возникал поперечный градиент скоростей течения с максимальной скоростью у внешней стенки. Рабочую скорость течения рассчитывали как среднюю по 14 замерам на глубинах 5 и 9 см в семи секторах, исключая сектор водоподачи (№ 7), на расстоянии 1 см от внешней стенки канала. Скорость и структуру потока в лимнозоне определяли, применяя видеосъёмку воды, подкрашенной перманганатом калия. Для расчёта скорости течения фиксировали время прохождения окрашенной водой полного круга по периметру лимнозоны. При всех использованных в установке скоростях течения наблюдали круговое течение в лимнозоне с максимальной скоростью у стенки: 0.016 (при рабочей скорости течения в установке 0.3 Vk), 0.031 (при 0.45 Vk) и 0.045 м/с (при 0.8 Vk).
Тесты проводили на единичных особях. Стартовым сектором служила лимнозона, которую перед помещением в неё опытной рыбы изолировали от канала сеткой. Время акклимации составляло 20 мин, после чего сетку стартового сектора убирали. Далее на протяжении 30 мин регистрировали движение особи видеокамерой смартфона Samsung Galaxy A10 SM-A105F (“Samsung Electronics”, Вьетнам), расположенного над установкой. При просмотре видеозаписей фиксировали направление и момент полного (всем телом) пересечения рыбой границы между секторами. Перемещения особи в пределах сектора не фиксировали.
Общая схема экспериментов. У каждой опытной рыбы вначале определяли реопреферендум и затем сразу – соотношение типов реореакции. Опыты по определению этих показателей проводили при трёх скоростях течения (0.3, 0.45 и 0.8 Vk). При каждой скорости течения тестировали разных особей. Ширина входа в лимнозону кольцевой установки в этих опытах была 19 см. Для выяснения влияния ширины входа в лимнозону на реореакцию тесты выполняли на других особях при скорости течения 0.45 Vk при четырёх вариантах ширины входа – 19, 10, 3 и 1 см. Объём материала по разным задачам исследования представлен в табл. 1.
Таблица 1.
Условия проведения и число опытов
| Предмет исследования | Установка. Показатель | Скорость течения, Vk | Ширина входа в лимнозону, см | Число индивидуальных опытов |
|---|---|---|---|---|
| Влияние скорости течения | Двухканальный лоток. Реопреферендум | 0.30 | 22 | |
| 0.45 | 29 | |||
| 0.80 | 22 | |||
| Кольцевой лоток. Соотношение типов реореакции | 0.30 | 19 | 22 | |
| 0.45 | 19 | 22 | ||
| 0.80 | 19 | 22 | ||
| Влияние ширины входа в лимнозону | Кольцевой лоток. Соотношение типов реореакции | 0.45 | 10 | 22 |
| 0.45 | 3 | 23 | ||
| 0.45 | 1 | 20 |
Примечание. Здесь и в табл. 2, 3 : Vk – критическая скорость течения (0.35 м/с).
По результатам экспериментов в кольцевом лотке рассчитывали следующие показатели.
1) Время (с) от начала опыта до первого выхода рыбы из лимнозоны.
2) Соотношение типов реореакции – соотношение экспериментальных оценок частот проявления типов реореакции (Pavlov et al., 2010). В кольцевых лотках эту частоту определяли по доле времени проявления реореакции определённого типа от длительности опыта (Павлов и др., 2020а). Расчёт проводили в два этапа.
На первом этапе определяли граничное время, которое необходимо для расчёта КТР, ПТР и ОТР (Павлов и др., 2020а). Граничное время (tg) – это оценка максимальной продолжительности прохода особью без остановок сектора кольцевого лотка. Его вычисляли отдельно для движения по течению и против него. Величину tg определяли по диаграмме частоты встречаемости времени, проведённого особью в секторе. Для этого классовый интервал увеличивали от 0.5 с до тех пор, пока частота первого класса не станет больше в семь или более раз (экспертная оценка), чем второго класса. Если особь покидала сектор после истечения граничного времени, мы считали, что она проявляла реореакцию компенсаторного типа в течение реального времени пребывания в секторе минус величина граничного времени.
На втором этапе вычисляли частоту проявления реореакции (Р) разных типов относительно длительности опыта по формулам: РПТР = (kutg + + ∑tu + ∑tr/2)/T, РОТР = (kdtg + ∑td + ∑tr/2)/T, РКТР = = ∑(t – tg)/T, РФТР = ∑tz/T, где T – длительность опыта, с; tg – граничное время, с; tu – время пребывания рыбы в секторе длительностью <tg при сквозном проходе сектора против течения, с; td – время пребывания в секторе длительностью <tg при сквозном проходе сектора по течению, с; tr – время пребывания в секторе длительностью <tg при развороте (вход и выход через одну границу сектора), с; t – время пребывания в секторе длительностью >tg, с; ku – число переходов в верхний по течению сектор после проявления КТР; kd – число переходов в нижний по течению сектор после проявления КТР; tz – время пребывания в лимнозоне, с.
3) Для расчёта показателей пути рыб точкой старта была граница между первым и вторым секторами. Общий путь (S) – произведение числа пройденных секторов на длину сектора – 0.275 м (длина центральной дуги сектора). Кроме того, аналогично рассчитывали длину пути по течению (Sd) и против него (Su). Передвижение рыб внутри секторов и в лимнозоне не учитывали.
4) Перемещение особи – разность пути против течения и по течению.
5) Для сравнимости с результатами других исследований рассчитывали индекс перемещения (Id) как частное от деления перемещения на общий путь: Id = (Su – Sd)/S. Величина индекса изменяется от +1 при движении рыбы только против течения, до −1 при движении рыбы только по течению.
6) Частоту смены типов реореакции рассчитывали как частное от деления числа изменений типа реореакции на время опыта в минутах. Чем меньше этот показатель, тем в среднем стабильнее у рыбы проявление реореакции исследованных типов.
7) Длительность движения рыб по разным участкам кругового канала определяли по видеозаписям. Оценивали время движения рыб у внутренней и внешней стенок (на расстоянии не более 1.5 см от стенки), а также в остальной (центральной) части канала. Показатель рассчитывали в процентах общего времени опыта.
Статистический анализ данных проводили с использованием пакета программ STATISTICA 10.0 (StatSoft, Inc., США). Обработку материала выполняли с применением критерия Стьюдента для долей по доле рыб с положительным или отрицательным реопреферендумом и по средним частотам типов реореакции (Лакин, 1973), H-критерия Краскела–Уоллиса, U-критерия Манна−Уитни и одновыборочного t-критерия Стьюдента по индивидуальным значениям показателей у каждой особи. Множественную проверку гипотез проводили, учитывая поправки Бонферрони или Холма (Kutner et al., 2005) в зависимости от числа сравниваемых субвыборок.
РЕЗУЛЬТАТЫ
Влияние скорости потока
В двухканальном лотке при скорости течения 0.3 Vk доля рыб, проявляющих положительный реопреферендум, была максимальна, а отрицательный – минимальна (рис. 3). Это указывает на привлекательность для данио малых скоростей течения. При скорости течения 0.45 Vk данио равновероятно выбирали каналы с течением и без него. Отказ от выбора зафиксирован всего у одной из 29 особей. При скорости 0.8 Vk данио избегали потока – доля рыб с отрицательным реопреферендумом была максимальна и превышала долю рыб с положительным значением данного показателя. Также максимальна была и доля рыб, отказавшихся от выбора.
Рис. 3.
Реопреферендум Danio rerio при разных скоростях течения: ( ) – положительный (доля рыб, выбравших канал с течением), (
) – положительный (доля рыб, выбравших канал с течением), ( ) – отрицательный (доля рыб, выбравших канал без течения), (
) – отрицательный (доля рыб, выбравших канал без течения), ( ) – отказ (доля рыб, не выбравших ни один канал за 5 мин опыта), Vk – критическая скорость течения (0.35 м/с). При скорости течения 0.30 и 0.80 Vk различия числа рыб с положительным и отрицательным реопреферендумом достоверны (р < 0.05, критерий Стьюдента для долей). При скорости 0.45 Vk различия недостоверны (р < 0.43).
) – отказ (доля рыб, не выбравших ни один канал за 5 мин опыта), Vk – критическая скорость течения (0.35 м/с). При скорости течения 0.30 и 0.80 Vk различия числа рыб с положительным и отрицательным реопреферендумом достоверны (р < 0.05, критерий Стьюдента для долей). При скорости 0.45 Vk различия недостоверны (р < 0.43).

Визуальные наблюдения в течение 7–10 мин после окончания опыта показали, что рыбы могли неоднократно заходить в каждый из каналов, возвращаться в стартовую зону и вновь выбирать один из каналов. Большинство рыб не задерживались надолго ни в одном из каналов. Также в течение опыта некоторые рыбы проявляли реореакцию компенсаторного типа – стояли у стенок установки или в середине лотка на границе потока. После окончания опыта такие рыбы могли выбрать один из каналов или продолжать указанное поведение.
В кольцевой установке длительность движения рыб по центру или у стенок кругового канала зависела от скорости потока: чем больше его скорость, тем дольше (p = 0.02, критерий Стьюдента для долей) рыбы находились у внутренней стенки канала и меньше у внешней (табл. 2). При движении вниз по течению рыбы были ориентированы головой как против течения, так и по нему.
Таблица 2.
Средняя длительность движения Danio rerio по центру и у стенок канала кольцевой установки, % времени опыта (30 мин)
| Участки движения рыб в круговом канале |
Скорость течения, Vk | ||
|---|---|---|---|
| 0.30 | 0.45 | 0.80 | |
| У внутренней стенки | 33.3 ± 7.0 | 54.1 ± 3.8 | 75.0 ± 0 |
| У внешней стенки | 42.1 ± 7.9 | 40.6 ± 4.9 | 25.0 ± 0 |
| По центру канала | 24.6 ± 3.5 | 5.3 ± 1.4 | 0 |
Время первого выхода из лимнозоны зависело от скорости течения в установке. При скоростях течения 0.3 и 0.45 Vk оно не превышало 1 мин и статистически не различалось (скорректированный р = 0.34, U-критерий Манна–Уитни). При скорости течения 0.8 Vk рыбы достоверно дольше задерживались в лимнозоне (скорректированный p < 0.001) (табл. 3 ).
Таблица 3.
Показатели реореакции Danio rerio при разных скоростях течения в кольцевом лотке с лимнозоной (M ± m)
| Показатель | Скорость течения, Vk | ||
|---|---|---|---|
| 0.30 | 0.45 | 0.80 | |
| Время первого выхода из лимнозоны, с | 58 ± 18 | 37 ± 11 | 328 ± 87 |
| Число повторных заходов в лимнозону за опыт | 5.55 ± 1.11 | 5.50 ± 0.76 | 3.09 ± 0.51 |
| Частота проявления реореакции динамических типов: | |||
| – ПТР | 0.27 ± 0.05 | 0.53 ± 0.04 | 0.17 ± 0.03 |
| – ОТР | 0.07 ± 0.03 | 0.13 ± 0.02 | 0.10 ± 0.02 |
| Частота проявления реореакции статических типов: | |||
| – КТР | 0.51 ± 0.06 | 0.24 ± 0.03 | 0.24 ± 0.04 |
| – ФТР | 0.15 ± 0.03 | 0.10 ± 0.02 | 0.49 ± 0.08 |
| Индекс перемещения | 0.36 ± 0.08 | 0.29 ± 0.07 | −0.23 ± −0.05 |
| Частота смены типов реореакции, мин−1 | 2.40 ± 0.46 | 4.09 ± 0.43 | 2.50 ± 0.41 |
Примечание. Частота проявления реореакции всех типов достоверно (p < 0.03, Н-критерий Краскела–Уоллиса) зависела от скорости течения в установке. Во всех случаях индекс перемещения достоверно отличался от нуля (одновыборочный t-критерий Стьюдента, p < 0.05). Здесь и в табл. 4: Типы реореакции: ПТР – положительный, ОТР – отрицательный, КТР – компенсаторный, ФТР – факультативный; M ± m – среднее значение и его ошибка.
На скорости течения 0.45 Vk динамические типы реореакции преобладали по частоте проявления (0.66 в сумме) преимущественно за счёт ПТР. При скоростях течения 0.3 и 0.8 Vk была больше частота проявления рыбами реореакции статических типов – соответственно 0.66 и 0.73. При скорости течения 0.3 Vk преобладал КТР, а при 0.8 Vk – ФТР (табл. 3 ). Индекс перемещения у данио оказался сходным (р = 0.62, U-критерий Манна–Уитни) на скорости течения 0.3 и 0.45 Vk и достоверно меньше на 0.8 Vk (скорректированный р < 2 × 10−6). Частота смены типов реореакции была максимальна при скорости 0.45 Vk и достоверно отличалась от таковой при других скоростях течения (p < 0.05, Н-критерий Краскела–Уоллиса).
Сравнение соотношения типов реореакции у рыб с положительным и отрицательным реопреферендумом (рис. 4) провели при скорости течения 0.45 Vk. Эта скорость течения не являлась для данио ни привлекательной, ни избегаемой – при ней рыбы равновероятно проявляли положительный и отрицательный реопреферендум.
Рис. 4.
Частота проявления реореакции разного типа у Danio rerio с положительным ( ) и отрицательным (
) и отрицательным ( ) реопреферендумом. Типы реореакции: ПТР – положительный, ОТР – отрицательный, КТР
– компенсаторный, ФТР – факультативный. ($ \bot - $) – ошибка средней.
) реопреферендумом. Типы реореакции: ПТР – положительный, ОТР – отрицательный, КТР
– компенсаторный, ФТР – факультативный. ($ \bot - $) – ошибка средней.

Оказалось, что соотношение типов реореакции не различается у рыб с положительным и с отрицательным реопреферендумом (р > 0.53 для всех типов реореакции, U-критерий Манна–Уитни).
Влияние лимнозоны и её доступности
Исследование проведено при рабочей скорости течения в установке 0.45 Vk. В лимнозоне структура и скорость течения различались в зависимости от ширины входа в неё (рис. 5). При малой ширине входа циркуляционное течение захватывало лишь часть объёма лимнозоны. Минимальная скорость течения была при ширине входа 3 см и наблюдалась по всей глубине, но только до половины периметра, а при ширине входа 1 см скорость была несколько выше, но её фиксировали только у поверхности.
Рис. 5.
Скорость течения по периметру лимнозоны в зависимости от ширины входа в неё. Круговое течение наблюдалось: 1 – во всей лимнозоне от поверхности до дна, 2 – от поверхности до дна, но только в части (половине) периметра, 3 – во всей лимнозоне, но только в поверхностных слоях воды.

В начале каждого опыта при любой ширине входа рыба выходила из лимнозоны в кольцевой канал. Зависимость времени первого выхода из лимнозоны от ширины входа не линейная. Доля рыб, не заходивших повторно в лимнозону, была обратно пропорциональна ширине (табл. 4). Эта доля при ширине входа 1 см достоверно (скорректированный р < 7.5 × 10−5, критерий Стьюдента для долей) отличалась от таковых при остальных величинах ширины входа.
Таблица 4.
Показатели реореакции Danio rerio в кольцевом лотке при разной ширине входа в лимнозону (M ± m)
| Показатель | Ширина входа в лимнозону, см | |||
|---|---|---|---|---|
| 19 | 10 | 3 | 1 | |
| Время первого выхода из лимнозоны, с | 37 ± 11.1 | 239 ± 57.0 | 203 ± 73.0 | 64 ± 21.0 |
| Число повторных заходов в лимнозону за опыт | 5.5 ± 0.76 | 3.5 ± 0.60 | 1.7 ± 0.52 | 0.1 ± 0.07 |
| Частота проявления реореакции динамических типов: | ||||
| – ПТР | 0.53 ± 0.04 | 0.21 ± 0.05 | 0.20 ± 0.04 | 0.22 ± 0.05 |
| – ОТР | 0.13 ± 0.02 | 0.09 ± 0.02 | 0.04 ± 0.06 | 0.04 ± 0.01 |
| Частота проявления реореакции статических типов: | ||||
| – КТР | 0.24 ± 0.03 | 0.41 ± 0.07 | 0.55 ± 0.06 | 0.68 ± 0.05 |
| – ФТР | 0.10 ± 0.02 | 0.29 ± 0.06 | 0.21 ± 0.06 | 0.06 ± 0.02 |
| Индекс перемещения | 0.59 ± 0.05 | 0.22 ± 0.10 | 0.52 ± 0.06 | 0.59 ± 0.06 |
| Частота смены типов реореакции, мин−1 | 4.09 ± 0.43 | 2.34 ± 0.22 | 1.62 ± 0.24 | 0.91 ± 0.14 |
| Доля рыб, не зашедших повторно в лимнозону, % | 9 | 18 | 35 | 90 |
Примечание. Во всех случаях индекс перемещения достоверно отличался от нуля (одновыборочный t-критерий Стьюдента, p < 0.05). Частота проявления реореакции всех типов достоверно (p $ \ll $ 0.001, Н-критерий Краскела–Уоллиса) зависела от ширины входа в лимнозону. Частота смены типов реореакции и число повторных заходов в лимнозону достоверно (скорректированный p < 0.03, U-критерий Манна–Уитни) сокращается при уменьшении ширины входа.
При максимальной ширине входа (19 см) суммарная частота динамических типов реореакции преобладала над частотой статических (0.66 против 0.34) (табл. 4). При всех остальных вариантах ширины входа преобладали статические типы. При уменьшении ширины входа в лимнозону снижалась частота ПТР и увеличивалась частота КТР. Частота смены типов реореакции и количество повторных заходов в лимнозону сокращалась при уменьшении ширины входа.
ОБСУЖДЕНИЕ
Перемещение рыб в водотоках определяется двумя комплексами поведения по отношению к течению – миграционным и резидентным. При миграционном поведении рыбы покидают один участок обитания и перемещаются в другой, то есть совершают миграции или кочёвки (локальные перемещения) (Lucas, Baras, 2008). При резидентном поведении рыбы перемещаются в пределах одного участка обитания.
Реореакция рыб, взятых из природных водоёмов, в гидродинамической установке отражает эти два комплекса поведения (Pavlov et al., 2010; Павлов и др., 2013, 2019; Звездин, 2016). Для различения миграционного и резидентного поведения в экспериментах мы предлагаем формальный критерий, который позволяет оценить, какое поведение рыбы проявляли в опыте.
Этот критерий основан на наблюдениях и результатах экспериментов с дикими рыбами. У рыб, совершающих в природе контранатантные или денатантные миграции, или кочёвки, в гидродинамической установке преобладает частота одного из динамических типов реореакции – ПТР или ОТР. Преобладание частоты статических типов (КТР и ФТР) соответствует резидентному поведению, при котором рыбы сохраняют свое местообитание. При этом индекс перемещения, который отражает вектор перемещения рыб, должен быть больше нуля при контранатантном перемещении, меньше нуля при денатантном и равен нулю при резидентном поведении. Следовательно, в кольцевом лотке с лимнозоной признаки миграционного поведения рыб могут быть выражены следующим образом: 1. (РПТР+ РОТР) > (РКТР+ РФТР); 2. Id ≠ 0.
Рассмотрим наши результаты с учётом это критерия.
Скорость 0.3 Vk (0.11 м/с) наиболее привлекательна для данио, поскольку при этой скорости в двухканальной установке рыбы чаще всего выбирают канал с течением, а в кольцевой установке бо́льшую часть времени проводят в потоке, проявляя КТР. На этой скорости течения у данио преобладают статические типы реореакции (суммарная частота 0.66, табл. 3 ); согласно предложенному критерию, первое условие не выполняется, следовательно, при этой скорости течения рыбы сохраняют местоположение в потоке. В природе данио встречаются в слабо текучих или стоячих водоёмах, низовьях ручьёв, каналах, канавах, прудах и рисовых чеках (Rahman, 1989; Talwar, Jhingran, 1991; McClure et al., 2006). Привлекательность течения в таких водоёмах связана, скорее всего, с питанием рыб. Данио питаются в основном зоопланктоном и упавшими в воду мелкими насекомыми. Известно, что питание рыб зоопланктоном в слабом потоке более эффективно, чем в стоячей воде (Михеев, Пакульская, 1988; Lupandin et al., 2003, 2004).
При скорости течения 0.45 Vk (0.16 м/с) у данио реопреферендум не выражен – в двухканальной установке они равновероятно выбирают каналы с течением и без него. Соотношение типов реореакции не различается у рыб, проявивших положительный и отрицательный реопреферендум. Поскольку выполняются оба условия критерия (суммарная частота (0.66) реореакции динамических типов больше, чем статических, и индекс перемещения не равен нулю), следовательно, при скорости 0.45 Vk поведение рыб было характерно для особей, мигрирующих против течения.
При скорости течения 0.8 Vk (0.28 м/с) в двухканальной установке реопреферендум у данио отрицательный, а преобладающий тип реореакции – ФТР (уход в лимнозону). В природе данио также избегают потоки с большими скоростями, хотя эти рыбы встречаются и в районах с сильным течением (возле порогов и водопадов). Но в этих местах данио держатся в более спокойных зонах – в прибрежье рядом с растительностью (Hora, 1930; Catalog …, 1998). На скорости 0.8 Vk у данио преобладают статические типы реореакции (в сумме 0.73). По предложенному критерию эти особи сохраняли своё место нахождения, т. е. данио сохраняли местообитание как при 0.8 Vk, так и при 0.3 Vk, однако в первом случае за счёт увеличения частоты ФТР, а во втором – КТР. Сравнение использованных показателей реореакции указывает, что методика определения реопреферендума позволяет оценить только предпочитаемые и избегаемые скорости потока.
Ранее было установлено, что личинки плотвы Rutilus rutilus в период покатной миграции делятся на две группировки – мигрантов и резидентов (Pavlov et al., 2001). Реопреферендум личинок адекватно отражал предпочтение потока или его избегание как у мигрантов, так и у резидентов в пределах суточного “окна миграции”. Покатная миграция личинок плотвы проходит в сумеречно-ночное время. При освещённости 1 лк и меньше мигранты проявляли положительный реопреферендум – выходили в русловой поток для миграции. Резиденты демонстрировали отрицательный реопреферендум, благодаря которому они уходили в участки прибрежья без течения и не мигрировали. При утреннем повышении освещённости реопреферендум мигрантов становился отрицательным, они прекращали миграцию и уходили из потока в зоны без течения. Резиденты, напротив, проявляли положительный реопреферендум и выходили для питания на течение у границы руслового потока и прибрежья. Определение реопреферендума позволило понять поведенческий механизм пространственного перераспределения резидентов и покатных мигрантов личинок плотвы в течение суток, которое обеспечивало их участие или неучастие в миграции. С другой стороны, по реопреферендуму невозможно различить резидентов, питающихся на границе потока, и контранатантных мигрантов. В светлое время суток такие мигранты так же, как и резиденты, проявляют положительный реопреферендум. Однако, войдя в поток, первые мигрируют, проявляя реореакцию положительного типа, а резиденты питаются, оставаясь на месте (проявление КТР). То есть по реопреферендуму можно судить только о переходах рыб из прибрежья в поток или обратно. Тогда как по соотношению типов реореакции можно судить и о таких переходах, и о поведении рыб в потоке. Поэтому реопреферендум менее полно, чем соотношение типов реореакции, характеризует последнюю.
Репрезентативность используемых методов можно оценить по количеству выборов, сделанных рыбой в потоке воды. При определении реопреферендума учитывали только единственный (первый) выбор рыбой варианта поведения. При проведении опытов по определению соотношения типов реореакции рыба могла многократно проявлять разные типы реореакции (Pavlov et al., 2010; Павлов и др., 2020а), т.е. она могла неоднократно выбирать разные варианты поведения в потоке воды. В среднем за время эксперимента рыба могла 12311 раза переключаться на разные типы реореакции, т.е. в опытах по определению соотношения типов реореакции число выборов рыбой вариантов поведения более чем в 100 раз превышало таковое в экспериментах по определению реопреферендума. Следовательно, соотношение типов реореакции более точно, чем реопреферендум, отражало реореакцию рыб.
В нашей предыдущей работе мы показали, что для оценки соотношения типов реореакции у разных видов рыб необходимо использовать скорости течений, которые лежат в диапазоне 0.2–0.7 Vk (Pavlov et al., 2010). Эти значения мы получили в опытах на рыбах, тестируемых в установках типа “Рыбоход”. В настоящей работе скорость течения, которое рыба избегает, составляла 0.8 Vk. Это значение выходило за пределы рекомендованных выше диапазонов, и при такой скорости течения рыба сохраняла своё местоположение. Такое же поведение рыб мы наблюдали при скорости течения 0.3 Vk, что близко к нижней границе рекомендованных скоростей. Можно предположить, что для рыбы, тестируемой в установках типа “кольцевой лоток с лимнозоной”, диапазон допустимых скоростей течения ýже, чем это было известно ранее, и скорости течения, предпочитаемые рыбами, в этот диапазон не входят.
Ранее мы исследовали влияние наличия лимнозоны на соотношение типов реореакции у данио с использованием нескольких гидродинамических установок – “Рыбоход” и кольцевых лотков с лимнозоной и без неё (Павлов и др., 2020б). Наличие у рыб возможности проявлять реореакцию факультативного типа существенно снижало частоту динамических типов реореакции и повышало статических типов. В установках без лимнозоны данио демонстрировали миграционное поведение, а при наличии лимнозоны – резидентное. В данной работе мы впервые изучили влияние степени доступности лимнозоны на проявление рыбами реореакции всех четырёх типов. При использованных размерах входа/выхода (19, 10, 3 и 1 см) все особи выходили из лимнозоны в начале опыта. При этом минимальное время на первый выход затратили рыбы при максимальной и минимальной ширине выхода (соответственно 37 и 64 с). Вероятно, это связано с тем, что гидравлические условия в лимнозоне при ширине выхода 1 см обеспечивали привлечение рыб к нему за счёт выраженного вертикального градиента скоростей: наибольшие скорости в лимнозоне были отмечены в верхних слоях. Уменьшение ширины входа в лимнозону существенно снижает возможность проявления реореакции факультативного типа – количество повторных заходов в лимнозону уменьшается прямо пропорционально ширине входа. При ширине входа 1 см число заходов сокращается в 55 раз по сравнению с максимальной шириной (19 см).
При небольшой (1 см) ширине входа в лимнозону рыбы преимущественно проявляют КТР у внутренней стенки кольцевого канала. При этом у них заметно сокращается время проявления реореакции положительного типа, вероятно, из-за отсутствия отдыха в лимнозоне. В результате уменьшения возможности проявления реореакции факультативного типа поведение данио кардинально изменяется – вместо миграционного поведения наблюдается резидентное. Эти результаты хорошо согласуются с ранее полученными данными. Как полное отсутствие лимнозоны (Павлов и др., 2020а), так и снижение её доступности вызывают изменение исходного поведения рыб (миграционного или резидентного) на противоположное. Полученные результаты имеют важное методическое значение: для корректного применения методики оценки соотношения типов реореакции необходимо использовать лимнозону с шириной входа не менее 19 см.
В качестве модельного объекта данио используют в широком круге исследований. При этом часто применяют поведенческие тесты, в том числе с использованием показателей, отражающих локомоторные возможности рыб. Однако поведение рыб в потоке прежде всего определяется физиологическим состоянием и выбором типа реореакции, которые быстро изменяются при изменении условий содержания рыб. Определение соотношения типов реореакции успешно использовано для оценки влияния ряда факторов (голод, рацион, гормональные и химические воздействия) на миграционное поведение рыб (Павлов Д. и др., 2010а, 2016, 2018, 2020б, 2020в; Пономарева, 2014; Павлов Е. и др., 2016). Было показано, что у рыб возникновение миграционного поведения может быть обусловлено неблагоприятными внешними условиями различной природы. Это позволяет использовать данную методику для выявления отношения особей к условиям обитания. Если условия воспринимаются особью как неблагоприятные, она стремится покинуть данное место.
В целом рассмотренные в данной работе методики могут дать новые возможности для быстрой диагностики изменений состояния рыб, находящихся под экспериментальным воздействием. Для оценки соотношения типов реореакции целесообразно использовать скорости течения ~ 0.5 Vk.
ВЫВОДЫ
1. Показатели реореакции данио – реопреферендум и соотношение типов реореакции – зависят от скорости течения. При скоростях течения 0.3 Vk реопреферендум рыб был положительным, при скорости 0.8 Vk – отрицательным. При этом соотношение типов реореакции на этих скоростях было характерным для резидентных рыб. При скорости течения 0.45 Vk у исследованных рыб реопреферендум не был выражен. На этой скорости соотношение типов реореакции изменялось, и у рыб отмечали поведение, характерное для особей, мигрирующих против течения.
2. Снижение доступности лимнозоны (при уменьшении ширины входа в неё), необходимой для проявления реореакции факультативного типа в кольцевом лотке, снижает прежде всего число возвращений рыб в неё и резко изменяет соотношение типов реореакции рыб. При максимальной ширине входа в лимнозону (19 см) соотношение типов реореакции данио отражает поведение, характерное для мигрирующих особей, при всех остальных вариантах ширины входа – для резидентных особей.
3. Реопреферендум рыб является менее полной и менее точной оценкой реореакции по сравнению с соотношением типов реореакции.
Список литературы
Звездин А.О. 2016. Реореакция ранней молоди нерки Oncorhynchus nerka (Walb.) в период расселения с нерестилищ: Автореф. дис. … канд. биол. наук. М.: ИПЭЭ РАН, 28 с.
Лакин Г.Ф. 1973. Биометрия. М.: Высш. шк., 352 с.
Михеев В.Н., Пакульская Д.С. 1988. Двигательная активность в пищедобывательном поведении личинок Hemigrammus caudovittatus // Вопр. ихтиологии. Т. 28. № 6. С. 1007–1013.
Павлов Д.С. 1979. Биологические основы управления поведением рыб в потоке воды. М.: Наука, 319 с.
Павлов Д.С., Скоробогатов М.А. 2014. Миграции рыб в зарегулированных реках. М.: Т-во науч. изд. КМК, 413 с.
Павлов Д.С., Костин В.В., Пономарева В.Ю. 2010а. Поведенческая дифференциация сеголеток черноморской кумжи Salmo trutta labrax: реореакция в год, предшествующий смолтификации // Вопр. ихтиологии. Т. 50. № 2. С. 251−261.
Павлов Д.С., Пономарева В.Ю., Веселов А.Е., Костин В.В. 2010б. Реореакция как один из механизмов формирования фенотипических группировок сеголеток атлантического лосося Salmo salar // Там же. Т. 50. № 4. С. 548–553.
Павлов Д.С., Звездин А.О., Костин В.В. 2013. Тип реореакции ранней молоди нерки Oncorhynchus nerka в реке Озерная и озере Курильское // Там же. Т. 53. № 1. С. 87–95. https://doi.org/10.7868/S0042875213010104
Павлов Д.С., Павлов Е.Д., Ганжа Е.В., Костин В.В. 2016. Реореакция молоди радужной форели при воздействии сурфагона // Онтогенез. Т. 47. № 2. С. 110–116. https://doi.org/10.7868/S0475145016020075
Павлов Д.С., Павлов Е.Д., Ганжа Е.В. и др. 2018. Влияние сурфагона на реореакцию молоди кумжи Salmo trutta // Вопр. ихтиологии. Т. 58. № 6. С. 740–744. https://doi.org/10.1134/S004287521806019X
Павлов Д.С., Костин В.В., Звездин А.О. и др. 2019. Реореакция молоди некоторых карповых рыб (Cyprinidae) в период осенней контранатантной миграции // Там же. Т. 59. № 6. С. 716–723. https://doi.org/10.1134/S0042875219060122
Павлов Д.С., Паршина В.Ю., Костин В.В., Прозоров Д.А. 2020а. Сравнение экспериментальных методов оценки мотивационной компоненты реореакции рыб (соотношения типов реореакции) // Там же. Т. 60. № 4. С. 478–487. https://doi.org/10.31857/S0042875220040189
Павлов Д.С., Костин В.В., Павлов Е.Д., Кравченко А.А. 2020б. Влияние изменения рациона на реореакцию молоди радужной форели Oncorhynchus mykiss // Там же. Т. 60. № 3. С. 364–367. https://doi.org/10.31857/S0042875220030157
Павлов Д.С., Павлов Е.Д., Ганжа Е.В., Костин В.В. 2020в. Изменение реореакции и содержания тиреоидных гормонов в крови молоди радужной форели Oncorhynchus mykiss при голодании // Там же. Т. 60. № 2. С. 229–234. https://doi.org/10.31857/S0042875220020186
Павлов Е.Д., Ганжа Е.В., Пономарева В.Ю. и др. 2016. Влияние метилтестостерона на физиологическое состояние и реореакцию радужной форели Parasalmo mykiss (=Oncorhynchus mykiss) при неблагоприятных условиях содержания // Там же. Т. 56. № 6. С. 740–752. https://doi.org/10.7868/S0042875216060072
Пономарева В.Ю. 2014. Поведенческие механизмы внутрипопуляционной дифференциации молоди некоторых лососевых рыб: Автореф. дис. … канд. биол. наук. М.: ИПЭЭ РАН, 28 с.
Пономарева В.Ю., Павлов Д.С., Костин В.В. 2017. Разработка и апробирование методики исследования соотношения типов реореакции рыб в кольцевом гидродинамическом лотке // Биология внутр. вод. № 1. С. 100–108. https://doi.org/10.7868/S0320965217010156
Eschmeyer W.N. 1998. Catalog of Fishes. V. 1–3. San Francisco: Calif. Acad. Sci., 2905 p.
Hora S. 1930. Ecology, bionomics and evolution of the torrential fauna, with special reference to the organs of attachment // Philos. Trans. R. Soc. Lond. B: V. 218. № 450–461. P. 171–282. https://doi.org/10.1098/rstb.1930.0005
Johnston M.E., Kelly J.T., Lindvall M.E. et al. 2017. Experimental evaluation of the use of vision and barbels as references for rheotaxis in green sturgeon // J. Exp. Mar. Biol. Ecol. V. 496. P. 9–15. https://doi.org/10.1016/j.jembe.2017.04.002
Kutner M.H., Nachtsheim C.J., Neter J., Li W. 2005. Applied linear statistical models. N.Y.: McGraw-Hill Irwin, 1396 p.
Lucas M., Baras E. 2001. Migration of freshwater fishes. Oxford: Blackwell Sci., 420 p. https://doi.org/10.1002/9780470999653
Lucon-Xiccato T., Bella L., Mainardi E. et al. 2021. An automated low-cost swim tunnel for measuring swimming performance in fish // Zebrafish. V. 18. № 3. P. 231–234. https://doi.org/10.1089/zeb.2020.1975
Lupandin A.I., Kirillov P.I., Pavlov D.S. 2003. Experimental study of feeding of the dace Leuciscus leuciscus at different current velocities // J. Ichthyol. V. 43. Suppl. 2. P. S255–S258.
Lupandin A.I., Kirillov P.I., Pavlov D.S. 2004. Foraging efficiency of chub Leuciscus cephalus at different flow rates and food concentration // Ibid. V. 44. Suppl. 2. P. S170–S174.
Maclean J.A., Gee J.H. 1971. Effect of temperature on movements of pre-spawning brook sticklebacks, Culaea inconstans, in the Roseau River, Manitoba // J. Fish. Res. Board Can. V. 28. № 6. P. 919–923. https://doi.org/DOI:10.1139/f71-133
McClure M.M., McIntyre P.B., McCune A.R. 2006. Notes on the natural diet and habitat of eight danionin fishes, including the zebrafish Danio rerio // J. Fish Biol. V. 69. № 2. P. 553–570. https://doi.org/10.1111/j.1095-8649.2006.01125.x
Pavlov D.S., Lupandin A.I., Kostin V.V. et al. 2001. Downstream migration and behavior of juvenile roach Rutilus rutilus (Cyprinidae) from two phenotypic groups // J. Ichthyol. V. 41. Suppl. 2. P. S133–S179.
Pavlov D.S., Kostin V.V., Zvezdin A.O., Ponomareva V.Yu. 2010. On methods of determination of the rheoreaction type in fish. // Ibid. V. 50. № 11. P. 977–984. https://doi.org/10.1134/S0032945210110020
Rahman A.K.A. 1989. Freshwater fishes of Bangladesh. Dhaka: Zool. Soc. Bangladesh et al., 364 p.
Talwar P.K., Jhingran A.G. 1991. Inland fishes of India and adjacent countries. V. 1–2. New Delhi: Oxford and IBH Publ., 1158 p.
Zens B., Glas M., Tritthart M. et al. 2018. Movement patterns and rheoreaction of larvae of a fluvial specialist (nase, Chondrostoma nasus): the role of active versus passive components of behaviour in dispersal // Can. J. Fish. Aquat. Sci. V. 75. № 2. P. 193–200. https://doi.org/10.1139/cjfas-2016-0276
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии