

Вопросы ихтиологии, 2023, T. 63, № 2, стр. 152-169
Биология и распространение звёздчатой Benthophilus stellatus и донской B. durrelli (Teleostei: Gobiidae) пуголовок
В. С. Болдырев 1, *, Ю. В. Басько 1, Е. В. Випхло 1
1 Волгоградский филиал Всероссийского научно-исследовательского института
рыбного хозяйства и океанографии – ВолгоградНИРО
Волгоград, Россия
* E-mail: neogobius@yahoo.com
Поступила в редакцию 11.04.2022
После доработки 18.05.2022
Принята к публикации 07.06.2022
- EDN: EXMTPC
- DOI: 10.31857/S0042875223010010
Аннотация
Приведена характеристика размерно-возрастной и половой структур аборигенных в Донском бассейне звёздчатой Benthophilus stellatus и донской B. durrelli пуголовок. Самки донской пуголовки созревают при длине около 35, звёздчатой – 50 мм. Для основной части (~74%) производителей донской пуголовки характерен единовременный нерест, звёздчатая откладывает три порции икры. Продолжительность периода размножения у этих видов составляет около 3.5 и 1.5 мес. соответственно, а индивидуальная абсолютная плодовитость самок – 0.2–0.6 и 1–3 тыс. икринок. Все производители донской пуголовки погибают после нереста на втором году жизни, незначительная часть особей звёздчатой пуголовки доживает до третьего года. Оценены спектры питания, размерная и биотопическая изменчивость, степень пищевого сходства, избирательность кормовых организмов у обоих видов. Для них характерны сходство питания и высокая пищевая пластичность. В Цимлянском водохранилище у пуголовок основными кормовыми объектами являются моллюск Lithoglyphus naticoides и личинки комаров, в нижнем течении р. Дон – различные ракообразные. Обосновывается предположение о том, что в настоящее время в водохранилищах Волжского каскада обитает три вида пуголовок (донская, звёздчатая и Махмудбекова B. mahmudbejovi). Донская пуголовка расселилась и в нижнем течении р. Волга. Её ареал помимо нативного Азовского и новоприобретённого Волжского бассейнов, по-видимому, включает в себя и Черноморский.
В настоящее время род Benthophilus включает в себя 21 вид демерсальных бентосоядных бычковых рыб (Boldyrev, Bogutskaya, 2007; Kovačić et al., 2021), три из которых нативно обитают в Азовcком бассейне. В нижнем течении р. Дон, его крупных притоках и Цимлянском водохранилище широко распространены звёздчатая B. stellatus и недавно описанная донская B. durrelli (Boldyrev, Bogutskaya, 2004; Болдырев и др., 2021) пуголовки.
Для пуголовок характерны дорсовентрально уплощённое в передней части тело, крупная, широкая голова и мелкие размеры. Максимальная стандартная длина тела (SL) большинства видов не превышает 80 мм. Чешуя отсутствует и заменена своеобразными покровными костными образованиями (бугорками и гранулами), форма, размеры, плотность и характер расположения которых на поверхности тела имеют важное таксономическое значение у этой группы рыб (Насека и др., 2012).
Пуголовки демонстрируют репродуктивную стратегию, типичную для всех представителей семейства Gobiidae, включая охрану кладки самцом. Их считают моноциклическими видами с порционным икрометанием, погибающими после нерестового периода на втором году жизни (Miller, 1986; The freshwater fishes …, 2004). В южной части Каспийского моря в силу относительной растянутости вегетационного сезона особи некоторых видов, по-видимому, созревают и нерестятся в осенне-зимний период уже в возрасте полугода (Рагимов, 1985). В местах, где численность пуголовок достигает значительных величин, они играют немаловажную роль в питании хищных видов рыб (Майский, 1955; Болдырев, 2007).
В последние десятилетия понто-каспийские бычки вызывают особый интерес, так как среди видов этой группы есть примеры успешных инвазий (Богуцкая и др., 2004). В некоторых крупных реках Черноморского бассейна с изменением их гидрорежима в результате зарегулирования наблюдается расселение пуголовок (Kottelat, Freyhof, 2007). В волжских водохранилищах и нижнем течении реки, вне границ своего исторического ареала, отмечена донская пуголовка (Boldyrev, Bogutskaya, 2007; Болдырев, 2021).
Биология пуголовок в целом мало изучена, а сведения об их образе жизни в Донском бассейне отсутствуют. Цель работы – охарактеризовать жизненный цикл, размножение, питание, распространение звёздчатой и донской пуголовок.
МАТЕРИАЛ И МЕТОДИКА
Материал собирали в 2001–2021 гг. в Цимлянском водохранилище и в 2003, 2004 гг. на нижнем участке Дона в границах Ростовской области. В водохранилище рыб отлавливали мальковым бимтралом и выбирали из дели промыслового невода на Некрасовской тоне, расположенной в верхней части водоёма – в 20 км выше г. Калач-на-Дону, а также в ходе мальковых (мальковые волокуша и бимтрал) и реже учётных траловых съёмок по всей его акватории (Вехов и др., 2014). Сведения о cкате молоди пуголовок через Волжскую (Волгоградскую) ГЭС получены из отчётов Волгоградского контрольно-наблюдательного пункта Нижневолжского филиала Главного бассейнового управления по рыболовству и сохранению водных биологических ресурсов (Нижневолжрыбвод)11.
В Цимлянском водохранилище основным орудием лова был самодельный мальковый бимтрал с рамой 80 × 42 см. Траления проводили со скоростью 1.6–2.2 км/ч. Глубины, на которых проводили лов, варьировали в пределах 3–20 м. Размер ячеи дели малькового бимтрала – 3 мм, промыслового невода и учётного трала – 30–45 мм. Длина мальковой волокуши 25 м, ячея – 4 мм.
На нижнем участке Дона материал собирали в мае и июле в рейсах, организованных Азово-Черноморским филиалом ВНИРО, из уловов бимтрала (бимтрал-Д), имеющего раскрытие 2.0 × 0.6 м и кутец из хамсороса (ячея 4–6 мм) длиной 6 м, на участке от станицы Мелиховская до устья.
Пуголовок в уловах разбирали по видовому, половому, количественному и размерно-массовому составу. За стандартную длину тела принимали расстояние от вершины рыла до конца гипурального комплекса. В отдельных случаях приводится общая длина (TL). Для сопоставления с данными литературы, в которых приводится только TL, для донской пуголовки использовали соотношение SL = 0.84TL − 1.48 (n = 92 экз., R2 = 0.99), для звёздчатой – SL = 0.83TL − 0.63 (n = 134, R2 = = 0.99). Промерено 1.6 тыс. экз. звёздчатой и 2.3 тыс. экз. донской пуголовок. Объёмы конкретных выборок по участкам водоёмов и по времени сбора указаны в соответствующих таблицах и рисунках. При изготовлении фотографий рыб подкрашивали ализарином.
Кроме того, в работе приведены сведения о пуголовках, собранных авторами в Волжском бассейне в 2001–2015 гг., а также о полученных в 2011 г. от А.Я. Мошу и В.К. Романеску (Институт зоологии АН Молдовы) 16 экз. SL 28–49 мм, выловленных в 2006 г. в низовье р. Днестр на участке реки между с. Устье и г. Криулень.
Обработку данных по плодовитости проводили с помощью методики для порционно нерестящихся рыб (Спановская, Григораш, 1976). Устанавливали сроки нереста, количество порций икры и число икринок в каждой из них. Для анализа динамики созревания половых продуктов самок использовали коэффициент зрелости (КЗ) – отношение массы гонад к массе тела, выраженное в процентах. Для установления числа икринок в очередной созревающей порции у самок с половыми продуктами на III–IV стадии зрелости проводили подсчёт в гонадах крупных желтковых ооцитов диаметром 0.7–1.5 мм при их чётком размерном обособлении от ооцитов диаметром <0.6 мм. Всего подсчитано число икринок в очередных порциях у 88 экз. звёздчатой и 143 экз. донской пуголовок.
Состав пищи исследовали счётно-весовым методом (Методическое пособие …, 1974). В пищевом комке выделяли следующие группы организмов: моллюски (Mollusca), планктонные ракообразные (Cladocera, Copepoda), мизиды (Mysida), гаммариды (Gammaridae), корофииды (Corophiidae), личинки комаров (Chironomidae, Culicidae), черви (Oligochaeta, Polychaeta), рыбы и прочие. Только моллюсков, рыб и некоторых насекомых определяли до рода или вида, остальные организмы – до указанных таксономических категорий. Отдельно учитывали непищевые объекты. Моллюсков, мизид и гаммарид по возможности промеряли. Всего было обработано 574 желудочно-кишечных тракта (ЖКТ) пуголовок обоих видов из Цимлянского водохранилища (район Некрасовской тони (337 экз.), приплотинный участок (9)) и нижнего течения Дона (228). Для характеристики спектра питания использовали такие показатели, как частота встречаемости (F, %), доля каждого компонента пищи по массе (P, %) и общий индекс наполнения ЖКТ (Ind, ‱) – отношение массы всего пищевого комка к общей массе рыбы.
Для сравнения спектров питания двух видов пуголовок использовали индекс пищевого сходства (ИПС) Шорыгина (1952). Индексы избирания (Е) рассчитывали по формуле Ивлева (1955). Для определения степени перекрывания их пищевых ниш рассчитывали индекс Хорна: ${{C}_{\lambda }} = 2\sum {{x}_{i}}{{y}_{i}}{\text{/}}\left( {\sum x_{i}^{2} + \sum y_{i}^{2}} \right)$, где xі и yі – значения отдельных компонентов в пищевых комках рыб, %. Индекс равен нулю при полном различии пищевых ниш и единице – при полном их совпадении. Значение индекса >0.6 расценивали как биологически значимое перекрывание пищевых ниш.
Для характеристики донных ценозов верхнего участка Цимлянского водохранилища в 2009–2015 гг. в пределах нескольких километров выше и ниже Некрасовской тони проводили сбор проб бентоса на глубинах 2–14 м дночерпателем Петерсена с площадью захвата 0.025 м2. Всего обработано 22 пробы. В каждой пробе оценивали качественно-количественный состав важнейших групп беспозвоночных.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Размерно-возрастной и половой состав
Размерный состав звёздчатой и донской пуголовок, собранных в разные сезоны в одних и тех же местообитаниях, существенно различается, что обусловлено различиями в темпе роста и сроках нереста. Донская пуголовка сравнительно небольшая. Максимальные стандартная длина и масса годовалых (1.) самцов в конце весны составляют 66 мм (TL 82 мм) и 9.5 г, самок – 54 мм (TL 66 мм) и 4.6 г. Соотношение длина–масса у этого вида аппроксимируется степенным уравнением: W = 10−5SL3.184 (n = 383, R2 = 0.96). Все производители после нереста погибают. Последних двухлеток (1+) отмечают в августе. С сентября в Цимлянском водохранилище присутствуют только сеголетки (0+) (рис. 1), хотя, судя по размерному составу пуголовок в севернее расположенных водоёмах новоприобретённого ареала, каким является Куйбышевское водохранилище (Кудерский и др., 1995), двухлетних производителей в значительном количестве отмечают и в сентябре, видимо, из-за сдвига сроков нереста. Как и у других представителей азово-черноморских бычков (Калинина, 1976) самцы в росте опережают самок (рис. 2). Зависимость длины от возраста (в сутках) донской пуголовки (возраст 0+, 1.) описывается уравнением: для самцов – SL = 14.302 ln(x) − − 40.881 (n = 679, R2 = 0.98), для самок – SL = = 11.538 ln(x) − 29.026 (n = 895, R2 = 0.99).
Рис. 1.
Размерный состав (стандартная длина SL) звёздчатой Benthophilus stellatus (а–е) и донской B. durrelli (ж–м) пуголовок из уловов различными орудиями лова в Цимлянском водохранилище и нижнем
течении р. Дон в разные месяцы 2002, 2003, 2013, 2015 и 2021 гг.: ( ) – ювенильные особи, (◼) – самцы, (◻) – самки. Некрасовская тоня Цимлянского водохранилища,
бимтрал: а, ж – апрель, n (соответственно) = 349 и 110 экз.; д – сентябрь, n = 60; и – июль, n = 292; л, м – соответственно сентябрь, n = 169 и декабрь, n = 131. Нижний Дон, бимтрал-Д: б, з – май, n = 48 и 50. Водохранилище: в, г – бимтрал, соответственно июль, n = 81 и август, n = 334; е – трал, декабрь, n = 17; к – волокуша, июль, n = 73. Стрелками указаны малозаметные на графике доли рыб.
) – ювенильные особи, (◼) – самцы, (◻) – самки. Некрасовская тоня Цимлянского водохранилища,
бимтрал: а, ж – апрель, n (соответственно) = 349 и 110 экз.; д – сентябрь, n = 60; и – июль, n = 292; л, м – соответственно сентябрь, n = 169 и декабрь, n = 131. Нижний Дон, бимтрал-Д: б, з – май, n = 48 и 50. Водохранилище: в, г – бимтрал, соответственно июль, n = 81 и август, n = 334; е – трал, декабрь, n = 17; к – волокуша, июль, n = 73. Стрелками указаны малозаметные на графике доли рыб.
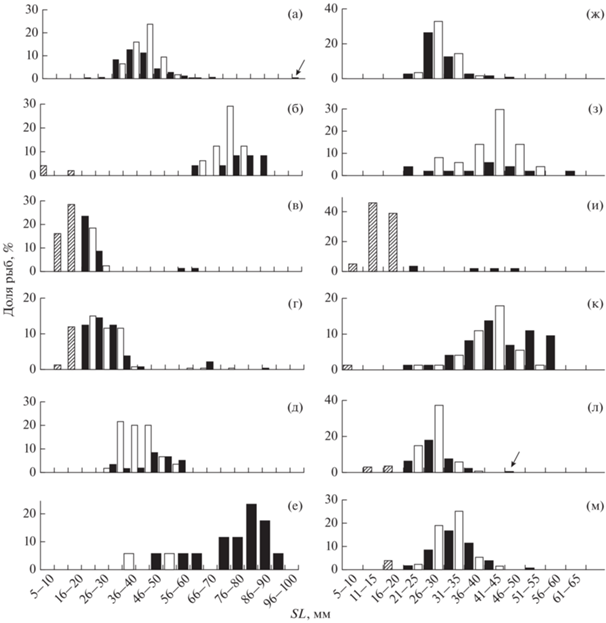
Рис. 2.
Линейный рост (стандартная длина SL) донской пуголовки Benthophilus durrelli (возраст 0+, 1.) верхнего участка Цимлянского водохранилища. Выборки: 1 – апрель 2003 г. (самцы/самки – 30/28 экз.); 2–8: 2013 г.: 2 – май (53/97), 3 – июль (ювенильные особи – 288 экз.), 4 – август (33), 5 – сентябрь (67/109 экз.), 6 – октябрь (59/109), 7 – ноябрь (96/134), 8 – декабрь (53/97). (—) – ювенильные особи, (– − –) – самцы, (― · ―×) – самки; линии соединяют средние значения, ( ) – пределы варьирования показателя.
) – пределы варьирования показателя.

Звёздчатая пуголовка значительно крупнее донской. Длина и масса самых больших самцов, отмеченных в ранневесенний период наблюдений, составляли 98 мм (TL 117 мм) и 32 г, самок – 83 мм (TL 104 мм) и 17 г. Соотношение длина–масса аппроксимируется уравнением: W = 2 × 10−5SL3.081, (n = 339, R2 = 0.98). Максимальные размеры самцов (TL 135 мм) и самок (TL 110 мм) этого вида, указываемые для Азовского моря, незначительно больше. Ранее считали, что все производители погибают после нереста на втором году жизни (Ильин, 1927а, 1927б). В водохранилище основная часть производителей погибает годовиками, но в водоёме отмечают отдельных особей двухлеток и двухгодовиков (2.). Так, вместе с сеголетками (рис. 3а), пойманными в конце декабря 2015 г., отмечены, судя по чёткому зимнему кольцу на отолитах, и двухлетние самцы SL 83–94 мм (рис. 3б), а редкие экземпляры SL > 90 мм, отловленные весной, являлись двухгодовиками (рис. 1). Особи обоих полов старше двухлетнего возраста отмечены и у пуголовки Берга B. leobergius в южной части Каспийского моря (Patimar et al., 2019). Максимальная SL этих видов достигает ~110 мм, что значительно больше, чем у большинства представителей рода (~33–80 мм) (Boldyrev, Bogutskaya, 2007). Самцы опережают самок в росте (рис. 4). Размерно-возрастная зависимость у сеголеток–годовиков описывается уравнением: для самцов – SL = = −0.0011x2 + 0.5672x − 2.8816 (n = 379, R2 = 0.99), для самок – y = −0.0007x2 + 0.3804x + 2.8377 (n = = 442, R2 = 0.99).
Рис. 3.
Отолиты сеголетки SL 79 мм (а) и двухлетки SL 83 мм (б) звёздчатой пуголовки Benthophilus stellatus (декабрь 2015 г.), (→) зимнее кольцо.
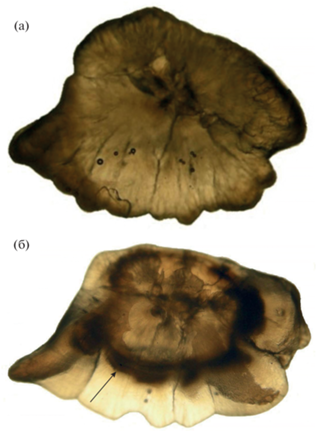
Рис. 4.
Линейный рост (стандартная длина SL) звёздчатой пуголовки Benthophilus stellatus (возраст 0+, 1.) Цимлянского водохранилища. Выборки: 1–3 (Некрасовская тоня): 1 – апрель 2003 г. (самцы/самки – 34/55 экз.), 2 – июнь 2001 г. (ювенильные особи – 93 экз.), 3 – июль 2001 г. (22 экз.); 4–5 (все плёсы водохранилища): 4 – июль 2015 г. (79 экз.), 5 – август 2013 г. (самцы/самки – 116/109 экз.); 6–8 (Некрасовская тоня): 6 – август 2013 г. (16/30 экз.), 7 – сентябрь 2013 г. (16/44 экз.), 8 – ноябрь 2013 г. (3/10 экз.). Обозначения см. на рис. 2.

Среди неполовозрелых пуголовок обоих видов доля самок выше, чем самцов (рис. 5). Это, видимо, обусловлено тем, что в период размножения в гнёзда, охраняемые самцами, как это известно для других видов понто-каспийских бычков, могут откладывать икру несколько самок. С началом нереста гибель самок опережает гибель самцов.
Размножение
Показателем порционности икрометания обоих видов пуголовок является наличие в гонадах самок разноразмерных ооцитов прото- и трофоплазматического роста в преднерестовый и в разгар нерестового периодов. В яичниках IV стадии зрелости диаметр желтковых ооцитов у обоих видов на конечной фазе развития составляет 1.3–1.6 мм. Икра пуголовок в период эмбрионального развития, как и у других бычков, имеет яйцевидную форму, которую она приобретает на последнем этапе созревания, ещё находясь в целомической жидкости (рис. 6).
Рис. 6.
Ооциты разных стадий развития в гонадах звёздчатой пуголовки Benthophilus stellatus перед откладыванием очередной порции икры. Масштаб: 1 мм.

У донской пуголовки КЗ в апреле низкий и не превышает значения 5.8 (рис. 7а). Нереститься, судя по поимкам самок с высокими показателями КЗ (до 24.6), вид начинает в первой половине мая, когда вода в водохранилище прогревается до 15–17°С. Продолжительность нерестового периода составляет ~3.0–3.5 мес. и обусловлена разновременностью созревания и икрометания самок различных размерных групп. Минимальная длина годовиков в середине апреля составляет 16 мм, в середине мая – 25 мм. Самки же начинают нереститься при достижении SL ~ 35 мм. С мая по июль в уловах всегда регистрируют особей с низкими значениями КЗ (0.1–5.0), по-видимому, ещё не участвовавших в нересте, а в августе – уже с КЗ только >5.0. Молодь первых дней жизни SL 6–9 мм отмечают в водоёмах до конца августа–начала сентября. Сеголетки от раннего нереста в середине июля в водохранилище уже достигают SL 22 мм, а в нижнем течении Дона – 28 мм. Самая мелкая молодь, учтённая в конце сентября, имеет SL 11 мм. В сентябре производителей в наших уловах уже не отмечали (рис. 1).
Рис. 7.
Изменение коэффициента зрелости самок донской Benthophilus durrelli (а) и звёздчатой B. stellatus (б) пуголовок в Цимлянском водохранилище и нижнем течении р. Дон. B. durrelli: водохранилище: 1 – март 2001 г. (38 экз.), 3 – май 2013 г. (26), 4 – июнь 2013 г. (28), 6 – июнь 2021 г. (26), 7 – август 2013 г. (33); р. Дон (2004 г.): 2 – май (12), 5 – июнь (14). B. stellatus: водохранилище: 1 – март 2014 г. (24 экз.), 2 – апрель 2001 г. (15), 3 – апрель 2003 г. (12), 6 – май 2013 г. (8), 8 – август 2013 г. (1); р. Дон: 4 – май 2003 г. (34), 5 – май 2004 г. (20), 7 – июль 2013 г. (3). Каждый бокс включает 5 горизонтальных линий, которые обозначают 10, 25, 50 (медиана), 75 и 90% данных (от 25 до 75% данных заключены в прямоугольник); (⚪) – значения за пределами 10 и 90% данных.
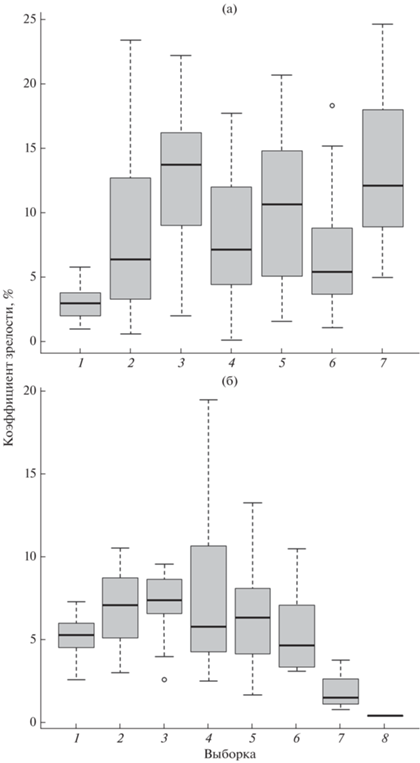
Продолжительность нерестового периода у звёздчатой пуголовки по сравнению с донской невелика – ~1.0–1.5 мес. Нереститься вид начинает с конца апреля при прогреве воды до 13–14°С. Бóльшая часть годовиков к началу сезона размножения уже достигает половозрелости (рис. 7б). Самки начинают нереститься при достижении SL ~ 50 мм. В марте и первой половине апреля значение КЗ у них не превышает 10. С конца апреля и весь май, когда продолжается нерест, отмечают особей с КЗ до 20. В июне происходит массовая посленерестовая гибель производителей, но в июле–августе в уловах единично продолжают регистрировать крупных самок с КЗ 0.4–4.0. Позже самок-двухлеток, в отличии от самцов, уже не отмечают (рис. 1). В конце мая 2003 г. на нижнем участке Дона учитывали молодь SL 7–19 мм. В середине июня 2001 г. на Некрасовской тоне её SL варьировала в пределах 5–14 мм, а в начале июля – 10–16 мм. В сборах по Цимлянскому водохранилищу в середине июля 2015 г. присутствовала молодь уже SL 12–29 мм, в начале августа 2013 г. – 15–42 мм.
У рыб с асинхронным типом вителлогенеза и многопорционным икрометанием установление индивидуальной абсолютной плодовитости, т.е. числа зрелых икринок, вымётываемых одной самкой за один нерестовый сезон, расчётным методом затруднено, так как в её формировании в течение всего нерестового периода могут принимать участие резервные ооциты. Поэтому оценка числа выметанных в текущем сезоне икринок разовым подсчётом желтковых ооцитов может приводить к заниженному результату (Овен, 1976), особенно если количество откладываемых порций превышает две–три. С другой стороны, известно, что на III стадии зрелости гонад формируется потенциальная плодовитость рыб, когда число половых клеток, вступивших в стадию трофоплазматического роста, в гонадах максимально (Иванков, 2001). Впоследствии, по мере созревания половых продуктов и расхода питательных веществ, часть ооцитов резорбируются. Большинство работ, содержащих данные по плодовитости понто-каспийских бычков, не учитывают эти факторы (Богуцкая и др., 2004). Имеющиеся данные по индивидуальной абсолютной плодовитости представителей рода Benthophilus получены либо подсчётом крупных желтковых ооцитов, размерно изолированных от сравнительно мелких, что в реальности соответствует количеству икринок в очередной порции (Ильин, 1927а; Трифонов, 1955; Гавлена, 1973), либо суммированием первых и сравнительно мелких ооцитов без обоснования границы разрыва с резервными (Рагимов, 1985). Часто в работах, посвящённых установлению плодовитости бычков, предположительное число порций икры выводят из полимодальности размерного ряда икринок в гонадах самок в преднерестовый период. Такой подход почти всегда приводит к выводу о 2–3-порционном нересте (Калинина, 1976), что не всегда соответствует действительности. Так у кругляка, наиболее хорошо изученного вида понто-каспийских бычков, число откладываемых порций икры, как показали результаты экспериментов в природных и лабораторных условиях, может доходить до пяти–шести в зависимости от продолжительности нерестового периода, обусловленного температурным режимом водоёма и обеспеченностью пищей самок. Порции откладываются с интервалом, по одним исследованиям, 17–20 сут (Ращеперин, 1964), по другим – 14–28 (Куликова, Фандеева, 1975). Важным выводом является значительное снижение числа икринок во второй порции по сравнению с первой и незначительное у каждой последующей по сравнению с предыдущей. Такая закономерность характерна и для некоторых видов рыб из других таксономических групп, вымётывающих икру несколькими порциями (Биология …, 1970; Решетников и др., 2016).
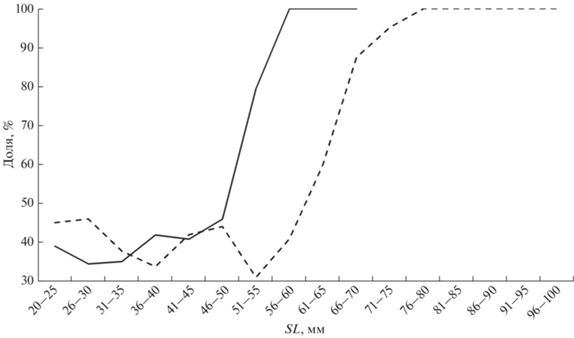
Распределение числа желтковых ооцитов старшей генерации у самок донских пуголовок (рис. 8), отловленных в разные месяцы и годы, позволяет предположить, что основная часть рыб этого вида является единовременно нерестующими. На рис. 8а выделены две совокупности рыб по распределению числа ооцитов старшей генерации. В первую, помимо всех майских рыб, собранных в разные годы в преднерестовый период и содержащих поэтому только икринки первой порции, попадает и бóльшая часть особей (74%), пойманных в июне–августе. Причём в августовской выборке таких 87%. Амплитуда варьирования числа икринок в каждом размерном классе рыб с шагом 1 мм составляет ~200 шт. Вторая совокупность значительно меньше по объёму и, по-видимому, отображает число икринок во второй порции. В силу сравнительно небольшого объёма выборки их число в каждом размерном классе варьирует значительно меньше, чем в первой порции, – в пределах 50–100 ооцитов. Из всего этого следует, что бо́льшая часть пуголовок погибает после откладывания первой порции икры, число ооцитов в которой увеличивается с ростом рыб и описывается линейным уравнением: y = 23.387SL − 604.88 (n = 118, R2 = 0.71). Относительно низкая плодовитость компенсируется растянутостью нерестового сезона и, соответственно, снижением рисков гибели ранней молоди от неблагоприятных факторов в условиях нестабильности среды обитания.
Рис. 8.
Число желтковых ооцитов старшей генерации у донской Benthophilus durrelli (а) и звёздчатой B. stellatus (б) пуголовок в Цимлянском водохранилище и нижнем течении р. Дон в зависимости от
стандартной длины тела (SL). B. durrelli: водохранилище: (+) – май 2013 г., ( ) – июнь 2013 г., ($ \times |$) – июль 2021 г., (⚫) – август 2013 г.; р. Дон: (◇) – май 2003 г., (△) – май 2004
г., (×) – июль 2004 г., (◻) – июль 2003 г. B. stellatus: водохранилище: (+) – март 2014 г., (△) – апрель 2001 г., (×) – апрель 2003 г., (⚪)
– апрель 2002 г., (⚫) – май 2013 г.; р. Дон: (◼) – май 2004 г., (◆) – май 2003 г.,
(▲) – июль 2004 г.; (---) – условная граница между первой и последующими генерациями ооцитов.
) – июнь 2013 г., ($ \times |$) – июль 2021 г., (⚫) – август 2013 г.; р. Дон: (◇) – май 2003 г., (△) – май 2004
г., (×) – июль 2004 г., (◻) – июль 2003 г. B. stellatus: водохранилище: (+) – март 2014 г., (△) – апрель 2001 г., (×) – апрель 2003 г., (⚪)
– апрель 2002 г., (⚫) – май 2013 г.; р. Дон: (◼) – май 2004 г., (◆) – май 2003 г.,
(▲) – июль 2004 г.; (---) – условная граница между первой и последующими генерациями ооцитов.
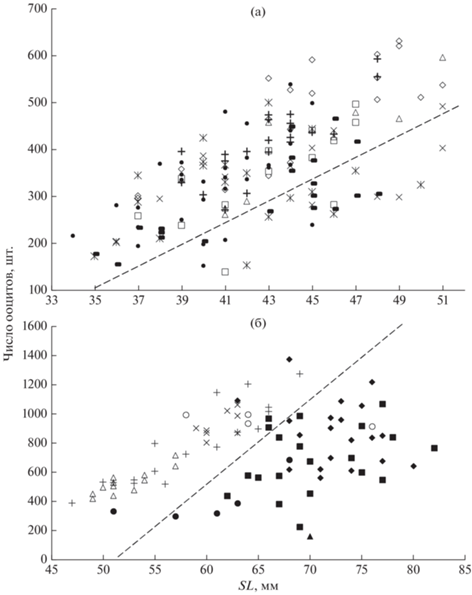
У звёздчатой пуголовки распределение числа зрелых ооцитов старшей генерации в разных выборках иное (рис. 8б). У экземпляров, собранных в марте и апреле, уже хорошо дифференцирована первая порция икринок. В эту совокупность попадают и отдельные особи, собранные в начале мая. Амплитуда варьирования числа икринок в каждом размерном классе с шагом 1 мм составляет ~200–300 шт. Основная часть майских экземпляров и одного июльского образует сравнительно обособленную совокупность, варьирование количества ооцитов на единицу длины в которой доходит до 600. Такая значительная амплитуда вкупе с тем, что продолжительность нереста этого вида длится ~1.0–1.5 мес., а созревание очередной порции икры у пуголовок составляет по некоторым оценкам 15–20 сут (Макеева и др., 2011), позволяют предположить, что основная часть самок звёздчатой пуголовки откладывает за сезон три порции икры. Зависимость числа ооцитов от длины рыб описывается уравнением: в первой порции – y = = 39.649SL − 1528.6 (n = 45, R2 = 0.84), во второй и третьей – y = 24.23SL − 1028.4 (n = 43, R2 = 0.27).
Питание
Основу питания обоих видов в верхнем участке Цимлянского водохранилища у особей SL 20–55 мм составляют личинки комаров и мелкие брюхоногие моллюски Lithoglyphus naticoides (табл. 1, рис. 9), в меньшей степени – черви и ракообразные. Среди последних преобладают мизиды. Остальные группы кормовых организмов в питании обоих видов играют незначительную роль. Из насекомых помимо личинок комаров в желудках встречаются подёнки (Ephemera sp.), личинки равнокрылых стрекоз (Zygoptera) и жуков (Coleoptera), водяные клопы (Corixidae) и водяные клещи (Hydrachnidia). Среди низших ракообразных преобладают ветвистоусые рачки, из рыб – молодь бычков родов Babka, Neogobius и самих пуголовок. Единично отмечаются мелкие шаровки (Sphaerium sp.) и пиявки (Hemiclepsis marginata).
Таблица 1.
Состав пищи и другие показатели звёздчатой Benthophilus stellatus и донской B. durrelli пуголовок Цимлянского водохранилища и нижнего течения р. Дон
| Компонент пищи и другие показатели | Верхний участок водохранилища | Нижнее течение Дона | ||||||
|---|---|---|---|---|---|---|---|---|
| B. stellatus | B. durrelli | B. stellatus | B. durrelli | |||||
| P | F | P | F | P | F | P | F | |
| Моллюски | 30.7 | 35.7 | 33.2 | 33.3 | 5.9 | 13.9 | 2.8 | 2.8 |
| Черви | 2.6 | 22.9 | 9.8 | 47.5 | 17.2 | 32.2 | 6.5 | 29.0 |
| Планктонные ракообразные | 0.4 | 7.0 | 0.4 | 19.2 | 0.2 | 5.5 | 0.3 | 6.2 |
| Корофииды | 1.9 | 11.5 | 2.2 | 35.0 | 16.4 | 76.5 | 22.2 | 60.9 |
| Гаммариды | 1.0 | 4.5 | 0.5 | 0.8 | 17.0 | 48.7 | 53.3 | 47.8 |
| Мизиды | 15.2 | 23.6 | 5.1 | 15.0 | 28.1 | 33.0 | 7.9 | 13.0 |
| Личинки комаров | 31.8 | 81.5 | 40.6 | 75.0 | 0.2 | 13.0 | 0.4 | 8.7 |
| Рыбы | 7.6 | 4.5 | 0.1 | 0.8 | 1.2 | 13.0 | 0.6 | 4.3 |
| Прочие | 2.1 | 20.4 | 3.5 | 10.8 | 0.8 | 5.2 | 1.0 | 7.2 |
| Грунт, слизь | 6.7 | 20.4 | 4.6 | 50.0 | 13.0 | 38.3 | 5.0 | 31.9 |
| SL, мм* | $\frac{{20.1{\kern 1pt} --{\kern 1pt} 87.0}}{{45.9}}$ | $\frac{{20.0{\kern 1pt} --{\kern 1pt} 61.6}}{{38.6}}$ | $\frac{{20.3{\kern 1pt} --{\kern 1pt} 89.5}}{{53.3}}$ | $\frac{{20.3{\kern 1pt} --{\kern 1pt} 61.2}}{{39.7}}$ | ||||
| Масса, г* | $\frac{{0.2{\kern 1pt} --{\kern 1pt} 23.7}}{{3.6}}$ | $\frac{{0.2{\kern 1pt} --{\kern 1pt} 8.0}}{{1.8}}$ | $\frac{{0.2{\kern 1pt} --{\kern 1pt} 24.9}}{{6.3}}$ | $\frac{{0.2{\kern 1pt} --{\kern 1pt} 7.5}}{{2.3}}$ | ||||
| Общий индекс наполнения ЖКТ, ‱ | $\frac{{1.1{\kern 1pt} --{\kern 1pt} 1290.3}}{{316.6}}$ | $\frac{{6.5{\kern 1pt} --{\kern 1pt} 1444.4}}{{344.4}}$ | $\frac{{3.6{\kern 1pt} --{\kern 1pt} 675.9}}{{256.1}}$ | $\frac{{8.1{\kern 1pt} --{\kern 1pt} 718.8}}{{244.1}}$ | ||||
| Число ЖКТ/из них пустые | 193/0 | 144/5 | 140/0 | 88/0 | ||||
Рис. 9.
Спектр питания звёздчатой Benthophilus stellatus (а) и донской B. durrelli (б) пуголовок в зависимости от их стандартной длины (SL) в верхней части Цимлянского водохранилища: 1 – моллюски, 2 – гаммариды, 3 – мизиды, 4 – корофииды, 5 – личинки комаров, 6 – рыбы, 7 – черви, 8 – прочие, 9 – грунт.
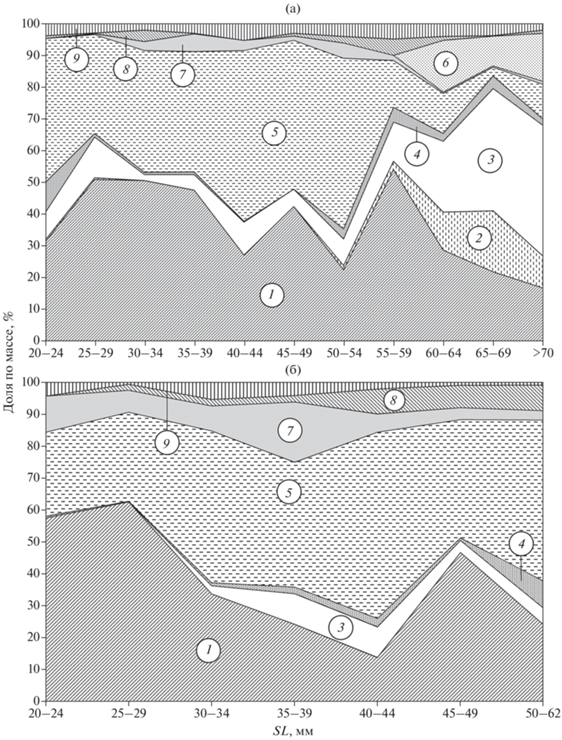
У крупных особей звёздчатой пуголовки SL > > 55 мм по сравнению с донской в питании наблюдается снижение доли моллюсков и личинок комаров за счёт увеличения потребления относительно крупных объектов, таких как мизиды, гаммариды и молодь рыб (рис. 9). О пластичности в отношении объектов питания свидетельствует тот факт, что почти у всех звёздчатых пуголовок в небольшой выборке из приплотинного участка (SL 58–85 мм, 9 экз.), в отличие от особей из верхней части водохранилища, в пищевом комке совсем отсутствовали моллюски и в значительном количестве отмечены кумовые рачки (Cumacea) (P = 21%, F = 67%).
Питание пуголовок нижнего течения Дона и водохранилища существенно различается (табл. 1). В реке основными потребляемыми организмами являются ракообразные и в меньшей степени черви. Роль личинок комаров и моллюсков незначительна. Моллюски L. naticoides в пищевом комке отсутствуют, чаще встречаются живородки (Viviparus viviparus), реже – мелкие дрейссены (Dreissena sp.), прудовики (Lymnaea sp.) и лунки (Theodoxus sp.). Единично отмечены личинки ручейников (Trichoptera), среди рыб – молодь бычков родов Babka, Neogobius и самих пуголовок. Случайной пищей являются редкие фрагменты и семена растений. Грунт, видимо, заглатывается вместе с бентическими организмами и отмечается довольно редко, обычно вместе с червями. В питании звёздчатых пуголовок в сравнении с донскими бо́льшую роль играют мизиды, меньшую – гаммариды.
Состав пищи самок и самцов пуголовок сходен. Обращает на себя внимание, что в весенней выборке 2003 г. из 12 экз. звёздчатых пуголовок, в пищевом комке которых имелись сеголетки бычковых, представители родов Babka и Neogobius отмечались исключительно у самок (9 экз.), а молодь пуголовок – у крупных самцов (3 экз.). Для представителей бычковых характерно то, что их самцы в период эмбрионального развития икры остаются на кладке, охраняя, очищая и аэрируя её. Отмечаемый каннибализм, видимо, связан с сопряжённой повышенной концентрацией именно самцов с их ранней молодью на нерестилищах в мае.
В целом различия в питании двух видов пуголовок незначительны. Их пищевое сходство в нижнем течении Дона можно охарактеризовать как высокое (ИПС равен 52.4, Cλ = 0.63), а в Цимлянском водохранилище как очень высокое (75.2, 0.94). Сходство отмечено у обоих видов и в синхронной избирательности кормовых объектов. В водохранилище высокие значения Е имеют L. naticoides (у звёздчатой – 0.67, у донской – 0.64) и мизиды (0.76, 0.35). Оба вида при необходимости легко переключаются на доступные кормовые объекты. Отмеченные различия обусловлены не видовой специализацией, а относительно мелкими размерами донской пуголовки по сравнению со звёздчатой. В питании последней с ростом возрастает значение таких сравнительно крупных объектов, как мизиды, гаммариды и молодь рыб (рис. 9а). Кроме того, пуголовки с ростом проявляют размерную избирательность по некоторым объектам питания (рис. 10).
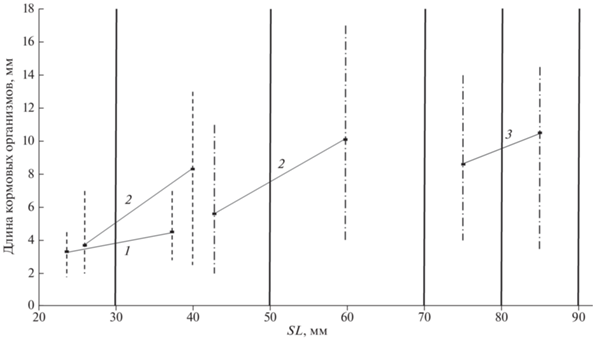
Рис. 10.
Размерный состав кормовых организмов в питании пуголовок Цимлянского водохранилища
в зависимости от их стандартной длины (SL): средние значения ( ) и пределы варьирования показателя у донской Benthophilus durrelli (
) и пределы варьирования показателя у донской Benthophilus durrelli ( ) и звёздчатой B.stellatus (
) и звёздчатой B.stellatus ( ) пуголовок, (
) пуголовок, ( ) – границы размерных групп рыб, 1 – Lithoglyphus naticoides (55 экз.), 2 – гаммариды (158 экз.), 3 – мизиды (88 экз.).
) – границы размерных групп рыб, 1 – Lithoglyphus naticoides (55 экз.), 2 – гаммариды (158 экз.), 3 – мизиды (88 экз.).
Полученные данные демонстрируют, что для обоих видов пуголовок характерны широкий спектр питания и способность переориентироваться в разных биотопах на самые различные кормовые объекты. Это обусловлено их неспособностью в силу мелких размеров и малой двигательной активности к протяжённым миграциям в поисках корма. Подтверждением пластичности пуголовок служат данные по изменчивости питания этих рыб в зависимости от характера грунтов и структуры донных ценозов только в пределах Таганрогского залива в 1920-е гг. (Киналев, 1937). Так, на песках его восточной части основу рациона пуголовок составляли полихеты рода Nereis, в средней части – личинки Chironomidae, на илах – молодь двустворчатого моллюска рода Abra и тому подобное. В целом в начале прошлого столетия в восточной части Азовского моря помимо указанных объектов рацион пуголовок состоял из корофиид, молоди брюхоногих (Hydrobia) и двустворчатых (Cerastoderma) моллюсков (Ильин, 1927а), а в 1950-е гг. в Таганрогском заливе – преимущественно из представителей родов Cerastoderma и Nereis (Желтенкова, 1964). Следствием участившихся во второй половине ХХ в. заморов на Азовском море (Александрова и др., 1998) главным образом стало существенное сокращение площадей биоценозов, включающих оксифильных представителей рода Cerastoderma, молодь которых до этого играла важнейшую роль в питании пуголовок и некоторых других видов бычков (Костюченко, 1960), и увеличение численности и биомассы замороустойчивых видов, среди которых были и Hydrobia sp. (Студеникина и др., 1998). Именно эти моллюски стали позже основным компонентом в рационе взрослых пуголовок в Азовском море (Рейх, 1976).
Для черноморских пуголовок также показательна зависимость между характером биотопа и преобладанием в рационе тех или иных кормовых объектов. Чаще у взрослых рыб это ракообразные (бокоплавы, мизиды), мелкие свободноживущие брюхоногие и двустворчатые моллюски, личинки Chironomidae и молодь рыб (Ульман, 1970; Смирнов, 1986; Sindilariu, Freyhof, 2003).
В 1950–1960-е гг. пуголовок случайно вселили в волжские водохранилища. В этих водоёмах тоже отмечают сезонные и локальные изменения в их питании. На разных участках Чебоксарского, Куйбышевского и Саратовского водохранилищ ведущими кормовыми объектами у пуголовок могут являться личинки Chironomidae, бокоплавы, брюхоногие моллюски (чаще L. naticoides) или молодь двустворчатых моллюсков (чаще дрейссена) (Гавлена, 1973; Ермолин, 1980; Кудерский и др., 1995; Касьянов, Клевакин, 2011; Семенов, 2011; Шемонаев, Кириленко, 2011).
Обращает на себя внимание то, что в отдельных локальностях в волжских водоёмах довольно значительную долю в питании пуголовок составляет молодь дрейссены. В то же время в нашей выборке из верхнего участка Цимлянского водохранилища, где количественные показатели численности (205.5 экз/м2) и биомассы (311.1 г/м2) этого двустворчатого моллюска достаточно высокие, он в их питании отсутствует. Представители семейства Dreissenidae имеют свободноживущую личиночную стадию (велигер), а для взрослых особей характерен прикреплённый образ жизни. Поселяться они могут на любых твёрдых предметах, при оседании же на ил велигеры погибают. Среди русловых донных биотопов в местах отлова пуголовок каменистые и песчаные россыпи отсутствуют, а преобладают сильно заиленные пески с высокой численностью и биомассой моллюсков (соответственно 510.9 экз/м2 и 2298.5 г/м2), и субстратом для закрепления дрейссены служат раковины крупных представителей родов Anodonta и Unio. Таким образом, для дрейссенников на этом участке характерна высокая степень агрегированности. Отсутствие представителей рода Dreissena в питании пуголовок объясняется тем, что последние имеют слабый челюстной аппарат и не способны в отличие от некоторых других представителей понтокаспийских бычковых (Богуцкая и др., 2004) отрывать отдельных моллюсков от друз. Довольно значительная роль молоди дрейссены, как компонента питания пуголовок в волжских водохранилищах, объясняется, по-видимому, тем, что они достаточно эффективно собирают отдельных моллюсков, разреженно оседающих и закрепляющихся на песчаном субстрате.
Как было показано, одним из основных компонентов питания у пуголовок в Цимлянском водохранилище является L. naticoides. Для этих свободноживущих моллюсков характерно отсутствие унгулятной крышечки, закрывающей устье раковины, в ротовой полости пуголовки их не дробят, и в пищеварительный тракт они поступают в целом состоянии. Лишь в заднем отделе кишечника раковины настолько истончаются в процессе пищеварения, что отдельные из них начинают распадаться (Богачик, 1967; наши данные). В пищевом комке отмечали экземпляры высотой 2–7 мм (рис. 10). Численность и биомасса этого моллюска составляют соответственно 81.8 экз/м2 и 1.7 г/м2. Высокая степень избирательности по отношению к этому виду и отсутствие в пищевом комке других брюхоногих моллюсков такого же размерного диапазона из родов Viviparus (83.9 экз/м2, 5.9 г/м2) и Theodoxus (20.0 экз/м2, 1.0 г/м2), по-видимому, объясняется наличием у них устьевой крышечки и более толстой раковины, усложняющих их переваривание.
Распространение пуголовок в пресноводных водоёмах
В нижнем течении Дона с отдельными притоками, а также в Цимлянском водохранилище с первых лет существования водоёма, образованного в результате зарегулирования реки в 1952 г. в 309 км от устья, распространены звёздчатая и донская пуголовки (Болдырев, 2002; Болдырев и др., 2021), одними из основных диагностических признаков для идентификации которых являются форма, размеры, плотность и характер расположения покровных костных образований (Kottelat, Freyhof, 2007; Насека и др., 2012) (рис. 11а, 11б). В крупнейшем притоке – р. Северский Донец – в 1920-х гг. представителей рода Benthophilus отмечали у г. Святогорск на расстоянии до 540 км от Дона и ~ 700 км от его устья (Солодовников, 1930). Судя по размерному составу сеголеток на конец июля–начало августа 1927 г. (7–16 мм (16 экз.)) (Шандиков, Гончаров, 2008), это молодь донской пуголовки. Выше Цимлянского водохранилища пуголовок не отмечали. Указание на их присутствие в конце XIX в. в Дону у Воронежа (~1400 км от устья), видимо, ошибочно (Ильин, 1927б; Boldyrev, Bogutskaya, 2007). Только в устьевой части реки встречается азовская пуголовка B. magistri (наши данные).
Рис. 11.
Покровные костные образования на верхней поверхности головы и передней части спины пуголовок из Цимлянского водохранилища, август 2012 г. (а, б) и авандельты р. Волга, сентябрь 1926 г. (в): а, б – соответственно Benthophilus stellatus SL 45 мм и B. durrelli SL 48 мм; в – B. mahmudbejovi SL 47 мм.

В Черноморском бассейне обитает близкая к звёздчатой голая пуголовка B. nudus. Это, по-видимому, самый крупный вид рода Benthophilus. Его максимальная TL составляет 150 мм (Дренски, 1951), а ареал, судя по большим размерам указываемых особей (Bănărescu, 1964; Мовчан и др., 2003; наши данные), включает в себя бассейны рек Днепр, Днестр, Дунай и Южный Буг. По крайней мере, в первых двух реках, как показывают последние исследования, видимо, встречается и донская пуголовка (Boldyrev, Bogutskaya, 2007; Романеску, 2015).
В Волге до её зарегулирования каскадом плотин пуголовок Махмудбекова B. mahmudbejovi (рис. 11в), зернистую B. granulosus, Абдурахманова B. abdurahmanovi, каспийскую B. macrocephalus и Берга B. leobergius отмечали только в дельте, преимущественно в её нижней части и авандельте (Коблицкая, 1966; Boldyrev, Bogutskaya, 2007; Насека и др., 2012). Выше других, по наблюдениям 1970-х гг., встречали пуголовок зернистую и Махмудбекова (Рагимов, 1981). Указание для нижнего течения реки пуголовки Световидова B. svetovidovi (Васильева, 1983) требует подтверждения.
В 1957–1965 гг. в волжские водохранилища (Рыбинское, Куйбышевское, Кутулукское и Волгоградское) для улучшения кормовой базы рыб заселяли беспозвоночных, перевозимых из дельты Дона, Цимлянского водохранилища и дельты Волги (Иоффе, 1968). В ходе проведения этих мероприятий в ряд водоёмов были вселены пуголовки. В 1970-е гг. их стали отмечать в Куйбышевском, Саратовском и Волгоградском водохранилищах (Гавлена, 1973; Ермолин, 1984; Шашуловский, Ермолин, 2005). Рост численности пуголовок в этих водоёмах, по-видимому, был весьма интенсивным, так как в эти же годы фиксировали скат их молоди через Волгоградскую плотину22. Уже в 1972–1973 гг. пуголовки были отмечены на незарегулированном участке реки у Волгограда (Рагимов, 1981). К настоящему времени они расселились вверх по Волге до Рыбинского водохранилища (Кодухова и др., 2016), встречаются и в таких отдельных крупных притоках, как реки Ока, Кама и Сура (Иванчев, Иванчева, 2010; Шакирова и др., 2015; личное сообщение О.А. Полумордвинова, Пензенский государственный педагогический университет).
Видовой состав рода Benthophilus в волжских водохранилищах требует уточнения. Чаще пуголовок в этих водоёмах формально называют звёздчатыми. К концу прошлого века, как показали недавние исследования (Boldyrev, Bogutskaya, 2007), под этим названием фигурировали четыре вида – нативные в бассейне Азовского моря звёздчатая и донская, Чёрного моря – голая и Каспийского – Берга. Несколько экземпляров из Волгоградского и Саратовского водохранилищ, изученные в ходе ревизии рода Benthophilus, были идентифицированы как B. durrelli, они, очевидно, попали сюда из Донского бассейна с акклиматизируемыми беспозвоночными. Стандартная длина более 1 тыс. промеренных из этих водоёмов особей пуголовок (Гавлена, 1973; Кудерский и др., 1995; Касьянов, Клевакин, 2011; Семенов, 2011; Шемонаев, Кириленко, 2011; Шакирова и др., 2015; Кодухова и др., 2016; личные сообщения В.П. Иванчева, Окский государственный заповедник и Д.П. Карабанова, Институт биологии внутренних вод), не превышала максимального значения для этого вида в Донском бассейне – 66 мм. Указание на самок SL 42–47 мм (Гавлена, 1973) и 33–48 мм (Кудерский и др., 1995) со зрелыми половыми продуктами в Куйбышевском водохранилище исключает возможность того, что это особи звёздчатой пуголовки в силу их мелких размеров, что давало основания предполагать: в волжских водохранилищах обитает только донская пуголовка. Кроме того, представители именно этого вида были позже отмечены в питании налима из нижнего бьефа Волгоградской ГЭС (Болдырев, 2021). Личинка пуголовки SL 5.5 мм, пойманная нами икорной сетью в Волге в 40 км ниже Волгоградской плотины 10 июля 2015 г. при температуре воды 23°С, судя по мелким размерам и, следовательно, относительно позднему нересту, тоже, видимо, принадлежала к B. durrelli. Подтверждением присутствия донской пуголовки в различных волжских водохранилищах является также частая встречаемость в них особей с четырьмя лучами в первом спинном плавнике (табл. 2). Соотношение особей с тремя и четырьмя лучами у этого вида в нативном ареале близко к равному (116 : 115), у звёздчатой пуголовки четыре луча в первом спинном плавнике отмечают очень редко (182 : 1). Этот признак хорош тем, что вероятность несоответствия результатов, полученных различными исследователями при подсчёте, в отличие от других показателей (Решетников, Попова, 2015) минимальна.
Таблица 2.
Выборки пуголовок из Азово-Черноморского бассейна и водохранилищ Волжского каскада с разным числом лучей в первом спинном плавнике (D1)
| Вид | Бассейн, водоём (годы сбора) | Число лучей в D1 | Источник информации | |||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |||
| B. nudus | Черноморский бассейн | 3 | 127 | 14 | Boldyrev, Bogutskaya, 2007 | |
| Benthophilus sp. | Р. Днепр, Каховское вдхр. (1979) | >18 | Смирнов, 1986 | |||
| B. stellatus | Азовское море | 8 | Манило, 2011 | |||
| То же | Азовский бассейн | 1 | 6 | 92 | 1 | Boldyrev, Bogutskaya, 2007 |
| » | Р. Дон, Цимлянское вдхр. (2014) | 2 | 8 | 90 | Наши данные | |
| B. durrelli | Азовский бассейн | 73 | 58 | Boldyrev, Bogutskaya, 2007 | ||
| То же | Р. Дон, Цимлянское вдхр. (2013) | 43 | 57 | Наши данные | ||
| Benthophilus sp. | Р. Волга, Рыбинское вдхр. (2014) | 1 | Кодухова и др., 2016 | |||
| То же | Р. Волга, Чебоксарское вдхр. (2007) | 2–4 (3.3) | Касьянов, Клевакин, 2011 | |||
| » | Р. Волга, Куйбышевское вдхр. (1970–1971) | 3–4 | Гавлена, 1973 | |||
| » | Р. Волга, Куйбышевское вдхр. (2004–2010) | 3–4 (3.2) | Семенов, 2011 | |||
| » | Р. Волга, Куйбышевское вдхр. (2003–2004) | 3–4 | Шемонаев, Кириленко, 2011 | |||
| » | То же | 1 | 4 | Кодухова и др., 2016 | ||
| » | Р. Волга, Саратовское вдхр. | 1 | 1 | То же | ||
| B. durrelli | Р. Волга, Саратовское вдхр. (2001) | 1 | Наши данные | |||
| То же | Р. Волга, Волгоградское вдхр. (2002) | 3 | 6 | То же | ||
| Benthophilus sp. | Р. Днепр, верховья (2011–2012) | 1 | Rizevsky, 2013 | |||
| То же | Р. Днепр, верховья (2014) | 2 | 4 | Григорчик, 2019 | ||
| B. durrelli | Р. Днестр, низовья (2006) | 3 | 13 | Романеску, 2015; наши данные | ||
| B. mahmudbejovi | Каспийский бассейн | 13 | 47 | Boldyrev, Bogutskaya, 2007 | ||
Молекулярно-генетический анализ различных представителей рода Benthophilus (Karabanov et al., 2022) с использованием в качестве маркера для определения видовой принадлежности фрагмента митохондриальной ДНК, включающего ген первой субъединицы цитохром оксидазы c (COI), продемонстрировал обособленность с незначительной генетической дистанцией, помимо нескольких видов (зернистая, Абдурахманова, Берга), двух сестринских клад. Первая (4 экз.) объединяет в себе депонированный в базе данных NCBI (https://www.ncbi.nlm.nih.gov) под номером FJ526834 экземпляр B. mahmudbejovi, две особи из Саратовского водохранилища и одну из Волги у Астрахани; вторая (8 экз.) – пуголовок из Чебоксарского и Волгоградского водохранилищ, Волги (без указания локализации), Камы, нижнего течения Волги и дельты Днестра (FJ526835). Ранее опубликованные материалы (Boldyrev, Bogutskaya, 2007; Болдырев, 2021), а также имеющиеся данные по размерному составу и соотношению лучей в первом спинном плавнике у пуголовок из волжских водоёмов позволяют предположить, что экземпляры второй группы относятся к B. durrelli, которая к настоящему времени расселилась и на незарегулированном участке нижней Волги, а первой – к B. mahmudbejovi. Последняя наиболее морфологически и, видимо, экологически близка по сравнению с другими видами рода к B. durrelli. Она ещё мельче, максимальная SL не превышает 55 мм. Число лучей в первом спинном плавнике у неё также, в отличие от B. stellatus, часто равно 4 (табл. 2). Присутствие в кладе с B. durrelli пуголовки из Днестра, вероятно, подтверждает ранее сделанное предположение о том (Boldyrev, Bogutskaya, 2007), что ареал этого вида включает помимо Азовского и Черноморский бассейн. Эти выводы нуждаются в подтверждении.
Кроме донской и предположительно пуголовки Махмудбекова к настоящему времени имеются факты присутствия в водоёмах Волжского каскада и чужеродной в Каспийском бассейне звёздчатой пуголовки. Об этом свидетельствует указание на поимки отдельных экземпляров SL до 76 мм в Чебоксарском (Клевакин, 2003), SL 68 мм в Саратовском (Кудерский и др., 1995) и SL более 70 мм в Куйбышевском (личное сообщение и фотоматериалы И.Ф. Галанина, Казанский (Приволжский) федеральный университет, 2013 г.) водохранилищах. Имеются сведения, требующие подтверждения, о поимке этого вида в Оке у г. Муром (2019 г.). Улов пуголовок в Чебоксарском водохранилище в октябре 2007 г., где в значительном количестве присутствовали самки SL 48–59 мм, по-видимому, тоже частично состоял из звёздчатых пуголовок (Касьянов, Клевакин, 2011). Предположительное отсутствие представителей этого вида, указанное в работе Карабанова с соавторами (Karabanov et al., 2022), по-видимому, свидетельствует об относительно низкой численности звёздчатых пуголовок в водоёмах Волжского бассейна по сравнению с пуголовками донской и Махмудбекова.
Список литературы
Александрова З.В., Семёнов А.Д., Ромова М.Г. и др. 1998. Режим кислорода и содержание биогенных веществ Азовского моря в многолетнем аспекте // Основные проблемы рыбного хозяйства и охраны рыбохозяйственных водоёмов Азово-Черноморского бассейна. Ростов н/Д.: Изд-во АзНИИРХ. С. 34–48.
Вехов Д.А., Науменко А.Н., Горелов В.П. и др. 2014. Современное состояние и использование водных биоресурсов Цимлянского водохранилища (2009−2013 гг.) // Рыбохозяйственные исследования на водных объектах европейской части России. СПб.: Изд-во ГосНИОРХ. С. 116–145.
Биология и промысловое значение рыбцов (Vimba) Европы. 1970. Вильнюс: Минтис, 516 с.
Богачик Т.А. 1967. Морфологические адаптации челюстно-глоточного аппарата бычков (Gobiidae) // Вопр. ихтиологии. Т. 7. № 1 (42). С. 108–116.
Богуцкая Н.Г., Болдырев В.С., Насека А.М. 2004. Бычки Neogobiinae (Teleostei, Gobiidae) в экосистемах Евразии и североамериканских Великих озёр // Биологические инвазии в водных и наземных экосистемах. М.: Т-во науч. изд. КМК. С. 297–320.
Болдырев В.С. 2002. Видовой состав бычков (Gobiidae) в Волгоградском и Цимлянском водохранилищах // Рыбохозяйственные исследования в бассейне Волго-Донского междуречья на современном этапе. СПб.: Квинта Северо-Запад. С. 98–105.
Болдырев В.С. 2007. Роль бычков (Gobiidae) в питании хищных рыб Цимлянского и Волгоградского водохранилищ // Матер. Междунар. науч.-практ. конф. “Состояние, охрана, воспроизводство и устойчивое использование биологических ресурсов внутренних водоемов”. Волгоград: Изд-во Волгоград. отд. ГосНИОРХ. С. 28–35.
Болдырев В.С. 2021. Особенности биологии налима Lota lota (Gadidae) нижнего течения Волги // Вопр. ихтиологии. Т. 61. № 4. С. 423–431. https://doi.org/10.31857/S0042875221040019
Болдырев В.С., Вехов Д.А., Хоружая В.В. и др. 2021. Ихтиофауна Цимлянского водохранилища // Вопр. ихтиологии. Т. 61. № 1. С. 36–45. https://doi.org/10.31857/S0042875221010057
Васильева Е.Д. 1983. Остеологический анализ некоторых каспийских пуголовок в связи с систематикой рода Benthophilus (Gobiidae) // Вопр. ихтиологии. Т. 23. № 4. С. 544–556.
Гавлена Ф.К. 1973. Звёздчатая пуголовка в Куйбышевском водохранилище // Вопр. ихтиологии. Т. 13. № 1 (78). С. 174–175.
Григорчик А.П. 2019. Морфометрическая характеристика пуголовки звёздчатой Benthophilus stellatus (Sauvage, 1874) из реки Днепр (Беларусь) // Изв. НАН Беларуси. Сер. биол. наук. Т. 64. № 4. С. 492–498. https://doi.org/10.29235/1029-8940-2019-64-4-492-49
Дренски П. 1951. Рибите в България. София: Изд-во Бълг. АН, 270 с.
Ермолин В.П. 1980. Питание и степень использования рыбами кормовых ресурсов водохранилища // Тр. Саратов. отд. ГосНИОРХ. Т. 18. С. 63–77.
Ермолин В.П. 1984. Экология питания рыб и пути повышения рыбопродуктивности Саратовского водохранилища: Автореф. дис. … канд. биол. наук. Л.: ГосНИОРХ, 24 с.
Желтенкова М.В. 1964. Межвидовые и внутривидовые взаимоотношения бентосоядных рыб Азовского моря // Вопр. ихтиологии. Т. 4. № 3 (32). С. 545–561.
Иванков В.Н. 2001. Репродуктивная биология рыб. Владивосток: Изд-во ДВГУ, 224 с.
Иванчев В.П., Иванчева Е.Ю. 2010. Круглоротые и рыбы Рязанской области и прилежащих территорий. Рязань: НП “Голос губернии”, 292 с.
Ивлев В.С. 1955. Экспериментальная экология питания рыб. М.: Пищепромиздат, 252 с.
Ильин Б.С. 1927а. Биология азовских пуголовок (Benthophilus, Pisces, Gobiidae) // Изв. Гос. ин-та опытн. агрономии. Т. 5. № 4. С. 308–309.
Ильин Б.С. 1927б. Определитель бычков (Fam. Gobiidae) Азовского и Чёрного морей // Тр. Азово-Черном. науч.-промысл. экспедиции. № 2. С. 128–143.
Иоффе Ц.И. 1968. Обзор выполненных работ по акклиматизации кормовых беспозвоночных для рыб в водохранилищах // Изв. ГосНИОРХ. Т. 67. С. 7–29.
Калинина Э.М. 1976. Размножение и развитие черноморско-азовских бычков. Киев: Наук. думка, 120 с.
Касьянов A.Н., Клевакин А.А. 2011. Звёздчатая пуголовка Benthophilus stellatus (Sauvage, 1874) Чебоксарского водохранилища // Рос. журн. биол. инвазий. Т. 4. № 3. С. 2–6.
Киналев Н.М. 1937. Питание бычков в Северном Каспии // Зоол. журн. Т. 16. Вып. 4. С. 755–772.
Клевакин А.А. 2003. Звёздчатая пуголовка Benthophilus stellatus (Sauvage, 1874) – новый вид рыб отряда окунеобразных (Perciformes) Чебоксарского водохранилища // Тез. докл. междунар. конф. “Экологические проблемы крупных рек – 3”: Тольятти: Изд-во ИЭВБ РАН. С. 120.
Коблицкая А.Ф. 1966. Определитель молоди рыб дельты Волги. М.: Наука, 166 с.
Кодухова Ю.В., Боровикова Е.А., Карабанов Д.П. 2016. Первая находка звёздчатой пуголовки Benthophilus stellatus (Sauvage, 1874) (Actinopterygii: Gobiidae) в Рыбинском водохранилище // Биология внутр. вод. № 4. С. 94–96.
Костюченко В.А. 1960. Питание бычка-кругляка и использование им кормовой базы Азовского моря // Тр. АзНИИРХ. Т. 1. № 1. С. 341–360.
Кудерский Л.А., Гончаренко К.С., Дрозжина К.С. и др. 1995. Звёздчатая пуголовка в водохранилищах волжского каскада // Результаты работ по акклиматизации водных организмов. СПб.: Изд-во ГосНИОРХ. С. 136–146.
Куликова Н.И., Фандеева В.Н. 1975. О порционности икрометания бычка-кругляка (Gobius melanostomus Pallas) // Тр. ВНИРО. Т. 96. Вып. 4. С.18–27.
Майский В.Н. 1955. Питание и кормовая база судака в Азовском море // Тр. ВНИРО. Т. 31. Вып. 1. С. 337–355.
Макеева А.П., Павлов Д.С., Павлов Д.А. 2011. Атлас молоди пресноводных рыб России. М.: Т-во науч. изд. КМК, 383 с.
Манило Л.Г. 2011. Пуголовка азовская, Benthophilus magistri (Gobiidae, Perciformes) — редкий вид в ихтиофауне Украины // Збірник праць Зоологічного музею. № 42. С. 92–99.
Методическое пособие по изучению питания и пищевых отношений рыб в естественных условиях. 1974. М.: Наука, 254 с.
Мовчан Ю.В., Манило Л.Г., Смирнов А.И. и др. 2003. Каталог коллекций зоологического музея ННПМ НАН Украины. Круглоротые и рыбы. Киев: Изд-во Зоомузея ННПМ НАНУ, 241 с.
Насека А.М., Болдырев В.С., Сподарева В.В. и др. 2012. Каталог фондовой коллекции Зоологического института РАН. Класс лучепёрые рыбы (Actinopterygii). Отряд окунеобразные (Perciformes), подотряд бычковидные (Gobioidei). Семейство бычковые (Gobiidae) черноморско-каспийского бассейна // Исследования фауны морей. Т. 73 (81). СПб.: Т-во науч. изд. КМК, 123 с.
Овен Л.С. 1976. Особенности оогенеза и характер нереста морских рыб. Киев: Наук. думка, 132 с.
Рагимов Д.Б. 1981. Распространение и численность бычковых (Gobiidae) в Северном Каспии // Вопр. ихтиологии. Т. 21. № 2 (127). С. 223–231.
Рагимов Д.Б. 1985. Материалы по размножению некоторых каспийских видов пуголовок рода Benthophilus Eichwald (Gobiidae) // Вопр. ихтиологии. Т. 25. № 2. С. 242‒247.
Ращеперин В.К. 1964. Особенности порционного икрометания бычка-кругляка Азовского моря и численность его молоди // Труды молодых ученых. М.: Пищепромиздат. С. 70–74.
Рейх Е.М. 1976. Питание молоди некоторых азовоморских бычков // Биологические ресурсы Азовского моря. Ростов н/Д: Изд-во РГУ. С. 26–34.
Решетников Ю.С., Попова О.А. 2015. О методиках полевых ихтиологических исследований и точности полученных результатов // Тр. ВНИРО. Т. 156. С. 114–131.
Решетников Ю.С., Попова О.А., Кияшко В.И. и др. 2016. Обыкновенный ёрш Gymnocеphalus cernuus (Linnaeus, 1758). М.: Т-во науч. изд. КМК, 279 с.
Романеску В. 2015. Первая находка Benthophilus durrelli Boldyrev et Bogutskaya, 2004 (Perciformes: Gobiidae) в реке Днестр // Чтения памяти Л.Л. Попа. Тирасполь: Изд-во ПГУ. С. 102–108.
Семенов Д.Ю. 2011. Данные о морфологии и биологии звёздчатой пуголовки Benthophilus stellatus (Perciformes, Gobiidae) // Пробл. регион. экологии. № 1. С. 73–77.
Смирнов А.И. 1986. Фауна Украины. Т. 8. Рыбы. Вып. 5. Киев: Наук. думка, 313 с.
Солодовников С.В. 1930. Риби, зібрані Донецькою Науковою Експедицією (р. 1927) // Труди Донецької Наукової Експедиції. № 1. Харків: Пролетарий. С. 33–40.
Спановская В.Д., Григораш В.А. 1976. К методике определения плодовитости единовременно и порционно икромечущих рыб // Типовые методики исследований продуктивности видов рыб в пределах их ареалов. Ч. 2. Вильнюс: Мокслас. С. 54–75.
Студеникина Е.И., Толоконникова Л.И., Воловик С.П. и др. 1998. Характеристика донных сообществ в современный период // Основные проблемы рыбного хозяйства и охраны рыбохозяйственных водоёмов Азово-Черноморского бассейна. Ростов н/Д.: Изд-во АзНИИРХ. С. 67–77.
Трифонов Г.П. 1955. Биология размножения азовских бычков // Тр. Карадаг. биостанции. № 13. С. 5–45.
Ульман Э.Ж. 1970. Бычки Каховского водохранилища и их биологическое значение // Автореф. дис. … канд. биол. наук. Ростов н/Д.: РГУ, 20 с.
Шакирова Ф.М., Северов Ю.А., Латыпова В.З. 2015. Современный состав чужеродных видов рыб Куйбышевского водохранилища и возможности проникновения новых представителей в экосистему водоёма // Рос. журн. биол. инвазий. Т. 8. № 3. С. 77–98.
Шандиков Г.О., Гончаров Г.Л. 2008. Редкие виды рыб бассейна Северского Донца северо-восточной Украины // Вестн. Харьков. нац. ун-та. Сер. биол. Вып. 8. № 828. С. 65–90.
Шашуловский В.А., Ермолин В.П. 2005. Состав ихтиофауны Волгоградского водохранилища // Вопр. ихтиологии. Т. 45. № 3. С. 324–330.
Шемонаев Е.В., Кириленко Е.В. 2011. Данные о морфологии и биологии пуголовки звёздчатой Benthophilus stellatus (Sauvage, 1874) Куйбышевского водохранилища // Вестн. СамГУ. № 5 (86). С. 182–186.
Шорыгин А.А. 1952. Питание и пищевые взаимоотношения рыб Каспийского моря. М.: Пищепромиздат, 268 с.
Bănărescu P. 1964. Fauna Republicii Populare Române, Pisces−Osteichthyes. V. 13. Bucureşti: Acad. R.P. Române, 959 p.
Boldyrev V.S., Bogutskaya N.G. 2004. A description of two new species of tadpole-gobies (Teleostei: Gobiidae: Benthophilus) // Zoosyst. Rossica. V. 13. № 1. P. 129–135.
Boldyrev V.S., Bogutskaya N.G. 2007. Revision of the tadpole-gobies of the genus Benthophilus (Teleostei: Gobiidae) // Ichthyol. Explor. Freshwat. V. 18. № 1. P. 31–96.
Karabanov D.P., Bekker E.I., Pavlov D.D. et al. 2022. New Sets of Primers for DNA Identification of Non-Indigenous Fish Species in the Volga-Kama Basin (European Russia) // Water. V. 14. № 3. Article 437. https://doi.org/10.3390/w14030437
Kottelat M., Freyhof J. 2007. Handbook of European freshwater fishes. Cornol; Berlin: Kottelat and Freyhof, 646 p.
Kovačić M., Esmaeili H.R., Zarei U.K. et al. 2021. A new species of tadpole-goby, Benthophilus persicus sp. nov. (Teleostei: Gobiidae) from the southern Caspian Sea // Zoo-taxa. V. 4980. № 1. P. 45–63. https://doi.org/10.11646/zootaxa.4980.1.3
Miller P.J. 1986. Gobiidae // Fishes of the North-eastern Atlantic and the Mediterranean. V. 3. Paris: UNESCO. P. 1019–1085.
Patimar R., Qaranjiki A., Bahalkeh A. 2019. Life history traits of the Caspian stellate tadpole-goby Benthophilus leobergius Berg, 1949 (Teleostei: Gobiidae) from the southeastern Caspian Sea, Iran // Iran. J. Ichthyol. V. 6. № 4. P. 254–263. https://doi.org/10.22034/iji.v6i4.361
Rizevsky V., Leschenko A., Ermolaeva I. et al. 2013. First record of the Ponto-Caspian stellate tadpole-goby Benthophilus stellatus (Sauvage, 1874) from the Dnieper River, Belarus // Bioinvasions Rec. V. 2. № 2. P. 159–161. https://doi.org/10.3391/BIR.2013.2.2.12
Sindilariu P., Freyhof J. 2003. Food overlap of benthic fishes in the Danube Delta, with special respect to two invasive gobiids (Teleostei: Gobiidae, Percidae, Cyprinidae) // Lauterbornia. № 46. P.149-157.
The freshwater fishes of Europe. 2004. V. 8/II. Gobiidae. 2. Weibelsheim: AULA-Verlag, 478 p.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии