

Вопросы ихтиологии, 2023, T. 63, № 2, стр. 233-237
Первое нахождение личинок неописанного вида из рода Lyconus (Lyconidae) в водах Северной Пацифики
Я. Ю. Большакова 1, *, С. А. Евсеенко 1
1 Институт океанологии РАН – ИО РАН
Москва, Россия
* E-mail: yanusrunaa@mail.ru
Поступила в редакцию 02.06.2022
После доработки 21.06.2022
Принята к публикации 24.06.2022
- EDN: EXYCGZ
- DOI: 10.31857/S0042875223020030
Аннотация
Приведено первое иллюстрированное описание двух личинок редкого глубоководного рода Lyconus из северной части Тихого океана. Исследованные экземпляры отличаются от ранее описанных личинок L. pinnatus полной и равномерной пигментацией тела. Новые материалы дополняют существующие представления о географическом распространении представителей семейства Lyconidae – это первая их находка в водах Северной Пацифики.
Представители рода Lyconus – редкие глубоководные рыбы (Cohen et al., 1990). До недавнего времени род провизорно рассматривали в составе семейства Merlucciidae в силу недостаточности знаний о морфологии входящих в его состав видов из-за редкости поимок (Okamura, 1989; Cohen, 1990; Lloris et al., 2005). Согласно последним молекулярно-генетическим данным, род Lyconus признают единственным членом самостоятельного семейства Lyconidae, образующем сестринскую кладу с семействами Bathygadidae и Macruronidae (von der Heyden, Matthee, 2008; Roa-Varón et al., 2021). Род включает два вида: L. pinnatus Gunther, 1887 из вод Южного полушария и L. brachycolus Brauer, 1906 из вод Атлантического океана (Fricke et al., 2022). Также имеются сведения о ещё двух не описанных видах: Lyconus sp., пойманном у о-ва Ява (Евсеенко, Сунцов, 1995), и Lyconus sp., обнаруженном на плато Кэмпбелл (южнее Новой Зеландии) (Inada, 1990). Иллюстрированные описания ранних стадий развития известны только для L. pinnatus (Евсеенко, Сунцов, 1995; Евсеенко и др., 2014). Новые личинки обнаружены в сборах северо-восточной части Тихого океана – сведения о нахождении здесь ликонусов отсутствуют. Цель предлагаемой статьи – документировать новое место поимки, дать первое описание и иллюстрацию личинок этого вида, а также сравнить их с ранее описанными личинками L. pinnatus.
МАТЕРИАЛ И МЕТОДИКА
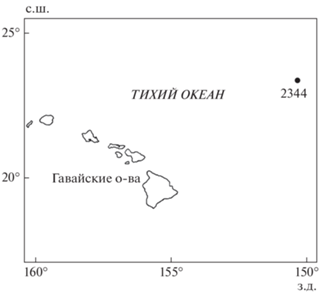
Две личинки Lyconus sp. общей длиной TL 37+ и 39+ мм были обнаружены в сборах 22-го рейса научно-исследовательского судна “Академик Мстислав Келдыш” в северную часть Тихого океана. Для удобства обсуждения мы обозначим этих личинок по месту их поимки – L. sp. “hawaiiensis”. Они были выловлены 09.09.1990 г. на станции 2344 (23°11′07′′ с.ш., 150°25′01′′ з.д.) к северо-востоку от Гавайских о-вов при облове слоя 0–1000 м (рис. 1). Сборы проводили разноглубинным тралом Айзекса–Кидда в модификации Самышева–Асеева – РТАКСА (длина 25 м, площадь устья 6 м2, ячея 5 мм) с концевой вставкой из нейлоновой сетки 500 мкм. Экземпляры зафиксированы в 4%-м растворе формальдегида. Личинку TL 37+ мм окрасили ализарином по стандартной методике (Taylor, van Dyke, 1985). В настоящее время личинки хранятся в ихтиологической коллекции ИО РАН (IORAS 04469).
Рис. 1.
Местоположение станции (⚫) отлова личинок Lyconus sp. “hawaiiensis” 09.09.1990 г. в ходе 22-го рейса научно-исследовательского судна “Академик Мстислав Келдыш” в северо-западную часть Тихого океана.
В работе использованы следующие сокращения: TL – общая длина тела, с – длина головы, H – максимальная высота тела; aA, aD, aV – антеанальное, антедорсальное и антевентральное расстояния; ао – длина рыла, о – горизонтальный диаметр глаза, po – длина заглазничного отдела головы, io – межглазничное расстояние, lmx – длина верхней челюсти; D (D1, D2), А, Р, V – число лучей соответственно в спинном (первом и втором), анальном, грудном и брюшном плавниках; vert.а., vert.с. – число туловищных и хвостовых позвонков; Br. – число бранхиостегий, sp.br. – число жаберных тычинок на первой жаберной дуге.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Описание. TL 37+ (с 7.4 мм) и 39+ (с 7.5 мм) мм (кончик хвостового отдела отсутствует у обеих личинок), D1 8–9, D2 77+, А 60+, P 15, V 9, vert.а. 16, vert.с. 60+, sp.br. 3 + 11 = 14, Br. 7, число зубов на сошнике 0.
Морфология. Тело личинок массивное, высокое в передней части, в каудальном направлении плавно сужается (рис. 2, 3). Туловищный отдел массивный, брюхо сильно раздуто. Высота тела за грудными плавниками составляет 96% с. Голова крупная и высокая, верхний профиль головы с заметным прогибом, глаза деформированы; рот косой, угол рта достигает вертикали переднего края глазницы; челюсти с мелкими зубами, на сошнике и на нёбных костях зубы отсутствуют. Жаберные тычинки на внешнем крае 1-й жаберной дуги относительно длинные и тонкие, на внутреннем крае – короткие с шипами на дистальном крае. На крышечной кости имеются два расположенных под углом друг к другу тонких гребня. Грудные плавники расположены по горизонтальной оси на уровне глаз. Брюшные плавники сдвинуты вперёд относительно грудных, их основания вытянутой треугольной формы. Спинной и анальный плавники длинные, лучи спинного плавника заметно длиннее лучей анального. Начало первого спинного плавника над основанием грудных плавников. Птеригиофор первого луча второго спинного плавника заметно выступает над краем тела (сильнее, чем последний птеригиофор первого спинного плавника), при этом расстояние между последним лучом D1 и первым лучом D2 не отличается от дистанции между лучами D1. Анальный плавник начинается на уровне 14–15-го луча второго спинного плавника. Кишечник короткий, мешковидный, брюхо личинок сильно раздуто от пищи. Анус располагается примерно посередине тела. Некоторые промеры тела личинки TL 39+ мм, в % с: H 96, aA 226, aD 113, aV 113, ао 28, о 31, po 41, io 24, lmx 51.
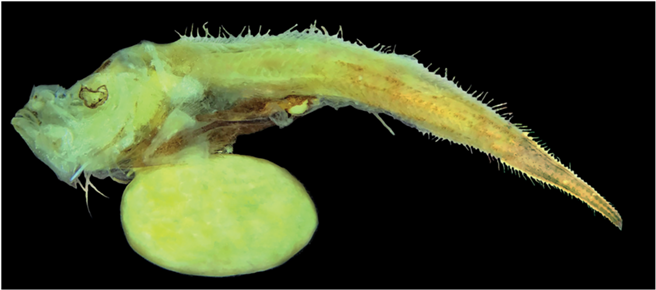
Рис. 3.
Внешний вид личинки Lyconus sp. “hawaiiensis” TL 37+ мм; реконструирована брюшная часть (рисунок Я.Ю. Большаковой).
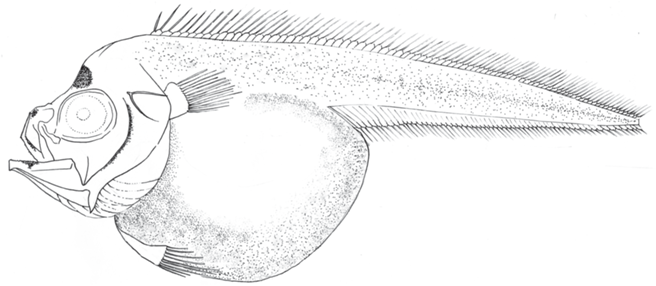
Пигментация. Личинки умеренно пигментированы (рис. 2, 3). Точечные меланофоры равномерно рассеяны по всему телу. Брюхо личинок окрашено интенсивнее, наиболее густо пигмент расположен в вентролатеральной части. Перитонеум прозрачный. Лучи во всех плавниках и основания парных плавников не окрашены. На голове пигмент концентрируется над средним мозгом, позади предкрышечной кости, у обонятельной капсулы, латерально на хряще (mesethmoid cartilage). Имеются меланофоры вдоль praemaxillare, а также ряд на нижней челюсти. Подбородочная часть равномерно пигментирована точечными меланофорами. Внутренние пигментные пятна, супротивно расположенные вдоль тел позвонков, просматриваются в дистальной половине хвостовой части тела.
Сравнительные замечания. Заметная особенность личинок из Северо-Восточной Пацифики – раздутое брюхо. Такая черта характерна для ранних стадий развития некоторых представителей отряда Gadiformes: Macrouridae (Merrett, 1978, 1989), Moridae (Matsuura, Aizawa, 1990; Meléndez, Markle, 1997; Евсеенко, Трунов, 2004) и Lyconidae (Евсеенко, Сунцов, 1995; Евсеенко и др., 2014). Спинной и анальный плавники предположительно сливаются с хвостовым, а промежуток между спинными плавниками практически отсутствует, что характерно для представителей семейств Bathygadidae, Lyconidae и Steindachneriidae (Cohen et al., 1990). Описание личинки TL 24 мм единственного вида семейства Steindachneriidae – Steindachneria argentea Goode et Bean, 1896 – в литературе имеется (Fahay, Markle, 1984). При TL 24 мм личинка уже демонстрирует признаки, характерные для взрослых особей этого вида, а именно: сформирован светящийся орган, а между анусом и урогенитальной папиллой есть заметное расстояние. Кроме этого, личинка S. argentea отличается более удлинённым телом, меньшим антеанальным расстоянием и пигментацией. В литературе имеются иллюстрированные описания пелагических личинок батигадин всего двух видов. Это личинка TL 30+ мм, предварительно идентифицированная как Gadomus sp. (Fahay, Markle, 1984), и личинка Bathygadus cottoides Günther, 1878 TL 22+ мм (Большакова, Евсеенко, 2016). Обе личинки имеют сходные особенности морфологии, которые отличают их от описанных личинок рода Lyconus: брюшные плавники сдвинуты назад относительно грудных и расположены латерально на брюхе, основания грудных плавников стебельчатые, на предкрышке имеются два кожистых выроста. Кроме этого, личинок Lyconidae можно отличить от Bathygadidae по меристическим признакам: по числу туловищных позвонков (11–13 у Bathygadidae против 14–20 у Lyconidae) и числу жаберных тычинок (sp.br. >18 у Bathygadidae против <17 у Lyconidae) (Cohen et al., 1990; Iwamoto, Anderson, 1994; Евсеенко, Сунцов, 1995; Iwamoto, 2002).
Личинки L. sp. “hawaiiensis” отличаются по меристическим признакам от известных видов рода Lyconus (таблица) – числом туловищных позвонков (меньше, чем у L. pinnatus и L. brachycolus, но больше, чем у L. sp. с о-ва Ява); отсутствием зубов на сошнике (напротив, имеются у L. pinnatus, L. brachycolus и L. sp. с о-ва Ява). Исходя из того факта, что у личинки L. pinnatus TL 17.5 мм (взрослые особи которого имеют зубы на сошнике) зубы уже имеются, отсутствие зубов у личинок L. sp. “hawaiiensis” вряд ли связано с размером особей. Отсутствие или наличие зубов на сошнике (но не их количество), по всей вероятности, является важным видоспецифичным признаком. Особи, описанные с плато Кэмпбелл (Inada, 1990), также не имеют зубов на сошнике, но при этом у них больше число лучей в грудных плавниках, чем у личинок L. sp. “hawaiiensis”. В целом по комплексу меристических признаков описанные новые личинки отличаются от всех известных на данный момент видов ликонусов.
Основные меристические признаки всех известных видов рода Lyconus
| Вид | vert.a. | D1 | P | V | sp.br. | Число зубов на сошнике |
|---|---|---|---|---|---|---|
| L. pinnatus | 19–20 | 12–13 | 15–17 | 9–10 | 3–4 + 11–13 | 1–2/1–3 |
| L. brachycolus | 17–18 | 9–10 | 13–14 | 8–9 | 3–5 + 9–12 | 1–10/2–8 |
| L. sp. (о. Ява) | 14 | – | 15 | 9 | 3 + 10 | 6/6 |
| L. sp. (плато Кэмпбелл) | – | – | 17–18 | 10 | 0–2 + 11–12 | 0 |
| L. sp. “hawaiiensis” | 16 | 8–9 | 15 | 9 | 3 + 11 | 0 |
Примечание. vert.a. – число туловищных позвонков, D1, Р, V – число лучей соответственно в первом спинном, грудном и брюшном плавниках; sp.br – число жаберных тычинок на первой жаберной дуге, “–” – нет данных. Источники информации: Cohen et al., 1990; Inada, 1990; Евсеенко, Сунцов, 1995; Lloris et al., 2005.
Описание ранних стадий развития имеется только для L. pinnatus (Евсеенко, Сунцов, 1995; Евсеенко и др., 2014). Габитуально личинки L. sp. “hawaiiensis” очень похожи на личинок L. pinnatus по форме тела, верхнему профилю головы, раздутому брюху, формой и длиной жаберных тычинок. Положение брюшных плавников подвержено индивидуальной изменчивости (Евсеенко, Сунцов, 1995), а у личинок зависит, по-видимому, от степени наполнения желудка. Личинки L. sp. “hawaiiensis” наиболее заметно отличаются от личинок L. pinnatus по характеру пигментации (рис. 2, 3): тело первых пигментировано равномерно и полностью, в то время как у вторых задняя треть тела лишена пигмента, кроме разрозненных меланофоров. Такая окраска характерна как для самых мелких личинок TL 13.0 и 17.5 мм, так и для крупного предмалька TL 50+ мм. Помимо этого различия хорошо заметны и в счётных признаках (таблица).
Учитывая различия в меристических признаках между исследованными личинками и известными видами ликонусов, а также их поимку далеко за пределами установленных ареалов видов, мы предполагаем, что новые особи могут принадлежать к ещё не описанному виду рода Lyconus.
Список литературы
Большакова Я.Ю., Евсеенко С.А. 2016. О видовом составе ихтиопланктона Китового хребта (Южная Атлантика) // Вопр. ихтиологии. Т. 56. № 6. С. 685–697. https://doi.org/10.7868/S0042875216060035
Евсеенко С.А., Сунцов А.В. 1995. Личинки Lyconus pinnatus (Merlucciidae) из юго-западной Пацифики с замечаниями о диагностике видов рода Lyconus // Вопр. ихтиологии. Т. 35. № 4. С. 469–477.
Евсеенко С.А., Трунов И.А. 2004. Пелагические мальки моровой рыбы Guttigadus kongi (Gadiformes, Moridae) из юго-западной Атлантики // Вопр. ихтиологии. Т. 44. № 4. С. 480–483.
Евсеенко С.А., Большакова Я.Ю., Маликова Д.Ю. 2014. О поимке предмалька редкого глубоководного вида Lyconus pinnatus (Merlucciidae) в центральных водах Срединно-Атлантического хребта // Вопр. ихтиологии. Т. 54. № 3. С. 368–370. https://doi.org/10.7868/S0042875214030035
Cohen D., Inada T., Iwamoto T., Scialabba N. 1990. Gadiform fishes of the world (Order Gadiformes). An annotated and illustrated catalogue of cods, hakes, grenadiers and other gadiform fishes known to date // FAO Fish. Synop. V. 10. № 125. 442 p.
Fahay M.P., Markle D.F. 1984. Gadiformes: development and relationships // Ontogeny and systematics of fishes. Lawrence: Allen Press. P. 265–283.
Fricke R., Eschmeyer W.N., van der Laan R. (eds.). 2022. Eschmeyer’s catalog of fishes: genera, species, references (http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp. Version 06/2022).
Inada T. 1990. Merlucciidae // Fishes collected by the R/V Shinkai Maru around New Zealand. Tokyo: Jpn. Mar. Fish. Resour. Res. Center. P. 163–167.
Iwamoto T. 2002. Bathygadidae // The living marine resources of the Western Central Atlantic. V. 2: Bony fishes. Part 1 (Acipenseridae to Grammatidae). Rome: FAO. P. 988–990.
Iwamoto T., Anderson M.E. 1994. Review of the grenadiers (Teleostei: Gadiformes) of southern Africa, with descriptions of four new species // Ichthyol. Bull. V. 61. 28 p.
Lloris D., Matallanas J., Oliver P. 2005. Hakes of the World (Family Merlucciidae): an annotated and illustrated catalogue of hake species known to date. № 2. Rome: FAO, 57 p.
Matsuura K., Aizawa M. 1990. Moridae, genus and species undetermined // Fishes collected by the R/V Shinkai Maru around New Zealand. Tokyo: Jpn. Mar. Fish. Resour. Res. Center. P. 158–162.
Meléndez R.C., Markle D.F. 1997. Phylogeny and zoogeography of Laemonema and Guttigadus (Pisces; Gadiformes; Moridae) // Bull. Mar. Sci. V. 61. № 3. P. 593–670.
Merrett N.R. 1978. On the identity and pelagic occurrence of larval and juvenile stages of rattail fishes (Family Macrouridae) from 60°N, 20°W and 53°N, 20°W // Deep Sea Res. V. 25. № 2. P. 147–160. https://doi.org/10.1016/0146-6291(78)90002-4
Merrett N.R. 1989. The elusive macrourid alevin and its seeming lack of potential in contributing to intrafamilial systematics // Papers on the systematics of gadiform fishes. Nat. Hist. Mus. of Los Angeles County. Sci. Ser. № 32. P. 175–185.
Okamura O. 1989. Relationships of the suborder Macrouroidei and related groups, with comments on Merlucciidae and Steindachneria // Ibid. № 32. P. 129–142.
Roa-Varón A., Dikow R.B., Carnevale G. et al. 2021. Confronting sources of systematic error to resolve historically contentious relationships: a case study using gadiform fishes (Teleostei, Paracanthopterygii, Gadiformes) // Syst. Biol. V. 70. № 4. P. 739–755. https://doi.org/10.1093/sysbio/syaa095
Taylor W.R., van Dyke G.C. 1985. Revised procedures for staining and clearing small fishes and other vertebrates for bone and cartilage study // Cybium. V. 9. № 2. P. 107–119.
von der Heyden S., Matthee C.A. 2008. Towards resolving familial relationships within the Gadiformes and the resurrection of the Lyconidae // Mol. Phylogenet. Evol. V. 48. № 2. P. 764–769. https://doi.org/10.1016/j.ympev.2008.01.012
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии