

Вопросы ихтиологии, 2023, T. 63, № 2, стр. 132-143
Зоогеографический анализ особенностей температурных предпочтений рыб европейской части России
А. К. Смирнов *
Институт биологии внутренних вод РАН – ИБВВ РАН
Ярославская обл., пос. Борок, Россия
* E-mail: smirnov@ibiw.ru
Поступила в редакцию 30.06.2022
После доработки 20.07.2022
Принята к публикации 20.07.2022
- EDN: FAFKLS
- DOI: 10.31857/S0042875223020248
Аннотация
Представлены результаты анализа температурных предпочтений 54 видов рыб и круглоротых, населяющих пресные водоёмы европейской части России. Показано, что сведения по основным температурным критериям фундаментальной экологической ниши (оптимум, лимиты и нерестовая температура) не могут быть использованы в качестве чёткого признака принадлежности видов к тому или иному фаунистическому комплексу. Тем не менее эти критерии являются, по всей видимости, определённым отражением климатических условий, существовавших во время возникновения и становления видов. Среди изученных рыб и круглоротых выделены две обособленные группы “северных” холодолюбивых и “южных” теплолюбивых видов. Их наличие обусловлено историческими особенностями формирования ихтиофауны региона. Климатическая тенденция к росту температуры воды, наметившаяся в последние десятилетия, вероятно, благоприятствует распространению на север представителей теплолюбивой “южной” группы видов, что может привести к дальнейшей перестройке конкурентных отношений в популяциях рыб и вызвать изменения границ их ареалов.
В рамках теории фаунистических комплексов, предложенной Никольским (1956, 1980), включение видов в ту или иную фаунистическую группу базируется на единстве географического происхождения (географическая зона) и экологической специфики, изучив которую можно восстановить условия, в которых протекало их развитие. Обоснованность подобного объединения вытекает из общности возникновения и длительного периода устойчивого сосуществования видов внутри фаунистических групп и, как следствие, их совместной адаптации к некоему единому комплексу абиотических и биотических условий. Эти представления неоднократно подвергали критике (Яковлев, 1964; Решетников 1981; Рыбы Монгольской …, 1983), связанной с их недостаточным подкреплением палеонтологическим материалом; трудностью определения первичных ландшафтно-климатических условий для видов с обширными ареалами; выделением комплексов видов не по единству возникновения, а по длительности совместного сосуществования; нетождественностью фаунистических ассоциаций, объединяемых по общности ландшафтно-географического происхождения с их исторической судьбой. Тем не менее эта теория дала мощный толчок к изучению зоогеографии рыб. По словам Яковлева (1964. С. 14), “теория фаунистических комплексов, являющаяся синтезом экологического и исторического методов, представляется нам исключительно плодотворной для зоогеографического анализа”.
Трудно переоценить влияние температуры на все аспекты жизнедеятельности эктотермных (пойкилотермных) животных. Её непосредственное воздействие на интенсивность обмена веществ у этой группы организмов напрямую обусловливает темпы их роста и развития, а также эффективность воспроизводства (Angilletta et al., 2002). Очевидно, что именно температура часто выступает основным фактором, влияющим на процветание и возможности расселения эктотермных видов (Pörtner, 2002; Bennett et al., 2019). Поэтому температурные критерии (оптимум, лимиты, диапазон температур репродукции) являются важной характеристикой видовой специфики, изучив которую, согласно вышеуказанной теории Никольского, можно сделать выводы о совокупности ландшафтно-климатических условий во время возникновения отдельной филогенетической единицы, а также оценить возможности её распространения и акклиматизации.
Значения температурных критериев фундаментальной экологической ниши (определяемой в лабораторных экспериментах) и реализованной ниши (занимаемой видом в естественных условиях обитания) зачастую не совпадают. Это обусловлено тем, что поведение и распределение особей в естественной водной среде определяется единовременным воздействием множества абиотических (содержание кислорода, течение, укрытия и так далее) и биотических (трофические условия, конкуренция, хищники и так далее) факторов, также способных оказать влияние на выбор тех или иных условий. Кроме того, показатели реализованной температурной ниши для видов с обширными ареалами могут различаться в южных и северных популяциях. Однако фундаментальная температурная ниша более стабильна и, очевидно, полнее отражает эколого-историческую специфику вида. А, как отмечено выше, именно экологическая специфика служит одним из критериев объединения видов в фаунистические комплексы. В связи с этим целью настоящей работы стало: сгруппировать виды по их отношению к температуре среды; провести комплексный анализ имеющихся собственных и литературных данных по температурным критериям жизнедеятельности рыб и миног, встречающихся в европейской части России; сопоставить результаты с принадлежностью видов к выделяемым фаунистическим комплексам; оценить вероятное воздействие изменения климата на рыб с разными температурными предпочтениями.
МАТЕРИАЛ И МЕТОДИКА
Проанализированы температурные критерии жизнедеятельности рыб и круглоротых, населяющих пресные водоёмы европейской части России. Выбор региона обусловлен лучшей изученностью его ихтиофауны и, как следствие, возможностью задействовать в анализе наибольшее количество видов. Привлечён как собственный материал, так и обширный массив данных из источников литературы, содержащий сведения о температурных предпочтениях рыб. Поиск производили по трём важнейшим критериям видовой фундаментальной температурной ниши: окончательная избираемая температура (ОИТ), верхняя окончательная летальная температура (ОЛТ) и нерестовая температура (НТ) (Fry, 1947; Beitinger et al., 2000). Согласно определению Фрая (Fry, 1947), под ОИТ подразумевали температурную зону, в которой рыбы будут окончательно скапливаться независимо от предшествующих температурных условий (температуры акклимации). Для оценки ОЛТ в первую очередь отбирали данные экспериментов, выполненных методами “хронического летального максимума (chronical lethal maximum)” или “окончательных пороговых летальных температур (ultimate incipient lethal temperature)” (Beitinger et al., 2000). Несмотря на методические различия (динамический и статический нагрев), значения температурной устойчивости, установленные в обоих случаях, близки к видовому максимуму и сопоставимы (Cooper et al., 2008). В настоящей работе осознанно не использовали информацию по холодоустойчивости рыб, так как она доступна лишь для весьма ограниченного числа видов. Однако можно допустить, что нижние значения ОЛТ для большинства рассматриваемых видов находятся в температурной зоне, близкой к 0°С. При сборе информации предпочтение отдавали данным, приводимым для неполовозрелых особей. Как было показано ранее, температурные характеристики молоди более стабильны вследствие меньшего влияния сопутствующих факторов, связанных с репродукционной активностью (Голованов, 2013). Температуру, благоприятную для нереста особей какого-либо вида, как правило, указывают как некий диапазон, ограниченный температурами начала нерестовой активности и её угнетения. Поэтому в работе использовали средние значения НТ как некий нерестовый оптимум.
К сожалению, искомая информация в полном объёме имеется лишь для относительно небольшого числа рыб, населяющих пресные воды европейской части России. Поэтому для отдельных видов приходилось использовать близкие по смыслу, но несколько иные температурные критерии. Так, вместо ОИТ в некоторых случаях приведены значения, полученные в краткосрочных экспериментах (от нескольких часов до нескольких суток). В том случае, когда и эти сведения были недоступны, привлекали данные по оптимальным температурам роста, которые, как правило, близки к предпочитаемым (Jobling, 1981). В последнем случае для расчётов использовали верхние значения из известных диапазонов. По наблюдениям автора настоящей публикации, они в большей степени соответствуют зоне ОИТ. При отсутствии информации по ОЛТ привлекали данные опытов, выполненных с использованием методов “критического термического максимума” (critical thermal maximum) и “начальных летальных температур” (incipient lethal temperature) (Beitinger et al., 2000), и предпочтение также отдавали максимальным значениям. Собранные согласно вышеуказанным критериям температурные данные для 54 видов рыб и круглоротых обобщены в Приложении с указанием источников информации.
Обработку данных и графические построения выполняли в статистическом пакете STATISTICA (“StatSoft”, США). Для группировки изученных рыб и миног по температурным критериям жизнедеятельности применяли анализ главных компонент (Айвазян и др., 1989). Статистическую оценку различий между фаунистическими комплексами, а также между выделенными по результатам работы группами рыб выполняли с использованием критерия серий Вальда–Вольфовица (Sprent, Smeeton, 2007). Для выявления связи между отдельными температурными критериями применяли непараметрический корреляционный анализ по Спирмену (Sokal, Rolph, 1995). Проверку нормальности распределений температурных критериев осуществляли с использованием критерия Шапиро–Уилка (Sokal, Rolph, 1995).
РЕЗУЛЬТАТЫ
Большинство исследованных видов принадлежит к числу наиболее массовых и, как следствие, лучше изученных по отношению к температурному фактору. По оценке автора настоящей работы, они составляют ~65% общего числа рыб и круглоротых рассматриваемого региона. Охвачены фактически все семейства аборигенных и вселённых рыб, но уровень их представленности весьма различен. Так, доли (%) задействованных в анализе видов в общем числе видов своего семейства составили следующие значения: Petromyzontidae – 60, Acipenseridae – 50, Clupeidae – 33, Cyprinidae – 68, Nemacheilidae – 100, Cobitidae – 50, Siluridae – 100, Esocidae – 100, Osmeridae – 100, Salmonidae – 72, Lotidae – 100, Gasterosteidae – 67, Cottidae – 10, Percidae – 60, Odontobutidae – 100 и Gobiidae – 33. Наиболее представленным по видовому составу оказался бореальный равнинный фаунистический комплекс (14 видов), а наименее – понтический морской (2 вида).
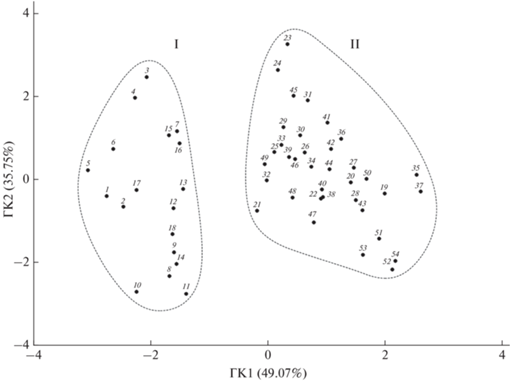
Статистический анализ имеющегося материала не выявил чёткого соответствия между температурными критериями жизнедеятельности видов и выделяемыми фаунистическими комплексами. Этот анализ включал в себя как важнейшие показатели фундаментальных температурных ниш (ОИТ, ОЛТ и НТ), так и их соотношения (разности между ОЛТ и ОИТ, ОИТ и НТ). Тем не менее были выявлены две обособленные группы видов, различающиеся по отношению к температуре среды обитания (рис. 1). Этим группам могут быть присвоены условные наименования “северная” (холодолюбивая) и “южная” (теплолюбивая). Первая из них ограничена представителями арктического пресноводного, арктобореального, а также бореального предгорного фаунистических комплексов. Из всей совокупности исследованных видов сюда вошли миноги (Petromyzontidae) – 3 вида, а также рыбы семи семейств: Salmonidae (8 видов), Osmeridae (2), Gasterosteidae (2), Balitoridae (1), Cottidae (1), Lotidae (1) и Cyprinidae (1). Вторая из выделенных групп объединила представителей бореального равнинного, древнего третичного, пресноводного и морского понтических, а также китайского равнинного фаунистических комплексов. Здесь можно отметить преобладание рыб семейства Cyprinidae (22 вида), остальные семейства представлены значительно меньшим числом видов: Percidae (3), Acipenseridae (3), Cobitidae (2), Esocidae (1), Siluridae (1), Gobiidae (1), Clupeidae (1) и Odontobutidae (1). Примечательно, что только одно семейство Cyprinidae присутствует одновременно как в “северной” (единственный представитель – обыкновенный гольян Phoxinus phoxinus (Linnaeus, 1758)), так и в “южной” группе видов. Рассчитанные на основе корреляций факторные нагрузки представлены в таблице. Их анализ позволяет сделать вывод о том, что основополагающим критерием для выделения двух вышеуказанных групп служит ОИТ (Фактор 1), а также разность между ОИТ и НТ (Фактор 2).
Рис. 1.
Распределение изученных видов рыб и круглоротых из пресных вод европейской части России в пространстве главных компонент (ГК) исходя из их температурных ниш. Группа: I – северные холодолюбивые виды (арктический пресноводный, арктобореальный и бореальный предгорный фаунистические комплексы), II – южные теплолюбивые виды (бореальный равнинный, древний третичный, понтический пресноводный, понтический морской и китайский равнинный фаунистические комплексы). 1 – Osmerus eperlanus, 2 – Osmerus mordax, 3 – Coregonus albula, 4 – Coregonus lavaretus, 5 – Salvelinus lepechini, 6 – Lota lota, 7 – Coregonus peled, 8 – Petromyzon marinus, 9 – Lampetra fluviatilis, 10 – Lampetra planeri, 11 – Gasterosteus aculeatus, 12 – Pungitius pungitius, 13 – Phoxinus phoxinus, 14 – Barbatula barbatula, 15 – Thymallus thymallus, 16 – Salmo salar, 17 – Salmo trutta, 18 – Cottus gobio, 19 – Carassius auratus complex, 20 – Carassius carassius, 21 – Gobio gobio, 22 – Squalius cephalus, 23 – Leuciscus idus, 24 – Leuciscus leuciscus, 25 – Rhynchocypris percnura, 26 – Rutilus rutilus, 27 – Cobitis taenia, 28 – Tinca tinca, 29 – Esox lucius, 30 – Gymnocephalus cernua, 31 – Perca fluviatilis, 32 – Acipenser baerii, 33 – Acipenser ruthenus, 34 – Misgurnus fossilis, 35 – Silurus glani, 36 – Sander lucioperca, 37 – Cyprinus carpio, 38 – Rhodeus sericeus, 39 – Acipenser gueldenstaedtii, 40 – Acipenser stellatus, 41 – Abramis ballerus, 42 – Abramis brama, 43 – Alburnus alburnus, 44 – Blicca bjoerkna, 45 – Chondrostoma nasus, 46 – Barbus barbus, 47 – Scardinius erythrophthalmus, 48 – Proterorhinus marmoratus, 49 – Clupeonella cultriventris, 50 – Perccottus glenii, 51 – Ctenopharyngodon idella, 52 – Mylopharyngodon piceus, 53 – Hypophthalmichthys nobilis, 54 – Hypophthalmichthys molitrix.
Факторные нагрузки на основе корреляций
| Переменная | Фактор | |||
|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |
| ОИТ | –0.960 | 0.280 | –0.028 | 0.000 |
| ОЛТ | –0.838 | –0.300 | –0.455 | 0.000 |
| НТ | –0.814 | –0.491 | 0.312 | –0.001 |
| Разность между ОЛТ и ОИТ | 0.380 | –0.770 | –0.512 | –0.000 |
| Разность между ОИТ и НТ | –0.098 | 0.908 | –0.407 | –0.001 |
На рис. 2 наглядно видно, что для комплексов северной группы средние значения ОИТ весьма близки и составляют 15–16°С (межкомплексные различия недостоверны, p > 0.05). Для комплексов южной группы характерно большое разнообразие средних значений ОИТ – 23–27°C (достоверные отличия только для понтического морского комплекса, p < 0.05). В целом различия между ОИТ для видов, входящих в северную и южную группы достоверны (разность 10.5°C, p = 0). Для северной группы средние значения ОЛТ составляют 28–33°C (отмечены достоверные различия между арктобореальным и бореальным предгорным комплексами, p = 0.03), а для южной – 31–40°C (достоверные отличия для понтического морского и китайского равнинного комплексов, p < 0.05). Межгрупповые различия ОЛТ достоверны, но менее выражены (разность 5.9°C, p = 0). Средние значения НТ для комплексов северной группы сравнительно невысоки и составляют интервал 7–14°C (различия недостоверны, p > 0.05), что ниже такового для южной группы – 14–23°C (отмечены достоверные различия между бореальным и китайским равнинными комплексами, p = 0.01). Как и в двух предыдущих случаях, различия между выделяемыми группами достоверны (разность 7.8°C, p = 0). Любопытен тот факт, что только в северной холодолюбивой группе присутствуют виды, у которых нерест проходит в осенне-зимний сезон года.
Рис. 2.
Температурные критерии жизнедеятельности входящих в состав разных фаунистических комплексов рыб и круглоротых: АП – арктический пресноводный, АБ – арктобореальный, БП – бореальный предгорный, БР – бореальный равнинный, ДТ – древний третичный, ПП – понтический пресноводный, ПМ – понтический морской, КР – китайский равнинный. Критерии: 1 – окончательная летальная температура (ОЛТ), 2 – окончательно избираемая температура (ОИТ), 3 – нерестовая температура (НТ). Представлены средние значения ± стандартная ошибка (◻) и стандартное отклонение (I).
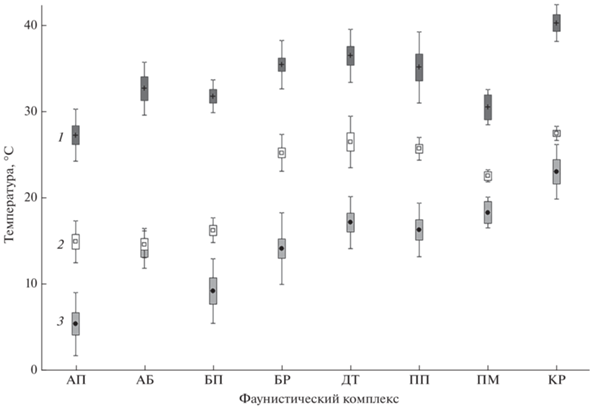
Анализ данных выявил интересную особенность: у большинства видов северной группы верхняя граница термоустойчивости располагается на большем удалении от зоны ОИТ по сравнению с южной группой. Так, для первой группы рыб эта разность составила в среднем 14.6°C, а для второй только 10.2°C (p = 0.00). При этом минимальная разность между верхними значениями ОЛТ и ОИТ характерна для понтического морского, а максимальная – для арктобореального комплексов.
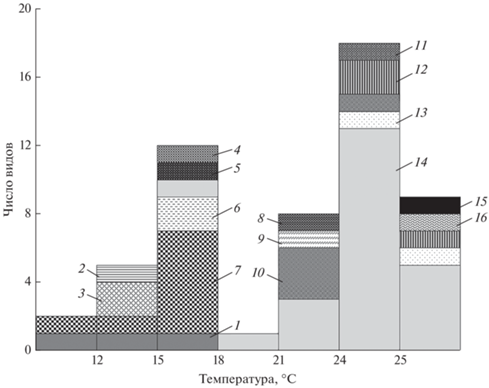
Между значениями ОИТ и ОЛТ отмечена достоверная сильная корреляционная связь (r = 0.8, p = 0). В то же время коэффициенты корреляции между НТ и ОИТ, а также ОЛТ были средними (соответственно r = 0.6 и 0.7 при p = 0). В исследуемой выборке теплолюбивые виды (>63% с ОИТ > 21°С) преобладали над холодолюбивыми (35% с ОИТ < 18°С). С условно промежуточными значениями ОИТ (18–21°С) был представлен только один вид – пескарь Gobio gobio (Linnaeus, 1758), а распределение значений этого критерия отличалось от нормального (p = 0) и носило бимодальный характер (рис. 3). Однако при рассмотрении ОЛТ аналогичные тенденции не выявлены, а распределение данных было близко к нормальному (p = 0.49). То же самое отмечено и для НТ (p = 0.56).
Рис. 3.
Распределение видов рыб исследованных семейств по температурным предпочтениям (окончательная избираемая температура) неполовозрелых особей. 1 – Petromyzontidae, 2 – Lotidae, 3 – Osmeridae, 4 – Cottidae, 5 – Nemacheilidae, 6 – Gasterosteidae, 7 – Salmonidae, 8 – Clupeidae, 9 – Gobiidae, 10 – Acipenseridae, 11 – Esocidae, 12 – Percidae, 13 – Cobitidae, 14 – Cyprinidae, 15 – Odontobutidae, 16 – Siluridae.
ОБСУЖДЕНИЕ
Температурные особенности рыб в свете климатических изменений
Расселение видов за пределы анцестральных зон (нативных ареалов) связано с массой ограничений как внутреннего (особенности физиологии, поведения, питания, размножения и другие), так и внешнего (физические ограничения среды, хищники, межвидовая конкуренция, наличие привычной пищи и другое) происхождения. Однако если ограничить рассмотрение расселения только группой эктотермов, то очевидно, что в новых условиях для подобных организмов доминирующим фактором будет температура. Именно температурные особенности поведения и физиологии у таких видов в подавляющем числе случаев определяют окно возможностей для их расселения и успешной акклиматизации (Bennett et al., 2019).
На протяжении последних десятилетий проявилась отчётливая тенденция по изменению термического режима водоёмов Северного полушария, в том числе центрального и северного регионов европейской части России (Василенко, 2019). Этот процесс хорошо изучен на примере Рыбинского водохранилища (Законнова, Литвинов, 2016) – за оценочный период (1976–2014 гг.) тенденция роста температуры поверхностного слоя воды в течение безлёдного периода составила 0.76°С/10 лет. Это вызвало подвижки в сроках установления и таяния ледового покрова, времени возникновения термоклина и водорослевых вспышек, а также продолжительности нагульного сезона для многих видов рыб (Рыбы Рыбинского …, 2015). Вместе с тем современные температурные значения мест обитания большинства видов рыб довольно далеки от летальных и не могут напрямую привести к их гибели от перегрева. Нельзя недооценивать возможности опосредованного влияния потепления на ихтиоценозы. Например, преждевременное развитие специфической водорослевой флоры, вызванное изменением термического режима водоёма (Сахарова, 2019), может обусловливать депрессию оксифильных видов, в частности ерша Gymnocеphalus cernua (Linnaeus, 1758). Ещё один пример действия температуры в качестве лимитирующего фактора – это снижение численности европейской корюшки (снетка) Osmerus eperlanus (Linnaeus, 1758) в водоёмах верхней Волги в конце XX в. (Карабанов, 2013). Депрессию популяций этого вида отмечали во всех водохранилищах указанного региона независимо от различий гидрологии. Это предполагает воздействие некоего общего негативного фактора и, вероятнее всего, именно смены термического режима в сторону потепления.
Наблюдения за морскими организмами показали, что даже относительно малые климатические сдвиги (увеличение среднего значения температуры воды на 1.13°С) способны негативно отражаться на функционировании их популяций (Pörtner, Knust, 2007). В первую очередь, это обусловлено повышением вероятности выхода значений температуры за некие критические рамки. Кроме того, в температурной нише каждого вида возможно наличие узких мест, весьма чувствительных к смене условий среды. В частности, нерестующие производители и эмбрионы рыб в большей степени уязвимы к воздействию температурного фактора по сравнению с неполовозрелыми и взрослыми особями (Pörtner, 2002; Dahlke et al., 2020; McKenzie et al., 2020). Последнее особенно актуально для короткоцикловых видов, поскольку череда неудачных нерестовых сезонов может приводить к резким изменениям их численности и популяционной структуры. По всей видимости, именно это имело место в популяциях снетка верхней Волги.
Пустующая экологическая ниша снетка в водоёмах верхней Волги спустя небольшой промежуток времени была успешно занята представителем южной группы – черноморско-каспийской тюлькой Clupeonella cultriventris (Nordmann, 1840) (Kiyashko et al., 2012). Зарегулирование р. Волга оказало положительное влияние на численность и возможности расселения этого представителя сельдевых (Карабанов, 2013). Другие примеры благополучия теплолюбивых видов в этом регионе – рост численности серебряного карася Carassius gibelio (Bloch, 1782) (Gerasimov et al., 2018) и линя Tinca tinca (Linnaeus, 1758) (Рыбы Рыбинского …, 2015), а также появление отдельных представителей Gobiidae (Карабанов и др., 2020).
Следует отметить, что успешное расселение отдельных видов рыб (приведённые выше примеры) было бы невозможно без ключевого участия человека. Само по себе зарегулирование рек значительно меняет их гидрологический режим, в том числе и температурную составляющую. Это создаёт дополнительные возможности для расселения одних видов, в первую очередь лимнофильных, а у других (реофилы) приводит к снижению численности и в некоторых случаях к полному исчезновению (Рыбы Рыбинского …, 2015). Кроме того, хозяйственная деятельность человека (рыболовство и рыбоводство, промышленные и сельскохозяйственные) в значительной мере способствует перераспределению ролей в рыбной части существующих сообществ. При этом можно проследить как прямое (вылов представителей ценных промысловых видов, вселение чужеродных для региона объектов аквакультуры), так и опосредованное воздействие через разрушение привычной среды обитания. Вместе с тем пример снетка и тюльки наглядно демонстрирует существование температурных лимитов для северных и южных видов, способных ограничивать их распространение.
Причины консерватизма и изменчивости температурных особенностей рыб
Не секрет, что термоадаптационные способности рыб, изучаемые на организменном уровне, обнаруживают значительную изменчивость, фенотипически и генотипически обусловленную (McKenzie et al., 2020). Однако рассматривая те же виды на уровне популяций, мы выявим признаки большего температурного консерватизма. Особи из удалённых мест, подчас населяющие весьма различные ландшафтно-климатические зоны, при акклимации к стандартным лабораторным условиям демонстрируют близкие значения избираемых и летальных температур. Так, для отдельных видов рыб было показано фактически полное отсутствие межпопуляционных различий (Brown, Feldmeth, 1971; Elliott, Elliott, 1995; Lyytikäinen et al., 1997; Elliott, Klemetsen, 2002; Zakhartsev et al., 2003; Mulhollem et al., 2015), в то время как для других такие различия были зарегистрированы, но были сравнительно невелики (<2°С) (Fields et al., 1987; Konecki et al., 1995; Fangue et al., 2006; Chen et al., 2013). Примечательно, что отсутствие значимой взаимосвязи между верхней границей фундаментальной температурной ниши и текущими условиями обитания в большей степени присуще животным наземным, нежели водным (Sunday et al., 2012; Araújo et al., 2013). Устойчивость к эволюционным изменениям систем с высоким уровнем гомеостаза и отсутствие обязательной прямой взаимосвязи между параметрами окружающей среды и диапазоном толерантности организма уже давно дискутируются (Brown, Feldmeth, 1971).
Чем же обусловлена стабильность температурных черт популяций, нередко испытывающих весьма различное климатическое воздействие? На этот вопрос в настоящее время нет однозначного ответа. Вполне вероятно присутствие некоторых ограничений и компромиссов, препятствующих действию естественного отбора. При этом скорость и выраженность эволюционных изменений в ответ на смену температурных условий могут различаться как для отдельных групп животных, так и для разных физиологических процессов, вовлечённых в жизнедеятельность и приспособляемость организмов (Angilletta et al., 2002). Существует ряд работ, доказывающих наследуемость термотолерантности у рыб, что могло бы служить полем действия для естественного отбора, однако устойчивость к острому тепловому воздействию далеко не всегда напрямую определяет приспособляемость организма к условиям обитания (McKenzie et al., 2020). Предполагается, что температурный консерватизм в разной степени выражен у отдельных таксонов и в целом присущ высокомобильным, широко распространённым видам с интенсивным межпопуляционным взаимодействием (Bennett et al., 2019). В то же время малоподвижные виды с сильным уровнем генетической изоляции популяций будут демонстрировать бόльшую изменчивость температурных показателей в зависимости от адаптированности к конкретным условиям обитания. В геологическом масштабе эволюционные процессы, протекающие на фоне резких изменений климата (например, в Северной Европе), способствовали формированию видов с более широкими температурными нишами (виды-генералисты), что также обусловливает снижение их внутривидовой (межпопуляционной) изменчивости (Bennett et al., 2019).
Другая возможность для ослабления воздействия отбора на температурные черты организмов – это терморегуляционное поведение. Для подавляющего большинства естественных водоёмов на протяжении большей части года характерна гетеротермия. Температурная неоднородность среды обитания позволяет водным организмам и, в частности рыбам, активно выбирать наиболее комфортные зоны. При этом в их поведении проявляется определённая миграционная ритмика (суточная, сезонная и другие), чаще всего объясняемая компромиссом между температурными и трофическими условиями (Brett, 1971; Clark, Levy, 1988; Clark, Green, 1991; Clough, Ladle, 1997; Garner et al., 1998; Mehner, 2012; Armstrong et al., 2013). В таких случаях миграционное поведение у холодолюбивых и теплолюбивых видов, как правило, выглядит зеркально – первые значительную часть суток проводят в прохладной воде и лишь ненадолго заплывают для питания в тёплые, богатые пищей зоны, а вторые действуют противоположным образом. Вместе с тем в лабораторных термоградиентных условиях при достаточной обеспеченности кормовыми ресурсами и в отсутствие хищников рыбы устойчиво скапливаются в зоне ОИТ, снижая двигательную активность, что напрямую отражается на приростах (Смирнов, Смирнова, 2019). Это позволяет расценивать феномен суточных миграций как ответную реакцию на отклонение абиотических (температура, кислород и так далее) и/или биотических (кормовые объекты, пресс хищников, конкуренция и так далее) условий от оптимума.
Терморегуляционное поведение можно рассматривать как самую быструю ответную реакцию организма на изменения, происходящие в среде обитания (Слоним, 1971; Bicego et al., 2007). В тех случаях, когда поведенческого ответа на воздействие температурного фактора недостаточно либо он ограничен иными факторами, подключается эффективный набор физиолого-биохимических механизмов (особенности акклимации, изменения митохондриальной и мембраной структуры клеток, синтез изоформ ферментов, выработка белков теплового шока и так далее) (Озернюк, 2000). Примечательно, что в краевых популяциях отдельных видов рыб (на юге для холодолюбивых, а на севере для теплолюбивых) в условиях, когда невозможно избежать воздействия экстремальных температур, как правило, связанных также и с дефицитом кислорода, наблюдаются процессы оцепенения особей. Оцепенение наступает в течение зимнего (например, у карася, сазана Cyprinus carpio Linnaeus, 1758, линя, ротан-головешки Perccottus glenii Dybowski, 1877, вьюна Misgurnus fossilis (Linnaeus, 1758)) или летнего сезонов (у налима Lota lota (Linnaeus, 1758)) года.
Таким образом, внутривидовые значения температурных компонентов фундаментальной ниши весьма стабильны во времени. В этой связи избираемая (предпочитаемая) температура как интегральный показатель температурных оптимумов для огромного количества одновременно протекающих физиологических процессов может в большей степени отражать исторически сложившиеся температурные характеристики вида (температурную предысторию вида), нежели его адаптацию к текущим условиям обитания (Angilletta et al., 2002). Ранее к схожему выводу о термопреферендуме как о температуре района видообразования (зоны происхождения) привёл анализ температурных предпочтений клещей (Acari) и насекомых (Insecta) (Кауфман, 1985). Следовательно, температурные черты вида свидетельствуют не только о его филогенетических особенностях, но и об экологической специфике условий возникновения. В чём-то это согласуется с теорией об анцестральных экологических зонах происхождения (Ricklefs, 2006). Согласно ей виды лучше приспособлены к параметрам среды, существовавшей в период возникновения данной филогенетической группы, тогда как переход в новые экологические зоны потребовал бы от них значительных эволюционных изменений. Другими словами, при расселении виды сохраняют некие базовые адаптационные настройки, предпочитая в новой среде находить элементы прежней (выбор локаций с приближенными к анцестральным условиями, активность в течение небольшой части года или суток, миграции). Теоретически такой нишевый консерватизм мог бы помочь при прогнозировании успешности акклиматизации потенциальных видов-вселенцев в тех или иных местообитаниях. Вместе с тем изучение температурных компонентов фундаментальной ниши, весьма вероятно, способно хотя бы приближённо ответить на вопрос о первичности тех или иных ландшафтно-климатических условий для видов с обширными ареалами.
Фаунистические комплексы и температурные особенности рыб
Анализ трёх важнейших критериев видовой фундаментальной температурной ниши (оптимум, лимиты, нерестовая температура) не выявил их чёткого соответствия выделяемым фаунистическим комплексам. Это определялось прежде всего значительной неоднородностью температурных показателей для рыб, объединённых в состав единого комплекса. Например, для бореального равнинного фаунистического комплекса нетипично теплолюбивыми и теплоустойчивыми видами-исключениями являются щиповка Cobitis taenia Linnaeus, 1758 и серебряный карась, для древнего третичного – сом Silurus glanis Linnaeus, 1758 и сазан. Вместе с тем средние значения ОИТ и ОЛТ для видов бореального равнинного, древнего третичного и понтического пресноводного фаунистических комплексов оказались весьма близки (рис. 2). Ранее Яковлев (1964) высказал аргументированное представление о формировании в неогене на обширной территории Палеоарктики (от Западной Европы до Дальнего Востока) к северу от 44° с.ш. единого фаунистического комплекса, сходного по составу с тремя вышеперечисленными. За ним предлагалось сохранить общее название “бореальный равнинный”. Условия, в которых протекал процесс становления комплекса, существенно отличались от современных и соответствовали относительно выровненному умеренно-тёплому и достаточно влажному климату (Яковлев, 1964). Такая точка зрения хорошо объясняет сходную теплолюбивость рыб бореального равнинного, древнего третичного и понтического пресноводного комплексов.
Выполненный математический анализ выявил в пресноводной ихтиофауне европейской части России наличие двух мощных пулов – условно холодолюбивых северных и теплолюбивых южных видов. Выделенные группы хорошо различимы по набору температурных черт и, в первую очередь, по ОИТ (рис. 1). Интересен тот факт, что группы рыб, отнесённые к холодолюбивым и теплолюбивым, с точки зрения систематики не пересекаются даже на уровне семейств (исключение – обыкновенный гольян). Это согласуется с выводами о существовании отчётливого филогенетического сигнала в отношении температурных свойств (например, теплоустойчивости) рыб на уровне семейств, указывающего на общность температурного прошлого для входящих в них видов (McKenzie et al., 2020).
Виды, формирующие выделенные группы, достоверно различались по относительному запасу теплоустойчивости, то есть разности между ОИТ и ОЛТ (рис. 2). Ранее уже обращали внимание на тот факт, что тропические виды обладают меньшим запасом теплоустойчивости по сравнению с видами умеренных широт, поскольку обитают в условиях более высоких температур (оптимум), близких к летальной границе (McKenzie et al., 2020). Создаётся впечатление, что жизнедеятельность обитателей тропиков, а также выделенной нами южной группы ограничена неким общим температурным порогом толерантности. Действительно, устойчивость многоклеточных организмов к воздействию тепла, как правило, не превышает значений, близких к 45°C, что обусловлено резким нарастанием дисбаланса между потреблением кислорода и его поступлением в ткани тела (Pörtner, 2002). Интересно, что даже у отдельных видов карпозубых (Cyprinodon), населяющих экстремальные местообитания (горячие источники Долины Смерти с температурой воды до 44°С), термотолерантность сопоставима с таковой для многих других теплолюбивых видов (Brown, Feldmeth, 1971).
Среди исследованных в настоящей работе рыб обнаружен всего один вид, для которого значение ОИТ занимает условно среднее положение (рис. 3). В геологическом аспекте это можно объяснить значительной изменчивостью ландшафтно-климатических условий умеренного пояса европейской части России, вызванной регулярными оледенениями. По мнению палеонтологов (Яковлев, 1964), в периоды наступления ледников часть теплолюбивых видов рыб могла успешно сохраняться в не затрагиваемых оледенениями пресноводных рефугиумах, располагавшихся на юге Европейского континента. Впоследствии, после таяния льдов, именно оттуда могло осуществляться повторное расселение рыб по водоёмам средних широт. Заселение же пресных вод рассматриваемого региона холодолюбивыми видами происходило за счёт северной морской и пресноводной горной ихтиофаун (Яковлев, 1964; Рыбы Монгольской …, 1983), что и определило гораздо более низкие значения их температурных предпочтений.
Данные о рыбах и круглоротых европейской части России перекликаются с информацией о рыбах других географических областей. Так, предпочитаемые температурные значения рыб Северной Америки весьма схожим образом (бимодально) распределены по температурной шкале (Magnuson et al., 1979). Тем не менее авторы цитируемой работы выделили три условные группы по отношению к температурному фактору: холодноводные (предпочитаемая температурная зона 11.0–14.9°С), умеренно холодноводные (21.0–24.9°С) и тепловодные (27.0–30.9°С). Примечательно, что на графике, приведённом авторами, линии, показывающие две последние группы, фактически перекрываются, образуя единую группу (аналог выделенной нами южной). В этой работе также отмечена довольно строгая, за редким исключением (Cyprinidae), принадлежность разных семейств рыб к обозначенным группам (Magnuson et al., 1979). По всей видимости, параллели между европейскими и североамериканскими видами не случайны и свидетельствуют о схожести процессов формирования ихтиофауны на территории Голарктики в целом.
В настоящее время в европейской части России встречается немало видов-вселенцев из состава китайского равнинного фаунистического комплекса (Karabanov et al., 2022). Проанализированные в нашей работе представители этого комплекса (пёстрый Hypophthalmichthys nobilis (Richardson, 1845) и белый H. molitrix (Valenciennes, 1844) толстолобики, белый Ctenopharyngodon idella (Valenciennes, 1844) и чёрный Mylopharyngodon piceus (Richardson, 1846) амуры и ротан-головешка) составили наиболее теплолюбивую часть южной группы (Приложение) . При этом встречаемость толстолобиков и амуров ограничена водоёмами юга России (до 55° с.ш.) либо прудами-охладителями ГРЭС, ТЭЦ и АЭС, где в течение всего года температура воды значительно выше естественной. Это ещё один пример воздействия температуры как фактора, ограничивающего расселение. Вместе с тем ротан как обладатель одной из самых широких температурных ниш (Голованов и др., 2013) успешно заселил водоёмы высоких широт, в частности бассейн Белого моря (Рыбы …, 2010).
ВЫВОДЫ
1. В пресноводной ихтиофауне европейской части России можно выделить две обособленные группы: северную холодолюбивую (ОИТ 12–18°С) и южную теплолюбивую (ОИТ 22–31°С). При этом характерно почти полное отсутствие видов с ОИТ в диапазоне 18–21°С.
2. Температурные критерии жизнедеятельности рыб (оптимум, лимиты и нерестовая температура) не могут быть использованы в качестве чёткого признака принадлежности видов к тому или иному фаунистическому комплексу.
3. Климатическая тенденция роста температуры воды, наметившаяся в водоёмах умеренных широт, по всей видимости, благоприятствует распространению на север представителей теплолюбивой южной группы видов, что может привести к дальнейшей перестройке конкурентных отношений в популяциях рыб и вызвать изменения границ их ареалов.
Список литературы
Айвазян С.А., Бухштабер В.М., Енюков И.С., Мешалкин Л.Д. 1989. Прикладная статистика. Классификация и снижение размерности. М.: Финансы и статистика, 607 с.
Василенко А.Н. 2019. Современные представления о термическом режиме рек и его трансформациях в условиях изменяющегося климата // Матер. Междунар. науч.-практ. конф. “Глобальные климатические изменения: региональные эффекты, модели, прогнозы”. Т. 1. Воронеж: Цифровая полиграфия. С. 359–363.
Герасимов Ю.В., Смирнов А.К., Кодухова Ю.В. 2018. Оценка возможных причин изменений численности и половой структуры в популяциях серебряного карася (Carassius auratus gibelio Bloch., 1783) // Биология внутр. вод. № 1. С. 70–79. https://doi.org/10.7868/S0320965218010096
Голованов В.К. 2013. Температурные критерии жизнедеятельности пресноводных рыб. М.: Полиграф-Плюс, 300 с.
Голованов В.К., Капшай Д.С., Герасимов Ю.В. и др. 2013. Термоизбирание и термоустойчивость молоди ротана-головешки Perccottus glenii в осенний сезон // Вопр. ихтиологии. Т. 53. № 2. С. 246–250. https://doi.org/10.7868/S0042875213020033
Законнова А.В., Литвинов А.С. 2016. Многолетние изменения гидроклиматического режима Рыбинского водохранилища // Тр. ИБВВ РАН. Вып. 75 (78). С. 16–22. https://doi.org/10.24411/0320-3557-2016-10016
Карабанов Д.П. 2013. Генетические адаптации черноморско-каспийской тюльки Clupeonella cultriventris (Nordmann, 1840) (Actinopterygii: Clupeidae). Воронеж: Науч. книга, 179 с.
Карабанов Д.П., Павлов Д.Д., Никитин Э.В. и др. 2020. Анализ видового состава, проблемы идентификации и путей расселения чужеродных видов рыб в бассейне реки Волги // Вестн. АГТУ. Сер. Рыбн. хоз-во. № 3. С. 7–17. https://doi.org/10.24143/2073-5529-2020-3-7-17
Кауфман Б.З. 1985. Возможное эволюционное значение реакции термопреферендума пойкилотермных животных // Журн. общ. биологии. Т. 46. № 4. С. 509–515.
Кияшко В.И., Карабанов Д.П., Яковлев В.Н. и др. 2012. Становление и развитие популяции Черноморско-Каспийской тюльки Clupeonella cultriventris (Clupeidae) в Рыбинском водохранилище // Вопр. ихтиологии. Т. 52. № 5. С. 571–580.
Никольский Г.В. 1956. Рыбы бассейна Амура. Итоги Амурской ихтиологической экспедиции 1945–1949 гг. М.: Изд-во АН СССР, 551 с.
Никольский Г.В. 1980. Структура вида и закономерности изменчивости рыб. М.: Пищ. пром-сть, 183 с.
Озернюк Н.Д. 2000. Температурные адаптации. М.: Изд-во МГУ, 205 с.
Решетников Ю.С. 1981. Идеи Г.В. Никольского о фаунистических комплексах и их современное развитие // Современные проблемы ихтиологии. М.: Наука. С. 75–95.
Рыбы в заповедниках России. 2010. Т. 1. М.: Т-во науч. изд. КМК, 627 с.
Рыбы Монгольской Народной Республики. 1983. М.: Наука, 278 с.
Рыбы Рыбинского водохранилища: популяционная динамика и экология. 2015. Ярославль: Филигрань, 418 с.
Сахарова Е.Г. 2019. Фитопланктон озера Плещеево в 2014–2016 гг. // Тр. ИБВВ РАН. Вып. 86 (89). С. 23–33. https://doi.org/10.24411/0320-3557-2019-10009
Слоним А.Д. 1971. Экологическая физиология животных. М.: Высш. школа, 448 с.
Смирнов А.К., Смирнова Е.С. 2019. Поведение молоди окуня Perca fluviatilis (Percidae) в гетеротермальной среде при разной обеспеченности пищей // Зоол. журн. Т. 98. № 2. С. 182–192. https://doi.org/10.1134/S0044513419020168
Яковлев В.Н. 1964. История формирования фаунистических комплексов пресноводных рыб // Вопр. ихтиологии. Т. 4. Вып. 1 (30). С. 10–22.
Angilletta M.J., Niewiarowski P.H., Navas C.A. 2002. The evolution of thermal physiology in ectotherms // J. Therm. Biol. V. 27. № 4. P. 249–268. https://doi.org/10.1016/S0306-4565(01)00094-8
Araújo M.B., Ferri-Yáñez F., Bozinovic F. et al. 2013. Heat freezes niche evolution // Ecol. Letters. V. 16. № 9. P. 1206–1219. https://doi.org/10.1111/ele.12155
Armstrong J.B., Schindler D.E., Ruff C.P. et al. 2013. Diel horizontal migration in streams // Ecology. V. 94. № 9. P. 2066–2075. https://doi.org/10.1890/12-1200.1
Beitinger T.L., Bennett W.A., McCauley R.W. 2000. Temperature tolerances of North American freshwater fishes exposed to dynamic changes in temperature // Environ. Biol. Fish. V. 58. № 3. P. 237–275. https://doi.org/10.1023/A:1007676325825
Bennett S., Duarte C.M., Marbà N., Wernberg T. 2019. Integrating within-species variation in thermal physiology into climate change ecology // Phil. Trans. R. Soc. B. V. 374. № 1778. Article 20180550. https://doi.org/10.1098/rstb.2018.0550
Bicego K.C., Barros R.C.H., Branco L.G.S. 2007. Physiology of temperature regulation // Comp. Biochem. Physiol. A Mol. Integr. Physiol. V. 147. № 3. P. 616–639. https://doi.org/10.1016/j.cbpa.2006.06.032
Brett J.R. 1971. Energetic responses of salmon to temperature. A study of some thermal relations in the physiology and freshwater ecology of sockeye salmon (Oncorhynchus nerka) // Am. Zool. V. 11. № 1. P. 99–113. https://doi.org/10.1093/icb/11.1.99
Brown J.H., Feldmeth C.R. 1971. Evolution in constant and fluctuating environments // Evolution. V. 25. № 2. P. 390–398. https://doi.org/10.1111/j.1558-5646.1971.tb01893.x
Chen Z., Anttila K., Wu J. et al. 2013. Optimum and maximum temperatures of sockeye salmon (Oncorhynchus nerka) populations hatched at different temperatures // Can. J. Zool. V. 91. № 5. P. 265–274. https://doi.org/10.1139/cjz-2012-0300
Clark C.W., Levy D.A. 1988. Diel vertical migrations by juvenile sockeye salmon and the antipredation window // Am. Nat. V. 131. № 2. P. 271–290.
Clark D.S., Green J.M. 1991. Seasonal variation in temperature preference of juvenile Atlantic cod (Gadus morhua), with evidence supporting an energetic basis for their diel vertical migration // Can. J. Zool. V. 69. № 5. P. 1302–1307. https://doi.org/10.1139/z91-183
Clough S., Ladle M. 1997. Diel migration and site fidelity in a stream-dwelling cyprinid, Leuciscus leuciscus // J. Fish Biol. V. 50. № 5. P. 1117–1119. https://doi.org/10.1111/j.1095-8649.1997.tb01635.x
Cooper B.S., Williams B.H., Angilletta M.J. 2008. Unifying indices of heat tolerance in ectotherms // J. Therm. Biol. V. 33. № 6. P. 320–323. https://doi.org/10.1016/j.jtherbio.2008.04.001
Dahlke F.T., Wohlrab S., Butzin M., Pörtner H.O. 2020. Thermal bottlenecks in the life cycle define climate vulnerability of fish // Science. V. 369. № 6499. P. 65–70. https://doi.org/10.1126/science.aaz3658
Elliott J.M., Elliott J.A. 1995. The critical thermal limits for the bullhead, Cottus gobio, from three populations in north-west England // Freshw. Biol. V. 33. № 3. P. 411–418. https://doi.org/10.1111/j.1365-2427.1995.tb00403.x
Elliott J.M., Klemetsen A. 2002. The upper critical thermal limits for alevins of Arctic charr from a Norwegian lake north of the Arctic circle // J. Fish Biol. V. 60. № 5. P. 1338–1341. https://doi.org/10.1006/jfbi.2002.1934
Fangue N.A., Hofmeister M., Schulte P.M. 2006. Intraspecific variation in thermal tolerance and heat shock protein gene expression in common killifish, Fundulus heteroclitus // J. Exp. Biol. V. 209. № 15. P. 2859–2872. https://doi.org/10.1242/jeb.02260
Fields R., Lowe S.S., Kaminski C. et al. 1987. Critical and chronic thermal maxima of northern and Florida largemouth bass and their reciprocal F1 and F2 hybrids // Trans. Am. Fish. Soc. V. 116. № 6. P. 856–863. https://doi.org/10.1577/1548-8659(1987)116<856:CACTMO>2.0.CO;2
Fry F.E.J. 1947. Effects of the environment on animal activity // Univ. Toronto Stud. Biol. Ser. № 55. (Publ. Ontario Fish. Res. Lab. № 68). P. 1–62.
Garner P., Clough S., Griffiths S.W. et al. 1998. Use of shallow marginal habitat by Phoxinus phoxinus: a trade-off between temperature and food? // J. Fish Biol. V. 52. № 3. P. 600–609. https://doi.org/10.1111/j.1095-8649.1998.tb02020.x
Jobling M. 1981. Temperature tolerance and the final preferendum—rapid methods for the assessment of optimum growth temperatures // J. Fish Biol. V. 19. № 4. P. 439–455. https://doi.org/10.1111/j.1095-8649.1981.tb05847.x
Karabanov D.P., Bekker E.I., Pavlov D.D. et al. 2022. New sets of primers for DNA identification of non-indigenous fish species in the Volga-Kama basin (European Russia) // Water. V. 14. № 3. Article 437. https://doi.org/10.3390/w14030437
Konecki J.T., Woody C.A., Quinn T.P. 1995. Temperature preference in two populations of juvenile coho salmon, Oncorhynchus kisutch // Environ. Biol. Fish. V. 44. № 4. P. 417–421. https://doi.org/10.1007/BF00008256
Lyytikäinen T., Koskela J., Rissanen I. 1997. Thermal resistance and upper lethal temperatures of underyearling Lake Inari Arctic charr // J. Fish Biol. V. 51. № 3. P. 515–525. https://doi.org/10.1111/j.1095-8649.1997.tb01509.x
Magnuson J.J., Crowder L.B., Medvick P.A. 1979. Temperature as an ecological resource // Am. Zool. V. 19. № 1. P. 331–343. https://doi.org/10.1093/icb/19.1.331
McKenzie D.J., Zhang Y., Eliason E.J. et al. 2020. Intraspecific variation in tolerance of warming in fishes // J. Fish Biol. V. 98. № 6. P. 1536–1555. https://doi.org/10.1111/jfb.14620
Mehner T. 2012. Diel vertical migration of freshwater fishes – proximate triggers, ultimate causes and research perspectives // Freshw. Biol. V. 57. № 7. P. 1342–1359. https://doi.org/10.1111/j.1365-2427.2012.02811.x
Mulhollem J.J., Suski C.D., Wahl D.H. 2015. Response of largemouth bass (Micropterus salmoides) from different thermal environments to increased water temperature // Fish Physiol. Biochem. V. 41. № 4. P. 833–842. https://doi.org/10.1007/s10695-015-0050-0
Pörtner H.O. 2002. Climate variations and the physiological basis of temperature dependent biogeography // Comp. Biochem. Physiol. A Mol. Integr. Physiol. V. 132. № 4. P. 739–761. https://doi.org/10.1016/S1095-6433(02)00045-4
Pörtner H.O., Knust R. 2007. Climate change affects marine fishes through the oxygen limitation of thermal tolerance // Science. V. 315. № 5808. P. 95–97. https://doi.org/10.1126/science.1135471
Ricklefs R.E. 2006. Evolutionary diversification and the origin of the diversity–environment relationship // Ecology. V. 87. № sp7. P. S3–S13. https://doi.org/10.1890/0012-9658(2006)87[3:EDATOO]-2.0.CO;2
Sokal R.R., Rohlf F.J. 1995. Biometry. New York: W.H. Freeman, 899 p.
Sprent P., Smeeton N.C. 2007. Applied Nonparametric Statistical Methods. Boca Raton: Chapman and Hall; CRC, 530 p. https://doi.org/10.1201/b15842
Sunday J.M., Bates A.E., Dulvy N.K. 2012. Thermal tolerance and the global redistribution of animals // Nat. Clim. Change. V. 2. № 9. P. 686–690. https://doi.org/10.1038/nclimate1539
Zakhartsev M.V., Wachter B., Sartoris F.J. et al. 2003. Thermal physiology of the common eelpout (Zoarces viviparus) // J. Comp. Physiol. B. V. 173. № 5. P. 365–378. https://doi.org/10.1007/s00360-003-0342-z
Дополнительные материалы
- скачать ESM.docx
- Приложение 1.
Таблица S1. Температурные критерии жизнедеятельности пресноводных рыб и миног европейской части России, входящих в состав отдельных фаунистических комплексов
Список литературы
Инструменты
Вопросы ихтиологии