

Известия РАН. Серия биологическая, 2019, № 6, стр. 598-606
Роль спящих почек в трансформации травянистых биоморф (на примере видов рода Nepeta L. sect. Spicatae (Benth.) Pojark.)
А. Ю. Асташенков 1, *, В. А. Черемушкина 1
1 Центральный сибирский ботанический сад СО РАН
630090 Новосибирск, ул. Золотодолинская, 101, Россия
* E-mail: astal@bk.ru
Поступила в редакцию 27.04.2018
После доработки 19.09.2018
Принята к публикации 24.09.2018
Аннотация
На примере ряда видов рода Nepeta определена роль спящих почек в формировании и трансформации травянистых жизненных форм растений. Описаны различные варианты побегообразования и механизмы перестроек биоморф с участием спящих почек. Установлено, что регулярное и нерегулярное развитие спящих почек может привести к формированию укороченных, раздвинутоузловых и удлиненных резидов – основы построения новых побеговых структур (побеговые системы, укореняющиеся парциальные образования), характерных для длиннокорневищных биоморф.
Начиная с XIX в. в литературе впервые упоминаются спящие почки (Hartig, 1851; Büsgen, 1897). Сведения о них касаются исключительно древесных форм: их строения, продолжительности жизни и роста, а также расположения их на побегах и стволах. Вызванный к ним научный интерес привел к дальнейшему изучению спящих почек у различных пород деревьев, их анатомии и происхождения (Любименко, 1900; Jost, 1925; и др.), возможности и способов ветвления (Chandler, 1925; Aaron, 1946; Стешенко, 1956; и др.), физиологии “сна” и “пробуждения” (Goebel, 1928; Зёдинг, 1955). С помощью большой группы тропических растений подробно изучена каулифлория, при которой цветки и соцветия развиваются из спящих почек (Esser, 1887; Velenovsky, 1905; Haberlandt, 1926; и др). В середине ХХ в. было опубликовано много работ, авторы которых исследовали свойства спящих почек (Кожевников, 1950; Baldini, Mosse, 1956; Ляшенко, 1958, 1964; и др.). Сегодня определена роль спящих почек в становлении общей структуры тела древесного растения (Серебряков, 1955; Мазуренко, Хохряков, 1977; Мазуренко, 1986; Sabatier, Barthélémy, 2001; и др.). При описании общей архитектуры кроны деревьев и кустарников нередко оси возобновления дифференцируют по положению и характеру их возникновения, часть этих осей связана с реализацией спящих почек (Millet et al., 1998; Khutornoi et al., 2001; Nikolini et al., 2001; Kostina et al., 2015; Getmanets, 2015; и др.). Функциональная активность спящих почек в формировании общего габитуса растений может быть использована при создании имитационных моделей роста растений (Měch, Prusinkiewicz, 1996). В эволюции жизненных форм покрытосеменных растений и их филогенеза переходные формы от древесных к полудревесным рассматривали через сокращение роста главной лидирующей оси и функционирование спящих почек в базальной части растения (Серебряков, 1954).
Таким образом, в морфологии растений в течение продолжительного времени разными исследователями уделялось особое внимание биологии и функционированию спящих почек исключительно у деревьев. В литературе о роли спящих почек в жизни травянистых растений имеются только отрывочные или косвенные сведения (Серебряков, 1952; Серебрякова, 1983; и др.). Обычно раскрытие спящих почек у этой группы растений оценивают как один из механизмов поддержания жизнеспособности особей на завершающих этапах онтогенеза (Борисова, 1961; Смирнова и др., 1984; Нухимовский, 1997; Черемушкина, 2004; Османова, 2007; и др.). Роль спящих почек в образовании структурных элементов, приводящих к трансформации и эволюционным перестройкам травянистых биоморф, в настоящее время практически никем не определяется.
Цель исследования – на примере ряда видов показать роль спящих почек, реализация которых на разных этапах онтогенеза приводит к становлению и перестройке жизненной формы.
МАТЕРИАЛЫ И МЕТОДЫ
В качестве модельных объектов исследования выбраны некоторые среднеазиатские виды секции Spicatae (Benth.) Pojark. рода Nepeta L. (Lamiaceae): Nepeta mariae Regel, N. pulchella Pojark., N. schugnanica Lipsk., N. transiliensis Pojark. Выбор объектов основан на их филогенетическом родстве и практически полном отсутствии данных об их биологии и развитии. Предложенные виды отражают многообразие механизмов морфологической адаптации особей к высокогорным условиям Центральной Азии, а изучение способов трансформации биоморф в изменяющихся географических и эколого-ценотических условиях дает возможность описать основополагающие эволюционно-морфологические преобразования и предложить основные пути соматической эволюции биоморф, и в частности в роде Nepeta.
N. mariae – высокогорный эндемик среднеазиатских гор. Он распространен на хребтах широтного направления северно-западной части Памиро-Алая и западной части Тянь-Шаня. В условиях Памиро-Алая особи вида произрастают на Туркестанском и Зеравшанском хребтах в поясе арчовников и степей. В условиях Тянь-Шаня на хребтах Ферганском, Чаткальском, Пскемском, Таласском N. mariae распространена от верхней границы лесного до субальпийского и альпийского поясов. В условиях субальпийского пояса вид встречается на влажных лужайках, по берегам рек, нередко на каменистых россыпях (Пояркова, 1954). Материал собран на Зеравшанском хребте в ущелье р. Вору (Таджикистан) в ксерофитных условиях на мелкозёмистом склоне в степном поясе гор (39°13′31.1′′ с.ш., 67°56′16.4′′ в.д., 3016 м над уровнем моря (у. м.)).
N. pulchella – эндемик, растущий в западной части Таласского Алатау. Особи вида произрастают в субальпийском поясе и в верхней полосе лесного пояса в луговых ценозах (Пояркова, 1953). Наши исследования в бассейне р. Жабаглы показали, что N. pulchella предпочитает расселение на экспозиционных вариантах северного макросклона Таласского хребта. Верхняя граница распространения проходит по субальпийскому поясу осевой части хребта. Наибольшее обилие достигается в разнотравно-злаковых субальпийских лугах. При цветении N. pulchella создается синий аспект по бортам рукавов временных водотоков и микропонижениям. Нижняя граница обитания проходит по луговому среднегорью, где особи приурочены к открытым участкам увалистого мезорельефа, принимая незначительное участие в сложении лугово-степных ценозов (Astashenkov еt al., 2017). Материал собран на территории Аксу-Жабаглинского заповедника в ксеромезофитных условиях субальпийского пояса (42°23′45.0′′ с.ш., 70°37′43.8′′ в.д., 2300 м над у. м.).
N. transiliensis – высокогорный эндемик Тянь-Шаня, растущий только в Заилийском Алатау (Казахстан) и в восточных областях Синцзяня (Китай). Особи произрастают в составе изреженных растительных группировок на каменистых склонах гор, встречаются по берегам рек в луговых сообществах (Пояркова, 1954; Буданцев, 1990). Материал собран в альпийском поясе в мезофитных условиях на каменистом русле притока р. Тургень на хр. Карач (Заилийский Алатау) (46°16′43.6′′ с.ш., 77°52′08.1′′ в.д., 2700 м над у. м.).
N. schugnanica – узколокальный эндемик Памира. Сегодня известно единственное место его произрастания – окр. пос. Нишусп (Западный Памир) (Кочкарева, 1986). Материал собран в мезофитных условиях в нижней части подвижного крупнообломочного осыпного склона, примыкающего к урезу воды основного русла р. Бинджюс (37°17′51.2′′ с.ш., 71°30′21.0′′ в.д., 2360 м над у. м.).
Основные методы, использованные в работе, – онтогенетический и сравнительно-морфологический. Они основаны на изучении биоморфы в процессе ее становления в ходе реализации онтогенеза особей (Meusel, 1953; Серебряков, 1962; Смирнова и др., 1976; Серебрякова, 1980).
При описании жизненных форм особей модельных видов использован эколого-морфологический подход Варминга–Серебрякова (Серебряков, 1954, 1962), который в настоящее время включает в себя комплексный анализ, основанный на изучении структурных элементов биоморфы (Серебрякова, 1977; Шафранова, Гатцук, 1994) и способов их сочленения в теле растений (Hallé, Oldeman, 1970; Hallé et al., 1978; Марфенин, 1999; Нотов, 1999; Barthélémy, Caraglio, 2007). Тип биоморфы как естественная единица, воздействующая на среду обитания и имеющая элементарный источник фитогенного поля, выделен по характеру размещения структурных частей и степени их автономности (моноцентрическая, неявнополицентрическая, явнополицентрическая) (Смирнова, 1987). При изучении онтоморфогенеза использованы концепция дискретного описания онтогенеза, впервые предложенная в России (Работнов, 1950; Уранов, 1975), и представления о его поливариантности (Жукова, 1995) и дополненные нами (Черемушкина, 2012). Календарный возраст особей установлен с помощью прямого подсчета годичных колец на анатомическом срезе каудекса, а также по числу сохранившихся ежегодных резидов (термин Е.Л. Нухимовского) (Нухимовский, 1997) на каудексе и корневище. При описании побегообразования у модельных видов использовались теоретические разработки по архитектуре трав (Серебрякова, 1977; Серебрякова, Петухова, 1978; Савиных, 2012), основанные на представлениях об архитектурных моделях деревьев (Hallé, Oldeman, 1970; Hallé et al., 1978). Ход соматической эволюции применительно к высшим растениям, связанный с изменением их жизненных форм, рассмотрен с учетом мнений многих исследователей (Серебряков, 1955; Хохряков, 1981; Серебрякова, 1983; Рing Li, Dohnston, 2000).
Под спящей почкой мы понимаем пазушную или придаточную почку, находящуюся в состоянии относительного покоя на протяжении нескольких лет, которая после пробуждения формирует моноподиальный побег.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
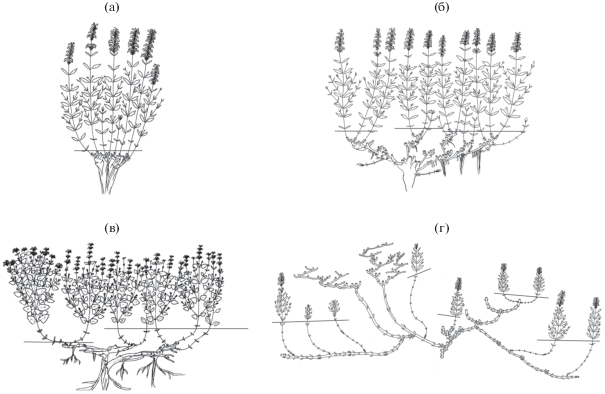
Модельные среднеазиатские виды Nepeta относятся к многолетним поликарпическим травам, развивающимся по симподиальной длиннопобеговой модели побегообразования. N. mariae – типичный стержнекорневой каудексовый многолетник. В течение всего онтогенеза у растений сохраняется моноцентрическая биоморфа (рис. 1а) (Асташенков, Черемушкина, 2016).
Рис. 1.
Основные типы биоморф модельных видов. а – стержнекорневая (модель N. mariae), б – короткокорневищно-стержнекорневая (N. pulchella), в – длиннокорневищно-стержнекорневая (N. schugnanica), г – длиннокорневищная (фрагмент клона, модель N. transiliensis).
Начиная с проростка и до виргинильного состояния, побегообразование осуществляется за счет функционирования почки возобновления, которая закладывается в базальной части ортотропного материнского побега в узле 2–3-го метамера (рис. 2a–г). Нижележащие почки не реализуются и становятся спящими. На 4–5-й год, переходя в виргинильное состояние, особи начинают ветвиться. Куст формируется за счет одновременного развертывания одной почки регулярного возобновления и одной спящей почки, причем расти начинает спящая почка резида позапрошлогоднего прироста (рис. 2д). В редких случаях реализуется почка, сохранившаяся на резидах первых годичных приростов. Начиная с этого состояния и до конца жизни растения, в побегообразовании помимо почки возобновления всегда участвует спящая почка. У растений, находящихся в генеративном состоянии, многолетняя основа куста образована ортотропными близко расположенными базальными частями побеговой системы. Как правило, первыми зацветают побеги, развернувшиеся из почек регулярного возобновления, а побеги, развившиеся из спящих почек, зацветают позже или остаются скрытогенеративными или вегетативными (рис. 2е). Такое развитие побегов приводит к растянутому по времени цветению особей в течение всего вегетационного сезона. Подобное нарастание и ветвление отмечено, например, у особей Lagochilus ilicifolius Bunge, растущих в степных сообществах Тувы (Черемушкина и др., 2016).
Рис. 2.
Механизмы формирования и пути трансформации биоморф модельных видов. а–и – основные морфологические структуры особей, формирующиеся на различных этапах онтогенеза, стрелки – пути развития биоморф и их переходы. 1 – почка возобновления; 2 – спящая почка; 3 – резид укороченный; 4 – удлиненный побег, возникший из почки возобновления; 5 – удлиненный побег, возникший из спящей почки; 6 – раздвинутоузловой резид; 7 – удлиненный резид; 8 – длиннокорневищно-удлиненный побег; 9 – побег корневищный полициклический, 10 – побег корневищный моноциклический; 11 – придаточные корни; 12 – надземная базальная часть побега (элемент эпигеогенного корневища); 13 – подземная базальная часть побега (элемент гипогеогенного коррневища).
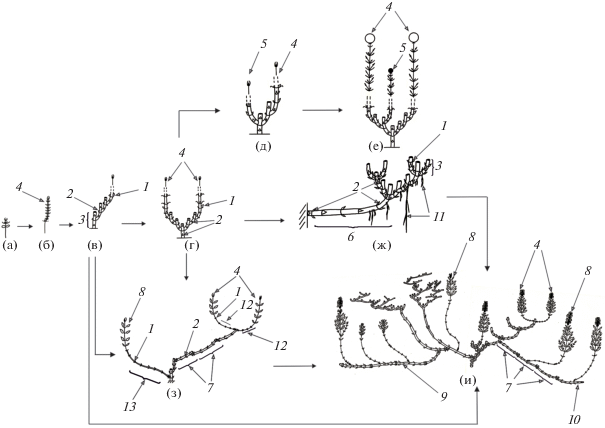
Детальный анализ биоморф особей N. pulchella показал, что растения формируют две жизненные формы: стержнекорневую каудексовую и короткокорневищно-стержнекорневую. Внутри стержнекорневой каудексовой биоморфы нами выделены две онтобиоморфы: компактнокаудексовая и рыхлокаудексовая (Astashenkov еt al., 2017). В течение онтогенеза происходит смена типа биоморфы с моноцентрической на неявнополицентрическую. Обычно стержнекорневая компактнокаудексовая онтобиоморфа формируется в крайних экологических условиях на границе верхнего предела произрастания – в криомезофитных лугах субальпийского пояса. Рыхлокаудексовая онтобиоморфа развивается на склонах гор по нижней границе распространения, обычно в лугово-степных сообществах. Побегообразование у особей этих биоморф происходит за счет функционирования почек регулярного возобновления, расположенных в узле третьего метамера. Чаще встречается короткокорневищно-стержнекорневая биоморфа (рис. 1б), которая формируется в условиях мезофитных разнотравных лугов и образуется в результате трансформации рыхлокаудексовой биоморфы. Становление короткокорневищно-стержнекорневой биоморфы начинается в молодом генеративном состоянии и связано с деятельностью спящих почек, развитие которых приводит к появлению раздвинутоузловых геофильных участков (рис. 2ж).
Побеги, развернувшиеся из спящих почек, удлиненные, моно-, реже дициклические, длина их геофильной части зависит от положения спящих почек на многолетних резидах (от 2 до 5 см), более длинные подземные участки отмечены у побегов, возникших из более старых почек. Обычно моноциклические побеги – вегетативные, дициклические – генеративные. В структуре дициклического побега первый годичный прирост состоит из плагиотропной геофильной части, второй – из генеративного ортотропного побега. Отмирание побегов происходит до геофильного участка с чешуевидными листьями. Почкой возобновления становится почка верхнего метамера сохранившегося резида. Первый раздвинутоузловой резид становится первым звеном симподиально-нарастающего плагиотропного гипогеогенного корневища, связанного с каудикулой каудекса.
Из верхней боковой почки удлиненного резида на следующий год развертывается новый моноциклический побег возобновления. После вегетации он отмирает до укороченной базальной части. Почками возобновления становятся почки второго-третьего метамера. В течение ряда лет развитие побегов следующих порядков осуществляется за счет почек, сохранившихся на укороченных резидах. Формируется парциальный куст – “центр закрепления”, состоящий в подземной сфере из системы симподиально-нарастающих укороченных резидов, которые образуют короткое корневище. На его многолетней части развивается большое число придаточных корней, которые углубляют в почву базальные части побеговых систем. Впоследствии один из придаточных корней утолщается, начинает лидировать в росте и после партикуляции особи функционально замещает главный корень – формируется придаточная корневая система с вторичным стержневым корнем. Новые “центры закрепления” появляются нерегулярно, что связано с периодичностью раскрытия новых спящих почек (от 2 до 5 лет). Число подобных “центров закрепления” может превышать 15. Моноцентрический тип биоморфы сменяется на неявнополицентрический. Корневище надстраивается резидами различной длины, что приводит к разрастанию осевых частей куста по периферии.
Особи вида N. schugnanica – длиннокорневищно-стержнекорневые растения (рис. 1в). В онтогенезе происходит смена типа биоморфы с моноцентрического на явнополицентрический.
На начальных этапах онтогенеза в побегообразовании особей участвуют только почки возобновления, расположенные в базальной части материнского побега годичного прироста (рис. 2a–в). Начиная с виргинильного состояния, в результате увеличения общих размеров особи базальная часть побега текущего прироста вместе с верхлежащими (2–3) удлиненными метамерами полегает и засыпается субстратом. Почки, сохранившиеся в узлах верхних метамеров, становятся почками возобновления, остальные почки не реализуются и переходят в спящее состояние. Погребенный участок побега представляет собой первое звено удлиненного эпигеогенного корневища (рис. 2з). Впоследствии одновременное развитие нескольких побегов приводит к формированию куртины с развитой придаточной корневой системой. В генеративном состоянии в побегообразование помимо почек возобновления включаются спящие почки, пробуждение которых приводит к формированию длиннокорневищноудлиненных побегов (термин Л.Е. Гатцук) (Гатцук, 1974). Эти побеги имеют геофильную часть с удлиненными метамерами, несущими чешуевидные листья, и надземную фотосинтезирующую часть также с удлиненными метамерами. Отмирание побегов происходит до геофильной части, сохранившийся удлиненный резид встраивается в общую структуру куртины – формируется комбинированное эпигипогеогенное корневище. Цикличность корневищных побегов зависит от положения спящих почек на многолетних частях растения, они могут быть моно-, ди-, реже трициклическими. Появление удлиненного корневища приводит к разрастанию особи, захвату и удержанию за ней территории. Подобное развитие отмечено нами у ряда видов Nepeta (N. bucharica, N. formosa), в том числе у особей N. sibirica (Водолазова и др., 2010).
N. transiliensis – типичный длиннокорневищный многолетник (рис. 1г). В онтогенезе особей меняется тип биоморфы с моноцентрического на явнополицентрический. До первого цветения особи находятся в виде компактных образований, смена биоморфы наступает в молодом генеративном состоянии. Начиная с проростка и до первого цветения, побегообразование осуществляется за счет раскрытия только почек возобновления, расположенных в базальной части побега. В структуре побега зона возобновления представлена тремя сильно сближенными и 1–2 удлиненными метамерами. В узлах последних закладываются почки возобновления, нереализованные почки становятся спящими. В молодом генеративном состоянии из спящих почек начинают развертываться длиннокорневищно-удлиненные и корневищные побеги, имеющие различную цикличность (рис. 2и). Корневищные побеги занимают плагиотропное направление роста, их метамерная протяженность не превышает семи метамеров, а их развитие, как правило, заканчивается отмиранием терминальной почки без выхода ее на поверхность почвы. Такой побег замещается новым корневищным побегом за счет развертывания одной боковой почки, расположенной в узле последнего, обычно шестого метамера. В результате подземного сочленения образуется симподиальное корневище, состоящее из 5–7 годичных приростов. В редких случаях корневищный побег способен нарастать моноподиально в течение 3‒4 лет. Каждый отдельный годичный прирост за счет раскрытия спящих почек ветвится. Побеги n + 1-го порядка характеризуются различной цикличностью от моно- до полициклических, а длина их геофильной части может достигать 30 см. Развитие корневищных побегов приводит к образованию куртины.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Анализ полученных результатов и имеющихся опубликованных данных по онтогенезу и сравнительной морфологии растений разных жизненных форм показал, что раскрытие спящих почек – один из определяющих механизмов изменения и формирования биоморф у некоторых видов растений. Нет сомнения, что реализация спящих почек приводит к формированию общего габитуса древесных форм, что убедительно отражено в ранее опубликованных работах (Серебряков, 1954; Мазуренко, Хохряков, 1977; Мазуренко, 1986; и др.). Также очевидна компенсаторная функция спящих почек, связанная с пережиданием неблагоприятных воздействий окружающей обстановки (Серебряков, 1959; Смирнова и др., 1984) или поддержанием жизнеспособности особи на финальных этапах ее развития (Серебряков, 1952; Борисова, 1961; и др.). С одной стороны, развитие спящих почек в онтогенезе можно рассматривать как результат реакции на изменяющиеся условия обитания, с другой – как генетически закрепленную программу побегообразования.
В некоторых литературных источниках приводятся прямые указания на изменения константной биоморфы в результате деятельности спящих почек у видов, оказавшиеся в нетипичных для них местообитаниях. Ранее было отмечено, что у некоторых древесных форм рода Potentilla L. развитие спящих почек приводит к формированию горизонтальных удлиненных ксилоризом (Гатцук и др., 1974). Так, при раскрытии спящих почек у особей Potentilla fruticosa L. развиваются удлиненные ксилоризомы, которые, выходя на поверхность почвы, образуют оси возобновления, а в результате кущения последних формируются парциальные кусты. У родственного вида Potentilla bifurca L. травянистой жизненной формы (Шафранова, 1970; Басаргин, 2011) на начальных этапах онтогенеза нарастающая в течение ряда лет моноподиально главная ось переходит в фазу куртины за счет развития спящих почек, в результате чего формируются горизонтальные гипогеогенные корневища с чешуевидными листьями. Подобный механизм формирования корневища описан также для Thermopsis lanceolata R. Br., Silene repens Patrin и др. (Басаргин, 2011). Такое побегообразование происходит регулярно и, скорее всего, закреплено генетически. Трансформация биоморфы за счет реализации спящих почек отмечена у некоторых видов рода Hedysarum L. (Karnaukhova, 2015). У особей Scutellaria supina L. описаны механизмы перестройки биоморфы из стержнекорневой в длиннокорневищно-стержнекорневую в результате появления из спящих почек удлиненных дициклических побегов замещения (Cheryomushkina, Guseva, 2015). Авторы рассматривают различные модификации биоморфы особей этого вида, в том числе с участием спящих почек, как образец возможных эволюционных морфологических трансформаций травянистой биоморфы в роде Scutellaria L.
Таким образом, на примере среднеазиатских представителей рода Nepeta, объединенных группой родства на уровне секции, показана роль спящих почек в побегообразовании особей, в становлении и трансформации биоморф.
Элементарной конструкционной единицей у модельных видов является резид, а в качестве базовой жизненной формы мы принимаем моноцентрическую стержнекорневую компактнокаудексовую (модель N. mariae). Характерная особенность становления такой биоморфы – регулярное функционирование спящих почек и построение многолетней основы тела растения укороченными ортотропными резидами. Нерегулярное пробуждение спящей почки и появление побега, имеющего раздвинутоузловой геофильный участок, приводит к образованию следующей структурной единицы – раздвинутоузлового резида. На основе такого резида формируется парциальный куст, который нарастает укороченными резидами n + 1-го порядка в течение нескольких лет, образуя короткое корневище (модель N. pulchella). Становление длиннокорневищно-стержнекорневой биоморфы (модель N. schugnanica) связано с регулярным пробуждением спящих почек, что приводит к формированию длиннокорневищноудлиненных побегов. Одновременно с этим происходит погружение удлиненных базальных частей текущих побегов, возникших из почек возобновления. Это сопровождается образованием удлиненного комбинированного эпигипогеогенного корневища. Регулярное участие в побегообразовании спящих почек и появление подземных корневищных побегов, которые в свою очередь становятся основой для развития разветвленной побеговой системы, характерно для типичной длиннокорневищной биоморфы с гипогеогенным удлиненным корневищем (модель N. transiliensis). Появление такой конструкционной единицы, как раздвинутоузловой резид, приводит к дезинтеграции особи в пространстве и смене биоморфы с моноцентрической на полицентрическую.
Скорее всего, повышенное увлажнение высокогорий и избыточный уровень водообеспечения растений в условиях прибрежных экотопов способствуют развитию длиннокорневищных биоморф с хорошо развитой системой придаточных корней. Мезофитные экологические условия дают предпосылки для формирования корневищных растений (Любарский, 1967).
Изменение типа биоморфы в ходе развития особей видов Nepeta от моноцентрической до неявно- и явнополицентрической за счет развития удлиненных конструкционных единиц приводит к захвату и удержанию пространства, что в эволюционном отношении могло оказаться одним из путей, обеспечивших устойчивое существование вида в современных эколого-фитоценотических условиях.
Реализация спящих почек в побегообразовании модельных видов через формирование побеговых систем и построение из их элементов общего габитуса растения являются адаптацией особей к высокогорным условиям обитания. Таким образом, функционирование спящих почек и развитие их в конструкционные элементы на примере видов Nepeta sect. Spicatae можно рассматривать как ароморфоз в эволюции жизненных форм растений, приводящий к усилению побегообразования и возникновению переходных и новых биоморф.
Работа выполнена по проекту Государственного задания Центрального Сибирского ботанического сада СО РАН №АААА-А17-117012610053-9.
Список литературы
Асташенков А.Ю., Черемушкина В.А. Архитектурная модель Nepeta mariae (Lamiaceae) // Раст. мир Азиат. Рос. 2016. № 4(24). С. 22–29.
Басаргин Е.А. Биоморфология некоторых длиннокорневищных видов растений и структура их ценопопуляций на юге Сибири. Дис. канд. биол. наук. Новосибирск:, 2011. 136 с.
Борисова И.В. Биология и основные жизненные формы двудольных многолетних травянистых растений степных фитоценозов Северного Казахстана // Тр. БИН АН СССР. Сер. 3. Геоботаника. М.; Л.: Наука, 1961. Вып. 13. С. 54–132.
Буданцев А.Л. Систематика рода Nepeta (Lamiaceae). Виды секции Spicatae // Ботан. журн. 1990. Т. 75. № 7. С. 1004–1013.
Водолазова С.В., Черемушкина В.А., Колегова Е.Б., Мяделец М.А. Онтогенез, структура ценопопуляций и эколого-ценотическая характеристика Nepeta sibirica (Lamiaceae) в Хакасии // Раст. ресурсы. 2010. Т. 46. Вып. 1. С. 3–16.
Гатцук Л.Е. К методам описания и определения жизненных форм в сезонном климате // Бюл. МОИП. Отд. биол. 1974. Т. 79. Вып. 3. С. 84–100.
Гатцук Л.Е., Дервиз-Соколова Т.Г., Иванова И.В., Шафранова Л.М. Пути перехода от кустарниковых форм к травянистым в некоторых таксонах покрытосеменных // Проблемы филогении высших растений. М.: Наука, 1974. С. 16–36.
Жукова Л.А. Популяционная жизнь луговых растений. Йошкар-Ола: Ланар, 1995. 224 с.
Зёдинг Г. Ростовые вещества растений. М.: Изд-во иностр. лит., 1955. 389 с.
Кожевников А.В. Весна и осень в жизни растений. М.: Моск. об-во испытателей природы, 1950. 238 с.
Кочкарева Т.Ф. Семейство Lamiaceae // Флора ТаджССР. Л.: Наука, 1986. Т. 8. С. 104–142.
Любарский Е.Л. Экология вегетативного размножения высших растений. Казань: КГУ, 1967. 184 с.
Любименко В.Н. О спящих почках // Тр. СПб. об-ва естествоиспытателей. Отд. ботан. 1900. Т. 30. Вып. 3. С. 195–259.
Ляшенко Н.И. О спящих почках кустарников // Ботан. журн. 1958. Т. 43. № 7. С. 1039–1047.
Ляшенко Н.И. Биология спящих почек. М.; Л.: Наука, 1964. 85 с.
Мазуренко М.Т. Биоморфологические адаптации растений Крайнего Севера: М.: Наука, 1986. 208 с.
Мазуренко М.Т., Хохряков А.П. Структура и морфогенез кустарников. М.: Наука, 1977. 151 с.
Марфенин Н.Н. Концепция модульной организации в развитии // Журн. общ. биологии. 1999. Т. 60. № 1. С. 6–17.
Нотов А.А. О специфике функциональной организации и индивидуального развития модульных объектов // Журн. общ. биологии. 1999. Т. 60. № 1. С. 60–79.
Нухимовский Е.Л. Основы биоморфологии высших растений. М.: Недра, 1997. Т. 1. 630 с.
Османова Г.О. Морфологические особенности особей и структура ценопопуляций Plantago lanceolata L. Йошкар-Ола: Мар. гос. ун-т, 2007. 184 с.
Пояркова А.И. Новые виды котовника // Ботан. мат. герб. БИН АН СССР. 1953. Т. 15. С. 282–320.
Пояркова А.И. Семейство Lamiaceae // Флора СССР. М.; Л.: Наука, 1954. Т. 20. С. 286–360.
Работнов Т.А. Жизненный цикл многолетних травянистых растений в луговых ценозах // Тр. БИН АН СССР. Сер. 3. Геоботаника. М.; Л.: Наука, 1950. 176 с.
Савиных Н.П. Архитектура трав // Актуальные проблемы современной биоморфологии: Матер. II Всерос. школы-семинара. Киров: Радуга-Прес, 2012. С. 342–354.
Серебряков И.Г. Морфология вегетативных органов высших растений. М.: Наука, 1952. 390 с.
Серебряков И.Г. Биолого-морфологический и филогенетический анализ жизненных форм покрытосеменных // Уч. зап. МГПИ им. В.П. Потемкина. 1954. Т. 37. Вып. 2. С. 21–89.
Серебряков И.Г. Основные направления эволюции жизненных форм у покрытосеменных растений // Бюл. МОИП. Отд. биол. 1955. Т. 60. № 3. С. 71–91.
Серебряков И.Г. Период покоя у некоторых травянистых и древесных растений Подмосковья // Уч. зап. МГПИ им. В.П. Потемкина. 1959. Т. 100. Вып. 5. С. 39–51
Серебряков И.Г. Экологическая морфология растений. М.: Высш. шк., 1962. 378 с.
Серебрякова Т.И. Об основных “архитектурных моделях” травянистых многолетников и модусах их преобразования // Бюл. МОИП. Отд. биол. 1977. Т. 82. Вып. 5. С. 112–128.
Серебрякова Т.И. Еще раз о понятии “жизненная форма” у растений // Бюл. МОИП. Отд. биол. 1980. Т. 85. Вып. 6. С. 75–86.
Серебрякова Т.И. Почка как этап развития побега // Тез. докл. VII Всесоюз. ботан. об-ва. Л.: Наука, 1983. С. 232.
Серебрякова Т.И., Петухова Л.В. “Архитектурная модель” и жизненные формы некоторых травянистых растений// Бюл. МОИП. Отд. биол. 1978. Т. 83. Вып. 6. С. 51–64.
Смирнова О.В. Структура травянистого покрова широколиственных лесов. М.: Наука, 1987. 206 с.
Смирнова О.В., Чистякова А.А., Истомина И.И. Квазисенильность как одно из проявлений фитоценотической толерантности растений // Журн. общ. биологии. 1984. Т. 45. № 2. С. 216–225.
Смирнова О.В., Заугольнова Л.Б., Торопова Н.А., Фаликов Л.Д. Критерии выделения возрастных состояний и особенности хода онтогенеза у растений различных биоморф. Ценопопуляции растений. М.: Наука, 1976. С. 14–44.
Стешенко А.П. Формирование структуры полукустарничков в условиях высокогорий Памира // Тр. Бот. ин-та. АН ТаджССР. 1956. 50 с.
Уранов А.А. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов // Науч. докл. высш. шк. Биол. науки. М.: Наука, 1975. № 2. С. 7–34.
Хохряков А.П. Эволюция биоморф растений. М.: Наука, 1981. 168 с.
Черемушкина В.А. Биология луков Евразии. Новосибирск: Наука, 2004. 279 с.
Черемушкина В.А. Особенности онтогенеза короткокорневищных растений // Растительный мир и его охрана: Матер. Междунар. науч. конф., посвященной 80-летию Института ботаники и фитоинтродукции, г. Алматы, 5–7 сентября 2012 года. Алматы: LEM, 2012. С. 396–398.
Черемушкина В.А., Асташенков А.Ю., Гусева А.А., Денисова Г.Р. Характеристика Lagochilus ilicifolius (Lamiaceae) в краевых популяциях Тувы // Раст. ресурсы. 2016. Т. 52. Вып. 3. С. 381–396.
Шафранова Л.М. О некоторых путях перехода от кустарников к травам в роде лапчатка (Potentilla L.): Автореф. дис. канд. биол. наук. М.: МГПИ, 1970. 30 с.
Шафранова Л.М., Гатцук Л.Е. Растение как пространственно-временная метамерная (модульная) система // Успехи экологической морфологии растений. М.: Прометей, 1994. С. 6–7.
Aaron I. Dormant and adventitious buds // Science. 1946. V. 104. 329 p.
Astashenkov A.Yu., Cheryomushkina V.A., Grebenjuk A.V., Dzumanov S.D. Transformation of life forms and ontogenetic structure of Nepeta pulchella Pojark. coenopopulations in Acsy-Zhabaglinsky nature reserve // Contem. Probl. Ecol. 2017. V. 10. № 6. P. 758–771.
Baldini E., Mosse B. Observations on the origin and development of spheroblasts in the apple // J. Hortic. Sci. 1956. V. 31. S. 156–162.
Barthélémy D., Caraglio Y. Plant architecture: a dynamic, multilevel and comprehensive approach to plant form, structure and ontogeny // Ann. Bot. 2007. № 99(3). P. 375–407.
Büsgen M. Bau und Leben unserer Waldbäume Illus. Jena. 1897. 230 s.
Chandler W. Fruit growing. Cambridge: Riverside Press, 1925. P. 14–19.
Cheryomushkina V.A., Guseva A.A. Life forms of Scutellaria supina L. (Lamiaceae) // Contem. Probl. Ecol. 2015. V. 8. № 5. P. 624–635.
Esser P. Die Entstehung der Blüten am alten Holze // Verh. Nat. Hist. Verein. Bonn: Jahrg, 1887. Bd 44(4). S. 69–112.
Getmanets I.A. Morphoadaptive determination of structural diversity of biomorphs of Salix L. species in Southern Urals // Contem. Probl. Ecol. 2015. V. 8. № 5. P. 574–583.
Goebel K. Organographie der Pflanzenzen, insbesondere der Archegoniaten und Samenpflanzen. Aufl. 3. Teil. 1. Jena: Fisher, 1928. 642 s.
Haberlandt G. Eine botanische Tropenreise. Aufl., 3. Leipzig: Endelmann, 1926. 292 s.
Hallé F., Oldeman R.A.A. Essai sur l architecture et la dynamique de croissance des arbres tropicaux. Paris: Masson, 1970. 178 p.
Hallé F., Oldeman R.A.A., Tomlinson P.B. Tropical trees and forests. Berlin; N.Y.: Springer, 1978. 441 p.
Hartig Th. Vollständige Naturgeschichte der Forstlichen Culturpflanzen Deutschlands. Berlin: A. Förstner'sche Verlagsbuchhandlung, 1851. 580 s.
Jost L. Über schlafende Knospen // Flora. 1925. Bd 118/119. S. 289–299.
Karnaukhova N.A. Ontogenesis and life forms of Hedysarum (Fabaceae) in south Siberia // Contem. Probl. Ecol. 2015. V. 8. № 5. P. 614–623.
Khutornoi O.V., Velisevich S.N., Vorob’ev V.N. Ecological variation in the morphological structure of the crown in Siberian stone pine at the Timberline // Russ. J. Ecol. 2001. V. 32. № 6. P. 393–399.
Kostina M.V., Barabanshikova N.S., Bityugova G.V. Yasinskaya O.I., Dubach A.M. Structural modifications of birch (Betula pendula Roth.) crown in relation to environmental conditions // Contem. Probl. Ecol. 2015. V. 8. № 5. P. 584–597.
Meusel H. Uber Wuchsformen, Verbreitung und Phylogenie einiger mediterranmitteleuropaischen Angiospermen – Gattungen // Flora. 1953. Bd 139. H. 3. S. 333–393.
Měch R., Prusinkiewicz P. Visual models of plants interacting with their environment // Computer Graphics Proceedings, Annual Conference Series, ACM SIGGRAPH. New Orleans, Louisiana: Assn for Computing Machinery, 1996. P. 367–410.
Millet J., Bouchard A., Ѐdelin C. Plagiotropic architectural development of four tree species of the temperate forest // Can. J. Bot. 1998. V. 76. № 12. P. 2100–2118.
Nikolini E., Chanson B., Bonne F. Stem growth and epicormic branch formation in understorey beech trees (Fagus sylvatica L.) // Ann. Bot. 2001.V. 87. P. 737–750.
Ping Li., Dohnston M.O. Heterochrony in plant evolutionary studies through the twentieth century // Bot. Rev. 2000. V. 66. P. 57–88.
Sabatier S., Barthélémy D. Bud structure in relation to shoot morphology and position on the vegetative annual shoots of Juglans regia L. (Juglandaceae) // Ann. Bot. 2001. V. 87. P. 117–123.
Velenovsky J. Vergleichende Morphologie der Pflanzen. V. 1. Praga: Rivnac, 1905. 274 s.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая