

Известия РАН. Серия биологическая, 2019, № 6, стр. 614-624
Система диагностических признаков покровных перьев птиц отряда ржанкообразных
О. Л. Силаева *
Институт проблем экологии и эволюции им. А.Н. Северцова
117071 Москва, Ленинский просп., 33, Россия
* E-mail: silaeva.o@gmail.com
Поступила в редакцию 06.02.2018
После доработки 26.04.2018
Принята к публикации 30.04.2018
Аннотация
Выявлены комплексы структурных признаков покровных перьев ржанкообразных видов птиц, имеющих таксономическое значение. Эти комплексы объединены в систему диагностических признаков, которая использована для идентификации таксонов птиц по одному покровному перу, по части пера и/или по мелким фрагментам перьев. Отмечено, что для успешного определения таксона необходимо использование признаков в рамках системы.
Отдельное перо птицы, будучи структурной составляющей оперения в целом, представляет собой системный комплекс тесно связанных между собой, частично коррелированных в линейно-метрических соотношениях микро- и макроструктурных элементов. Данные элементы могут обладать разными свойствами: показывать индивидуальную изменчивость и филогенетические связи между таксонами, а также служить критериями таксона.
Цель работы – выявление и описание системных макро- и микроструктурных признаков отдельного покровного пера, пригодных для идентификации таксона в исследованиях разной направленности. Определение видовой принадлежности птиц по перьям и их фрагментам востребовано в теоретических и практических сферах деятельности человека: в орнитологических исследованиях (миграции, линька, кольцевание птиц, питание хищных животных), в археологии и судебной науке (охрана природы), в определении загрязнения пищи, в криминалистике, в лесном хозяйстве, в таможенной службе и в музейном деле.
Основа практического идентификационного направления – фундаментальные классические работы (Nitzsch, 1840; Chandler, 1916; Lucas, Stettenheim, 1972), определившие место морфологии пера, или птилологии, как научного направления наряду с остеологией и морфологией других органов. Они обусловили разработку методик исследования перьевого материала для выявления микроструктурных отличий, имеющих таксономическое значение. Лейборн впервые начала определять мелкие останки птиц в интересах авиационной орнитологии (Laybourne, 1974), а позднее вместе с коллегами она усовершенствовала методику подготовки перьевых останков для проведения микроструктурного анализа. Метод исследования основан на структурных вариациях пуховых бородок и их элементов, принадлежащих к разным таксонам (Laybourne et al., 1992; Laybourne, Dove, 1994).
Значительный вклад в развитие диагностического направления внесли сотрудники Экспертного центра таксономической идентификации (Expert Center for Taxonomic Identification (ETI)), опубликовавшие фундаментальную мультимедийную базу данных по идентификации останков птиц (Bird remains identification system (BRIS)) (Brom, 1991; Prast, Shamoun, 1996; 2001). Данные BRIS стали активно применять в авиационно-орнитологических исследованиях (Rochard, Horton, 1977). Впервые с помощью мультивариантного анализа МANOVA и одновариантного ANOVA, а также дискриминантного анализа функций были изучены количественные характеристики микроструктурных признаков пера и было показано, что признаки микроструктуры с успехом могут использоваться в качестве таксономических (Dove, 1997).
В 1960–1970-х гг. исследования перьевого материала начали проводить как ученые АН СССР (Ильичев, 1961, 1963; Бородулина, 1964; Добринский, 1966; Яблоков, Валецкий, 1972), так и ученые отраслевой науки, в частности это были эксперты в области судебной медицины и криминалистики (Бокариус, 1961; Бордонос, Булыга, 1968). Значительный вклад в исследования микроструктуры пера был внесен авторами книги “Архитектоника перьев и ее диагностическое значение” (Чернова и др., 2006). Активное изучение микроструктурных особенностей перьевого материала продолжается в ИПЭЭ РАН, где исследуются как количественные, так и качественные характеристики перьевых структур (Силаева и др., 2012, 2013, 2015).
МАТЕРИАЛЫ И МЕТОДЫ
Для 43 видов семи семейств ржанкообразных (Charadriiformes) мы попытались выявить диагностические признаки и особенности архитектоники перьевых структур (табл. 1). Классификация соответствует конспекту орнитологической фауны (Степанян, 2003).
Таблица 1.
Исследованный материал по отряду Ржанкообразных
| Вид | Число | Вид | Число | ||
|---|---|---|---|---|---|
| особей | перьев | особей | перьев | ||
| Charadriidae | Glareolidae | ||||
| Pluvialis squatarola | 4 | 10 | Glareola pratincola | 4 | 11 |
| P. apricaria | 3 | 5 | Stercorariidae | ||
| Charadrius hiaticula | 3 | 6 | Stercorarius pomarinus | 3 | 10 |
| C. dubius | 3 | 11 | S.parasiticus | 10 | 52 |
| Vanellus vanellus | 3 | 14 | Laridae | ||
| Arenaria interpres | 3 | 12 | Larus ichthyaetus | 3 | 30 |
| Haematopodidae | L. minutus | 3 | 16 | ||
| Haematopus ostralegus | 4 | 13 | L. ridibundus | 3 | 20 |
| Scolopacidae | L. argentatus | 3 | 25 | ||
| Tringa ochropus | 3 | 10 | L. cachinnans | 3 | 148 |
| T. nebularia | 5 | 12 | L. hyperboreus | 3 | 5 |
| T. totanus | 4 | 10 | L. canus | 4 | 12 |
| Actitis hypoleucos | 12 | 44 | Rissa tridactyla | 3 | 6 |
| Xenus cinereus | 3 | 5 | Chlidonias niger | 3 | 13 |
| Phalaropus fulicarius | 4 | 14 | C. leucopterus | 3 | 9 |
| P. lobatus | 12 | 45 | Hydroprogne caspia | 3 | 6 |
| Philomachus pugnax | 3 | 10 | Sterna hirundo | 3 | 8 |
| Calidris temminckii | 4 | 17 | Alcidae | ||
| C. ferruginea | 4 | 17 | Uria lomvia | 4 | 20 |
| C. alpine | 5 | 27 | Cepphus grylle | 3 | 10 |
| Lymnocryptes minimus | 3 | 11 | Aethia cristatella | 3 | 33 |
| Gallinago gallinago | 6 | 14 | Cyclorrhynchus psittacula | 3 | 35 |
| G. media | 40 | 96 | |||
| Scolopax rusticola | 8 | 24 | |||
| Numenius arquata | 4 | 9 | |||
| Limosa limosa | 3 | 10 | |||
Исследовали лучи пуховых и комбинированных бородок базальной части полупуховых межлопаточных перьев (tectrices interscapularis). Полупуховым называется перо, пуховая часть которого составляет более двух третей опахал. Именно покровные перья птиц и их фрагменты чаще всего поступают на микроструктурную экспертизу для идентификации таксона птицы. Мы рассматриваем преимущественно качественные данные перьевых структур, не касаясь их морфометрических характеристик.
Исследования проводили с помощью визуального осмотра и методов электронной сканирующей и светооптической микроскопии: использовали сканирующий электронный микроскоп (СЭМ) Tescan Vega TS 5130 MM (Cam Scan MV 2300), светооптический микроскоп Leica DMR 2700 (Leica, Германия), оснащенный фотокамерой JVC 3 CCD C-MOUNT, микроскоп Am Scope ×40‒×2500 LED Lab Compound Microscope (Китай) with 3D Two-Layer Mechanical Stage с увеличением ×4/0.1, ×10/0.25, ×40/0.65, ×100/1.25 oil., бинокулярную лупу МБС 9 (СССР). Для обеспечения проводимости перед исследованием в СЭМ на биологические образцы напылялось золото S150A Sputter Coater в соответствии с методиками подготовки пера в ИПЭЭ РАН для исследования его архитектоники (Чернова и др., 2006, 2009). Обработка изображений проводилась в программе Adobe Photoshop CC 2014.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Узлы с зубцами как элементы пухового луча. Представители отряда имеют продолговатые узкие узлы, которые, как правило, плохо выделяются на фоне междоузлий, особенно в дистально-медиальной части луча; зубцы узлов здесь располагаются вдоль междоузлия. Четко видны узлы в проксимальных частях луча, где они раскрыты, тело узла широкое, а зубцы узлов расположены почти под прямым углом к междоузлиям (рис. 1б). Особенно это характерно для семейства чайковых (рис. 2в).
Рис. 1.
Элементы пухового луча. а – базальная часть луча степной тиркушки Glareola nordmanni, б – базальная часть луча моевки Rissa tridactyla, в – дистальная часть луча белобрюшки.

Рис. 2.
Сужения и искривления междоузлий пуховых лучей. а – сужение междоузлий пухового луча галстучника, б – круглоносого плавунчика, в – искривления междоузлий серебристой чайки.

Луч к вершинной части обычно утончается. В этой части луча у большинства представителей отряда тело узла уменьшается, редуцируется, а зубцы становятся длинными и заостренными. Число таких узлов варьирует у разных видов от 2‒3 до 6‒7. Тело узла покровного пера вальдшнепа в медиальной и проксимальной частях луча почти не выявляется на фоне междоузлия. Зубцы чаще всего сглажены настолько, что имеют вид едва заметных выступов и располагаются вдоль междоузлия.
У дупеля, камнешарки и большого кроншнепа на апикальных концах луча нет узлов с заостренными зубцами; зубцы остаются такими же короткими, как на медиальных участках. У ржанковых, тиркушковых и бекасовых тело узла короче, но более выпуклое по сравнению с таковыми у чайковых и чистиковых (рис. 1). У луговой тиркушки, большого веретенника и некоторых других видов этих семейств тело узла хорошо определяется, несмотря на то что на некоторых узлах зубцы загибаются вовнутрь к междоузлию, делая узел менее заметным. У кулика-сороки зубцы почти не выражены, они образуют лишь неровный кант.
Плохо выражены узлы на всех участках пуховых лучей представителей семейства чистиковых, в частности большой конюги, белобрюшки и чистика. У чистиковых в дистальных сегментах луча узлы различаются благодаря чрезвычайно длинным зубцам (рис. 1в).
У представителей семейства поморниковых узлы почти одинаковые во всех сегментах луча. В медиальных частях лучей зубцы одного узла сильно различаются между собой по длине. У чайковых в проксимальной части луча узлы широкие, вздутые, с расходящимися, отклоняющимися в стороны от междоузлия зубцами, образующими нечто похожее на розетки. Однако у некоторых видов, например у бургомистра, узлы в проксимальных частях луча не имеют ярко выраженных розеток. В медиальной части узел выглядит компактнее, а в дистальной части его тело редуцируется, зубцы вытягиваются и заостряются. В апикальной части луча обычно 5‒7 редуцированных узлов. Узлы у чаек по форме сходны с таковыми у крачек, но крупнее.
Макро- и микроструктурные характеристики исследованных нами представителей группы больших белоголовых чаек (серебристой чайки и чайки-хохотуньи) очень сходны между собой. Скорее всего, особенности их структур могут быть выявлены с помощью анализа многомерных данных с использованием микроструктурных промеров (Силаева и др., 2011, 2012, 2013, 2015; Силаева, Вараксин, 2016). Узлы преимущественно вздутые с неравномерно расположенными широкими, короткими зубцами. К вершине луча узлы уменьшаются, на апикальном конце имеется 3‒4 редуцированных узла.
Различия между структурой элементов лучей ржанковых, тиркушковых и бекасовых, с одной стороны, и чистиковых, с другой стороны, значительны. Они проявляются в конфигурации, размерах узлов и узловых зубцов в зависимости от сегментов луча, что позволяет дифференцировать представителей этих семейств.
Междоузлия как элементы пухового луча. К основным качественным характеристикам междоузлий относятся их сужения и искривления (рис. 2). Сужения междоузлий отмечаются в центральной части между двумя узлами или ближе к дистальной части. Они наблюдаются в следующих семействах отряда ржанкообразных: бекасовых (гаршнеп, белохвостый песочник, чернозобик, турухтан, круглоносый плавунчик), ржанковых (галстучник), чайковых (белокрылая крачка, серебристая чайка). Видно, что у представителей отряда искривления междоузлий распространены мозаично. Можно предположить, что крупные тяжелые узлы на тонких междоузлиях вызывают искривления последних.
Почти у всех представителей отряда ржанкообразных наблюдаются удлинения междоузлий к дистальному концу луча, однако удлинения необходимо оценивать на основе морфометрических данных. Междоузлия и узлы имеют продольную ребристость у всех представителей отряда, что наблюдается и у представителей других отрядов (Чернова и др., 2009).
Пигментация элементов пухового луча. Предполагается, что концентрация пигмента варьирует в зависимости от принадлежности пера к той или иной птерилии и не зависит от таксона, к которому относится данная особь (Prast, Shamoun, 1996). Следует также отметить, что степень пигментации по-разному просматривается в светооптическом и электронном сканирующем микроскопах. Однако очевидно, что на апикальных отрезках лучей пигментация редуцированных узлов почти у всех видов отсутствует. Наиболее пигментированные узлы всегда имеются в базальных частях лучей, и, как правило, это наиболее крупные узлы. Узлы ржанковых, тиркушковых и бекасовых лучше пигментированы, по крайней мере на медиальных и проксимальных участках луча, чем узлы чайковых, поморниковых и чистиковых (рис. 1).
Конусы и билатеральные полосы. Соотношения между контурными и пуховыми частями классических комбинированных бородок (бородки, несущие как контурные, так и пуховые лучи), а также соотношения между числом комбинированных, контурных и чисто пуховых бородок формируют конфигурацию опахал, которая визуально проявляется в сочетании билатеральных рисунков полос и/или (тупых/острых) конусов (1‒3 на рис. 3). Полосы образуются при сохранении на определенном промежутке вдоль стержня постоянных длин контурных и пуховых частей комбинированных бородок. В результате изменения соотношений между длинами контурных и пуховых частей комбинированных бородок в основном уменьшаются длины контурных частей по направлению к базальной части пера, на концах опахал образуются острые или тупые билатеральные конусы (1, 2 на рис. 3). В полупуховых перьях пуховые части комбинированных бородок образуют вполне симметричные билатеральные полосы; в контурных перьях за счет короткой непуховой части получается тупой конус. Под конусом, как правило, располагаются чисто пуховые бородки, в случае если конус с комбинированными бородками в базальной части оставляет для них место, т.е. не доходит вплотную до верхнего пупочного отверстия (1 на рис. 3).
Рис. 3.
Билатеральные полосы и конусы, образованные контурными частями бородок. 1 – бургомистр, 2 – гаршнеп, 3 – бекас Gallinago gallinago. Пропорции размеров перьев на рисунке не соблюдены.

Есть некоторые отличия в образовании конусных структур и билатеральных полос между семействами отряда ржанкообразных. У ржанковых, бекасовых, тиркушковых и куликов-сорок эти структуры чаще выглядят как конусы, а не как полосы. Однако у перевозчика, чернозобика, кулика-сороки, плосконосого плавунчика отмечены структуры в виде полос, оканчивающиеся острым или тупым конусом. У гаршнепа на ряде перьев отмечен острый конус с некоторым расширением на проксимальном конце (2 на рис. 3).
У чайковых, поморниковых и чистиковых контурные части комбинированных бородок образуют преимущественно широкие или узкие билатеральные полосы на опахалах, заканчивающиеся тупым или острым в зависимости от ширины контурных билатеральных полос конусом. При наличии узких билатеральных полос на проксимальном конце образуются острые конусы (3 на рис. 3). Довольно часто из-за разреженности комбинированных бородок конус и билатеральные полосы почти не просматриваются.
Дополнительные пуховые образования. К основным пуховым термоизолирующим образованиям мы относим классические комбинированные бородки и типичные чисто пуховые бородки с пуховыми лучами в базальных частях опахал. Дополнительные пуховые образования ‒ дополнительное перо (ДП) и пупочный пух (ПП), а также модифицированные лучи (МЛ) (рис. 4). ДП и основное перо (ОП) мы рассматриваем как перьевую пару, в которой ДП ‒ уменьшенная копия ОП. ДП отличается хорошо оформленным стержнем, к нему прикрепляются пуховые бородки, несущие пуховые лучи. Бородки ПП в отличие от ДП образуют пучок или ободок, который окаймляет дистальную часть очина основного пера в области верхнего пупочного отверстия. При наличии ДП бородки ПП могут отходить билатерально от его стержня (Силаева, 2011; Силаева, Горохова, 2017).
Рис. 4.
Дополнительные образования в перьях мородунки (а) и чайки-хохотуньи (б и в). 1 – дополнительное перо, 2 – пупочный пух, 3 – пуховая вставка с модифицированными лучами. Масштаб: 1 см.
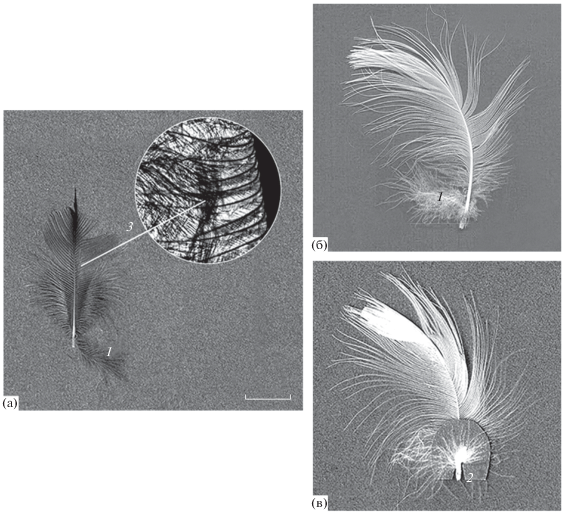
ДП и ПП распределяются мозаично в отрядах и семействах птиц, представляя собой комплексные структуры, входящие в систему диагностических признаков (СДП). ДП и ПП особенно хорошо развиты в семействах чайковых, поморниковых и чистиковых, встречаются почти на всех покровных перьях представителей данных семейств.
С помощью птерилографического анализа мы оценили наличие ДП и ПП в птерилиях чайковых (чайка-хохотунья), поморниковых (короткохвостый поморник) и чистиковых (толстоклювая кайра) птиц. Для каждой птерилии одного представителя семейства исследовалась выборка ~30 перьев.
Дополнительные образования в виде ДП и ПП есть на покровных перьях всех исследованных птерилий. Наверняка более плотное оперение, усиленное этими пуховыми структурами, необходимо для защиты жизненно важных и наиболее уязвимых органов от механических повреждений, переохлаждения и проникновения воды.
У всех трех представителей чайковых, поморниковых и чистиковых в оперении головы имеется 75‒92% перьев с ДП. При этом соблюдается определенное равновесие: именно стольких перьев достаточно, чтобы защитить голову птицы от переохлаждения и механических повреждений. Учитывается, что ДП как более плотная структура по сравнению с ПП утяжелила бы тело птицы.
В шейной птерилии чайки-хохотуньи дополнительные макроструктурные образования распределяются неравномерно: в дорзальной части ‒ 32% перьев с ДП и 68% с ПП, в вентральной ‒ 86% перьев с ДП и всего 14% с ПП.
Пупочный пух есть на ушных птерилиях всех трех видов. ДП и ПП на переднеушной и заднеушной птерилиях функциональны для слуха (Ильичев, 1961, 1963). На этих птерилиях в основном встречается ПП как более легкое образование по сравнению с ДП. Закономерность распределения ДО по таксонам и птерилиям пока не до конца ясна.
Модифицированные лучи (МЛ) с удлиненными опахальцами. Это особая пуховая структура, которая появляется на контурных бородках в дистальной части опахал, что делает эти бородки комбинированными. МЛ образуют пуховую вставку в медиально-дистальную контурную часть опахал, расположенную билатерально относительно стержня ОП. Пуховые вставки ржанкообразных по сравнению с таковыми воробьинообразных (Passeriformes) (семейство Passeridae) имеют обычно овальную форму. Пуховая вставка с МЛ не прикрывается соседними структурами и не подвергается интенсивному обнашиванию, как края пера (рис. 4а, 3).
МЛ распространены мозаично в следующих семействах отряда ржанкообразных: ржанковых (тулес, золотистая ржанка, галстучник, малый зуек, чибис, камнешарка), куликах-сороках (кулик-сорока), бекасовых (черныш, травник, мородунка, круглоносый плавунчик, плосконосый плавунчик, чернозобик, краснозобик, гаршнеп), тиркушковых (луговая тиркушка). В семействах чайковых, поморниковых и чистиковых МЛ отсутствуют.
Опахальца МЛ длиннее классических пуховых структур соответствующего вида; зубцы на узлах МЛ чрезвычайно длинные и острые (Силаева, 2011; Силаева и др., 2012, 2013, 2015).
Полупрозрачные базальные клетки МЛ длиннее таковых у типичных пуховых лучей. Однако плотность типичных пуховых и контурных лучей выше плотности МЛ. Таким образом, линейные размеры и формы микроструктурных элементов МЛ и типичных пуховых базальных лучей различаются. Удлиненные пуховые опахальца МЛ характеризуются более длинными и широкими междоузлиями по сравнению с таковыми у типичных пуховых лучей. МЛ, вероятно, представляют собой контурные лучи, преобразованные в пуховые.
Типы обнашивания пера. Форма покровных перьев у представителей отряда ржанкообразных преимущественно округлая или продолговатая. Форма дистального среза перьев довольно разнообразна: он может быть заостренным, округлым, скошенным косо в сторону более широкого опахала или прямо обрезанным.
Довольно часто встречающиеся у куликов и ржанок заостренные дистальные срезы перьев, т.е. симметрично срезанные по косой апикальные концы обоих опахал, возможно, приводят к особому типу обнашивания перьев, когда в дистальной части пера появляется выступающий за срез пера конец стержня с сильно укороченными бородками. У некоторых видов на межлопаточных и других покровных перьях вершинный конец стержня на 1‒3 мм выходит за дистальный срез опахал (2 на рис. 5). Выступающий конец стержня может иметь вид более или менее равнобедренного треугольника с контурными бородками, длина которых равномерно уменьшается по направлению к апикальному краю. Образование выступающего конца стержня происходит в результате укорачивания и/или потери вершинных бородок стержня пера при сохранении более прочного апикального конца стержня.

Тип обнашивания пера с образованием выступающего апикального конца стержня характерен для представителей отряда ржанкообразных: ржанковых (тулес, галстучник, малый зуек, камнешарка), куликов-сорок (кулик-сорока), бекасовых (травник, мородунка, плосконосый плавунчик, белохвостый песочник), тиркушковых (луговая тиркушка).
Такой срез встречается и на кроющих маховых и рулевых перьях. Крайне редко заостренный дистальный срез появляется на перьях с билатеральным скосом обоих опахал и на латерально изогнутых перьях. Тип обнашивания пера (2 на рис. 5) может быть рассмотрен как альтернатива обычному обнашиванию, выражающемуся в беспорядочно расположенных выщербинах на обношенном пере (3 на рис. 5) или примерно однородном истончении апикальных концов бородок опахал (1 на рис. 5). Для чайковых, поморниковых и чистиковых такой тип обнашивания пера не характерен (если он и встречается, то дистальный срез пера несимметричен).
Подотряд Charadrii с семействами ржанковых, бекасовых и тиркушковых по большинству признаков перьевых структур противопоставляется подотряду Lari с семействами поморниковых и чайковых. Подотряд Alcea по большинству признаков оперения приближается к подотряду Lari.
Рассмотрим комплексы структурных признаков, имеющих таксономическое значение и, следовательно, пригодных для включения в СДП. В первую очередь это ‒ элементы пухового луча: узлы, их форма, размер (тело узла и зубцы), плотность расположения на участках луча, распределение узлов по сегментам луча (Chandler, 1916; Lucas, Stettenheim, 1972; Brom, 1991; Силаева, 2011; Силаева и др., 2012, 2013, 2015). Для таких комплексов микроструктурных признаков, которыми обладает узел, важны также морфометрические исследования. Так, например, число зубцов на узлах разных таксонов варьирует, при этом нет критерия, какого размера отросток следует считать зубцом, а какой – неровностью дистального края узла. Для выработки критерия необходимо проведение микроизмерений.
Искривления, сужения и пигментацию междоузлий можно использовать вместе с другими макро- и микроструктурными признаками пера как таксономические признаки. Конфигурация билатеральных полос и конусов как макроструктурные признаки могут помочь в общем сравнительном анализе при идентификации таксона по покровным перьям. Типы обнашивания пера также можно использовать в таксономических целях. Эти признаки видны невооруженным глазом и не требуют применения метода микроскопирования для их выявления.
Дополнительные образования в виде ДП, ПП и МЛ, а также комплексные вариации их структур могут участвовать в таксономической идентификации высших и средних таксонов (Chandler, 1916; VanTyne, 1959; Ziswiler, 1962; Силаева, Горохова, 2017). Так, например, факт наличия у чайковых, поморниковых и чистиковых птиц ДП и ПП и отсутствие МЛ позволяют выявить принадлежности к определенному высшему или среднему таксону. При этом сочетание у представителей ряда семейств ржанкообразных птиц разных дополнительных образований можно использовать как показатель принадлежности особи к одному из данных семейств. Кроме того, комплексные характеристики дополнительных образований (размер, форма, структура МЛ, а также бородок ДП и ПП и т.д.) способствуют в некоторой степени выявлению птерилии, которой принадлежит перо. Факт наличия дополнительных образований устанавливается обычно без использования микроскопов, а у крупных перьев и некоторые структурные признаки можно увидеть с помощью лупы и бинокулярной лупы в отсутствие специального оборудования. Однако мозаичное распределение ДО в разных таксономических группах уменьшает их ценность как таксономических признаков и требует использования их непременно в общей СДП.
ЗАКЛЮЧЕНИЕ
Все выявленные комплексные характеристики могут иметь не только таксономическое, но и филогенетическое значение, а также показывать индивидуальную изменчивость перьевых структур в целом и их отдельных элементов. Для элементов луча – это формы узлов и зубцов на разных участках луча, для дополнительных образований – размеры и формы самого образования и составляющих его структур, лучей и бородок. Именно поэтому при идентификации важно рассматривать выявленные характеристики оперения в общей идентификационной схеме диагностических признаков, т.е. в комплексе с другими птилологическими признаками, что значительно облегчит процесс таксономического определения.
Выявленные признаки мы объединяем в идентификационную СДП, которая может быть использована для исследований в разных областях, в частности в авиационной орнитологии при расследовании случаев столкновений воздушных судов с птицами. Правильная идентификация останков повышает точность статистических данных по столкновениям и, таким образом, способствует повышению безопасности полетов. Подробные данные по таксону птицы позволят определить параметры риска и условия, с которыми может быть связано наибольшее число столкновений на соответствующем аэродроме. И тогда возможно принятие наиболее адекватных экологических мер и разработка наиболее действующих средств управления поведением птиц.
Все исследованные составляющие СДП требуют дальнейшего изучения для понимания, в какой степени их можно использовать в каждом из данных аспектов. При этом СДП уже успешно применяют для определения таксономической принадлежности птицы после столкновения с воздушным судном в лаборатории экологии и управления поведением птиц (единственной в России), где проводятся исследования определения таксона по микроструктуре пера. Теперь возможно определять семейство, а в ряде случаев и вид птицы по одному покровному перу, по части пера и по мелким обломкам перьев, например, при расследовании авиационных инцидентов. Извлеченные из двигателя перья выглядят примерно так, как это показано на рис. 6.
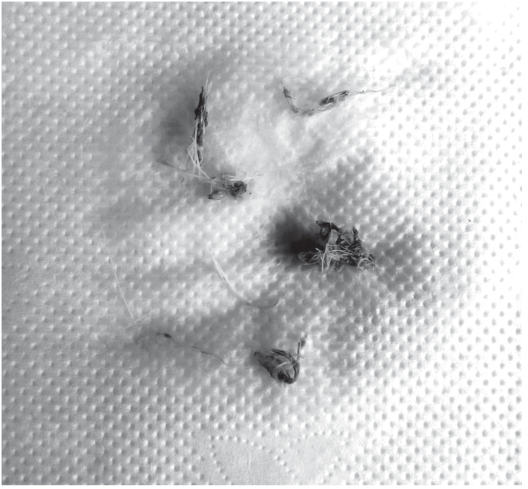
Опыт работы лаборатории по определению таксона птицы по микроструктуре подтверждает целесообразность использования каждого перьевого элемента в рамках системы диагностических признаков, а также насущную необходимость расширения составляющих этой системы.
Автор благодарит Ю.А. Богданову и Ю.А. Горохову (ИПЭЭ РАН) за помощь в микроскопировании.
Работа выполнена с использованием оборудования ЦКП “Инструментальные методы в экологии” при ИПЭЭ РАН.
Список литературы
Бокариус К.И. Экспертиза птичьего пуха // Практика криминалистической экспертизы. Киев: Высш. шк., 1961. Вып. 1. 94 с.
Бордонос Т.Г., Булыга Л.П. О некоторых особенностях строения перьев и пуха домашних животных // Криминалистическая и судебная экспертиза. Киев: Высш. шк., 1968. Вып. 5. С. 290–295.
Бородулина Т.Л. Строение кроющего оперения птиц в связи с их полетом // Зоол. журн. 1964. Т. 43. № 12. С. 1826–1836.
Добринский Л.Н. Географическая изменчивость варьирования морфологических признаков птиц и некоторые вопросы систематики // Внутривидовая изменчивость наземных позвоночных животных и микроэволюция. Свердловск: Урал. фил. АН СССР, 1966. С. 341–344.
Ильичев В.Д. Морфофункциональные особенности наружного уха птиц, ведущих сумеречный и ночной образ жизни // Докл. АН СССР. 1961. Т. 137. Вып. 6. С. 1485–1488.
Ильичев В.Д. Исследование тонкой структуры пера с помощью прибора ФМН‑2 // Зоол. журн. 1963. Т. 42. № 10. С. 1584–1585.
Силаева О.Л. Функциональное значение дополнительного пера в оперении птицы // В мире науч. открытий. 2011. № 4(16). С. 268–278.
Силаева О.Л., Вараксин А.Н. Внутривидовая изменчивость микроструктуры покровных перьев кряквы и хохлатой чернети // Междунар. науч.-исслед. журн. 2016. № 5(47). Ч. 5. С. 90–100.
Силаева О.Л., Горохова Ю.А. Дополнительные пуховые образования пера у птиц отряда Ржанкообразных (Сharadriiformes) // Междунар. науч.-исслед. журн. 2017. № 3(57). С. 20–26.
Силаева О.Л., Вараксин А.Н., Ильичев В.Д. Экспертиза перьевого материала с использованием методов анализа многомерных данных // Вестн. РУДН. 2011. № 4. С. 16–21.
Силаева О.Л., Ильичев В.Д., Чернова О.Ф. Определитель птиц по перу и его фрагментам. Отряд Воробьинообразные (Passeriformes), семейство Врановые (Corvidae). Саарбрюкен: LAP LAMBERT Acad. Publ., 2012. 316 с.
Силаева О.Л., Чернова О.Ф., Вараксин А.Н. Определитель птиц по перу и его фрагментам. Отряд Гусеобразные (Anseriformes). М.: ИПЭЭ РАН, 2015. Компакт-диск с цвет. ил. 269 с.
Силаева О.Л., Ильичев В.Д., Чернова О.Ф., Вараксин А.Н. Определитель птиц по перу и его фрагментам. Отряды: Курообразные (Galliformes), Голубеобразные (Columbiformes), Рябкообразные (Pterocletiformes). М.: ИПЭЭ РАН, 2013. 120 с. + CD-ROM.
Степанян Л.С. Конспект орнитологической фауны России и сопредельных территорий (в границах СССР как исторической области). М.: Академкнига, 2003. 808 с.
Чернова О.Ф., Ильяшенко В.Ю., Перфилова Т.В. Архитектоника перьев и диагностическое значение. М.: Наука, 2006. 98 с.
Чернова О.Ф., Перфилова Т.В., Фадеева Е.О., Целикова Т.Н. Атлас микроструктуры перьев птиц. М.: Наука, 2009. 163 ил. 173 с.
Яблоков А.В., Валецкий А.В. Изменчивость структур пера и окраски яиц у некоторых птиц // Зоол. журн. 1972. Т. 51. № 2. С. 248–258.
Brom T.G. The diagnostic and phylogenetic significance of feather structures. Published Ph.D. thesis. Amsterdam: Univ. Amsterdam, 1991. 279 p.
Chandler A.C. A study of the structure of feathers with reference to their taxonomic significance // Univ. Calif. Publ. 1916. V. 13. P. 243–446.
Dove C.J. Quantification of microscopic feather characters used in the identification of North American plovers // Condor. 1997. V. 99. № 1. P. 47–57.
Laybourne R.C. Collision between a vulture and an aircraft at an altitude of 37.000 feet // Wilson Bull. 1974. V. 86. P. 461–462.
Laybourne R.C., Dove C. Preparation of bird strikes remains for identification // Bird Strike Comm. Europe. 1994. V. 22. P. 531–534.
Laybourne R.C., Sabo B.A., Morningstar A. Basic techniques for preparation of down for examination with the scanning electron microscope // Auk. 1992. V. 109. № l. P. 195–197.
Lucas A.M., Stettenheim P.R. Avian anatomy. Integument.Washington: US Dept. Agricult., 1972. Pts 1, 2. 750 p.
Nitzsch Ch.L. System der Pterylographie. Halle: Eduard Anton, 1840. 226 S.
Prast W., Shamoun J. BRIS: A computer based bird remains identification system. Further developments // Birds Europe. CD‑ROM. Amsterdam: ETI., 1996.
Prast W., Shamoun J. Bird Remains Identification System (BRIS). Birds of Europe. CD-ROM, Amsterdam: ETI., 2001.
Rochard J.B.A., Horton N. Birds killed by aircraft in the United Kingdom 1966–76 // Bird Strike Comm. Europe. 1977. V. 12. P. 227–234.
Van Tyne J. Fundamentals of Ornithology. N.Y.: J. Wiley and Sons., Inc., 1959. 642 p.
Ziswiler V. Die Afterfeder der Voegel. Untersuchungen zur Morphogenese und Phylogenese des sogenannten Afterschaftes // Zool. Jahrb. Abt. F. Anat. 1962. Bd 80. S. 245–308.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая