

Известия РАН. Серия биологическая, 2021, № 3, стр. 241-248
Оогенез (профаза 1 мейоза) и митотические хромосомы партеногенетического вида Darevskia armeniaca (семейство Lacertidae)
Л. А. Куприянова 1, Л. Д. Сафронова 2, В. Б. Сычева 2, Ф. Д. Даниелян 3, В. Г. Петросян 2, *
1 Зоологический институт РАН
199034 Санкт-Петербург, Университетская наб., 1, Россия
2 Институт проблем экологии и эволюции им. А.Н. Северцова, РАН
119071 Москва, Ленинский просп. 33, Россия
3 Ереванский государственный университет
0025 Ереван, ул. Алека Манукяна, 1, Армения
* E-mail: vgpetrosyan@gmail.com
Поступила в редакцию 06.11.2019
После доработки 30.01.2020
Принята к публикации 23.05.2020
Аннотация
Впервые проведен анализ течения раннего оогенеза и мейоза партеногенетического вида Darevskia armeniaca. Показано, что число бивалентов синаптонемных комплексов (СК) на стадиях ранней пахитены–диплотены профазы 1 мейоза равно 19, на основе которых представлены СК-кариотипы. Приведены результаты сравнительного молекулярно-цитогенетического анализа митотических хромосом (2n = 38, 34A + 2m + Zw, половые хромосомы) партеногенетических самок и мейотических хромосом (n = 19 бивалентов) мужской особи D. armeniaca с инверсией пола. Отмечено отсутствие механизма эндоредупликации хромосом в премейотических митозах D. armeniaca, прохождение ранних стадий мейоза и формирование гаплоидного числа СК-бивалентов гомеологичных хромосом.
Открытие значительного числа рыб, амфибий и рептилий, размножающихся посредством гино-, гибридо- и партеногенеза, поставило перед исследователями ряд серьезных вопросов, в частности вопрос о цитологических особенностях оогенеза однополых видов. В первых работах было показано, что у таких форм в процессе оогенеза осуществляется мейоз и механизмы восстановления соматического числа хромосом у однополых позвоночных и беспозвоночных часто едины. Классификации механизмов диплоидизации отличаются одна от другой по цитологической картине нарушений оогенеза или по их генетическому эффекту (пре-, внутри- и постмейотический типы мейоза). Генетические последствия перечисленных типов мейоза различны, поэтому от механизмов восстановления соматического числа хромосом у однополых особей будут зависеть результаты мейоза и судьбы особей в целом. Эти последствия приобретают еще большее значение, если учесть, что в эволюционном отношении однополое размножение связано с гибридизацией и полиплоидией.
Сохранение и поддержание высокой степени гетерозиготности гибридов возможно при премейотическом типе диплоидизации, а именно при подавлении цитокинеза в последнем премейотическом митозе. Более того, этот механизм позволяет избегать многие трудности, связанные с синапсисом гомеологов гибрида и расхождением псевдобивалентов в течение мейоза гибридных триплоидных форм. По-видимому, именно этими обстоятельствами объясняется распространение этого механизма у однополых позвоночных и обнаружение такого типа мейоза у североамериканских партеногенетических гибридных диплоидных (2n) и триплоидных (3n) ящериц рода Aspidoscelis семейства Teiidae (Cuerllar, 1971; Lutes et al., 2010). У гибридных диплоидных партеногенетических cамок этого рода в мейоз входит тетраплоидная (4n) клетка, и в ходе профазы 1 мейоза на стадиях пахитены–диплотены в ооците происходит спаривание сестринских хромосом и формирование диплоидного числа псевдобивалентов (Lutеs et al., 2010). Однако в мейозе гибридных гиногенетических рыб Carassius gibelio такой механизм отсутствует (Черфас, 1969).
Кавказские партеногенетические ящерицы рода Darevskia семейства Lacertidae также имеют гибридное происхождение (Moritz et al., 1992b) и в отличие от североамериканских партеногенетических видов всегда характеризуются типичным для семейства диплоидным (2n) числом акроцентрических (А) хромосом, равным 38 с гетероморфными половыми Zw хромосомами (2n = 38А, Zw).
В предшествующих работах в ядрах ооцитов гибридных партеногенетических самок этого рода были изучены стадии ранней пахитены–диплотены–диакинеза и обнаружены фигуры, образованные гомеологичными хромосомами с концевыми хиазмами в проксимальных и дистальных районах мейотических хромосом (Cuellar, 1971; Kupriyanova, 1992, 1994, 2010). Кроме того, при анализе синаптонемных комплексов (СК) число бивалентов приближалось к 19, т.е. к гаплоидному числу (n = 19). Эти и другие косвенные факты позволяли допустить отсутствие механизма эндоредупликации хромосом в премейотическим митозе и возможность рекомбинации генетического материала, а следовательно, возникновение генетического разнообразия за счет рекомбинации (Kupriyanova, 1992, 1994, 2010; Куприянова, 1997).
Очевидно, что цитогенетические механизмы мейоза ключевые в эволюции партеногенеза гибридных видов. Для более детального описания и уточнения механизмов раннего мейоза гибридных партеногенетических видов рода Darevskia мы исследовали ооциты вида Darevskia armeniaca (Méhely, 1909), проанализировали структуру элементов мейотического ядра и морфологию СК-хромосом, на основании которых составили СК-кариотипы хромосом гибридного ядра. Такие СК-кариотипы были сопоставлены с полученными ранее митотическими хромосомами D. armeniaca, а также с характеристиками профазы 1 мейоза, а именно с хромосомами на стадии диакинеза у редких диплоидных мужских особей D. armeniaca с нарушенной фертильностью (Darevskii, Kupriyanova, 1982; Kupriyanova, 2010).
МАТЕРИАЛЫ И МЕТОДЫ
Самки D. armeniaca были собраны в разные годы в районах Армянских городов Цахкадзора (40°32′7′′ с.ш., 044°42′30′′ з.д.) и Дилижана (40°44′2′′ с.ш., 044°49′4′′ з.д.). Характеристики точек находок и современные ареалы партеногенетического вида D. armeniaca и родительских видов D. valentini, D. mixta были описаны ранее (Petrosyan et al., 2019а, b; Petrosyan et al., 2020).
Мейотические хромосомы. Для микроскопического анализа СК-препараты тотальных распластанных ядер ооцитов готовили из клеточной суспензии, полученной из зародышевой полости яичника самки методом Дрессера и Мозеса (Dresser, Moses, 1980). Распластывание (спредирование) ооцитов проводили на капле 0.5%-ного раствора NaCl. Препараты фиксировали 4%-ным параформальдегидом, а для визуализации СК их окрашивали 50%-ным раствором нитрата серебра, затем просматривали и фотографировали под световым микроскопом Leica (Германия). Длины СК-бивалентов ооцитов измерялись с помощью программы Leica Application Suite.
Митотические хромосомы. В работе использовали метод получения митотических хромосом с предварительным введением 0.1%-ного раствора фитогемагглютинина (ФГА P ПанЭко (Россия); 0.03 мл раствора на 10 г массы) и колхицина (Merck (Германия); 0.1 мл на 10 г массы). Препараты окрашивали красителем Giemsa по стандартной методике и методом дифференциального сравнительного С-окрашивания с последующим окрашиванием хромосом красителем, специфичным к АТ-парам оснований флуорохромом ДАПИ. Препараты просматривали и фотографировали под световым микроскопом Leica, длины хромосом измерялись с помощью программы Leica Application Suite.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
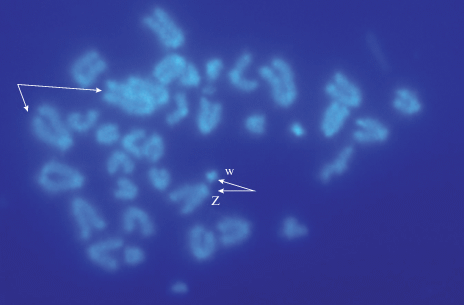
В кариотипе исследованных самок D. аrmeniaca присутствуют 38 акроцентрических (A) хромосом, среди которых 2 микрохромосомы (m) и половые Zw (2n = 38A: 34 A + 2m + Zw; n = 19). Сравнительное флуорохромное дифференциальное (С/СМА3/ДАПИ) окрашивание метафазных хромосом D. аrmeniaca показало, что большинство хромосом набора имеют мелкие блоки АТ-повторов в прицентромерных и GC-повторов в теломерных районах C-гетерохроматина (рис. 1, 2). Вместе с тем молекулярно-цитогенетический анализ выявил различия в структуре некоторых хромосом – гетероморфизм теломерных районов С-гетерохроматина хромосом. Например, одна первая хромосома первой пары кариотипа имеет в этих районах крупные яркие блоки, содержащие GC-повторы, в то время как у другой хромосомы первой пары эти блоки выражены значительно слабее. Яркий С-блок, включающий в себя GC-пары, присутствует также и в половой w-микрохромосоме, хотя по размеру и морфологии она практически не отличается от двух других микрохромосом кариотипа (рис. 1). Кроме того, одна макрохромосома D. аrmeniaca отличается по морфологии от других хромосом набора, поскольку она имеет короткие плечи и небольшие блоки, обогащенные АТ- и GC-повторами, в околоцентромерных и теломерных районах соответственно. По размерам эта непарная хромосома относится к средней группе хромосом, к пятой-шестой паре, и рассмотрена как половая Z-хромосома (рис. 1, 2). Важно отметить, что у разных видов этого семейства половая Z-хромосома пятой-шестой пары часто имеет короткие плечи, что объясняют ее нестабильностью. Ранний мейоз (профаза 1 мейоза) партеногенетического вида D. armeniaca (2n = 38А) изучался на препаратах распластанных ядер ооцитов размером 0.8–1.1 мм.
Рис. 1.
Метафазные пластинки клеток крови Darevskia armeniaca (самка) – С-исчерченность гетерохроматина после окрашивания специфичным G-C-флуорохромом (СМА3) (Kupriyanova, 2010, с изменениями) 2n = 38 A: 34A + 2m + Zw (половые хромосомы). Стрелки указывают на гетероморфные ауто- (первая пара) и половые хромосомы Z и w (пятая пара).
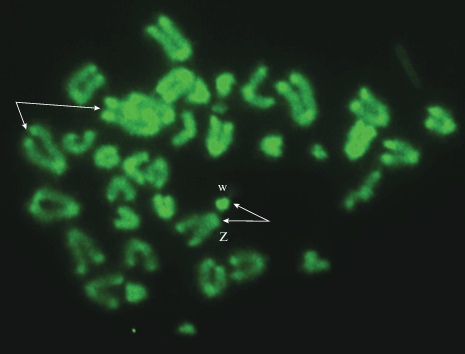
Рис. 2.
Метафазные пластинки клеток крови Darevskia armeniaca (самка) – С-исчерченность гетерохроматина после окрашивания специфичным АТ-флуорохромом (ДАПИ) 2n = 38 A: 34A + 2m + Zw (половые хромосомы). Стрелки указывают на гетероморфные и половые Z- и w-хромосомы.
Анализ СК на стадиях ранней пахитены и диплотены показал, что они полностью синаптированы и сформировали 19 СК-бивалентов. Этот анализ профазы мейоза проведен на пяти ядрах овоцитов (рис. 3a–г). Однако распластанность СК-бивалентов (комплекса) не происходит аналогично, как было показано при сперматогенезе (Сафронова, Куприянова, 2016), поскольку ядра овоцитов погружены в жидкую среду с большим количеством жира. Этот процесс препятствует хорошему распластыванию на стекле, что приводит к волнистости, а в некоторых случаях – к закручиванию концов. На рис. 3 нумерация СК-бивалентов представлена в порядке убывания их линейных размеров проведенных измерений. СК-бивалент № 1, сформированный первыми по размеру и гетероморфными по структуре хромосомами набора (первый СK), отличался по морфологии от остальных СК-бивалентов. Следует особо подчеркнуть, что на всех изученных препаратах распластанных ядер ооцитов число СК-бивалентов всегда равно гаплоидному числу хромосом вида, т.е. 19.

ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В ряде предшествующих комплексных работ было показано (Kupriyanova, 1989, 1992, 2010), что гибридный партеногенетический вид D. аrmeniaca (D. mixta х D. valentini), как и другие однополые и обоеполые виды рода Darevskia семейства Lаcertidae, характеризуется 38 одноплечими по морфологии и близкими по размерам акроцентрическими (А) хромосомами и стабильной структурой кариотипа (2n = 38 А, NF (основное число) = 38). Кроме того, этот вид, как и два других гибридных партеновида (D. dahli, D. unisexualis) рода Darevskia, имеет сходные числа и морфологию половых Zw-хромосом. В результате гибридизации самки D. аrmeniaca могли получить Z-хромосому от отцовского D. valentini и w-хромосому от материнского D. mixta родительских видов (Kupriyanova, 1992; Moritz et al., 1992а; Murphy et al., 2000). Для кариотипов Lаcertidae характерна локализация структурного гетерохроматина (С-окрашивание) в прицентромерных и теломерных районах хромосом и редко в интеркалярных районах (Olmo et al., 1986; Kupriyanova, 1994). Такое единообразие хромосом ограничивает возможность использования признаков кариотипа при анализе вопросов происхождения этих гибридных видов, процесса мейоза и их эволюции в целом. Именно по этой причине идентификация хромосом видов рода Darevskia в целом до сих пор – трудная задача. Однако эти результаты свидетельствовали и о цито- и генетической близости родительских видов рода Darevskia (Kupriyanova, 2010; Куприянова, 2014).
Тем не менее молекулярно-цитогенетический анализ одного из родительских видов для D. rostombekowi этого рода, D. raddei, продемонстрировал присутствие в кариотипе дицентрической хромосомы (Spangenberg et al., 2019). Результаты данной работы выявили некоторые различия в молекулярно-цитогенетической структуре теломерных районов хромосом D. аrmeniaca. К настоящему времени известно, что амплификация некоторых генов и образование тандемно расположенных высококопийных повторов, например, в прицентромерных районах хромосом – это частое геномное нарушение, встречающееся у гибридов. Прицентромерные и теломерные районы часто имеют ключевое значение для пространственной ориентации хромосом в ядре и очень важны для совпадения мест связи хромосом гибрида с ядерной оболочкой, а также для конъюгации хромосом в процессе мейоза. Крупная хромосомная реорганизация, системные мутации играют существенную роль в процессе сальтационного видообразования и затрагивают ранние этапы развития особи (Стегний, 2019).
Проведенный анализ четырех клеток, находящихся на стадии ранней профазы 1 мейоза, свидетельствует о том, что в раннем оогенезе партеногенетического гибридного вида D. armeniaca отсутствуют полиплоидные ооциты и, следовательно, механизм эндоредупликации хромосом в последних митотических делениях. Все полученные данные свидетельствует о том, что в раннем оогенезе диплоидные ядра ооцитов самки D. armeniaca (2n = 38) вступают в ранний мейоз (в профазу 1 мейоза) и на стадии ранней пахитены–диплотены формируют 19 СК-бивалентов. Картины раннего мейоза еще раз демонстрируют генетическую близость родительских видов гибридного однополого D. armeniaca.
Сравнительное дифференциальное флуорохромное (С/СМА3/ДАПИ) окрашивание хромосом D. armeniaca позволило также еще раз рассмотреть половые Zw-хромосомы видов рода Darevskia. Известно, что половая Z-хромосома ряда видов Lаcertidae отнесена по размеру к пятой-шестой хромосоме и часто имеет короткие плечи. Например, согласно молекулярно-цитогенетическому картированию хромосом Z-половая хромосома Lacerta agilis (2n = 38, Zw) отнесена по длине к пятой паре кариотипа и имеет частичную гомологию с шестой и девятой хромосомами курицы (Skikulnath et al., 2014). Предположительно пятая-шестая по длине половые хромосомы Z1 и W были обнаружены в криптической группе Zootoca vivipara (Куприянова, Руди, 1990; Odierna et al., 1998; Куприянова, Беме, 2012). Непарная пятая-шестая по длине хромосома D. аrmeniaca интерпретирована как половая Z-хромосома. Отметим, что половая Z-хромосома видов родов Takydromus, Gallotia, Eremias отнесена к средней размерной группе кариотипа, 12–13 по длине (Olmo et al., 1986; Lisachov et al., 2019). Перечисленные факты ясно указывают на трудности в решении вопросов идентификации половых хромосом лацертидных ящериц и на необходимость дальнейшего детального сравнительного анализа половых хромосом разных групп семейства и ящериц в целом.
Следует напомнить, что предшествующие цитогенетические и молекулярно-цитогенетические работы убедительно продемонстрировали существенную роль половых Zw-хромосом в филогенетических ограничениях возникновения партеногенеза в роде Darevskia (Kupriyanova, 1989, 1992, 2010; Куприянова, 1997, 1999). Эти факты указывали также на то, что “балансовая” гипотеза перехода гибрида к однополому способу размножения, предложенная ранее (Moritz et al., 1989, 1992a), не учитывала все факторы возникновения однополых ящериц (Куприянова, 1997, 1999).
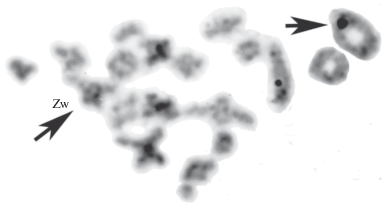
В связи с тем значением, которое имеют половые Zw-хромосомы в успешной гибридизации ящериц рода Darevskia и в переходе возникающих гибридов к партеногенетическому типу мейоза, мы вновь проанализировали опубликованные ранее данные о кариотипе редкой мужской особи партеногенетического гибридного вида D. аrmeniaca (Darevskii, Kupriyanova, 1982). В отличие от многочисленных триплоидных гибридов, появляющихся в симпатрических популяциях между партеногенетическими и обоеполыми видами, редкий самец был отловлен в “чистой” популяции D. аrmeniaca (район г. Степанаван, Армения). Хромосомные исследования показали полное совпадение кариотипа самца с таковым у самок этой и других популяций вида: диплоидное число хромосом 2n = 38, среди которых присутствовали половые Zw-хромосомы (рис. 4). В результате был сделан вывод о нарушении взаимодействия половых Zw-хромосом в гибридном геноме D. аrmeniaca и об инверсии пола у Zw-особи. Одновременно были установлены цитологические нарушения в течение мейоза особи: в расхождении гомеологов в мейотическом делении I и в образовании анеуплоидных сперматоцитов II порядка, сперматид и сперматозоидов и в нарушении процессов спермиогенеза. В течение мейоза особи на стадии диакинеза профазы 1 мейоза были сформированы 19 бивалентов. Среди них выделен пятый-шестой по размеру бивалент, предположительно образованный половыми хромосомами, и отмечены особенности его морфологии. Большой элемент “бивалента”, вероятно, представлен крупной Z-хромосомой и небольшой элемент, вероятно, w-микрохромосомой (рис. 5). Плотной конъюгации этих хромосом не наблюдается, можно отметить ассоциацию концов w микрохромосомы с интеркалярным районом Z хромосомы и значительный размер w микрохромосомы, возможно за счет ее деконденсации.
Рис. 4.
Метафазные пластинки клеток крови Darevskia armeniaca (самец) – стандартное окрашивание Giemsa (Darevskii, Kupriyanova, 1982, с изменениями) 2n = 38 A: 34A + 2m + Zw (половые хромосомы). Стрелка указывает на половую w-микрохромосому.

Рис. 5.
Мейотические пластинки гонад D. armeniaca (самец) – диакинез, стандартное окрашивание Giemsa (Darevskii, Kupriyanova, 1982, с изменениями) n = 19. Стрелки указывают на первый (меньшего размера) бивалент и пятый (большего размера) половой бивалент, образованный предположительно половыми хромосомами Z и w.
Следует напомнить, что определенный спектр цитогенетических, геномных и функциональных нарушений был установлен и у гибридных партеногенетических самок рода Darevskiа. В связи с этим был сделан важный вывод, что, несмотря на цито- и генетическую близость родительских видов, у гибридных партеногенетических видов наблюдается нестабильность их гибридных геномов. В результате “гибридная” нестабильность и рекомбинационный обмен в профазе 1 мейоза могут служить механизмами генетической изменчивости партеногенетических видов рода (Kupriyanova, 1989, 1992, 2010; Куприянова, 1997, 1999, 2014). Например, обнаруженные у D. аrmeniaca редкие аллели по белковым системам (MacCullach et al., 1995), вариабельность и мутации определенного типа по локусам микросателлитной ДНК этих партеногенетических видов (Ryskov, 2008; Vergun et al., 2014; Girnyk et al., 2018) могут быть объяснены указанными цитогенетическими механизмами (Kupriyanova, 2010; Куприянова, 2014).
ЗАКЛЮЧЕНИЕ
Полученные данные о раннем оогенезе у самок гибридного партеногенетического вида D. armeniaca (2n = 38) свидетельствуют о типичном течении ранних стадий мейоза и прохождении ранней профазы 1 мейоза. На стадии пахитены–диплотены происходит формирование 19 СК элементов и позднее 19 бивалентов гомеологичных хромосом. Это свидетельствует о том, что в мейоз вступает ооцит с диплоидным числом хромосом. Таким образом, полученные факты подтверждают немногочисленные сведения о том, что механизмы мейоза у партеногенетических ящериц родов Aspidoscelis и Darevskia различны. У ящериц рода Darevskia в ходе оогенеза в последних премейотических митозах не происходит эндоредупликации хромосом и, следовательно, удвоения числа хромосом до 4n. В течение раннего мейоза в диплоидном ооците на стадии пахитены–диплотены образуется гаплоидное число СК бивалентов (n = 19). Полученные данные вновь подтверждают цито- и генетическую близость родительских видов гибридного партеновида D. armeniaca, образование в мейозе СК-бивалентов гомеологичных хромосом и возможность рекомбинационных обменов. Вместе с тем они указывают на гетероморфность в цитогенетической структуре некоторых аутохромосом и половых митотических хромосом, особую морфологию их СК-бивалентов и у переопределенного самца – на специфическую морфологию и структуру полового бивалента с “деконденсированной” w-хромосомой и с типом ассоциации половых хромосом. Разные типы полоопределяющих механизмов отмечены у ящериц: у Aspidoscelis самки характеризуются гомоморфными ХХ-половыми хромосомами, тогда как у Darevskia – гетероморфными Zw-половыми хромосомами.
Цитологические характеристики и особенности мейоза более поздних стадий оогенеза D. аrmeniaca и других партеногенетических видов рода Darevskia до конца не исследованы. Механизмы восстановления диплоидного числа хромосом у кавказских партеногенетических видов, а также механизмы поддержания их гетерозиготности и системы половых Zw-хромосом – цель дальнейших исследований авторов.
Авторы выражают благодарность А.П. Рыскову за ценные замечания, внимание к работе и оказанную помощь при подготовке рукописи.
Работа выполнена при финансовой поддержке ЗИН РАН (грант № АААА-А19-119020590095-9) и РФФИ (гранты 17-00-00427, 17-00-00430).
Список литературы
Куприянова Л.А. Некоторые цитогенетические закономерности сетчатого видообразования однополых видов ящериц (Reptilia, Lacertidae) и других групп позвоночных животных // Цитология. 1997. Т. 39. № 12. С. 1089–1108.
Куприянова Л.А. Генетическое разнообразие гибридных у однополых видов и форм рода Lacerta (Reptilia, Lacertidae): его возможные цитогенетические механизмы, цитогенетика мейоза природных полиплоидных форм // Цитология. 1999. Т. 41. № 12. С. 1038–1047.
Куприянова Л.А. Концепция гибридогенного видообразования позвоночных животных: комплексные исследования однополых видов рептилий // Тр. ЗИН РАН. 2014. Т. 318. № 4. С. 382–390.
Куприянова Л.А., Бёме В. Живородящая ящерица (Zootoca vivipara (Lichtenstein,1823), Lacertidae) из северо-восточных и центральных районов Европы: внутривидовое кариотипическое разнообразие // Зоол. журн. 2012. Т. 91. № 11. С. 1428–1432.
Куприянова Л.А., Руди Е.Р. Сравнительно-кариологический анализ популяций живородящей ящерицы (Lacerta vivipara, Lacertidae, Sauria) // Зоол. журн. 1990. Т. 69. № 6. С. 93–101.
Сафронова Л.Д., Куприянова Л.А. Метафазные и мейотические хромосомы, синаптонемные комплексы (СК) живородящей ящерицы (Zootoca vivipara, Lacertidae) // Генетика. 2016. Т. 52. № 11. С.1311–1317.
Стегний В.Н. Генетика сальтационного видообразования и системные мутации. Томск: Издательский дом ТГУ, 2019. 264 с.
Черфас Н.Б. Основные итоги цитогенетического анализа однополой и двуполой форм серебряного карася // Генетика, селекция и гибридизация рыб. М.: Наука, 1969. С. 85–97.
Cuellar O. Reproduction and the mechanism of meiotic restitution in the parthenogenetic lizard Cnemidophorus uniparens // J. Morph. 1971. V. 133. № 2. P. 139–165.
Darevskii I.S., Kupriyanova L.A. Rare males in parthenogenetic lizard Lacerta armeniaca Méhely // Vertebr. Hung. 1982. V. 21. P. 69–75.
Dresser M., Moses M. Synaptonemal complex karyotyping in spermatocytes of the Chinesehamster (Cricetulus gricus). IV. Light and electron microscopy of synapsis and nucleolar development by silver staining // Chromosoma. 1980. V. 76. P. 1–22.
Girnyk A., Vergun A., Semyenova S.K., Guliaev A., Arakelyan M., Danielyan F., Martirosyan I., Murphy R., Ryskov A.P. Multiple interspecific hybridization and microsatellite mutations provide clonal diversity in the parthenogenetic rock lizard Darevskia armeniaca // BMC Genomics. 2018. V. 19. № 979. DOI.org/10.1186/s12864=018-5359-5.
Kupriyanova L. Cytogenetic evidence on genome interaction in hybrid Lacerta. Evolution and ecology of unisexual vertebrates // Bull. New York State Museum. Albany. 1989. V. 466. P. 236–239.
Kupriyanova L. Diversity in parthenogenetic lacertid lizards: cytogenetic studies // Proc. 6th Ordinary General Meeting of the Societas Europaea Herpetologica. Budapest: SHE, 1992. P. 273–279.
Kupriyanova L. Structure, localization and stability of chromosome in karyotype evolution in lizards of the Lacertidae family // Rus. J. Herpetol. 1994. V. 1. P. 1–12.
Kupriyanova L. Cytogenetic and genetic trends in the evolution of unisexual lizards // Cytogen. Gen. Res. 2010. V. 127. P. 273–279.
Lisachov A.P., Galkina S.A., Saifitdinova A.F., Romanenko S.A., Andreyushkova D.A., Trifonov V.A., Borodin P.M. Identification of sex chromosomes in Eremias velox (Lacertidae, Reptilia) using lampbrush chromosome analysis // Comp. Cytogen. 2019. V. 13. №. 2. P. 121–132. https://doi.org/10.3897/CompCytogen.v13i2.34116
Lutes A.A., Neaves W.B., Baumann D.P., Wiegrabe W., Baumann P. Sister chromosome pairing maintains heterozygosity in parthenogenetic lizards // Nature. 2010. V. 11. № 464(7286). P. 283–286.
MacCulloch R.D., Murphy R.W., Kupriyanova L.A., Darevsky I.S., Danielyan F.D. Clonal variation in the parthenogenetic rock lizard Lacerta armeniaca // Genome. 1995. V. 38. P. 1057–1060.
Moritz C., Wright J.W., Brown C.M. Mitochondrial DNA analysis and the origin and relative age of parthenogenetic Cnemidophorus: phylogenetic constraints on hybrid origins // Evolution. 1992a. V. 46. P. 184–192.
Moritz C., Uzzell T., Spolsky C., Hotz H., Darevsky I., Kupriyanova L., Danielyan F. The material ancestry and approximate age of parthenogenetic species of Caucasian rock lizards (Lacerta: Lacertidae) // Genetica. 1992b. V. 87. P. 53–62.
Moritz C., Brown W.M., Densmore L.D., Wright J., Vyas D., Donnellan S., Adams M., Baverstock P. Genetic diversity and the dynamics of hybrid parthenogenesis in Cnemidophorus (Teiidae) and Heteronotia, (Gekkonidae) // Bull. New York State Museum. Albany. 1989. V. 466. P. 87–112.
Murphy R., Darevsky I., Kupriyanova L., MacCulloch R., Fu J. A fine line between sex and unisexuality: the phylogenetic constraints on parthenogenesis in lacertid lizards // Zool. J. Linn. Soc. 2000. V. 130. P. 527–549.
Odierna G., Aprea G., Capriglione T. Progressive differentiation of the W sex chromosome between oviparous and viviparous populations of Zootoca vivipara (Reptilia, Lacertidae) // Ital. J. Zool. 1998. V. 65. P. 295–302.
Olmo E., Odierna G., Cobror O. C-band variability and phylogeny of Lacertidae // Genetica. 1986. V. 71. P. 63–74.
Petrosyan V.G., Osipov F.A., Bobrov V.V., Dergunova N.N., Danielyan F.D., Arakelyan M.S. New records of Darevskia armeniaca (Méhely, 1909) and Darevskia valentini (Boettger, 1892) (Squamata, Sauria, Lacertidae) from Armenia and updated geographic distribution maps // Check List. 2019a, V. 15. P. 21–31.
Petrosyan V., Osipov F., Bobrov V., Dergunova N., Kropachev I.I., Danielyan F., Arakelyan M. New records and geographic distribution of the sympatric zones of unisexual and bisexual rock lizards of the genus Darevskia in Armenia and adjacent territories // Biodiversity Data Journal, 2020. № 8. e56030.https://doi.org/10.3897/BDJ.8.e56030.
Petrosyan V., Osipov F., Bobrov V., Dergunova N., Nazarenko E., Omelchenko A., Danielyan F., Arakelyan M. Analysis of geographical distribution of the parthenogenetic rock lizard Darevskia armeniaca and its parental species (D. mixta, D. valentini) based on ecological modeling // Salamandra, 2019b. V. 55. № 3. P. 173–190.
Ryskov A.P. Genetically unstable microsatellite containing loci and genome diversity in clonally reproduced unisexual vertebrates / International Review of Cell and Molecular Biology / Ed. Jeon K.W. N.Y. Acad. Press, 2008. V. 270. P. 319–349.
Skikulnath K., Matsubara K., Uno Y., Nishida C., Olsson M., Matsuda Y. Identification of the linkage group of the Z sex chromosomes of the sand lizard (Lacerta agilis, Lacerttidae) and elucidation of karyotype evolution in lacertid lizards // Chromosoma. 2014. V. 123. P. 563–575.
Spangenberg V., Arakelyan M., Galoyan E., Pankin M., Petrosyan R., Stepanyan I., Grishaeva T., Danielyan F., Kolomiets O. Extraordinary centromeres: differences in the meiotic chromosomes of two rock lizards of species Darevskia portschenskii and Darevskia raddei. 2019. http://dx.doi.org/10.7717/peerj.6360
Vergun A.A., Martirosyan I.A., Semyenova S.K., Omelchenko A.V., Petrosyan V.G., Lazebny O.E., Tokarskaya O.N., Korchagin V.I., Ryskov A.P. Clonal diversity and clone formation in the parthenogenetic caucasian rock lizard Darevskia dahlia // PLoS One. 2014. e91674. https://doi.org/10.1371/journal.pone.0091674
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая