

Известия РАН. Серия биологическая, 2021, № 4, стр. 389-398
История растительности ближних островов алеутской гряды в голоцене
О. И. Смышляева 1, *, Е. Э. Северова 2, Б. Ф. Хасанов 1, О. А. Крылович 1, Е. А. Кузьмичева 1, А. Б. Савинецкий 1
1 Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский пр-т, 33, Россия
2 Московский государственный университет им. М.В. Ломоносова, биологический факультет
119191 Москва, Ленинские горы, 1, стр. 12, Россия
* E-mail: smyslyaevaol@gmail.com
Поступила в редакцию 24.08.2020
После доработки 04.09.2020
Принята к публикации 17.11.2020
Аннотация
Реконструирована история формирования растительности торфяника Алкан Крик на острове Шемья (Ближние острова Алеутской гряды). На плоском острове, лишенном вулканов, ~7000 лет назад преобладала осоковая тундра, которая сменилась болотом с заметным участием Sphagnum и Equisetum в период максимального голоценового потепления 7000–5000 лет назад. Разнотравные осоково-злаковые луга сформировались на месте болота 4700 лет назад в результате локального изменения увлажнения. В результате похолодания и процессов выщелачивания ~ 3400 лет назад на острове стали преобладать вересковые тундры.
Исследования истории растительного покрова в голоцене необходимы для анализа путей его формирования, современного состояния и путей будущего развития, они дают ключи к пониманию возможного ответа растительности на глобальные климатические изменения. Основным методом исследований истории растительности в региональном масштабе был и остается спорово–пыльцевой анализ аллювиальных, озерных и болотных отложений. Подкрепленный данными об абсолютном возрасте образцов этот метод дает обоснованную картину изменения растительности и климата прошлого.
Данная работа проведена на Алеутских островах – архипелаге вулканического происхождения, протяженностью почти 2000 км от Аляски до Камчатки. Для выбранного региона характерны мягкие зимы, прохладное лето, небольшой перепад среднегодовых температур, частые туманы, сильная облачность и высокая влажность (Byers, 1959). Атмосферная циркуляция и климатические изменения определяются в регионе, в основном, динамикой Алеутского минимума – области низкого атмосферного давления в районе Алеутских островов (Rodionov et al., 2007). Растительность островов лишена деревьев и может быть охарактеризована как арктическая тундра. Флора представлена преимущественно тихоокеанскими прибрежными видами, также многочисленны аркто-альпийские виды, распространенные циркумполярно (Hulten, 1968).
Историю растительности Алеутской гряды начали изучать в середине прошлого века, когда был проведен спорово-пыльцевой анализ погребенных почв на островах Танага и Уналашка (Anderson, Bank, 1952). Спектры почв с разных глубин показали, что в голоценовой истории о. Танага произошла как минимум одна смена растительных сообществ с вересковыми кустарничками на травяные сообщества. На данный момент изучены торфяники на островах Умнак (Heusser, 1973), Карлайл (Kuzmicheva et al., 2019), Атка (Heusser, 1990), две колонки из торфяника на о. Адак (Heusser, 1978; Noguchi et al., 2018), торфяник на о. Атту (Heusser, 1990) и на о. Беринга (Кирьянов и др., 1986). Обобщив полученные для разных частей гряды результаты, Хессер (Heusser, 1990) выдвинул предположение, что основным фактором, влиявшим на смены растительности в голоцене, был климат. При этом климатические закономерности могли отличаться в западной, центральной и восточной частях Алеутской гряды. Также Хессер (Heusser, 1990) обратил внимание на то, что выпадавшие в результате извержений вулканические пеплы незначительной толщины могли воздействовать на растительность, вызывая скорее изменения участия видов в сообществах, но не приводя к значительной смене их состава. Пеплы, выпадавшие в результате крупных извержений, могли практически полностью уничтожать растительные сообщества (Talbot et al., 2010). Вулканическая активность на западных островах в голоцене, предположительно, была слабее, чем в остальной части архипелага.
В данной работе для реконструкции динамики растительности западной части Алеутских островов был выбран плоский о. Шемья (Ближние Алеутские острова), лишенный вулканов. В отобранной для спорово-пыльцевого анализа колонке из торфяника Алкан Крик отсутствовали заметные пепловые слои, что позволило проследить независимое от вулканических извержений влияние климатических фаз на растительные сообщества острова в голоцене.
МАТЕРИАЛЫ И МЕТОДЫ
Остров Шемья (Ближние острова) представляет собой плоский, лишенный вулканов остров, расположенный на 52°43′27′′ с.ш. и 174°07′08′′ в.д., его длина составляет ~4.5 км, ширина – почти 7 км, а площадь – 15 км2. Современная растительность острова, в основном, представлена сообществами двух типов (Киселева и др., 2002). Растительность с доминированием злаков, осок и значительным участием разнотравья можно встретить в долинах ручьев, низинах и на склонах различных экспозиций. Наибольшего разнообразия такие сообщества достигают на хорошо дренированных склонах южной экспозиции. Второй тип сообществ – мохово-кустарничковые со значительным участием вересковых (Empetrum nigrum, Vaccinium vitis-idea, Linnea borealis), а также злаков и осок – встречается в разнообразных местообитаниях (Киселева и др., 2002). Кроме этих двух типов, на холмах недалеко от берега моря можно встретить сомкнутую растительность лугового типа с доминированием Elymus arenarius, Heracleum lanatum и Senecio pseudo-arnica (Savinetrsky et al., 2010). В условиях переувлажнения доминантами растительных сообществ являются осоки (Киселева и др., 2002).
В 1994 г. в рамках работы WAASP (Западно-Алеутский археологический и палеоэкологический проект) была отобрана колонка из торфяника Алкан Крик глубиной 270 см (рис. 1, табл. 1), слоями толщиной 10 см. Торфяник расположен в долине ручья, в северо-западной части острова, в 750 м от берега моря, на высоте 35 м н. у. м. (Savinetsky et al., 2010). Для определения возраста и скорости накопления отложения на радиоуглеродное датирование были отобраны растительные остатки из пяти слоев (табл. 2). Образцы обрабатывали по методике, описанной в работе Киселевой с соавт. (Киселева и др., 2002). Датирование проводили сцинтилляционным методом в лаборатории исторической экологии Института проблем экологии и эволюции им. А.Н. Северцова. Радиоуглеродные даты были откалиброваны для определения календарного возраста и построения модели роста отложения (рис. 2) в пакете “Bchron” 4.2.6 (Parnell, 2015) в статистической среде R v. 3.6.2 (R Core Team, 2019) с использованием “наземной” калибровочной кривой “IntCal13” (Reimer et al., 2013). Детали и математический аппарат, используемый в пакете, можно найти в работе Хаслетта и Парнелла (Haslett, Parnell, 2008). Ниже указаны калиброванные даты.
Рис. 1.
Карта района исследований. Алеутские острова. На врезке изображен остров Шемья, знаком отмечен торфяник Алкан Крик.

Таблица 1.
Полевое описание торфяника Алкан Крик (о. Шемья)
| Глубина, см | Описание торфа |
|---|---|
| 0–10 | Современные растения |
| 10–20 | Дернина черная, плотная, переплетена корнями |
| 20–80 | Торф черный, очень плотный, плохо разложившийся |
| 80–130 | Горизонт торфянистый, гомогенный, желтоватый, с включениями глины; растительные остатки плохо разложившиеся |
| 130–205 | Горизонт торфянистый, однородный, бурый, насыщен плохо разложившимися растительными остатками, с включениями глины. Слой 130-140 см более серый, с большим количеством минеральной фракции. На глубине 165 см есть тонкая прослойка песка |
| 205–250 | Горизонт аналогичен предыдущему, с желтоватым оттенком, встречаются небольшие черные вертикальные пятна |
| 250–260 | Горизонт глинистый, очень плотный, серый, пластинчатый, содержит много растительных остатков: в горизонте встречаются линзы песка и голубой глины |
| 260–264 | Горизонт глиняный, буроватый, с вкраплениями голубой глины |
| 264–270 | Горизонт глинистый, желтый, неоднородный, железистый, с включениями щебенки |
Таблица 2.
Радиоуглеродные датировки торфяника Алкан Крик (остров Шемья)
| Лабораторный номер | Глубина, см | Радиоуглеродный возраст, л. н. | Калиброванный возраст, л. н. | |
|---|---|---|---|---|
| медиана* | медиана ± 2σ | |||
| ИЭМЭЖ-1199 | 20–30 | 562 ± 83 | 830 | 570–1900 |
| ИЭМЭЖ -1201 | 80–90 | 3221 ± 75 | 3620 | 3340–4260 |
| ИЭМЭЖ -1200 | 130–140 | 4396 ± 198 | 5350 | 4700–6350 |
| ИЭМЭЖ -1202 & 1204 | 200–210 | 7562 ± 104 | 8425 | 8190–8810 |
| ИЭМЭЖ -1203 | 250–260 | 7924 ± 320 | 9365 | 8735–10 755 |
Рис. 2.
Модель скорости роста торфяника Алкан Крик. Темно-серый цвет – калиброванные даты, серый цвет – 95% доверительный интервал хронологии.

Для спорово-пыльцевого анализа было отобрано 28 образцов объемом 1 см3 из слоев толщиной 10 см. Химическая обработка проб включала в себя кипячение в 10%-ной HCl, 10%-ном KOH, промывку через сито, ацетолиз по стандартной методике (Faegri, Iversen, 1975) с некоторыми модификациями (Chambers et al., 2011). Для определения концентрации пыльцы и спор в процессе химической обработки добавляли специальные маркеры (Stockmarr, 1973). В каждый образец было добавлено по две таблетки спор Lycopodium clavatum из партии № 483216. Определение пыльцы и спор проводили с помощью атласов и ключей (Reille, 1998, 1999), электронных баз данных пыльцевых типов (https://globalpollenproject.org). Несмотря на наличие соответствующих определительных ключей (Warner, Chinnappa, 1986; Sarwar, Takahashi, 2014), пыльца Ericaceae была определена до уровня семейства, поскольку родовые особенности типов не всегда были различимы. Пыльца была посчитана под световым микроскопом Axioskop ZEISS с увеличением ×400, как минимум до 500 пыльцевых зерен в тех образцах, где это было возможно. Процентное соотношение пыльцевых зерен было рассчитано от суммы всей пыльцы в образце, а процентное соотношение спор от суммы пыльцевых зерен и спор в образце. Для построения спорово-пыльцевых диаграмм была использована программа Tilia software v.2.0.41 (Grimm, 2015). Пыльцевые зоны были выделены с помощью стратиграфически ограниченного кластерного анализа (CONISS) (Grimm, 2015).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Торфяник Алкан Крик начал формироваться ~9500 лет назад (рис. 2, табл. 2). Средняя скорость накопления – 0.27 мм/год. Отложение накапливалось относительно равномерно на протяжении всего времени. Только в начальный период формирования отложения 9200–8300 лет назад (250–200 см) торф накапливался несколько быстрее – средняя скорость накопления в это время составила 0.56 мм/год.
В результате спорово-пыльцевого анализа в отложении было выявлено 34 типа пыльцы и спор: 6 древесных палинотипов (“деревья и кустарники”), 21 травянистый палинотип (“травы”), 7 типов спор (рис. 3, рис. 4). Основные доминирующие таксоны – Ericaceae (вересковые), Cyperaceae (осоки), Poaceae (злаки). Участие Ericaceae варьировалось в пределах от единичных зерен до 88.4%, Cyperaceae – от 0.5 до 72.3%, Poaceae от 0.9 до 62.5% (рис. 3). Общая концентрация пыльцевых зерен в образце варьировала от 30 424 до 795 972 п. з. (пыльцевых зерен)/см3 (рис. 4).
Рис. 3.
Сокращенная процентная спорово-пыльцевая диаграмма торфяника Алкан Крик. AlCr-1a, AlCr-1b, AlCr-2 – пыльцевые зоны, выделенные на основании CONISS. Знаками + и ++ отмечены единично встретившиеся пыльцевые зерна.

Рис. 4.
Сокращенная диаграмма изменения концентрации пыльцевых зерен и спор торфяника Алкан Крик (п.з./см3). AlCr-1a, AlCr-1b, AlCr-2 – пыльцевые зоны, выделенные на основании CONISS.
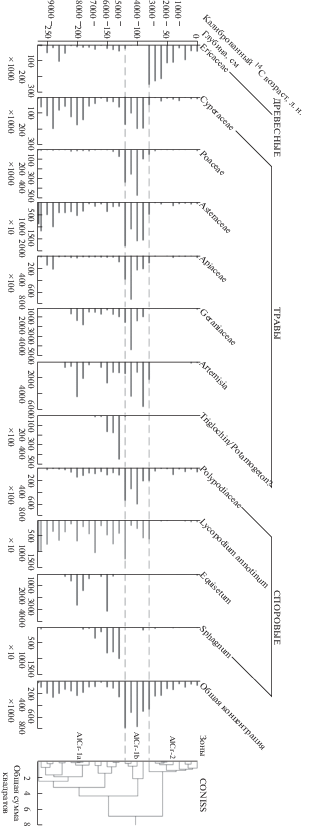
В самом основании торфяника, в двух первых образцах (~9500 лет назад) в спектрах доминировала пыльца Asteraceae (сложноцветные, 26.1%), Cyperaceae (25.2%), Poaceae (21.4%), а также споры Lycopodium annotinum (20.8%). Значение общей концентрации пыльцы – 35 642 п. з./см3.
В выделенной кластерным анализом зоне от 9500 до 4700 лет назад (AlCr-1a, 270–120 см) в процентном соотношении доминировала пыльца Cyperaceae – в среднем, 45.0% (максимальное значение концентрации – 200 972 п. з./см3). Второй по обилию пыльцевой тип – Ericaceae (рис. 3, 4), содержание которого составляет, в среднем, 17.6%. При этом разброс концентрации пыльцы Ericaceae в данной части составлял от 1355 до 106 900 п. з./см3. Среднее содержание пыльцы Poaceae составляет 12.4%, концентрация варьирует от 3708 п. з./см3 (6700 лет назад) до 346 882 п. з./см3 (4700 лет назад). Концентрации пыльцы Betula и Alnus имеют наиболее высокие значения концентрации в пределах этой зоны – до 4478 п. з./см3 8400 лет назад и 5033 п. з./см3 9200 лет назад соответственно. Около 8700 лет назад в спектрах появляются единичные споры Equisetum, концентрация которых достигает 3192 п. з./см3, 6500–6000 лет назад, а затем 5600 лет назад падает до нуля. Споры Sphagnum появляются в спектре ~8300 лет назад, увеличивают свое участие до 6.0% 6500–6000 лет назад, а затем практически исчезают из спектра ~5100 лет назад. В период 7800–5100 лет назад отмечено появление неопределенного палинотипа (безапертурные пыльцевые зерна с сетчатой скульптурой поверхности), содержание которого достигает 16.9% (5100 лет назад). Среди растений Алеутских островов (Hulten, 1968) такая пыльца может принадлежать растениям из родов Triglochin или Potamogeton.
Около 4700 лет назад (зона AlCr-1b, 120–80 см, 4700–3400 лет назад) в составе спорово-пыльцевых спектров увеличивается процент участия пыльцы Poaceae, а пыльцевые зерна Ericaceae единичны (рис. 3, 4). Концентрация пыльцы Apiaceae, Poaceae и Fritilaria возрастает до 74 332, 485 911 и 3097 п. з./см3 соответственно. Концентрация пыльцы Ranunculus-type резко увеличивается до 145 566 п. з./см3, к концу зоны (3400 лет назад) снижается до 7585 п. з./см3, затем исчезает. Изменения концентрации пыльцы 5100–3400 лет назад – 481 641–795 972 п. з./см3, значения максимальны для всего отложения. Споры Lycopodium annotinum и пыльца Asteraceae на верхней границе зоны AlCr-1b (3400 лет назад, 80 см) практически исчезают из состава спектра.
Пыльца Ericaceae в процентном соотношении начинает преобладать 3400 лет назад (зона AlCr-2, 3400–0 лет назад, 80–0 см) совместно с пыльцой Cyperaceae. При этом концентрация пыльцы осоковых сильно снизилась (рис. 4) по сравнению с предыдущей зоной (до 2549–26 346 п. з./см3). Концентрация пыльцы Ericaceae максимальна ~3400 лет назад – 252 577 п. з./см3, затем постепенно снижается к поверхности до 42 475 п. з./см3. На протяжении зоны AlCr-2 резко снижается общая концентрациия пыльцы – от 53 541 до 261 505 п. з./см3. Содержание пыльцы Poaceae, в среднем, составляет 5.1%, за исключением самого верхнего образца, где она достигает 16.4%.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Торфяник Алкан Крик начал формироваться в глубине острова Шемья ~9500 лет назад (рис. 1, 2). В это время растительные сообщества, преимущественно, были представлены разнотравно-осоковой тундрой с участием вересковых кустарничков (рис. 3). Злаково-осоковые и разнотравно-осоковые тундры на Алеутских островах распространены в более защищенных от действия ветра условиях, часто слабо дренированных (Heusser, 1973; Jordan, Krumhard, 2003). В состав первичных растительных сообществ других островов архипелага входили зонтичные и плауны, как и на о. Шемья (Heusser, 1973, 1978, 1990; Kuzmicheva et al., 2019). При этом различие доминантов первичных сообществ могли быть значительными, а состав зависел скорее от локальных условий формирования, чем климатической обстановки. На о. Умнак первичные сообщества были представлены сильно переувлажненными осоковыми тундрами (Heusser, 1973). На о. Адак также преобладали осоки, но локальные условия, вероятно, были более сухими (Heusser, 1978). Спорово-пыльцевые спектры острова Атка сходны со спектрами торфяника Алкан Крик, но в растительности о. Атка доминировали злаковые, что также отражает более сухие локальные условия (Heusser, 1990). На близком к острову Шемья о. Атту (Heusser, 1990) первичные сообщества были представлены разнотравьем с высоким участие зонтичных (Apiaceae) и сложноцветных (Asteraceae), которое формировалось в более защищенных от ветра условиях. На Алеутских островах, практически лишенных в голоцене наземных хищников, преобладание в спорово-пыльцевых спектрах зонтичных и сложноцветных может означать наличие рядом с торфяником крупной колонии морских птиц, поскольку отдельные виды этих семейств предпочитают богатые азотом субстраты (Kuzmicheva et al., 2019). Спектры первых сукцессионых стадий о. Карлайл заметно отличаются доминированием вересковых в связи с формированием растительности в условиях сильного воздействия ветра непосредственно в прибрежной зоне (Kuzmicheva et al., 2019). Аналогичные им современные сообщества можно также встретить на морском берегу или высоко на склонах (Heusser, 1990; Jordan, Krumhardt, 2003). Можно предположить, что локальные условия местообитаний, степень защищенности от ветра, дренированности и аэрированности субстрата, а также наличие птичьих колоний в совокупности определяли внешний облик и состав доминантов первичных растительных сообществ.
Около 7000 лет назад разнотравно-осоковая тундра с участием вересковых кустарничков на о. Шемья заметно заболачивается (рис. 3). Индикаторами интенсивного увлажнения служат споры Equisetum (Galka et al., 2018) и сфагновых мхов (Jordan, Krumhardt, 2003). Безапертурная пыльца, которая может принадлежать растениям родов Potamogeton или Triglochin (рис. 3, 4), также является индикатором местообитаний с избыточным увлажнением (Hulten, 1968). Период 7000–5000 лет назад для региона Берингова моря считается температурным максимумом голоцена (Kaufman et al., 2016), после чего температура постепенно понижается. В течение голоцена, в болотах арктической Аляски Sphagnum имел высокое участие в составе растительности в относительно более теплые климатические фазы (Galka et al., 2018). Можно предположить, что экспансия сфагновых мхов, способных удерживать большое количество влаги, привела к формированию на месте влажной тундры осокового болота. Эта тенденция хорошо прослеживается при анализе спорово-пыльцевого спектра о. Атту (Heusser, 1990), где 7000 лет назад также происходит заболачивание тундры благодаря участию Sphagnum (Heusser, 1990). Таким образом, к локальному увеличению увлажнения сообществ на Алеутских островах могли приводить не только периоды увеличения влажности в регионе, но и продолжительные периоды потепления за счет увеличения участия сфагновых мхов.
Около 4700 лет назад в ответ на наступившее за температурным максимумом похолодание Sphagnum исчезает из спектров, и осоковое болото зарастает разнотравными осоково-злаковыми лугами (рис. 3). На о. Атту (Heusser, 1990) также произошла смена переувлажненного осокового болота со Sphagnum на разнотравно-злаковые луга, но почти на тысячу лет раньше – 5500 лет назад. Несинхронность смен растительности на соседнем с о. Шемья острове можно связать с гористым рельефом о. Атту и наличием в отложении пепловых прослоек. На островах Адак, Атка, Карлайл и Умнак подобных смен растительности в ответ на последовавшее за максимумом температур похолодание не наблюдается (Heusser, 1973, 1978, 1990; Kuzmicheva et al., 2019). Изменения растительных сообществ на этих островах могли быть в большей степени обусловлены влиянием выпадавших вулканических пеплов.
Период 4700–3400 лет назад характеризуется максимальными значениями концентрации пыльцы в торфянике Алкан Крик (рис. 4). При этом увеличение концентрации пыльцы не могло быть связано с особенностями формирования отложения, поскольку, по данным модели роста (рис. 2), на протяжении почти всей истории, торф накапливался равномерно. Этот период характеризуется постепенным снижением температур, последовавшим за голоценовым максимумом (Kaufman et al., 2016). При этом для центральной и восточной частей Алеутских островов реконструированы относительно более сухие периоды 4000–2700 и 5000–3500 лет назад соответственно (Jones et al., 2009; Krawiec, Kaufman, 2014). Можно было бы предположить, что в фазу похолодания более сухие условия на океанических островах с большой облачностью были благоприятными для увеличения продуктивности многих травянистых таксонов, но ни на одном из изученных островов не отмечено повышения концентраций пыльцы в данный период времени. Вероятно, значения концентрации пыльцы оказались чувствительны к вулканическим извержениям, имевшим разное влияние на растительность вдоль всей Алеутской гряды в голоцене (Heusser, 1990). Выпадение в результате извержений пепловых прослоек разной толщины и химического состава могло приводить как к полному уничтожению растительности, так и к заметным сукцессионным сменам сообществ (Нешатаева, 2009), не позволяющим проследить влияние климата.
Около 3400 лет назад разнотравные осоково-злаковые луга достаточно резко сменяются сообществами с доминированием вересковых кустарничков (рис. 3, 4). Значения общей концентрации пыльцы и многих пыльцевых типов, за исключением Ericaceae, снижаются практически до нуля. Также постепенно снижается таксономическое разнообразие. Исходя из современного распространения реконструированных сообществ, можно предположить, что к смене сообществ могли привести похолодание, заметное усиление ветровой активности, общее уменьшение влажности в регионе или локальное изменение гидрологического режима. Возможно также взаимодействие всех выше перечисленных причин. Заметное похолодание началось в регионе Берингова моря, по разным данным, от 4000 до 3000 лет назад (Black, 1976, 1981; Thorson, Hamilton, 1986; Jordan, Krumhardt, 2003). Усиление штормовой активности (Jordan, Krumhardt, 2003) также могло способствовать доминированию вересковых кустарничков, более приспособленных к условиям сильного воздействия ветра (Heusser, 1978). На о. Атту ~3000 лет назад также наблюдается заметный пик участия верескового кустарничка Empetrum nigrum (Heusser, 1990), который образует сообщества на продуваемых хорошо дренированных склонах и в других регионах (Бреслина, 1987; Bunting, 1994). На других островах Алеутской гряды, растительность которых была подвержена более заметному влиянию вулканических извержений (Heusser, 1973, 1978, 1990), не происходило смены травяных сообществ на кустарничковые в связи с заметным похолоданием 4000–3000 лет назад.
Смену травяных сообществ кустарничковыми можно объяснить не только климатическими факторами. На океанических островах в силу избыточного увлажнения идет интенсивный процесс выщелачивания и вымывания почв и торфа (Birks, Peglar, 1979). Подобные процессы наблюдаются на Шетландских (Bunting, 1994) и Фарерских островах (Lawson et al., 2005). В условиях недостатка минерального питания вересковые кустарнички получают конкурентное преимущество по сравнению с травянистыми видами растений, благодаря эрикоидной микоризе (Макаров, 2019). Можно предположить, что на о. Шемья травяные сообщества сменяются кустарничковыми с доминированием вересковых в результате постепенного выщелачивания торфяника Алкан Крик в условиях океанического климата. На о. Адак (Heusser, 1978; Noguchi et al., 2018) вересковые также могли стать доминантами в силу схожих процессов выщелачивания, а не в ответ на более сухие условия (Vaillencourt, 2013). Похожая смена травяных сообществ на вересковые происходила на острове Атту (Heusser, 1990), который, как и о. Шемья, относится к западной части гряды. На острове Атту Ericaceae начинают играть существенную роль в составе спорово-пыльцевых спектров 6200 лет назад, что также может быть связано с гористым рельефом острова и более сильным воздействием ветра, а ~3000 лет назад их участие в составе спектров заметно возрастает (Heusser, 1990). Климатические факторы и интенсивное выщелачивание в условиях избыточного увлажнения могли влиять на растительность Алеутских островов в голоцене совместно.
Таким образом, на основании результатов спорово-пыльцевого анализа торфяника Алкан Крик была реконструирована история растительности острова Шемья (Ближние Алеутские острова). Торфяник формировался со средней скоростью накопления 0.27 мм/год. Период относительно более высокой скорости накопления отмечен 9200–8300 лет назад (0.56 мм/год). Около 9500 лет назад в защищенных от ветра и слабо дренированных условиях на острове преобладала разнотравно-осоковая тундра с участием вересковых кустарничков. В результате заметного потепления 7000 лет назад в растительных сообществах увеличилось участие Sphagnum. Сфагновые мхи, благодаря своей способности удерживать влагу, сформировали на месте влажной тундры условия для возникновения осокового болота с участием Equisetum. В ответ на последовавшее похолодание, ~4700 лет назад Sphagnum исчез из спорово-пыльцевых спектров, а на месте болота в более сухих условиях образовались разнотравные осоково-злаковые луга. Период 4700–3400 лет назад, отличавшийся сухим климатом и более низкой штормовой активностью, характеризуется максимальными значениями концентрации пыльцы в торфянике Алкан Крик. Около 3400 лет назад травяные сообщества на острове Шемья сменились сообществами с доминированием вересковых кустарничков, что могло быть связано с фазой похолодания в регионе Берингова моря, начавшейся ~3000 лет назад.
Авторы выражают благодарность Дикси Вест за организацию и совместное участие в экспедиции.
Работа выполнена при финансовой поддержке РФФИ (гранты 19-34-90071, 18-04-00782 А).
Список литературы
Бреслина И.П. Растения и водоплавающие птицы морских островов Кольской Субарктики. Л.: Изд-во" Наука", 1987. 200 с.
Кирьянов В.Ю., Егорова И.А., Литасова С.Н. Вулканические пеплы на острове Беринга (Командорские острова) от голоценовых извержений Камчатки // Вулк. и сейсм. 1986. № 6. С. 18–28.
Киселева Н.К., Савинецкий А.Б., Хасанов Б.Ф. Развитие природных процессов на о Шемья в голоцене // Изв. РАН, сер. Геогр. 2002. Т. 1. С. 97.
Макаров М.И. Роль микоризы в трансформации соединений азота в почве и в азотном питании растений (обзор) // Почв. 2019. №. 2. С. 220–233.
Нешатаева В.Ю. Растительность полуострова Камчатка. М.: Товарищество научных изданий КМК, 2009. 537 с.
Anderson S.T., Bank T.P. Pollen and radiocarbon studies of Aleutian soil profiles //Sci. 1952. V. 116. № 3004. P. 84–86.
Birks H.J.B., Peglar S.M. Interglacial pollen spectra from Sel Ayre, Shetland // New Phytol. 1979. V. 83. № 2. P. 559–575.
Black L.T. Volcanism as a factor in human ecology: the Aleutian case // Ethnohistory. 1981. P. 313–340.
Black R.F. Geology of Umnak Island, eastern Aleutian Islands as related to the Aleuts // Arct. Antarct. Alp. Res. 1976. V. 8. № 1. P. 7–35.
Bunting M.J. Vegetation history of Orkney, Scotland; pollen records from two small basins in west Mainland // New Phytolt. 1994. V. 128. № 4. P. 771–792.
Byers F.M. Investigations on Alaska volcanoes: Geology of Umnak and Bogoslov Islands, Aleutian Islands, Alaska // Geol. Soc. Am. Bull. 1959. 267–369.
Chambers F.M., van Geel B., van der Linden M. Considerations for the preparation of peat samples for palynology, and for countin of pollen and non-pollen palynomorphs // Mires and Peat. 2011. V. 7. P. 1–14.
Faegri K., Iversen J. Textbook of pollen analysis. // Munksgaard, Copenhagen. 1975. 295 p.
Gałka M., Swindles G.T., Szal M., Fulweber R., Feurdean A. Response of plant communities to climate change during the late Holocene: Palaeoecological insights from peatlands in the Alaskan Arctic // Ecol. Indic. 2018. V. 85. P. 525–536.
Grimm E.C. Tilia for windows: pollen spreadsheet and graphics program // Version. 2015. 2.41
Haslett J., Parnell A.C. A simple monotone process with application to radiocarbon-dated depth chronologies // J. R. Stat. Soc. Ser. C Appl. Stat. 2008. V. 57. № 4. P. 399–418.
Heusser C.J. Postglacial vegetation on Adak Island, Aleutian Islands, Alaska // Bull. Torrey Bot. Club. 1978. P. 18–23.
Heusser C.J. Postglacial vegetation on Umnak Island, Aleutian Islands, Alaska // Rev. of Palaeobot. and Palyno. 1973. V. 15. № 4. P. 277–285.
Heusser C.J. Late Quaternary vegetation of the Aleutian Islands, southwestern Alaska // Can. J. Bot. 1990. V. 68. № 6. P. 1320–1326.
Hultén E. Flora of Alaska and neighboring territories: a manual of the vascular plants. – Stanford University Press, 1968. 1008 p.
Jones M.C., Peteet D.M., Kurdyla, D., Guilderson T. Climate and vegetation history from a 14.000-year peatland record, Kenai Peninsula, Alaska // Quaternary Res. 2009. P. 72. № 2. P. 207–217.
Jordan J.W., Krumhardt A. Postglacial climate and vegetation on the western Alaska Peninsula // Alaska J. Anthropol. 2003. V. 1. P. 16–33.
Kaufman D.S., Axford Y.L., Henderson A.C., McKay N.P., Oswald W.W., Saenger C., Anderson R.S., Bailey H.L., Clegg B., Gajewski K., Hu F.S., Jones M.C., Massa C., Routson C.C., Werner A., Wooller M.J., Yu Z. Holocene climate changes in eastern Beringia (NW North America)–A systematic review of multi-proxy evidence // Quat. Sci. Rev. 2016. V. 147. P. 312–339.
Krawiec A.C.L., Kaufman D.S. Holocene storminess inferred from sediments of two lakes on Adak Island, Alaska // Quaternary res. 2014. V. 82. № 1. P. 73–84.
Kuzmicheva E.A., Smyshlyaeva O.I., Vasyukov D.D., Khasanov B.F., Krylovich O.A., Okuno M., West D.L., Hatfield V.L., Savinetsky, A.B. A 7300-yr-old environmental history of seabird, human, and volcano impacts on Carlisle Island (the Islands of Four Mountains, eastern Aleutians, Alaska) // Quaternary Res. 2019. V. 91. № 3. P. 934–952.
Lawson I.T., Church M.J., McGovern T.H., Arge S.V., Woollet J., Edwards K.J., Gathorne-Hardy F.J., Dugmore A.J., Cook G., Mairs K.-A., Thomson A.M., Sveinbjarnardotti G. Historical ecology on Sandoy, Faroe Islands: palaeoenvironmental and archaeological perspectives // Hum. Ecol. 2005. V. 33. № 5. P. 651–684.
Noguchi M., Fujiki T., Okuno M., Gualtieri L., Hatfield V.L., Sarata B., Torii M., Wada K., Nakamura T., West D. Vegetation Changes around Haven Lake, Adak Island, Central Aleutians, Alaska, Determined from Pollen Analysis // Radiocarbon. 2018. V. 60. № 5. P. 1483–1492.
Parnell A. Bchron: Radiocarbon dating, age-depth modelling, relative sea level rate estimation, and non-parametric phase modelling. R package version 4.1. 1; 2015.
R Core Team (2019). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/
Reille M. Pollen et Spores ďEurope et ďAfrique du Nord: Supplement 2. – Marseille, 1998. 521 p.
Reille M. Pollen et spores d’Europe et d’Afrique du Nord: index. – Marseille, 1999. 535 p.
Reimer P.J., Bard E., Bayliss A., Beck J.W., Blackwell P.G., Ramsey C.B., Buck C.E., Cheng H., Edwards R.L., Friedrich M., Grootes P.M., Guilderson T.P., Haflidason H., Hajdas I., Hatte C., Heaton T.J., Hoffmann D.L., Hogg A.G., Hughen K.A., Kaiser K.F., Kromer B., Manning S.W., Niu M., Reimer R.W., Richards D.A., Scott E.M., Southon J.R., Staff R.A., Turney C.S.M., van der Plicht J. IntCal13 and Marine13 radiocarbon age calibration curves 0–50.000 years cal BP // Radiocarbon. 2013. V. 55. № 4. P. 1869–1887.
Rodionov S.N., Bond N.A., Overland J.E. The Aleutian Low, storm tracks, and winter climate variability in the Bering Sea // Deep Sea Research Part II: Topical Studies in Oceanography. 2007. V. 54. № 23–26. P. 2560–2577.
Sarwar A.K.M., Takahashi H. Pollen morphology of the tribe Phyllodoceae (Ericoideae, Ericaceae) and its taxonomic significance // Bangl. J. Plant Taxon. 2014. V. 21. № 2. P. 129–137.
Savinetsky A.B., Kiseleva N.K., Khasanov B.F., Lefevre C., Corbett D. Paleoenvironment-Holocene Deposits from Shemya Island // The People at the End of the World: The Western Aleutians Project and the Archaeology of Shemya Island. Aurora Monograph Series. 2010. V. 7.
Stockmarr J. Determination of spore concentration with an electronic particle counter: Danmarks Geologiske Undersøgelse, 1973.
Talbot S.S., Talbot S.L., Walker L.R. Post-eruption legacy effects and their implications for long-term recovery of the vegetation on Kasatochi Island, Alaska // Arct. Antarct. Alp. Res. 2010. V. 42. № 3. P. 285–296.
Thorson R.M., Hamilton T.D. Glacial Geology of the Aleutian Islands: Based on the contributions of Robert F. Black., 1986.
Vaillencourt D.A. Five-thousand years of hydroclimate variability on Adak Island, Alaska inferred from δD of n-alkanoic acids (Doctoral dissertation, Northern Arizona University), 2013.
Warner B.G., Chinnappa C.C. Taxonomic implications and evolutionary trends in pollen of Canadian Ericales // Can. J. Bot. 1986. V. 64. № 12. P. 3113–3126.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая