

Известия РАН. Серия биологическая, 2022, № 1, стр. 93-101
Сезонная изменчивость показателей системы крови домовой мыши (Mus musculus L.) в условиях гор Центрального Кавказа
М. М. Емкужева 1, *, Ф. А. Темботова 1, Э. Ж. Темботова 1
1 Институт экологии горных территорий им. А.К. Темботова РАН
360051 Нальчик, ул. И. Арманд, 37а, Россия
* E-mail: emkugeva_m@mail.ru
Поступила в редакцию 31.07.2018
После доработки 25.05.2021
Принята к публикации 25.05.2021
- EDN: RHCGMP
- DOI: 10.31857/S1026347021060056
Аннотация
Исследована изменчивость функционирования кроветворной системы (эритропоэз костного мозга) и показателей крови домовой мыши (Mus musculus L.) в горах Центрального Кавказа (1800 м над ур. м.) в зависимости от сезона года. Выявлена выраженная сезонная динамика в деятельности костного мозга и периферической крови с выделением двух периодов: холодного (зима) и теплого (весна, лето, осень). Для вида наиболее энергозатратным года является зимний сезон, что проявляется в максимальной активности костного мозга, приводящей к повышению дыхательной функции крови. Наблюдаемое в этот период максимальное увеличение кислородной емкости крови, обусловливается выходом в периферическую кровь большого количества ретикулоцитов – молодых эритроцитов. При этом минимальные в годовом цикле размеры эритроцитов способствуют увеличению общей поглотительной поверхности для кислорода.
Сезонные изменения условий существования, которые обуславливают как климатические изменения, так и преобразования в кормовой базе животных – один из важнейших факторов эволюционной экологии грызунов. При этом оценка признаков системы крови, с одной стороны, может характеризовать реакцию организма на изменяющиеся условия среды обитания, с другой – характеризовать эти условия (Лохмиллер, Мошкин, 1999; Тарахтий, Давыдова, 2007). Особый интерес подобные исследования представляют в условиях горных территорий, где на сезонную цикличность накладывается пониженное парциальное давление кислорода. При адаптации горные популяции широко распространенных видов используют все механизмы компенсации кислородной недостаточности (Большаков, 1972). Так, домовая мышь (Mus musculus L.), относящаяся к группе настоящих синантропов (Кучерук, 1988), освоила широкий спектр природных и антропогенных ландшафтов Центрального Кавказа, в том числе и в высотном направлении (выше 2000 м над ур. м), причем в естественных биотопах выше 1500 м над ур. м. не встречается (Темботов, Шхашемишев, 1984 и наши данные). К числу лимитирующих факторов, определяющих возможность существования зверьков в жилищах человека, по данным литературы, относятся низкие температуры на севере и высота над уровнем моря в горах (Домовая мышь, 1994). Приспособление вида к условиям среднегорий Центрального Кавказа обеспечивается высокой эритропоэтической активностью костного мозга, большим числом ретикулоцитов, поступающих в периферическую кровь, что сопровождается более высоким содержанием гемоглобина в крови, за счет более крупных по объему эритроцитов, без увеличения их количества, по сравнению с предгорьями. При этом изменения индекса сердца не выявлено. Данный показатель имеет высокие значения как в предгорьях, так и среднегорьях (Темботова и др., 2007а, 2007б; Емкужева, 2013). Г. Маклин и А. Ли (Maclean, Lee, 1973) показали зависимость состояния крови домовой мыши Австралии от сезона года, температуры среды и двигательной активности: концентрация гемоглобина, гематокрит и объем циркулирующей крови зимой существенно выше, чем летом. В предгорьях Центрального Кавказа у вида также выявлена сезонность в изменчивости показателей красной крови (Темботов и др., 2009). В горах Кавказа сезонная изменчивость системы крови домовой мыши не изучена. В этой связи цель работы – оценить количественно-качественные морфологические показатели кроветворной ткани и периферической крови домовой мыши (M. musculus L.) и их сезонную изменчивость в условиях среднегорий Центрального Кавказа.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования, согласно типизации А.К. Темботова (Соколов, Темботов, 1989), проводили в условиях субальпийского пояса эльбрусского варианта Центрального Кавказа: пос. Эльбрус, высота местности – 1800 м над ур. м., GPS координаты – 43°15′ с.ш., 42°38′ в.д. В работе использовали 169 особей домовой мыши (Mus musculus L.) с охватом всех сезонов года: зима ♂♂ – 17, ♀♀ – 14; весна ♂♂ – 14, ♀♀ – 12; лето ♂♂ – 31, ♀♀ – 28; осень ♂♂ – 23, ♀♀ – 30. Животных отлавливали стандартным методом ловушко-линий (Карасева, Телицина, 1996) в домах и хоз. постройках местных жителей. Изучены взрослые особи, возраст которых определяли на основании комплекса признаков стертости зубов, массы тела, состояния генеративных органов. Беременные самки включены в анализ, поскольку при сравнении изученных параметров значимых различий между беременными и небеременными самками не выявлено. Домовая мышь в условиях гор Центрального Кавказа размножается круглогодично, пик приходится на весенне-летний период и снижение – в осенне-зимний период. Доля беременных от общего числа самок составляет: зимой – 11.1%, весной – 58.3%, летом – 33.3%, осенью – 2.7%.
Исследование показателей системы крови проводили в течение суток после отлова. Костный мозг получали из бедренной кости (Тодоров, 1968). Препараты костного мозга приготовляли на предметных стеклах (Воробьев, 1985), и окрашивали комбинированным методом (по Май-Грюнвальд и Романовскому-Гимза). Морфологический анализ клеток костного мозга (соотношение клеток лейкоцитарного и эритробластного ряда, эритробластограмма) проводили с помощью микроскопа Микмед-5, окуляр – 10, объектив – 100 с масляной иммерсией (Берчану, 1985; Воробьев, 1985; Меньшиков, 2012). Ретикулоциты крови (RET) и их парциальный состав изучали по Е.Н. Мосягиной (1962) на микропрепаратах, окрашенных 1%-ным раствором бриллиант-крезилблау в физиологическом растворе (pH 7.3). Расчет проводили на 10 тыс. эритроцитов. По степени зрелости различали 5 групп ретикулоцитов (RET 0, RET I, RET II, RET III, RET IV), по Л. Гейльмейеру (Тодоров, 1968). Концентрацию гемоглобина (HGB, г/л) определяли унифицированным гемоглобин цианидным методом на гемоглобинометре фотометрическом Мини ГЕМ-540, гематокритное число – соотношение плазмы и форменных элементов (HCT, об. %) – микрометодом на гематокритной центрифуге СМ-70, подсчет количества эритроцитов (RBC, млн в 1 мкл) проводили в камере Горяева. Диаметр эритроцитов (микрометры) измерялспрямым микрометрическим методом с помощью винтового окуляр-микрометра МАВ 1–16× на сухих окрашенных препаратах крови, с использованием микроскопа Микмед-5 с масляной иммерсией. Измеряли диаметр 100 различных эритроцитов, результаты распределяли по группам (по величине диаметра эритроцитов).
Рассчитывали среднее содержание гемоглобина в эритроците (MCH) в пикограммах (пг), среднюю концентрацию гемоглобина в эритроците (MCHC) в г/л, средний объем эритроцита (MCV) в кубических микрометрах (мкм3), (Кост, 1975; Берчану, 1985; Воробьев, 1985; Иванова, 1995; Риган и др., 2000). Среднюю площадь эритроцита рассчитывали по Берчану (1985) и выражали в мкм2.
Статистическую обработку данных проводили с использованием пакета программ “Statistica-10 for Windows”. Влияние факторов проанализировано многофакторным дисперсионным анализом. При оценке различий показателей использовали Tukey-тест. Для всех статистических тестов принят 5% уровень значимости.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Двухфакторный дисперсионный анализ (Factorial ANOVA) показал, что система крови домовой мыши в условиях среднегорья Центрального Кавказа претерпевает значительные сезонные изменения. Влияние пола животных незначимо (табл. 1).
Таблица 1.
Результаты двухфакторного многомерного анализа
| Эффекты | F | P | df |
|---|---|---|---|
| Пол | 1.8 | 0.087 | 10 |
| Сезон | 12.0 | 0.000 | 30 |
| Взаимодействие факторов | 2.1 | 0.000 | 30 |
Система крови домовой мыши в зимний период (январь–февраль). Активная часть паренхимы костного мозга у всех изученных домовых мышей в зимний период была густо заполнена кроветворными клетками. Эритроидные клетки на препаратах равномерно располагались между другими кроветворными элементами лейкоцитарного ряда. На каждое поле зрения их приходилось до 20–30, среди которых 2–7 клеток эритроидного ряда находились в стадии митоза. Жировые клетки встречались не в каждом поле зрения. В парциальном соотношении клеточных групп костного мозга эритроидные клетки составили 10% у самцов и 20.26% у самок, оставшиеся ~80% приходятся на лейкоцитарный ряд (табл. 2).
В эритрограмме выявлен сдвиг в левую сторону за счет молодых клеток (эритробластов и базофильных нормобластов). Содержание ретикулоцитов (молодых эритроцитов) в периферической крови высокое и составляет 40.2‰ у самцов и 39.3‰ у самок. В парциальном составе преимущественное содержание ретикулоцитов второй и третьей стадий. Число эритроцитов в 1 мкл крови у самцов – 8.975 млн и у самок – 8.870 млн, при этом они небольших размеров (5.45 и 5.51 мкм соответственно). Содержание гемоглобина, в среднем, высокое, у особей обоих полов (табл. 3). Также высока респираторная функция крови. Средняя площадь эритроцитов зимой составила 91.42 мкм2 у самцов, у самок – 93.00 мкм2.
Таблица 2.
Сезонная динамика параметров костного мозга домовой мыши (Mus musculus) в условиях среднегорий Центрального Кавказа
| Параметры | Пол | Среднее значение | p < 0.05 | ||||
|---|---|---|---|---|---|---|---|
| зима (1) | весна (2) |
лето (3) |
осень (4) |
||||
| Эритробластный ряд в % ко всем ядерным клеткам костного мозга | ♂ | 20.10 | 18.69 | 19.54 | 19.63 | 1-2, 2-3 | |
| ♀ | 20.26 | 19.11 | 21.00 | 18.70 | 1-2, 2-3, 3-4 | ||
| Эритробластограмма | Эритробласты, % | ♂ | 3.64 | 2.87 | 2.00 | 3.17 | 1-2, 2-3, 3-4, 1-3, |
| ♀ | 3.33 | 2.66 | 2.17 | 3.20 | 1-2, 2-3, 3-4, 1-3 | ||
| Базофильные Нормобласты, % |
♂ | 20.23 | 17.19 | 15.76 | 15.60 | 1-2, 2-3, 1-3 | |
| ♀ | 19.31 | 16.38 | 15.45 | 17.52 | 1-2, 3-4, 1-3 | ||
| Полихроматофильные нормобласты, % | ♂ | 30.91 | 31.35 | 30.29 | 29.03 | 2-3, 3-4 | |
| ♀ | 32.06 | 31.97 | 30.71 | 29.52 | 1-3, 1-4 | ||
| Оксифильные нормобласты, % | ♂ | 52.35 | 52.30 | 53.62 | 55.41 | 3-4 | |
| ♀ | 52.80 | 51.21 | 52.68 | 56.96 | 3-4 | ||
| Соотношение лейкопоэз/эритропоэз | ♂ | 3.97 : 1 | 4.35 : 1 | 4.11 : 1 1 | 4.09 | ||
| ♀ | 3.94 : 1 | 4.23 : 1 | 3.76 : 1 | 4.34 : 1 | |||
Таблица 3.
Сезонная динамика параметров периферической крови домовой мыши (Mus musculus L.) в условиях среднегорий Центрального Кавказа
| Параметры | Пол | Среднее значение | p < 0.05 | ||||
|---|---|---|---|---|---|---|---|
| зима (1) | весна (2) | лето (3) | осень (4) |
||||
| Общее количество ретикулоцитов (RET, ‰) | ♂ | 39.98 | 31.82 | 39.53 | 37.84 | 1-2, 2-3 | |
| ♀ | 39.34 | 36.10 | 39.80 | 39.32 | 1-2, 2-3 | ||
| Парциальный состав | RET 0 (‰) | ♂ | – | – | – | – | |
| ♀ | – | – | – | – | |||
| RET I (‰) | ♂ | 1.29 | 0.56 | 0.47 | 1.13 | 1-2, 2-3, 3-4, 1-3 | |
| ♀ | 0.75 | 0.69 | 0.46 | 0.76 | 3-4 | ||
| RET II (‰) | ♂ | 17.12 | 4.22 | 7.76 | 5.19 | 1-2, 2-3, 1-3 | |
| ♀ | 16.49 | 4.42 | 7.47 | 5.45 | 1-2, 2-3, 1-3 | ||
| RET III (‰) | ♂ | 13.63 | 17.41 | 16.53 | 16.29 | 1-2, 2-3, 1-3 | |
| ♀ | 13.88 | 20.18 | 16.33 | 17.67 | 1-2, 1-3 | ||
| RET IV (‰) | ♂ | 7.56 | 9.42 | 8.96 | 10.06 | 1-2, 2-3,1-3 | |
| ♀ | 7.97 | 12.20 | 9.10 | 9.63 | 1-2, 3-4, 1-3 | ||
| HGB, г/л | ♂ | 170.94 | 161.43 | 159.37 | 160.39 | 1-2, 1-3 | |
| ♀ | 173.77 | 160.92 | 160.64 | 161.23 | 1-2, 2-3, 1-3 | ||
| RBC, млн в 1 мкл | ♂ | 8.975 | 8.510 | 8.129 | 8.102 | 1-3 | |
| ♀ | 8.870 | 7.858 | 8.048 | 8.115 | 1-2 | ||
| HCT, об/% | ♂ | 55.47 | 53.08 | 51.29 | 52.35 | 1-2, 1-3 | |
| ♀ | 54.54 | 52.82 | 51.13 | 52.52 | 1-3 | ||
| Диаметр эритроцитов, мкм | ♂ | 5.45 | 5.56 | 5.90 | 5.67 | 1-2, 2-3, 3-4, 1-3 | |
| ♀ | 5.51 | 5.64 | 5.83 | 5.68 | 1-2, 2-3, 3-4, 1-3 | ||
| MCH, пг | ♂ | 19.38 | 19.33 | 19.77 | 19.54 | ||
| ♀ | 20.17 | 20.52 | 19.80 | 19.91 | |||
| MCV, мкм3 | ♂ | 61.68 | 61.91 | 64.87 | 65.38 | ||
| ♀ | 63.24 | 64.93 | 64.18 | 64.83 | |||
| MCHC, г/л | ♂ | 31.01 | 31.27 | 31.05 | 30.65 | ||
| ♀ | 31.89 | 31.09 | 31.26 | 30.92 | |||
| F, усл. ед. | ♂ | 45.75 | 41.35 | 44.88 | 41.22 | 1-2, 3-4 | |
| ♀ | 47.73 | 40.50 | 43.30 | 42.82 | 1-2 | ||
| Средняя площадь эритроцитов, мкм2 | ♂ | 91.42 | 93.33 | 99.29 | 96.18 | 2-3, 3-4 | |
| ♀ | 93.00 | 95.95 | 97.27 | 95.32 | 1-3 | ||
По системе крови в зимний период по полу различия отмечены по эритробластам (p < 0.021), ретикулоцитам I стадии (p < 0.015) и диаметру эритроцитов (p < 0.015).
Морфологический анализ эритроцитов на препаратах крови показал, что они нормохромные, и процесс их поступления интенсивен, о чем свидетельствует наличие 6–14 полихроматофильных эритроцитов в каждом поле зрения. В эритроцитах обнаружены также включения телец Хауелла-Жолли (у 47.37% изученных животных). Все это свидетельствует о высокой активности эритропоэза в зимний период.
Система крови домовой мыши в весенний период (апрель–май). В весенний период (как и зимой) на препаратах костного мозга на фоне зрелых эритроцитов равномерно располагались кроветворные клетки. Эритроидные элементы встречались в каждом поле зрения в количестве до 10–20, 2–3 из которых на стадии митоза. Жировые клетки встречались реже, чем зимой. Полученные результаты свидетельствуют о снижении активности эритропоэза в весенний период, в сравнении с зимним, не только за счет уменьшения общего количества эритроидных клеток в костном мозге, но и за счет снижения доли более молодых форм – эритробластов и базофильных нормобластов – у обоих полов (табл. 2). При этом весной также происходят изменения в содержании ретикулоцитов в периферической крови при сравнении с зимними показателями. Так, общее количество ретикулоцитов весной уменьшается, снижаются клетки первой стадии (достоверно у самцов, на уровне тенденции – у самок), а ретикулоциты третьей (у самцов p < 0.001, у самок p < 0.0004) и четвертой стадий (p < 0.005 и p < 0.0001 соответственно) – увеличиваются (табл. 3).
В периферической крови содержание гемоглобина, число эритроцитов, гематокритный показатель снижаются весной по отношению к зиме. Что касается диаметра эритроцитов, то этот показатель весной достоверно увеличивается у обоих полов (табл. 3). Средняя площадь эритроцитов увеличивается, и более значительно у самцов. Среднее содержание гемоглобина в эритроцитах, средний объем эритроцитов и средняя концентрация гемоглобина в эритроцитах остаются на уровне зимних показателей (табл. 3). Респираторная функция весной снижается по отношению к зиме (p < 0.033, как у самок, так и у самцов). В дискриминантном анализе по показателям костного мозга в качестве различающих весенний и зимний сезоны признаков выступают базофильные нормобласты (F = 23.08 при p < 0.033 у самцов; F = 19.26 при p < 0.004 у самцов). По периферической крови – общее количество ретикулоцитов (F = 114.89 при p < 0.000), ретикулоциты III стадии (F = 57.49 при p < 0.000) и диаметр эритроцитов (F = 19.59 при p < 0.031) у самцов, и ретикулоциты II стадии (F = 17.85 при p < 0.001) у самок. Качество различения во всех моделях высокое (83–90%).
Морфологический анализ эритроцитов на препаратах крови в весенний период свидетельствует, что поступающие клетки (эритроциты) из костного мозга нормохромные. Полихроматофильные эритроциты, которые встречались в каждом поле зрения, свидетельствуют о нормальном процессе обновления эритроцитов. В эритроцитах (как и зимой) обнаружены тельца Хауелла-Жолли, но в процентном содержании их значительно больше (весной – 79.31%). Половые различия по параметрам системы крови в данном сезоне слабо выражены.
Система крови домовой мыши в летний период (июль–август). Морфологический анализ костного мозга летом показал равномерное распределение на препаратах кроветворных клеток. Эритроидные клетки встречались в количестве до 30 в каждом поле зрения. В стадии митоза встречались 2–5 клеток в поле зрения. В летний период в кроветворной системе домовой мыши происходят дальнейшие адаптивные преобразования в сравнении с весенним периодом. Летом активизируется эритропоэз костного мозга, повышается общее количество эритроидных клеток в костном мозге у обоих полов. Их количество составило 19.54% у самцов и 21.00% – у самок (табл. 2). В эритрограмме меняется соотношение эритроидных клеток. Снижается количество эритробластов (у обоих полов p < 0.000), базофильных нормобластов (достоверно у самцов, у самок – на уровне тенденции). Численные данные по более зрелым формам – полихроматофильным и оксифильным нормобластам – очень близки с весенними показателями. Летом в периферическую кровь по результатам дисперсионного анализа поступает большее, по сравнению с весной, количество ретикулоцитов (табл. 3). Увеличивается и доля молодых стадий (вторая стадия), а третья и четвертая стадии по своим численным данным близки с весенними показателями (табл. 3). Эритроциты летом значительно увеличиваются в размерах у обоих полов, по сравнению с весной (p < 0.0003), диаметр их составляет 5.90 мкм у самцов и 5.83 мкм у самок, площадь этих клеток летом достигает 99.29 мкм2 у самцов и 97.27 мкм2 у самок. Содержание гемоглобина в крови, число эритроцитов и гематокритный показатель остаются близкими весенним данным (табл. 3). Близки летние и весенние показатели по среднему содержанию гемоглобина, средней концентрации гемоглобина в эритроците и среднему объему эритроцитов. Респираторная функция крови у обоих полов имеет тенденцию к повышению по отношению к весенним данным. Морфологический анализ эритроцитов на препаратах крови показал, что они нормохромные, как и в предыдущие сезоны, процесс поступления молодых эритроцитов (полихроматофильных) увеличен. В эритроцитах, как и весной, обнаружены тельца Хауелла-Жолли у 57% изученных животных, что значительно меньше, чем в весенний период. Половые различия по изученным параметрам системы крови в данном сезоне не отмечены. В результате дискриминантного анализа различающими признаками между сезонами (весна–лето) по системе крови установлены: эритробласты (F = 26.06 при p < 0.0002 у самцов), общее количество ретикулоцитов (у самцов F = 14.39 при p < 0.0004 у самок F = 14.46 при p < 0.003), и ретикулоциты II стадии 14.46 при p < 0.0003, самок F = 13.26 при p < 0.005). Корректная дискриминация и качество моделей – 60–70%.
Система крови домовой мыши в осенний период. Осенью на препаратах костного мозга также наблюдалось равномерное распределение кроветворных клеток. Эритроидные клетки встречались в каждом поле зрения в количестве до 20–25, отмечены клетки в стадии митоза (1–3 в каждом поле зрения). По результатам дисперсионного анализа соотношение клеток эритроидного и лейкоцитарного ряда в костном мозге остается близким летним значениям у самцов, у самок отмечается понижение этих показателей (p = 0.000). Меняется качественный состав эритрограммы: повышается содержание эритробластов, базофильных (достоверно лишь у самцов) и оксифильных нормобластов нормобластов. Осенью поступление ретикулоцитов в кровь у домовой мыши имеет тенденцию к снижению у обоих полов по сравнению с летними данными. Изменения наблюдаются и в парциальном составе ретикулоцитов: увеличиваются клетки первой стадии. В крови осенью содержание гемоглобина, число эритроцитов, гематокритный показатель близки к летним значениям. Значительные сезонные изменения в осенний период происходят в крови с диаметром и площадью эритроцитов у особей обоих полов. Осенью эти показатели достоверно снижаются (p < 0.0001) по отношению к летним данным. Морфологический анализ эритроцитов на препаратах крови показал, что они нормохромные и процесс поступления молодых эритроцитов (полихроматофильных) не нарушен. Тельца Хауелла-Жолли в эритроцитах встречаются, но меньше, чем летом – 49.18%. Различия по полу в системе крови осенью отмечены по эритропоэтической активности. Дискриминирующими признаками по системе крови выступают эритробласты (у самцов 25.39 при p < 0.0001, у самок F = 25.51 при p < 0.0004) и диаметр эритроцитов (у самцов 9.43 при p < 0.0004, у самок F = = 12.34 при p < 0.010) у обоих полов. Корректная дискриминация высока во всех случаях сравнения (82–91%).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
У домовой мыши в условиях в среднегорий Центрального Кавказа система крови подвержена сезонной изменчивости. Анализ динамики кроветворения показал высокую активность эритропоэза во все сезоны года, при этом максимальные значения отмечаются зимой и летом, а в переходные сезоны – весной и осенью – более низкие. Сходный вектор изменчивости наблюдается и в динамике ретикулоцитов. Следует отметить, что сезонный ритм изменений проявляется не только в количественных характеристиках эритропоэза, но и в качественном составе клеток. Так, при высокой активности эритропоэза, в эритрограмме костного мозга в зимний период содержание эритробластов и базофильных нормобластов, а также количество клеток в стадии митоза достоверно выше, чем летом, что свидетельствует о более активном эритропоэзе костного мозга зимой. Это подтверждается данными по ретикулоцитам. При большом количестве ретикулоцитов в периферической крови, как и летом, увеличение в их парциальном составе I и II стадий (более молодых форм) также указывает на напряжение эритропоэза. Тем самым, в периферическую кровь домовой мыши поступает большое количество эритроцитов, но меньших размеров. Тренды сезонной изменчивости количества эритроцитов, гематокритной величины и концентрации гемоглобина в крови сходны: максимальные значения достигаются в зимний период и падение до минимальных – в летний. Респираторная функция крови, а соответственно, уровень потребления кислорода у домовой мыши также зимой высокие, кислородная емкость периферической крови в единице объема максимальна. Циркулирование в крови “мелких” эритроцитов в осенне-зимний период объяснимо с позиций общего физико-химического закона: чем больше площадь соприкосновения двух фаз, тем выше скорость диффузии. Большое количество “мелких” эритроцитов в 1 мкл крови не препятствует прохождению крови через капиллярное ложе и увеличивает общую поглотительную поверхность для кислорода. Чем меньше абсолютные размеры эритроцитов, тем больше скорость поглощения кислорода гемоглобином при прохождении крови через легкие, что необходимо для нормального поддержания организма в холодное время года и согласуется с данными литературы (Holland, Forster, 1966; Тарахтий и др., 2009).
Кластерный анализ по показателям системы крови домовой мыши на основании дистанции Махаланобиса (рис. 1) подтверждает выше сказанное и, как видно из рисунка, наиболее обособленной является зимняя выборка, что справедливо для особей обеих полов.
Рис. 1.
Дендрограмма сходства (Ward s method, 1-Pearson – r) между выборками M. musculus по показателям системы крови на основании дистанции Махаланобиса в разные сезоны года.
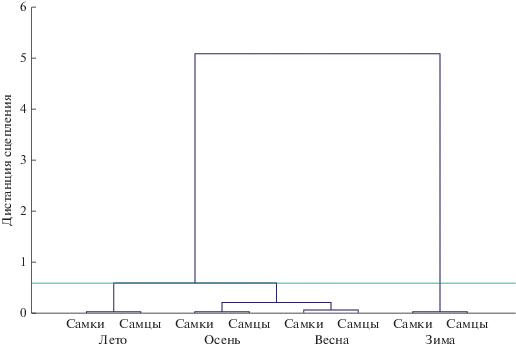
Учитывая, что средняя предпочитаемая температура у домовой мыши, согласно данным В.А. Стальмаковой (1978), значительно выше, чем у ряда других грызунов, обитающих в естественных условиях (рыжая и обыкновенная полевки, лесная и желтогорлая мыши), также как и чувствительность к холоду (Калабухов, 1969), можно предположить, что зимний период для вида в изученных условиях среднегорий также является наиболее неблагоприятным и энергозатратным. Высокий эритропоэз с продукцией большого количества мелких эритроцитов в зимний период года, когда температура среды значительно снижается, был выявлен в природе у ряда грызунов (Rewkieweir-Dziarska, 1975; Темботова и др., 1979, 1982; Темботова, Темботов, 1987, 1988; Темботов и др., 2009). Следует отметить, что степень активности эритропоэза в зимний период у разных видов разная, что, видимо, можно объяснить видоспецифичностью.
В летний период в периферическую кровь, в результате активного эритропоэза, из костного мозга поступает большое количество молодых эритроцитов (ретикулоцитов), что свидетельствуют о значительном обновлении крови. Эти эритроциты качественно другие в сравнении с зимними: они более крупные по своим размерам (диаметр, средняя площадь), но концентрация и содержание гемоглобина в них поддерживаются на уровне зимних значений. Количество эритроцитов, содержание гемоглобина в 1 мкл крови и гематокритный показатель летом ниже, по сравнению с зимними данными, и достигают минимальных значений в годовом цикле (за редким исключением). Анализ гематокритной величины показывает увеличение плазмы крови (~5%) летом.
Наиболее стабильными в течение года из изученных показателей системы крови домовой мыши в среднегорьях Центрального Кавказа являются средний объем эритроцитов, концентрация и содержание гемоглобина в эритроцитах.
Обнаруженные в эритроцитах домовой мыши в условиях гор Центрального Кавказа тельца Хауелла-Жолли (у 70% особей весной, в другие сезоны – до 50%) являются мелкими остатками ядерного материала (Гольдберг, Гольдберг, 1975; Риган и др., 2000). Присутствие их, возможно, объясняется повышенной скоростью продуцирования эритроцитов в момент регенеративного ответа. Их наличие в эритроцитах также отмечено нами ранее у малой лесной мыши на высоте 2000 м над ур. м. и домовой мыши (700 м над ур. м.) на Центральном Кавказе (Темботов и др., 2005; Темботова и др., 2007а). Риган с соавт. (2000) связывают наличие телец Хауелла-Жолли с неспособностью макрофагов полностью удалять ядерный материал из эритроцитов.
Сравнение сезонной динамики показателей периферической крови домовой мыши в среднегорьях и предгорьях Центрального Кавказа показало, что механизм адаптации к сезонным изменениям сходный: увеличение кислородной емкости крови зимой. Однако в ее способах обеспечения, а соответственно и в трендах изменчивости показателей крови у вида на двух высотных уровнях имеются некоторые различия. Как и в среднегорьях, зимой максимальные в предгорьях значения отмечены по содержанию гемоглобина, количеству эритроцитов, гематокритной величины, минимальные – по диаметру эритроцитов (Темботов и др., 2009). Стабильные в среднегорьях качественные характеристики содержания гемоглобина в эритроцитах (MCH, MCHC) претерпевают сезонные изменения: в зимний период увеличиваются, достигая максимальных в году значений. Следует отметить, что показатели красной крови (содержание гемоглобина, его концентрация и содержание в эритроците) в предгорьях даже зимой не достигают значений, характерных для среднегорий. Обитание домовой мыши в среднегорьях Кавказа вызывает напряжение со стороны эритрона и ускоренную регенерации крови (Емкужева, 2013). Можно полагать, что обитание в горах требует высокого энергообмена в течение всего года.
* * *
В функционировании системы крови синантропного вида – домовой мыши в условиях среднегорий (1800 м над ур. м.) Центрального Кавказа в течение годового цикла обнаружены следующие адаптивные преобразования.
У вида выявлена выраженная сезонная динамика деятельности костного мозга и состояния периферической крови. Годовой цикл делится на три периода: холодный (зима), теплый (лето) и межсезонье (весна, осень). Наиболее энергозатратным сезоном года для вида, как и у грызунов обитающих в естественной среде, является зимний период, что проявляется в максимальной активности костного мозга, приводящей к повышению респираторной функции крови и, соответственно, увеличению ее кислородной емкости. Как и у ряда других грызунов, активность работы костного мозга в зимний период сопровождается выработкой наибольшего числа эритроцитов с наименьшим их диаметром и высоким уровнем гемоглобина, что способствует более эффективному обеспечению организма кислородом. Летом состояние красной крови качественно иное, менее активный костный мозг вырабатывает наиболее крупные по диаметру эритроциты, но число их меньше.
В течение четырех месяцев с конца зимы к началу лета происходит значительное обновление эритроцитов в периферической крови, в результате эритродиереза большое число мелких эритроцитов замещается крупными, но в меньшем количестве. Осенью происходит обратный процесс.
Растянутый период размножения, охватывающий весь годовой цикл, практически не сказывается на состоянии системы крови. Установленные закономерности характерны для животных обоих полов.
В результате выявленных количественно-качественных сезонных изменений в кроветворении и периферической крови домовой мыши поддерживаются стабильные показатели количества гемоглобина в эритроцитах и в значительной степени обусловливает устойчивое функционирование организма в условиях гор Центрального Кавказа.
Из полученных данных можно заключить, что несмотря на круглогодичное обитание домовой мыши в жилищах человека в среднегорьях Центрального Кавказа система крови домовой мыши сохраняет сезонность.
Список литературы
Берчану Шт. Клиническая гематология. Бухарест: Медицинское изд-во, 1985. 1222 с.
Большаков В.Н. Пути приспособления мелких млекопитающих к горным условиям. М. : Наука, 1972. 199 с.
Воробьев А.И. Руководство по гематологии. М. 1985. Т. 1. 353 с.
Гольдберг Д.И., Гольдберг Е.Д. Справочник по гематологии. Томск: Томский гос. ун-т. 1975. 278 с.
Домовая мышь: Происхождение, распространение, систематика, поведение. М.: Наука, 1994. 267 с.
Емкужева М.М. Сравнительный анализ адаптивных реакций системы крови и интерьерных признаков дикоживущих и синантропных грызунов семейства MURIDAE к условиям гор центральной части Северного Кавказа // автореф. дис. канд. биол. наук; Саратов, 2013. 20 с.
Иванова Н.Т. Система крови. Материалы к сравнительной морфологии системы крови человека и животных. Ростов-на-Дону. 1995. 154 с.
Калабухов Н.И. Периодические (сезонные и годичные) изменения в организме грызунов, их причины и последствия. Л.: Наука, Ленингр. отд. 1969. 249 с.
Карасева Е.В., Телицина А.Ю. Методы изучения грызунов в полевых условиях: Учеты численности и мечение. М.: Наука, 1996. 227 с.
Кост Е.А. Справочник по клиническим лабораторным методам исследования. М.: Медицина, 1975. 436 с.
Кучерук В.В. Грызуны – обитатели построек человека и населенных пунктов различных регионов СССР // Общая и региональная териология. Сб. науч. тр. М.: Наука, 1988. С. 165–237.
Лохмиллер Р.П., Мошкин М.П. Экологические факторы и адаптивная значимость изменчивости иммунитета мелких млекопитающих // Сиб. экол. журн. Т. 6. № 1. 1999. С. 37–58.
Меньшиков В.В. Стандартизация аналитических технологий лабораторной медицины. М.: Лабора, 2012. 367 с.
Мосягина Е.Н. Эритроцитарное равновесие в норме и патологии. М.: Медицина, 1962. 298 с.
Обыкновенная полевка: виды-двойники. (Microtus arvalis Pallas, 1779 M. rossiaemeridionalis. Ognev, 1924), 1994. Отв. ред. Соколов В.Е., Башенина Н.В. М.: Наука, 431 с.
Риган В., Сандерс Т., Деникола Д. Атлас ветеринарной гематологии. М.: ООО “Аквариум ЛТД”, 2000. 136 с.
Соколов В.Е., Темботов А.К. Позвоночные Кавказа. Млекопитающие. Насекомоядные. М.: Наука, 1989. 547 с.
Стальмакова В.А. О некоторых физиологических особенностях домовой мыши – Mus musculus L. // Фауна и экология животных Таджикистана. Сб. Таджикистан, 1978. С. 81–91
Тарахтий Э.А., Давыдова Ю.А. Сезонная изменчивость показателей системы крови рыжей полевки (Clethrionomus glareolus) разного репродуктивного состояния // Изв. РАН. Сер. биол., № 1. 2007. С. 14– 25.
Тарахтий Э.А., Сумин М.Н., Давыдова Ю.А. Изменчивость показателей “красной” крови системы рыжей полевки (Clethrionomus glareolus) в зависимости от сезона и репродуктивного состояния // Успехи совр. биологии. 2009. Т. 129. № 2. С. 191–197.
Темботов А.К., Темботова Э.Ж., Берсекова З.А., Емкужева М.М. Влияние градиента высоты местности на гематологические показатели одного из широкораспространенных видов грызунов – малой лесной мыши (MURIDAE, RODENTIA) на Центральном Кавказе // Материалы междунар. конф. “Млекопитающие горных территорий”. М.: Т-во научных изданий КМК, 2005. С. 169–174.
Темботов А.К., Темботова Э.Ж., Темботова Ф.А., Емкужева М.М. Изменчивость гематологических параметров домовой мыши (MUS MUSCULUS L.) в предгорьях Центрального Кавказа // Успехи совр. биологии. 2009. Т. 129. № 4. С. 370–378.
Темботов А.К., Шхашамишев Х.Х. Животный мир Кабардино-Балкарии. Нальчик: Эльбрус. 1984. 190 с.
Темботова Э.Ж., Барагунова Е.А., Василенко В.Н. Сезонные изменения эритроидного ростка костного мозга и диаметра эритроцитов лесной мыши в условиях лесостепного пояса Центрального Кавказа // Межвед. сб. науч. тр. “Проблемы горной экологии”. Нальчик. 1982. С. 106–111.
Темботова Э.Ж., Василенко В.Н., Темботова Ф.А. Сезонные изменения гематологических показателей у Apodemus sylvaticus в условиях равнины Кабардино-Балкарской АССР // Межвед. сб. науч. тр. Нальчик. “Фауна, экология и охрана животных Северного Кавказа”. Нальчик. 1979. Вып. 4. С. 119–129.
Темботова Э.Ж., Емкужева М.М., Темботова Ф.А. Эколого-физиологический анализ эритрона домовой мыши (Mus musculus L.) в условиях высотной поясности Центрального Кавказа // Материалы междунар. конф. “Млекопитающие горных территорий”. М: Т-во научных изданий КМК, 2007а. С. 315–323.
Темботова Э.Ж., Емкужева М.М., Темботова Ф.А. Внутрипопуляционная изменчивость (сезонная, половая) морфофизиологических параметров домовой мыши (MURIDAE, RODENTIA) в предгорьях Центрального Кавказа // Горные экосистемы и их компоненты. Труды междунар. конф. М: Т-во научных изданий КМК, 2007б. C. 121–126.
Темботова Э.Ж., Темботов А.К. Система крови полевой мыши и ее сезонные изменения в условиях лесостепного пояса КБАССР // Межвед. сб. науч. тр. “Фауна и экология млекопитающих Кавказа”. Нальчик. 1987. С. 144–162.
Темботова Э.Ж., Темботов А.К. Система крови обыкновенной полевки (Microtus arvalis) и ее сезонные изменения в условиях предгорья КБАССР // Межвед. сб. науч. тр. “Вопросы горной экологии”. Нальчик. 1988. С. 125–145.
Тодоров И. Клинические лабораторные исследования в педиатрии. София: Медицина и физкультура. 1968. 874 с.
Holland R.A., Forster R.E. Effect of size of red cells of the kinetics of their oxygen uptake in different species. Federat. Proc. V. 2. 1966. P. 727–742.
Maclean G.S., Lee A.K. Effects of season, temperature and activity on some blood parameters of feral house mice Mus musculus // J. Mam., 1973. V. 54. № 3. P. 660.
Kostelecka-Myrcha A. Variation of morpho-physiological indexes of blood in Clethrionomus glareolus (Shreber, 1780) // Acta teriologica. 1967. V. XII. 13. P. 191–222.
Rewkieweir-Dziarska A. Seasonal changes in hemoglobin and erythrocyte indexes in Microtus arvalis (Pallas, 1779) // Bull. Acad. Polon Sci; Ser Sci: Biol. 1975. V. 23. № 7. P. 481–486.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая