

Известия РАН. Серия биологическая, 2022, № 2, стр. 151-159
Первые данные о личинках трематод у моллюска Hydrobia acuta в Черном море
Ю. В. Белоусова *
ФГБУН ФИЦ “Институт биологии южных морей им. А.О. Ковалевского РАН”
299011 Севастополь, пр. Нахимова, 2, Россия
* E-mail: julls.belousova@gmail.com
Поступила в редакцию 10.07.2020
После доработки 15.10.2020
Принята к публикации 15.10.2020
- EDN: DVDDXM
- DOI: 10.31857/S1026347022020044
Аннотация
Представлены сведения о видовом составе личинок трематод у брюхоногого моллюска Hydrobia acuta в Черном море. Впервые в акватории Севастополя обнаружено 7 видов гельминтов от моллюска Hydrobia: пять видов церкарий (Gynaecotyla adunca, Maritrema misenense, Timoniella imbutiformis, Cryptocotyle sp., Haplosplanchnus pachysoma, и три вида метацеркарий (Paratimonia sp., H. pachysoma, Proctoeces maculatus) Приведена морфологическая характеристика и показатели инвазии личинок трематод у исследуемого вида гастропод. Впервые моллюск H. acuta зарегистрирован как промежуточный хозяин для трематоды P. maculatus.
Черноморский моллюск Hydrobia acuta (Draparnaud, 1805) – наиболее массовый представитель малакофауны на распресненных мелководьях. Несмотря на свои небольшие размеры, этот вид моллюсков занимает видное место в бентосе, являясь важным источником пищи для многих видов рыб в морских заливах и бухтах.
В Черном море, в акватории Тулигульского лимана, у моллюсков H. ventrosa (Montagu, 1803) ранее отмечали несколько видов церкарий семейства Microphallidae: Сercaria xiphidocercaria (II Sordi, 1959), C. microphallidarum (Reimer, 1963), C. caribbea Cable. 1956 (Долгих, 1969), а у берегов Новороссийской бухты у этого же вида гастропод – церкарий С. kowalewski (Dolgikh, 1966) и С. xiphidocercaria (Долгих, 1966).
Литературные сведения о гельминтах H. acuta в акватории Черного моря отсутствуют, поскольку специальных гельминтологических исследований этого вида моллюсков не проводили.
Целью настоящей работы было показать видовой состав личиночных форм гельминтов, паразитирующих у наиболее массового представителя малакофауны H. acuta в распресненной акватории (эстуарий реки Черная) и мелководье (бухта Казачья) Севастополя.
МАТЕРИАЛЫ И МЕТОДЫ
Материалом для данной работы служили собственные сборы моллюсков из устья реки Черной (Севастопольская бухта) и бухты Казачья в период с ноября 2011 г.–ноябрь 2012 г. Методом полного гельминтологического вскрытия исследовано 6790 экз. гастропод H. acuta. Все ткани моллюсков обследованы компрессорным методом (Быховская-Павловская, 1969) под бинокуляром МБС-10 при увеличении ×98. Окрашивали трематод уксусным кармином по стандартной методике (Роскин, Левинсон, 1957) и после дегидратации спиртами (70–100°) и просветления в гвоздичном масле гельминтов заключали в канадский бальзам. Измерения гельминтов приведены в пределах: минимума – максимума, среднее и стандартная ошибка. Все рисунки выполнены в редакторе векторной графики в программе Inkscape 0.48.2.-1 (Scalable Vector Graphics, 2011). Использовали следующие показатели инвазии: экстенсивность инвазии (ЭИ), интенсивность инвазии (ИИ), индекс обилия (ИО)
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
У 5053 экз. H. acuta в обследованной акватории Севастополя обнаружено 7 видов личинок трематод: церкарии Maritrema misenense (Palombi, 1940; Prévot, Bartoli and Deblock, 1976), Gynaecotyla adunca (Linton, 1905; Yamaguti, 1939), Timoniella imbutiformis (Molin, 1859; Brooks, 1980), Haplosplanchnus pachysoma (Eysenhardt, 1829; Looss, 1902), Cryptocotyle sp. (Luhe, 1899) и метацеркарии H. pachysoma, Paratimonia sp. (Prevot and Bartoli, 1967; Proctoeces maculatus (Looss, 1901; Odhner, 1911).
Ниже приведены морфологические описания личинок гельминтов, наиболее часто встречаемых у исследуемых моллюсков H. аcuta.
Сем. Microphallidae (Ward. 1901)
Maritrema misenense (Palombi, 1940; Prévot, Bartoli and Deblock, 1976).
Локализация: гонада; oписание: основано на 20 экземплярах церкарий.
Показатели инвазии: ЭИ – 1%, ИИ 25 ± ± 5 экз./особь, ИО 0.01 ± 0.01.
Спороцисты Maritrema misenense овальной формы, содержали 15–30 зрелых церкарий. Родильная пора не обнаружена.
Мелкие церкарии имеют удлиненное тело, сплющенное в дорсо-вентральном направлении. Длина тела 80 ± 3 µm при ширине 51 ± 3 µm. Кутикула покрыта очень мелкими шипиками. Ротовая присоска, размером 20 ± 2 × 18 ± 1, субтерминальная. Брюшная присоска (10 ± 1 × 9 ± 0.8 µm) в два раза меньше ротовой и расположена посередине тела трематоды. Ротовая присоска снабжена стилетом размером 13 ± 1 µm цилиндрической формы. Коническая часть стилета немного короче, чем его основание (5 ± 0.3 µm), идущим из цента ротовой присоски. Поперечная щель в области ротовой присоски указывает на местоположения будущего рта.
Четыре пары желез проникновения открываются в передней части тела. Каналы желез открываются парами в области ротовой присоски. Железы проникновения, заполненные гранулированными включениями, достигают уровня брюшной присоски. Одноклеточные железы крупных размеров. Границы между клетками плохо выражены. Протоки желез представлены извитыми каналами. Пищеварительная система не развита. Выделительный пузырь может иметь щелевидную или пузыревидную форму. Хвост простого строения длиной 60–105 µm, сужается к своему концу (рис. 1).
Рис. 1.
Микрофотография церкарии Maritrema misenense в акватории бухты Казачья: 1 – стилет, 2 – ротовая присоска, 3–зачатки брюшной присоски, 4 – железистые клетки, 5 – выделительный пузырь. Масштаб 50 µm.
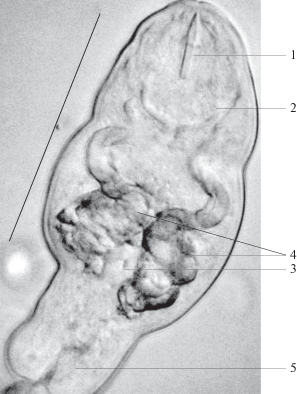
Двигаются церкарии телом вперед, однако, что характерно, могут с легкостью отбрасывать свой хвост, ползая с помощью присосок.
Gynaecotyla adunca (Linton, 1905; Yamaguti, 1939).
Локализация: гонада. Описание основано на 10 экз. церкарий. Показатели инвазии личинками: ЭИ – 1%, ИИ-1 – 15 экз./особь, ИО – 0.007 ± 0.004.
Партениты обнаружены в гонадах брюхоногих моллюсков. Представлены несколькими генерациями спороцист, имеющих в зависимости от сезона различный состав гемипопуляций. Материнская спороциста не найдена.
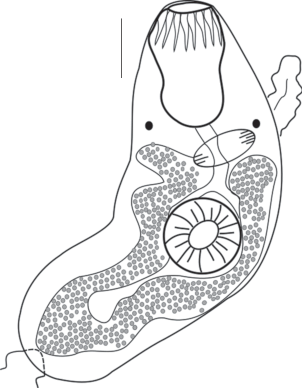
Церкарии мелкие. Тело церкарий овальной формы. Длина их тела 84 ± 3 µm, ширина – 36 ± 2 µm. Ротовая присоска размером 16 ± 1 × 15 ± 0.5 µm, расположена субтерминально и вооружена крупным стилетом (10 ± 0.5 µm), идущим из центра ротовой присоски. Острие стилета может немного выступать над передним краем тела церкарии. Две пары стилетных желез открываются в передней части тела у начала ротовой присоски. Зачатки брюшной присоски просматриваются между железами проникновения. Пищеварительная система не развита. Выделительный пузырь имеет Y-образную форму. Выделительная система трематоды хорошо дифференцируется на живых объектах. Формула экскреторной системы 2 (2 + 2) = 8. Хвост длиной 78 ± 3 µm, сужается к своему концу (рис. 2).

Сем. Cryptogonimidae (Ward, 1917)
Timoniella imbutiformis (Molin, 1859; Brooks, 1980). Локализация: печень. Описание по 2 экз. церкарий. Показатели инвазии личинками: ЭИ – 2%, ИИ – 1-1 экз./особь, ИО – 0.007 ± 0.004.
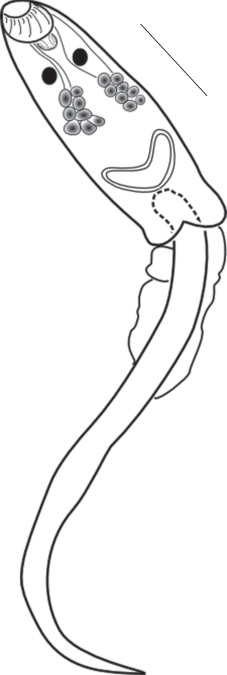
Тело личинки вытянутое, сплющено в дорсо-вентральном направлении. Длина тела 425 ± 25 µm, ширина 176 ± 10 µm. Воронкообразная ротовая присоска размером 132 ± 8 × 73 ± 3 µm, вооружена 18 шипами, в два раза крупнее брюшной (73 ± 3 × × 73 ± 3 µm). Префаринкс относительно мелкий, фаринкс небольших размеров (30 ± 2 × 73 ± 3 µm), короткий пищевод разветвляется над брюшной присоской. Кишечные ветви достигают заднего конца тела. На уровне фаринкса расположены два пигментных глазка. Формула выделительной системы 2 [(2 + 2) + (2 + 2)] = 16. Выделительный пузырь Y –образной формы (рис. 3).
Сем. Heterophyidae (Leiper, 1909)
Cryptocotyle sp. (Lűhe, 1899). Локализация: пищеварительная железа.
Показатели зараженности: ЭИ – 7%, ЭИ – 25 ± 5 экз./особь, ИО – 0.08 ± 0.02. Описание основано на 20 экз. церкарий от моллюсков.
Длина тела фиксированных церкарий 143 ± ± 24 µm, ширина – 57 ± 10, длина ротовой присоски 22 ± 5 µm, ширина – 23 ± 5. Фаринкс мелкий, ярко выраженный. На уровне глотки расположены 2 пигментных глазка. Пищевод и кишечник отсутствуют. Особое строение системы желез проникновения характерно для церкарии рода Cryptocotyle. Железы проникновения (8 пар) находятся между глоткой и выделительным пузырем. Выделительный пузырь чашевидной формы. Длина хвоста церкарии 200 ± 60 µm, он в 1.5 раза превышает длину тела и по бокам снабжен плавательной мембраной (рис. 4)
Церкарии Cryptocotyle после выхода из моллюска-хозяина энергично плавают. Процесс их движения отчетливо разделяется на две чередующиеся фазы: активную и пассивную. В ходе первой личинка активно работает хвостом, что обеспечивает ее поступательное движение вперед. Во время второй фазы она замирает и в таком состоянии парит в воде, медленно опускаясь вниз.
Сем. Haplosplanchnidae (Poche, 1926)
Haplosplanchnus pachysoma (Eysenhardt, 1829; Looss, 1902).
Локализация: полость тела. Описание: по 3 экз. адолескарий.
Адолескарии крупных размеров, длина тела 1000 ± 40 µm при ширине 360 ± 40 µm. Тело трематоды имеет треугольную форму. Брюшная присоска (VS) очень мощная, размером 294 × 168 µm. Ротовая присоска (OS) размером 120 ± 10 × 130 ± 10 µm, у живых особей размещена субтерминально, у фиксированных — несколько смещена назад от переднего конца тела. Отношение 2.5 ≤ VSL/OSL ≤ 3. Префаринкс короткий, длиной 70 ± 10 µm. Фаринкс овальной формы, хорошо развит, 70 ± 5 × × 70 ± 8 µm. Кишечник непарный, длиной 215 ± ± 1 µm, простирается до заднего конца брюшной присоски. Между фаринксом и передним краем брюшной присоски располагается большое количество относительно мелких железистых клеток. Возможно, что это цистогенные железы. Зачатки половых желез не отмечены. (рис. 5а). Циста метацеркарии не обнаружена.
Рис. 5.
Haplosplanchnus pachysoma в эстуарии реки Черная. (а) – личинка (рисунок живого червя), масштаб 500 µm; (б) – микрофотография церкарии (живой гельминт): 1 – ротовая присоска, 2 – брюшная присоска, 3 – кишечник, 4 – желeзистые клетки. Масштаб 100 µm.
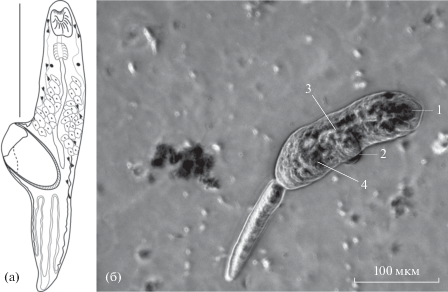
Церкария H.pachysoma (1 экземпляр) была отмечена единожды в июле 2019 г. (рис. 5б).
Тело личинки вытянутой формы. Ротовая присоска небольших размеров, несколько смещена от переднего конца тела. На уровне ротовой присоски имеются два пигментных глазка. Брюшная присоска мощнее ротовой. Префаринкс очень короткий, переходит в хорошо выраженный овальный фаринкс. Кишечник непарный, простирается до заднего конца брюшной присоски. Между фаринксом и брюшной присоской располагается большое количество относительно мелких железистых клеток. Экскреторный тонкостенный мочевой пузырь объeмной V-образной формы. Плавают церкарии телом вперед при помощи простого хвоста.
сем. Fellodistomidae (Nicoll, 1909)
Proctoeces maculatus (Looss, 1901; Odhner, 1911). Локализация: полость тела. Показатели инвазии: ЭИ – 1%, ИИ – 2 ± 1 экз./особь, ИО – 0.002 ± 0.001. Описание основано на 5 экз. прогенетических метацеркарий.
Тело трематоды удлиненное, суженное к переднему и заднему концам. Длина тела 1631 ± 75 µm, ширина 550 ± 68 µm. Кутикула гладкая. Ротовая присоска, длиной 141 ± 6 µm, шириной 163 ± 7 µm, расположена субтерминально. Мускулистая брюшная присоска, длиной 243 ± 6 µm, шириной 256 ± 6 µm, располагается в передней трети тела. Фаринкс хорошо развит. Длина фаринкса 113 ± ± 7 µm, ширина – 113 ± 7 µm. Пищевод короткий. Кишечные ветви почти достигают заднего конца тела. При фиксации трематода несколько изогнулась, поэтому половые органы слегка сместились. Овальные цельнокрайние семенники, длиной 146 ± ± 11 µm, шириной 95 ± 5 µm, располагаются один над другим. Округлый яичник находится впереди семенников. Длина яичника 119 ± 24 µm, ширина 101 ± 23 µm. Матка сильно развита и заполняет всю часть тела за брюшной присоской Яйца удлиненно- овальной формы, желтоватого окраса. Длина яиц составляет 48 ± 2 µm, ширина – 24 ± 1 µm (рис. 6).
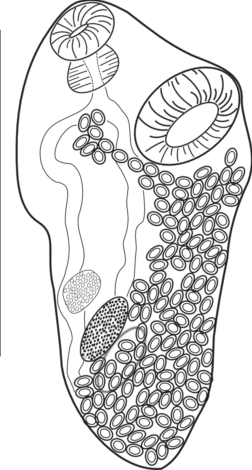
Cем. Monorchiidae (Odhner, 1911)
Paratimonia sp. (Prevot and Bartoli, 1967). Локализация: полость тела.
Показатели инвазии: ЭИ – 1%, ИИ – 1.33 ± ± 0.33 экз./особь, ИО – 0.014 ± 0.009. Описание основано на двух метацеркариях.
Обнаруженные нами метацеркарии Paratimonia sp. имеют овальную форму тела. Длина тела – 868–882 µm, ширина – 364–560 µm. У живых особей просматриваются мелкие шипики на покровах. Мощная брюшная присоска длиной 252–280 µm и шириной 182–266 µm в два раза больше ротовой (длина – 126–140 µm, ширина – 154–182). Префаринс короткий, фаринкс крупный, круглой или овальной формы. Короткий пищевод разветвляется над брюшной присоской. Кишечные ветви достигают нижней границы верхнего семенника. На уровне фаринкса и ротовой присоски расположены два крупных пигментных глазка, являющиеся систематическим признаком рода. Экскреторный пузырь вытянутой формы, в виде ампулы. Семенники лежат в диагональ друг над другом, между ветвями кишечника. Яичник лежит над семенниками и сдвинут вправо. В задней части тела четко просматривается сформированная половая бурса. Половая пора видна (рис. 7).
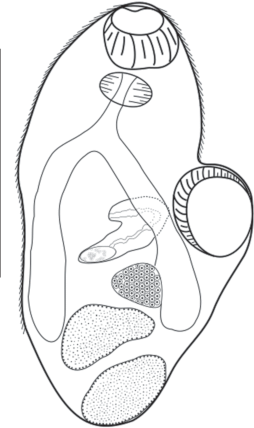
Наши исследования этой личинки показали, что, по всей вероятности, для этого вида трематод возможно ускорение жизненного цикла, при котором происходит преждевременное развитие всех систем органов, в особенности половой системы, характеризующих взрослую особь трематоды. Брюхоногие моллюски H. acuta, выступающие в таком случае в качестве паратенического хозяина, видимо, могли заразиться в результате контакта с церкариями, покинувшими первого промежуточного хозяина (двустворчатых моллюсков Abra segmentum, разделяющих с гидробией общий биотоп).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Целенаправленные фаунистические исследования трематод черноморских моллюсков естественных поселений в акватории Севастополя впервые были начаты в 1909–1911 гг. Д.О. Синицыным (Синицын, 1911). Им была опубликована первая сводка “Партеногенетическое поколение трематод и его потомство в черноморских моллюсках”. В данной работе отмечено паразитирование 25 видов церкарий и метацеркарий у 11 видов черноморских моллюсков и приведены морфологические описания найденных личинок трематод. Только через полвека данные исследования были возобновлены А.В. Долгих (Долгих, 1965). Было обследовано 40 видов моллюсков из 20 семейств, (17% от черноморской фауны моллюсков), и у 21 вида зарегистрировано 32 вида личинок трематод (2 вида из сем. Bucephalidae, 1 вид – из сем. Diplostomatidae, 4 вида – из сем. Opecoelidae, 1 вид – из сем.Acanthocolpidae, 1 вид – из сем. Zoogonidae, 4 вида –из сем. Fellodistomatidae, 2 вида из сем. Gymnophallidae, 1 вид – из сем. Plagiorchioidea, 3 вида – из сем. Microphalidae, 2 вида – из сем. Opisthorchioidae, 1 вид – из сем. Monorchidae, 2 вида –из сем. Lepocreadiidae, 1 вид – из сем. Haploporidae, 7 видов – из сем. Hemiuroidae), в т. ч. церкарии 27 видов трематод и метацеркарии 5 видов трематод (из них 2 – прогенетические). Из них 12 видов личинок трематод были описаны как новые для науки; 9 видов отмечены ею впервые у моллюсков Черного моря. (Долгих, 1965) Из устного сообщения А.В. Гаевской (Долгих) известно, что в 1960-х гг. были начаты работы по изучению трематод H. acuta, но результаты ее исследований не были опубликованы. Таким образом, настоящая работа является продолжением гельминтологического мониторинга с одной стороны и первой работой по изучению трематодофауны H. acuta для данной акватории.
Из сем. Heterophyidae нами отмечены церкарии рода Cryptocotyle, впервые описанные Ф. Креплином (Creplin, 1825). Однако наибольший вклад в изучение особенностей морфологии и жизненного цикла этой трематоды внес Г. Стэнкард (Stunkard, 1930). В своей, ставшей классической, работе Стэнкард рассмотрел историю рода, жизненный цикл, описал ряд стадий паразита, паразито-хозяинные взаимоотношения, морфологические особенности церкарии и метацеркарии. Им был поставлен ряд опытов с целью описания ранних стадий развития паразита. Обнаруженные нами церкарии наиболее близки Cryptocotyle по основным морфологическим признакам (форма тела, строение хвоста, строение системы желез проникновения) и промерам.
Из сем. Microphallidae нами обнаружены церкарии двух видов – Maritrema misenense и Gynaecotyla adunca. Последние впервые были описаны Паломби (Palombi, 1940) от моллюсков Cerithium vulgatum из Средиземного моря. Ф. Анкел (Ankel, 1962) отмечает этих церкарий как Cercaria misenensis у 2 видов гидробий – Hydrobia ulvae (Pennant, 1777) и H. ventrosa (Montagu, 1803) у берегов Дании. Похожие по морфологическим особенностям трематоды были обнаружены М. Сорди (Sordi, 1959) от Paludestrina salinasi Aradas Calcara, 1842 из окрестностей Ливорно (Италия). В Черном море до нас микрофаллидную церкарию G. adunca находила А.В. Долгих (1965) под названием C. misenensis. Церкарии G. adunca (syn. Cercaria sevillana) из Португалии от моллюска Nassarius reticulatus (Linnaeus, 1758) (Russel-Pinto, Bartoli, 2002) идентичны церкариям, найденным Долгих (1965) в том же виде моллюска в акватории Черного моря, однако ошибочно названы Cercaria misenensis, которые являются паразитами моллюсков Cerithium rupeste, Risso, 1826. Cercaria sevillana отличается от Cercaria misenensis формой и размером выделительного пузыря, длиной и формой хвоста, а также поведенческими реакциями. Эти морфологические отличия были доказаны в работе португальских авторов (Russel-Pinto, Bartoli, 2002). Наша находка церкарии по всем параметрам совпадает c последним морфологическим описанием (Russel-Pinto, Bartoli, 2002): Y-образная форма мочевого пузыря обнаруженной нами церкарии G.adunca vs грушевидной формы С. misenensis, (Долгих, 1965), ребристая структура хвоста vs гладкой структуры C.misenensis (Долгих, 1965), положение и форма стилета.
Обнаруженные нами церкарии второго вида микрофаллид Maritrema misenense по размеру и основным морфологическим признакам, включая форму стилета, конфигурация которого является важнейшим диагностирующим признаком церкарий микрофаллид, сходна с формой стилета особей M. misenense ранее описанных в литературе (Долгих, 1965): положение и форма стилета, грушевидная форма тела, гладкая структура vs ребристой структуры хвоста у других представителей сем. Microphallidae, (Долгих, 1965). Указанные морфологические характеристики и позволяют нам отнести обнаруженную микрофаллидную церкарию к виду Maritrema misenense.
Личиночная стадия H. pachysoma впервые зарегистрирована нами в акватории Севастополя (Севастопольская бухта, река Черная). Личинка на стадии церкарии и жизненный цикл H. pachysoma были впервые подробно изучены Кэбэлом (Cable, 1954). Позднее Ферес и Мэйлрейд (Fares, Maillard, 1975) описали жизненный цикл H. pachysoma. Авторы отмечают, что церкарии развиваются в брюхоногом моллюске H. ventrosa, через некоторое время церкарии, покинув своего первого промежуточного хозяина, инцистируются. Для трематоды H. pachysoma кефалевые являются окончательным хозяином и заражение рыб происходит через адолескарий (Fares, Mailard, 1975). У исследованных моллюсков H. acuta в начале весны и осенью были найдены крупные неинцистированные личинки трематод, а в июле 2019 г. зарегистрирована личинка на стадии церкарии у этого же вида моллюсков. Морфология обоих видов личинок соответствовала диагнозу рода Haplosplanchnus: удлиненно-овальная ротовая присоска, крупная брюшная присоска своеобразного строения (она глубоко погружена в тело, при рассмотрении сбоку имеет мешковидную или цилиндрическую форму, снабжена мускулистым сфинктером), непарный кишечник.
В акватории Севастополя зарегистрированы церкарии Timoniella imbutiformis, вторым промежуточным хозяином которых являются бычки Neogobius melanostomum. Первое описание церкарий этого вида было сделано Ребеком в 1960 г. (Rebecq, 1960) из Средиземного моря. В работе Мэйларда (Maillard, 1973) упоминается, что первым промежуточным хозяином для этого вида трематод являются брюхоногие моллюски рода Hydrobia, а вторым промежуточным хозяином для T. imbutiformis являются 4 вида бычков (Квач, 2004). Последняя по времени ревизия рода Timoniella (Kvach., 2017) показала, что для идентификации вида T. imbutiformis в качестве основных диагностирующих признаков указывают количество шипов на ротовой присоске (17–20) и расположение кишечной ветви. Обнаруженные нами церкарии этого вида трематод наиболее близки к виду T. imbutiformis количеством шипиков на ротовой присоске (18) и локализацией кишечной ветви, которая слепо заканчивается и достигает заднего конца тела.
Краткое описание метацеркарий и половозрелых особей Paratimonia в Черном море впервые приведено Н.Н. Найденовой в 1970 г. (Найденова, 1974). В Черном море зарегистрированы мариты одного вида этого рода – Paratimonia gobii. Последние сведения о морфологии марит рода Paratimonia приведены в работе Брэя с соавт. (Bray et al., 2008) Согласно литературным данным (Найденова, 1974; Bray et al., 2008): отсутствие префаринкса, наличие короткого пищевода и кишечных ветвей, достигающих нижней границы семенника, строение половой системы позволяют нам отнести обнаруженных метацеркарий к роду Paratimonia.
В условиях Черного моря промежуточным хозяином для Proctoeces maculatus зарегистрирована мидия Mytillus galloprovincialis (Мачкевский, 1984). Дополнительными хозяевами выявлены бентосные моллюски, такие как: Rissoa labiosa, R. splendida. Tricolia pulla, M. galloprovincialis (Долгих, 1965) и полихеты: Nereis diversicolor, N. succinea. Дефенетивными хозяевами являются в Черном море являются рыбы семейства Labridae и Gobiidae. Впервые в 1959 г. (Stunkard, Uzmann, 1969) была описана прогенетическая стадия Proctoeces maculatus из мидии Mytillus edulis. В одной и той же особи мидии авторами были отмечены спороцисты с церкарями, неинцистированные метацеркарии и мариты. О случаях находок прогентических метацеркарий у моллюсков сообщается и в других работах (Dolfus, 1964; Долгих, 1965, 1967). У берегов Крымского побережья прогенетические метацеркарии Proctoeces maculatus впервые обнаружены Долгих (1967) у моллюсков Rissoa splendida. H. acuta впервые регистрируется в качестве промежуточного хозяина для P. maculatus
Таким образом, в результате исследования гастропод H. acuta получены новые сведения о составе их трематодофауны в Черном море. Установлено, что в акватории Севастополя фауна трематод исследуемых брюхоногих моллюсков представлена 7 видами, относящихся к 6 семействам: Microphallidae, Monorchiidae, Cryptogonimidae, Fellodistomidae, Haplosplanchidae, Heterophyidae.
Значительную часть из всех трематод моллюсков составляют церкарии, для которых окончательными хозяевами являются рыбы. Наиболее часто инвазируют гидробий личинки трематод сем. Heterophyidae и Microphallidae, крайне редко встречаются личинки Paratimonia sp. и Haplosplanchnus pachysoma.
Благодарности. Автор выражает благодарность М.В. Макарову (ФИЦ ИнБЮМ) за помощь в сборе и определении видового состава моллюсков и Ю.М. Корнийчук за научную консультацию при подготовке статьи к печати.
Конфликт интересов: автор рукописи заявляет об отсутствии конфликта интересов.
Этические нормы: Это исследование было проведено в соответствии с этикой и благополучием животных. Все исследования выполняли в соответствии с Законом о ветеринарной и медицинской деятельности и Национальным законом о защите животных.
Финансирование. Работа выполнена в рамках государственного задания ФГБУН ФИЦ ИнБЮМ им. А.О Ковалевского РАН (тема № 121030100028-0).
Список литературы
Быховская–Павловская И.Е. Паразитологическое исследование рыб. Ленинград: Наука, 1969. 108 с.
Долгих А.В. Личинки трематод паразиты моллюсков крымского побережья Черного моря: Автореферат дис. канд. биол. наук. Севастополь-Львов, 1965. 24 с.
Долгих А.В. К изучению гельминтофауны моллюсков Новороссийской бухты // Гельминтофауна животных южных морей. Киев: Наукова думка. 1966. С. 114–133.
Долгих А.В. О прогенезе у трематод рода Proctoeces Odhner, 1911 // Паразитология. 1967. Т. 1. Вып. 3. С. 219–221.
Долгих А.В. О зараженности гидробии выпуклой Hydrobia ventrоsa (Mont.) Тилигульского лимана личинками микрофаллид // Вестник зоологии. 1969. № 4. С. 32–35.
Квач Ю.В. Фауна гельмінтів бичкових риб (Gobiidae) Тилігульського лиману Чорного моря // Вісник Львівського унів. Серія біологiчна. 2004. Выпуск 37. С. 144–148.
Мачкевский В.К. Биология и экология трематоды Proctoeces maculatus – паразита черноморских мидий. Автореферат дис. … канд. биол. наук. Москва, 1984. 24 с.
Найденова Н.Н. Паразитофауна рыб семейства бычковых Черного и Азовского морей. Киев: Наукова думка, 1974. 182 с.
Синицын Д.О. Партеногенетическое поколение трематод и его потомство в черноморских моллюсках. Санкт-Петербург, 1911. 127 с.
Роскин Г.И., Левинсон Л.Б. Микроскопическая техника: учеб. пособие для вузов. Под ред. Г.И. Роскина. 3-е изд. М.: Совет. Наука, 1957. 466 с.
Ankel F. Hydrobia ulvae Pennant and Hydrobia ventrosa Montagu als Wirte larvaler Trematoden // Videnskabelige Meddelelser Dansk Naturhistorisk Forening. 1962. Bd 124. S. 1–100.
Bray R.A., Gibson D., Jones A. Keys to the Trematoda. V. 3. CAB International and Natural History Museum, 2008. 824 p.
Cable R.M. Studies on marine digenetic trematodes of Puerto Rico. The life cycle in the family Haplosplanchnidae // J. Parasitology. 1954. V. 40. P. 71–76.
Creplin F.C. Observationes de entozois. Pars I. Gryphiswaldiae, 1825. 86 p.
Dollfus R. Metacercaria: Proctoeces progeneticus (Trematoda: Digenea) chez une Gibbula (Gastropoda prosobranchiata) de la cote atlantique du Maroc. Observations sur la famille Fellodistomatidae. Ann. Parasitol. Hum. Comp. 1964. V. 39. № 6. P. 755–774.
Fares A., Maillard C. Cycle evolutis de Haplosplanchnus pachysomus (Eyesenhardt, 1829). Loos, 1902 (Trematoda, Haplosplanchnidae), parasite de Mugilides (Teleostei) // Bulletin du Museum national d’histoire naturelle. 1975. V. 312. P. 837–844.
Inkscape 0.48.2.-1 2. Scalable Vector Graphics (SVG) (WEB: http://www.inkscape.org), 2011.
Kvach Y. The taxonomic and phylogenetic status of digeneans from the genus Timoniella (Digenea: Cryptogonimidae) in the Black and Baltic seas // J. Helminthology. 2017. V. 92. P. 596–603.
Maillard C. Etude du cycle évolutif du Trématode: Acanthostomum imbutiforme (Molin, 1859) Gohar, 1934, parasite de Morone labrax (Linné, 1758) // Ann. Parasitol. Hum. Comp. 1973. V. 48. P. 33–46.
Palombi A. Gli stadi larvali dei trematodi del Golfo di Napoli. 1°. Contributo allo studio della morfologia, biologia e sistematica delle cercarie marine // Pubblicazioni Stazione Zool. Napoli. 1940. V. 14. P. 51–94.
Rebecq J. Timoniella atherinae nov. gen., nov. sp. (Trematoda, Acanthostomatidae), metacercaire parasite d’Atherina mochon C.V. In: Libro homenaje al Dr. Eduardo Gaballero y Gaballero, jubileo 1930–1960. Institute Politecnico Nacional. Mexico. P. 257–262.
Russell–Pinto F., Bartoli P. Cercaria sevillana n. sp., a new cercaria (Digenea: Microphallidae) from Nassarius reticulatus (L.) (Mollusca: Prosobranchia) in Portugal // Syst. Parasitology. 2002. V. 53. P. 175–182.
Sordi M. Ricerche sulla biologia delle acque salmastre del Livornese. Trematodi parassiti di Paludestrina salinasi .Aradas Caleara 1842. Monitore zoologico.italiano. 1959. V. 66. P. 1.
Stunkard H.W. The life history of Cryptocotyle lingua (Creplin), with notes on the physiology of the metacercariae // J. Morphology and Physiology. 1930. V. 50. P. 143–193.
Stunkard H., Uzmann J. The life-cycle of the Digenetic trematode Proctoeces maculatus (Looss, 1901) Odhner, 1911 (syn. P. subtenuis (Linton, 1907) Hanson, 1950) and the description of Cercaria adranocerca. Biol. Bull. 1969. V. 116. № 1. P. 184–193.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая