

Известия РАН. Серия биологическая, 2022, № 6, стр. 661-667
О периферической крови у трех видов рыб с разной экологией (Scombridae и Berycidae)
А. И. Никитенко 1, *, Г. И. Пронина 2, 3, А. М. Орлов 6, 8, 7, 5, 9, Д. В. Артеменков 4, А. Н. Строганов 10, В. А. Беляев 4
1 Филиал по пресноводному рыбному хозяйству ФГБНУ ВНИРО
141821 Московская область, пос. Рыбное, Дмитровский р-н, Россия
10 Московский государственный университет имю М.В. Ломоносова
119991 Москва, Россия
2 ВНИИ ирригационого рыбоводства
142460 Московская обл., Ногинский р-н, пос. Воровского, Россия
3 Российский государственный аграрный университет – МСА им. акад К.А.Тимирязева
127550 Москва, ул. Тимирязевская 49, Россия
4 ВНИИ рыбного хозяйства и океанографии
105187 Москва, Окружной проезд 19, Россия
5 Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский просп. 33, Россия
6 Дагестанский государственный университет
367000 Махачкала, ул. Гаджиева 43а, Россия
7 Прикаспийский институт биологических ресурсов ДНЦ РАН
367000 Махачкала, ул. Гаджиева 45, Россия
8 Томский государственный университет
634050 Томск, просп. Ленина 36, Россия
9 Институт океанологии им. П.П. Ширшова РАН
117218 Москва, Нахимовский просп. 36, Россия
* E-mail: alexey_nikitenko90@mail.ru
Поступила в редакцию 14.05.2021
После доработки 08.06.2021
Принята к публикации 10.06.2021
- EDN: YFWFCM
- DOI: 10.31857/S1026347022060117
Аннотация
На основании материалов научного мониторинга на промысловых судах в акватории исключительных экономических зон Марокко и Мавритании в 2004–2017 гг. впервые представлены гематологические параметры периферической крови скумбрии Scomber colias, пеламиды Sarda sarda и берикса Beryx splendens. При описании клеток эритропоэтического ряда отмечен факт большего количества незрелых клеток у берикса, что, вероятно, связано с его пищевыми вертикальными миграциями на большие глубины с высоким давлением и пониженным содержанием кислорода. Анализ эритропоэза и лейкоцитарной формулы рыб позволил выявить особенности физиологического и иммунологического характера. Так, у пеламиды отмечено более высокое содержание моноцитов, чем у скумбрии и берикса, что свидетельствует о высоком уровне врожденного клеточного иммунитета, представленного фагоцитозом.
Кровь является информативным индикатором состояния организма рыб, отражающим влияние физико-химических факторов среды, сезона и экологии вида, как для отдельно взятой особи, так для популяций в целом (Головина, Тромбицкий, 1989; Камышников, 2004). В условиях различных экологических зон Мирового океана особую актуальность приобретают исследования закономерностей физиологических адаптаций рыб к изменяющимся внешним факторам. Клетки эритропоэтического ряда и лейкоцитов выполняют разнообразные физиологические и иммунологические функции, защищают организм от чужеродных тел и обеспечивают адаптацию рыб к изменяющимся внешним факторам (Микряков, Балабанова, 1979; Secombes, 1996; Галактионов, 2005). Особенности вида и среды обитания отражаются на составе эритроцитов и лейкоцитов рыб, основными типами которых являются зрелые и базофильные эритроциты, нормобласты, а также лимфоциты, моноциты, нейтро-, эозино- и базофилы и небольшая доля незрелых форм лейкоцитов. Соотношение отдельных типов клеток может свидетельствовать о физиологическом состоянии (или статусе) рыб и наличии биотических и абиотических стресс-факторов (Иванова, 1983; Parish et al., 1986; Головина, Тромбицкий, 1989; Житенева и др., 1989; Точилина, 1994; Микряков и др., 2001).
Достаточно много исследований посвящено изучению клеточного состава крови рыб, обитающих в пресноводных и солоноватоводных экосистемах (Иванова, 1983; Головина, Тромбицкий, 1989; Точилина, 1994). Несмотря на свое широкое распространение и значительную численность в Мировом океане, скумбрия (Scomber colias Gmelin, 1789), пеламида (Sarda sarda Bloch, 1793) и берикс (Beryx splendens Lowe, 1834) обитают в различных экологических условиях и отличаются своеобразным пищевым поведением (Nielsen, 1979; Maul, 1990; Collette, 2003). Они давно привлекают внимание специалистов, которыми в большей степени исследованы процессы роста, особенности морфологии, физиологии, биохимии, химического состава тела и т.д. Однако к настоящему времени сведения об особенностях морфофизиологических перестроек форменных элементов крови в процессах адаптации к изменяющимся внешним факторам, отмеченных у трех видов рыб семейств скумбриевые Scombridae и бериксовые Berycidae, в целом крайне малочисленны (Аминева, Яржобек, 1984; Shimizu, 1984; Collette, 2003).
Скумбрия S. colias, пеламида S. sarda и берикс B. splendens – условно хищные и хищные рыбы средних и крупных размеров, обитающие в умеренных и субтропических водах. Первый вид является прибрежным пелагическим, поэтому втречается на глубинах от поверхности до 300 м, обычен в диапазоне глубин до 110 м. Ареал скумбрии расположен в теплых водах восточной части Атлантического океана и в Средиземном и Черном морях (Collette, Nauen 1983; Парин, 1988). Пеламида является исключительно эпипелагическим видом, встречается в диапазоне глубин до 200 м и иногда заходит в устья. Пеламида распространена преимущественно в восточной части Атлантического океана от Норвегии до Южной Африки, в том числе Средиземном и Черном морях, встречается в западной Атлантике от Канады до северной части Мескиканского залива, также зарегистированы поимки у берегов Колумбии и Венесуэлы (Collette, 1986; Maigret, 1986). Последний вид распространен по всему миру в регионах континетального склона в Мексиканском заливе и на подводных горах в Атлантическом, Индийском и Тихом океанах, за исключением восточной и северной части Тихого океана. Берикс обычно обитает на глубинах от 150 до 800 м, встречается – от 10 до 1300 м (Heemstra, 1986; Paxton, 1999; Ramos et al., 2001; Козлов, 2004; Moffitt et al., 2004).
Цель данной работы – исследование гематологических параметров периферической крови трех видов рыб с различной экологией (S. colias, S. sarda, B. splendens) семейств скумбриевые (Scombridae) и бериксовые (Berycidae) в центрально-восточной части Атлантического океана.
МАТЕРИАЛЫ И МЕТОДЫ
Рыб отлавливали во время научного мониторинга на промысловых судах в акватории исключительных экономических зон Марокко и Мавритании между 16°0′–23°58′ с.ш., 16°25′–17°50′ з.д. в феврале-декабре 2004–2017 гг. на глубине от 35 до 270 м разноглубинным тралом типа Sirius 4, вертикальным раскрытием 55–80 м, горизонтальным – 140–160 м, канатная и сетная части которого изготовлены из современных облегченных материалов, размеры ячеи в крыльях 50–70 мм и кутце 16–24 мм. При проведении биологического анализа ежедневно из улова отбирали случайную пробу, в которой определяли и измеряли: длину рыб по Смитту, массу тела, пол, стадию зрелости гонад, балл наполнения желудков с указанием преобладающих объектов питания (Изучение экосистем …, 2004). За весь период исследований обработано 7812 экз. трех видов рыб (S. colias, S. sarda, B. splendens).
Отобранные особи берикса в биоанализе имели размер от 11 до 27 см и массу от 26 до 544 г, что отличается от литературных данных, где отмечены особи достигшие размеров 70 см в Мексиканском заливе, 70 см в Атлантическом, 44 см в Индийском и 50 см в Тихом океанах, обычный размер составляет 40 см (Parin et al., 1995; Lehodey, Grandperrin, 1996; Sommer et al., 1996; Santamaria et al., 2006). Пеламида может достигать длины 61 см в Средиземном море (Campo et al., 2006; Zorica, 2008) и 91 см в Центрально-Восточной Атлантике, обычный размер – 50 см (Collette, Nauen 1983; Гущин, Кортен, 2016). Проанализированные экземпляры в период научного мониторинга – от 23 до 65 см; масса тела – от 250 до 3455 г. Особи скумбрии в биоанализе – от 12 до 46 см; масса от 123 до 1440 г, по литературным данным скумбрия может достигать 50 см, обычный размер составляет 30 см (Torres et al., 2012; Гущин, Кортен, 2017).
Для проведения клинических исследований рыб в 2017 г. взяты 10 проб из уловов 4 тралений (скумбрия – 4 экз., пеламида – 3 экз., берикс – 3 экз., табл. 1). У свежевыловленных рыб осуществляли взятие проб крови из хвостовой артерии. Сразу изготавливали препараты на предметных стеклах в 2-х повторностях на одну пробу. После изготовления мазки крови высушивали в темном проветриваемом месте до исчезновения влажного блеска. Высушенные мазки хранили при комнатной температуре, обернув мазки в бумагу, и транспортировали в лабораторию для анализа (Иванов и др., 2013).
Таблица 1.
Характеристика объектов исследования хищных рыб семейств Scombridae (скумбрии, пеламиды) и Berycidae (берикса)
| Вид | Длина, см | Масса, г | Пол | Глубина, м | Стадия половозрелости | Степень наполнения желудка |
|---|---|---|---|---|---|---|
| Scomber colias | 27.9 | 268 | m | 50 | 2 | 3 |
| 28.2 | 290 | m | 50 | 2 | 3 | |
| 29.8 | 280 | f | 50 | 2 | 2 | |
| 26.8 | 222 | f | 100 | 2 | 1 | |
| Sarda sarda | 44.0 | 1190 | f | 50 | 2 | 0 |
| 44.5 | 1262 | f | 50 | 2 | 1 | |
| 47.3 | 1578 | f | 50 | 2 | 1 | |
| Beryx splendens | 21.8 | 240 | f | 50 | 4 | 4 |
| 22.3 | 224 | m | 50 | 3 | 4 | |
| 22.9 | 270 | m | 50 | 2 | 0 |
Состав эритроцитов и лейкоцитов определяли в мазках периферической крови, окрашенных по Романовскому–Гимза (Иванов и др., 2013). В каждом мазке определяли содержание основных типов клеток подсчетом по методу Горяева под цифровым микроскопом Optika DM-15 (Польша) с увеличением 10 × 60, далее производили расчет относительного их количества от суммы всех клеток в эротропоэтическом ряду и в лейкоцитарной формуле. Результаты исследований обработаны статистически с использованием t-теста при уровне достоверности p ≤ 0.05 стандартным пакетом программ (Statistica v.12).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Места обитания отмеченных видов рыб характеризуются различными экологическими условиями, в которых у рыб наблюдаются морфологические особенности и своеобразное пищевое поведение. Во время научного мониторинга в содержимом пищеварительного тракта скумбрии обнаружены мелкие пелагические виды рыб (ставриды, миктофиды), беспозвоночные (кальмары, креветки, копеподы, эвфаузииды, оболочники) и неидентифицированные переваренные остатки пищи. В исследованиях по питанию скумбрии дополнительно отмечены рыбы семейств анчоусовые, сельдевые, веретенниковые, горбылёвые, некоторые беспозвоночные (мизиды, гиперииды, полихеты), фитопланктон и водоросли (Maigret, Ly 1986; Castro, 1993; Гущин, Кортен, 2017). Доля рыбы в составе пищи скумбрии в зависимости от сезона и района может составлять от 29 до 80%. По анализу полученных данных спектр питания пеламиды состоит исключительно из рыб (сардинелла, ставрида и скумбрия). Но в других исследованиях в содержимом пищеварительного тракта регистрируются беспозвоночные (моллюски, копеподы и оболочники), в различных районах обитания и сезонах в составе пищи пеламиды встречаются анчоус, смарида, алоза, сардина, менола, каранкс (Collette, 2003; Campo et al., 2006; Гущин, Кортен, 2016).
Состав пищи берикса в акватории Марокко и Мавритании представлен миктофидами и беспозвоночными (копеподы, эвфаузииды). Также в Центрально-Восточной Атлантике исследователи питания берикса фиксируют в их пищеварительном тракте рыб семейств гоностомовые и стомиевые, а также беспозвоночных (креветки, полихеты, кальмары) (Dudockin, Kotlyar 1989; Козлов, 2004; Heemstra, Heemstra 2004). В других регионах пища берикса не отличается и состоит преимущественно из ракообразных и мезопелагических рыб. Доля рыбы в составе пищи берикса в зависимости от сезона колеблется от 11 до 58% (Lehodey, Grandperrin, 1996; Durr, Gonzalez, 2002; Horn et al., 2010; Kells, Carpenter, 2011). Особенности питания видов и их среды обитания отражаются на составе эритроцитов и лейкоцитов рыб, а соотношение отдельных типов клеток может характеризовать физиологическое состояние.
Морфология и размеры клеток эритропоэтического ряда у трех рассматриваемых видов (рис. 1) аналогична другим костистым видам рыб (Иванова, 1983; Головина, Тромбицкий, 1989). Среди типов клеток были отмечены гемоцитобласты и эритробласты (за исключением берикса), нормобласты, базофильные эритроциты и зрелые полихроматофильные эритроциты (табл. 2). На долю созревающих клеток изучаемых рыб приходилось 7.4–14.0%. Известно, что некоторое количество незрелых клеток циркулирует в периферической крови, так как у рыб отсутствует красный костный мозг и эритропоэз у них происходит в нескольких органах и тканях и завершается в сосудистой системе (Иванова, 1983; Головина, Тромбицкий, 1989).
Рис. 1.
Клетки крови скумбрии Scomber colias (а), пеламиды Sarda sarda (б) и берикса Beryx splendens (в). Масштаб: 50 мкм. Обозначения: 1 – эритробласты; 2 – нормобласты; 3 – базофильные эритроциты; 4 – зрелые и полихроматофильные эритроциты; 5 – миелоциты; 6 – метамиелоциты; 7 – палочкоядерные нейтрофилы; 8 – эозинофилы; 9 – лимфоциты.
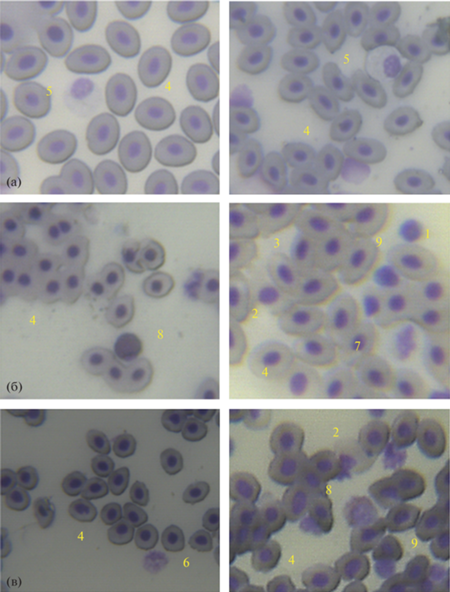
Таблица 2.
Параметры периферической крови трех видов рыб семейств Scombridae и Berycidae
| Показатели | Cкумбрия (Scomber colias) | Пеламида (Sarda sarda) | Берикс (Beryx splendens) |
|---|---|---|---|
| Эритропоэз, % | |||
| Гемоцитобласты, эритробласты | 0.1 ± 0.03 | 0.3 ± 0.13 | – |
| Нормобласты | 1.5 ± 0.50СБ | 2.0 ± 0.58 | 4.0 ± 1.00СБ |
| Базофильные эритроциты | 5.8 ± 2.14 | 9.7 ± 1.20 | 10.0 ± 1.53 |
| Сумма зрелых и полихроматофильных эритроцитов | 92.6 ± 2.48СБ | 88.0 ± 1.15 | 86.0 ± 1.15СБ |
| Лейкоцитарная формула, % | |||
| Миелоциты | 1.8 ± 0.63 | 1.0 ± 0.50 | 0.7 ± 0.67 |
| Метамиелоциты | 0.3 ± 0.25СП | 1.3 ± 0.33СП | 0.7 ± 0.33 |
| Палочкоядерные нейтрофилы | 1.0 ± 0.50 | 2.7 ± 1.20 | 1.7 ± 0.88 |
| Сегментоядерные | 1.0 ± 0.58 | 4.7 ± 2.33 | 1.7 ± 1.20 |
| Эозинофилы | 1.0 ± 0.50 | 0.3 ± 0.13 | 0.7 ± 0.33 |
| Базофилы | 0.5 ± 0.29 | 0.3 ± 0.13 | 1.3 ± 0.88 |
| Моноциты | 1.0 ± 0.41СП | 2.3 ± 0.33СП | 1.3 ± 0.67 |
| Лимфоциты | 93.5 ± 2.22 | 87.3 ± 3.71 | 92.0 ± 1.15 |
У берикса примерно в два раза выше процент нормобластов чем у скумбрии и пеламиды за счет снижения зрелых эритроцитов, которые были самой многочисленной группой клеток у рыб. Тем не менее, у берикса в отличие от других изучаемых рыб, отсутствует в крови эритро- и гемоцитобласты.
По результатам исследований состава клеток эритроидного ряда следует отметить, что эритропоэз у трех исследованных видов семейств скумбриевые и бериксовые различался (табл. 2). Факт большего количества незрелых клеток у берикса, вероятно, связан с возможными пищевыми вертикальными миграциями на большие глубины с высоким давлением и пониженным содержанием кислорода, поэтому у берикса эритропоэз происходит более интенсивно. Высокое содержание незрелых клеток эритропоидного ряда в отличие от скумбрии наблюдается у пеламиды, которая не испытывает больших перегрузок с высоким давлением, но являясь активным хищником в эпипелагической зоне может создавать риск пониженного содержания кислорода в тканях.
Предположительно, различие в содержании зрелых эритроцитов и пониженное их количество в составе эритропоэза берикса относительно скумбрии, может быть объяснено более высоким содержанием гемоглобина в крови берикса чем у скумбрии. При этом, содержание гемоглобина в крови у берикса (Аминева, Яржобек, 1984; Graham, Dickson, 2004) также выше чем у пеламиды, хотя сумма зрелых эритроцитов не отличается статистически между бериксом и пеламидой.
Рассматриваемые виды рыб имеют аналогичную другим видам рыб морфологию и размеры лейкоцитов: лимфоциты (Ø 5.0 мкм), моноциты, нейтрофилы и эозинофилы (Ø 11.0–12.0 мкм), бластные клетки (Ø 10.0 мкм). Гранулоциты этих видов, как и у большинства представителей отряда Perciformes, представлены четырьмя типами: базофилы, эозинофилы, палочкоядерные и сегментоядерные нейтрофилы (Иванова, 1983; Головина, Тромбицкий, 1989; Балабанова, 2002). При изучении лейкограмм морских костистых рыб из отрядов Clupeiformes, Beloniformes, Gadiformes, Perciformes, Pleuronectiformes и семейств Serranidae, Labridae и Myctophidae были обнаружены следующие типы клеток: гранульные микрофаги (нейтрофилы, эозинофилы, базофилы) и их незрелые формы, лимфоциты, моноциты (Точилина, 1994, Гордеев и др., 2014).
Результаты изучения лейкоцитов периферической крови скумбрии, пеламиды и берикса свидетельствуют, что их морфофункциональные характеристики гетерогенны и представлены разными по структуре клетками: лимфоциты, сегментоядерные и палочкоядерные нейтрофилы, базофилы, эозинофилы, моноциты, метамиелоциты и миелоциты. Основную долю в лейкограмме составляют лимфоциты.
У пеламиды отмечена высокая доля метамиелоцитов в лейкограмме по сравнению со скумбрией и моноцитов по сравнению с обоими изучаемыми видами рыб. Метамиелоциты являются стадией гранулоцитов (микрофагов), участвующих в реакциях фагоцитоза (Галактионов, 2005). Моноциты обладают большой миграционной способностью и активно поглощают не только бактерии, но и продукты распада клеток и тканей (Житнева и др., 1989; Головина, Тромбицкий, 1989). Следовательно, факт повышенного содержания этих клеток (метамиелоцитов и моноцитов) у пеламиды свидетельствует о большем потенциале врожденного иммунитета.
Повышенное содержание базафилов у берикса относительно пеламиды и скумбрии, может быть связано с физиологическим состоянием исследуемых особей. У особей берикса, взятых на клинические исследования, наблюдается 4-я и 3-я стадии половозрелости относительно 2-ой для остальных особей. Исследовались здоровые особи, о чем может свидетельствовать факт, что показатель не выходил за пределы референтных значений для рыб. Доля базофилов в норме у млекопитающих и большинства костистых рыб составляет от 0 до 3% (Иванов и др., 2013; Пронина, Корягина, 2015).
Исследованные показатели крови позволили отметить экологичекие особенности обитания трех видов рыб семейств скумбриевые Scombridae и бериксовые Berycidae. Так, высокий уровень эритропоэзa судя по значительному количеству незрелых клеток эритроидного ряда у берикса, вероятно, связан с его пищевыми вертикальными миграциями на большие глубины с высоким давлением и пониженным содержанием кислорода. У пеламиды отмечается более высокое содержание моноцитов, чем у скумбрии и берикса, что свидетельствует о высоком уровне врожденного клеточного иммунитета, представленного фагоцитозом.
Список литературы
Аминева В.А., Яржобек А.А. Физиология рыб. М.: Легкая и пищевая пром. 1984. 200 с.
Балабанова Л.В. Ультраструктура гранулоцитов некоторых видов окунеобразных рыб // Биология внутр. вод. 2002. № 1. С. 79–84.
Галактионов В.Г. Эволюционная иммунология: учебное пособие. М.: Академкнига, 2005. 408 с.
Головина Н.А., Тромбицкий И.Д. Гематология прудовых рыб. Кишинев: Штиинца, 1989. 156 с.
Гордеев И. И., Микряков Д.В., Балабанова Л. В., Микряков В.Р. Состав лейкоцитов периферической крови антарктического клыкача Dissostichus mawsoni (Nototheniidea) // Вопр. ихтиол. 2014. Т. 54. № 3. С. 1–4. https://doi.org/10.7868/S0042875214030047
Гущин А.В., Кортен А. Питание пелагических рыб в водах Мавритании. 2. Представители семейств Carangidae, Scombridae, Pomatodidae, Trichiuridae // Вопр. ихтиол. 2016. Т. 56. № 1. С. 68–75. https://doi.org/10.7868/S0042875216010057
Гущин А.В., Кортен А. Питание пелагических рыб вод Мавритании. 3. Восточная скумбрия Scomber colias, европейская ставрида Trachurus trachurus, ставрида-треке Trachurus trecae // Вопр. ихтиол. 2017. Т. 57. № 3. С. 308–321. https://doi.org/10.7868/S0042875217030079
Житенева Л.Д., Полтавцева Т.Г., Рудницкая О.А. Атлас нормальных и патологически измененных клеток крови рыб. Ростов н/Д.: Ростов. книж. изд-во, 1989. 111 с.
Иванов А.А., Пронина Г.И., Корягина Н.Ю, Петрушин А.Б. Клиническая лабораторная диагностика в аквакультуре. Методические указания. М.: Изд-во ТСХА, 2013. 50с.
Иванова Н.Т. Атлас клеток крови рыб. М.: Лег. и пищ. пром-сть, 1983. 184 с.
Изучение экосистем рыбохозяйственных водоемов, сбор и обработка данных о водных биологических ресурсах, техника и технология их добычи и переработки. Госкомрыболовство России. Москва. ВНИРО. 2004. 299 с.
Камышников В.В. Справочник по клинико-биохимическим исследованиям лабораторной диагностике. М.: МЕДПресс-информ, 2004. С. 56–60.
Козлов Д.А. Биология и промысел низкотелого берикса (Beryx splenens Lowe, 1833) на подводных возвышенностях северной части Атлантики. Автореф. дис. … канд. биол. наук. Калининград, 2004. 23 с.
Микряков В.Р., Балабанова Л.В. Клеточные основы иммунитета у рыб // Физиология и паразитология пресноводных животных. Л.: Наука, 1979. С. 57–64.
Микряков В.Р., Балабанова Л.В., Заботкина Е.А., Лапирова Т.Б., Попов А.В., Силкина Н.И. Реакция иммунной системы рыб на загрязнение воды токсикантами и закисление среды // М.: Наука, 2001. 126 с.
Парин Н.В. Рыбы открытого океана. М.: Наука, 1988. 272 с.
Пронина Г.И., Корягина Н.Ю. Референтные значения физиолого-иммунологических показателей гидробионтов разных видов // Вестн. Астрахан. гос. тех. ун-та. № 4. 2015. С. 103–108.
Точилина Л.В. Лейкоцитарная формула морских рыб // Гидробиол. журн. Т. 30. № 3. 1994. С. 50–57.
Campo D., Mostarda E., Castriota L., Scarabello M.P., Andaloro F. Feeding habits of the Atlantic bonito, Sarda sarda (Bloch, 1793) in the southern Tyrrhenian sea // Fisheries Research. 2006. V. 81. P. 169–175. https://doi.org/10.1016/j.fishres.2006.07.006
Castro J.J. Feedting ecology of chub mackerel Scomber japonicas in the Canary Islands area // S. Afr. J. mar. Sci. 1993. V. 13. P. 323–328. https://doi.org/10.2989/025776193784287400
Collette B.B. Family Scombridae Rafinesque 1815 – mackerels, tunas, and bonitos // Calif. Acad. Sci. 2003. V. 19. 28 p.
Collette B.B. Scombridae (including Thunnidae, Scomberomoridae, Gasterochismatidae and Sardidae). Fishes of the north-eastern Atlantic and the Mediterranean. Paris: UNESCO, 1986. V. 2. P. 981–997.
Collette B.B., Nauen C.E. Scombrids of the world. An annotated and illustrated catalogue of tunas, mackerels, bonitos and related species known to date // FAO Species Catalogue. Rome: FAO, 1983. V. 2. 137 p.
Dudochkin A.S., Kotlyar A.N. On the feeding of alfoncino (Beryx splendens) // J. Ichthyol. 1989. V. 29(5) P. 1–8.
Durr J., Gonzalez J.A. Feeding habits of Beryx splendens and Beryx decadactylus (Berycidae) off the Canary Islands // Fisheries Res. 2002. V. 54. P. 363–374. https://doi.org/10.1016/S0165-7836(01)00269-7
Graham J.B.; Dickson K.A. Tuna Comparative Physiology // The J. Experimental Biology. 2004. Vol. 207 (23). P. 4015–4024. https://doi.org/10.1242/jeb.01267
Heemstra P., E. Heemstra Coastal Fishes of Southern Africa // National Inquiry Services Centre. Eastern Cape, 2004. 488 p.
Heemstra P. Berycidae // Smiths’ Sea Fishes. Berlin: Springer-Verlag, 1986. p. 409–410.
Horn P.L., Forman J., Dunn M.R. Feeding habits of alfonsino Beryx splendens // J. Fish Biology. 2010. Vol. 76. P. 2382–2400. https://doi.org/10.1111/j.1095-8649.2010.02630.x
Kells V., Carpenter K.E. A Field Guide to Coastal Fishes. Baltimore: John Hopkins University Press, 2011. 448 p.
Lehodey P., Grandperrin R. Age and growth of the alfonsino Beryx splendens over the seamounts off New Caledonia // Marine Biol. 1996. V. 125 P. 249–258. https://doi.org/10.1007/BF00346305
Maigret J., Ly B. Les poissons de mer de Mauritanie // Compiègne: Science Nat., 1986. 213 p.
Maul G.E. Berycidae // Check-list of the fishes of the eastern tropical Atlantic (CLOFETA). Paris: UNESCO, 1990. V. 2. P. 626.
Moffitt R.B., Kobayashi D.R., Dinardo G.T. Status of the Hawaiian Bottomfish Stocks // Honolulu: Pacific Islands Fisheries Science Center, 2004. Report H-06-01. 45 p.
Nielsen J. Berycidae // Check-list of the fishes of the north-eastern Atlantic and of the Mediterranean (CLOFNAM). Paris: UNESCO, 1979. V. 1. P. 337.
Parin N.V., Shcherbachev Y.N., Pakhorukov N.P. Bottom and near-bottom fishes of the Rio Grande Rise (Southwest Atlantic Ocean) // J. Ichthyol. 1995. V. 35(9). P. 205–219.
Parish N., Wrathmell A., Hart S., Harris J. The leucocytes of the elasmobranch Scyliorhinus vanicula L. A morphological study // J. Fish. Biol. 1986. V. 28 (5). P. 545–561. https://doi.org/10.1111/j.1095-8649.1986.tb05192.x
Paxton J.R. Berycidae. Alfonsinos // FAO species identification guide for fishery purposes. The living marine resources of the WCP. Rome: FAO, 1999. V. 4. P. 2218–2220.
Ramos A., Moya F., Salmerón F., García P., Carroceda A., Fernández L., González J.F., Tello O., Sánz J. L., Ballesteros M. Demersal Fauna on Deep Seamounts of Sierra Leone Rise (Gulf of Guinea, Africa) // NAFO. 2001. CR Doc. 01/149. 4 p.
Santamaría M.T.G., López-Abellán L.J., Gonzalez J.F. Growth of alfonsino Beryx splendens Lowe 1834 in the South-West IndianOcean // African J. of Marine Science. 2006. V. 28 (1). P. 33–40. https://doi.org/10.2989/18142320609504131
Secombes C.J. The nonspecific immune system: cellular defense // The fish immune system: organism, pathogen and environment. London: Acad. Press., 1996. P. 63–105.
Shimizu T. Berycidae // FAO species identification sheets for fishery purposes. Western Indian Ocean fishing area 51. 1984. V. 1. 6 p.
Sommer C., Schneider W., Poutiers J.M. FAO species identification field guide for fishery purposes // The living marine resources of Somalia. Rome: FAO, 1996. 376 p.
Torres M.A., Ramos F. Sobrino I. Length-weight relationships of 76 fish species from the Gulf of Cadiz (SW Spain) // Fish. Res. 2012. V. 127. P. 171–175. https://doi.org/10.1016/j.fishres.2012.02.001
Zorica B., Sinovcic G. Biometry, length-length and length-weight relationships of juveniles and adults of Atlantic bonito, Sarda sarda, in the eastern Middle Adriatic Sea // Acta Adriatica. 2008. V. 49(1). P. 65–72.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая