

Известия РАН. Серия биологическая, 2023, № 6, стр. 659-668
Суточная активность желтого суслика Spermophilus fulvus Licht. (Sciuridae): первый опыт инструментального исследования
Н. А. Васильева 1, *, А. А. Лупырёв 1, 2, Н. С. Васильев 1, 2
1 Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский просп., 33, Россия
2 Московский государственный университет им. М.В. Ломоносова, биологический ф-т
119234 Москва, Ленинские горы, 1, Россия
* E-mail: ninavasilieva@gmail.com
Поступила в редакцию 20.09.2022
После доработки 09.11.2022
Принята к публикации 09.11.2022
- EDN: UBZLPS
- DOI: 10.31857/S1026347022600777
Аннотация
Большая часть информации о суточной активности животных основана на лабораторных данных. Для получения сведений из природной популяции мы впервые провели инструментальное исследование суточного цикла активности желтого суслика – крупного зимоспящего грызуна. Восемь особей были оснащены радиопередатчиками с акселерометрами и датчиками освещенности летом 2021 г. Полученные данные подтвердили, что желтые суслики покидают норы только днем; уход суслика в нору соответствовал переходу в состоянии покоя. По мере приближения залегания в спячку время пребывания на поверхности сокращалось. Наземная активность сусликов была однофазной, в отличие от близких видов с двумя пиками активности. Возможно, желтые суслики более интенсивно используют дневные часы из-за дефицита времени, который является следствием длительной спячки .
Особенности суточной активности у животных входят в число ключевых характеристик биологии вида. Они тесно связаны с экологическими свойствами вида: способами поиска пищи, использованием пространства, характеристиками местообитания и др. При этом, суточным циклам подвержены ключевые физиологические процессы в организме животного: цикл сна и бодрствования, активность сердечно-сосудистой, эндокринной, пищеварительной систем, температуры тела и др. (Froy, 2007; Refinetti, 2019). Известно, что млекопитающие демонстрируют широкое разнообразие паттернов суточной активности, в том числе, различные 24-часовые ритмы (дневной, ночной, круглосуточный), а также краткосрочный ритм активности; для многих видов показано, что синхронизация этих ритмов осуществляется в соответствии с интенсивностью освещения (Ashby, 1972). При этом, изменчивость паттернов суточной активности внутри вида может быть очень значительной и определяться полом (Schmidt, 1999; Everts et al., 2004; Vieira et al., 2010), физическим состоянием, стадией репродуктивного цикла (Clutton-Brock et al., 1982; Kenagy, Hoyt, 1989), внешними условиями (Рашитов, 2018; Podolski et al., 2013; Allen et al., 2018) и др.
Традиционно, изучение режима суточной активности животных ограничивалось лабораторными исследованиями, проводимыми, зачастую, при неестественном режиме освещения. Инструментальные методы регистрации активности до последнего времени применялись лишь на небольшом числе видов и в условиях неволи (напр., Daan, Aschoff, 1975; Bartness, Albers, 2000; Refinetti, 2019). В дикой природе информация об изменениях в активности животных в течение суток ограничивалась данными, полученными в ходе визуальных наблюдений, которые особенно ненадежны в случае мелких, скрытных и осторожных видов животных. Такое положение дел препятствовало получению релевантных данных от широкого спектра видов и проведению сравнительного анализа. В последние годы стремительное развитие технологий позволило перевести исследования ритмов активности животных на новый уровень: представителей разных видов начали оснащать разнообразными датчиками, в том числе, в дикой природе. Появление мощной вычислительной аппаратуры позволило разработать подходы к анализу больших массивов информации, получаемых с таких регистраторов. Тем не менее, инструментальные исследования в природе до сих пор крайне немногочисленны.
Зимоспящие млекопитающие, которые значительную часть года проводят в состоянии глубокого угнетения жизненных процессов, представляют особый интерес с точки зрения исследования суточных ритмов. Он связан с тем, что механизмы регуляции процессов, связанных со спячкой (в том числе, циклов пробуждения во время спячки) могут иметь общую природу с механизмами регуляции суточных циклов активности (Heller, Ruby, 2004; Malan, 2010). Наземные беличьи (суслики, сурки) – группа норных зимоспящих грызунов, которые ведут дневной образ жизни (Шилова, 2000; Dobson, 1984; Armitage, 1986; Waterman, 2007). Они являются традиционной модельной группой в поведенческий экологии, поскольку широкое разнообразие социальных систем и репродуктивных стратегий у этих грызунов сочетается с сравнительно малыми морфологическими, физиологическими и экологическими различиями между видами. Это дает возможность на модели этой группы видов исследовать эволюцию формирования самых разных признаков у млекопитающих (Шилова, 2000; Шилова, 2004, Васильева, Чабовский, 2017).
Норный образ жизни разделяет активность наземных беличьих на две составляющие: пребывание в норе и пребывание на поверхности земли, что определяет специфику исследований активности этих животных. С одной стороны, возникает возможность упрощенного подхода к исследованию активности: животное, находящееся на поверхности земли, автоматически можно считать активным, а под землей – неактивным (см. напр., Bronson, 1962; Hut et al., 1999; Long et al., 2005). С другой стороны, возникают очевидные методические трудности для получения информации о животном из норы, в особенности в условиях дикой природы. На сегодня инструментальные исследования паттернов суточной активности были проведены лишь на единичных видах наземных беличьих, в первую очередь, на европейском (Spermophilus citellus) (Hut et al., 1999) и арктическом (Urocitellus parryii) (Long et al., 2005; 2007; Chmura et al., 2020) сусликах, для изучения соотношения наземной и подземной активности которых были использованы светочувствительные датчики.
Желтый суслик (Spermophilus fulvus Licht., 1823) – самый крупный из сусликов, самцы перед спячкой могут достигать массы 2 кг (Млекопитающие…, 1969; Васильева и др., 2009). Этот вид населяет сухие степи, полупустыни, пустыни и характеризуется экстремально длительной зимней спячкой – до 9 мес. (Шилова, 2000; Васильева, Чабовский, 2017). При столь коротком периоде активной наземной жизни можно ожидать присутствия специфических черт режима суточной активности у этого вида. Однако до сих пор информация об этом виде крайне ограничена, имеются лишь сведения, полученные в ходе визуальных наблюдений, свидетельствующие о том, что это строго дневные животные (Орлов, 1925; Материалы…, 1929; Огнев, 1947). Некоторые авторы утверждают, что наземная активность (т.е., пребывание вне норы в течение дня) у желтого суслика двухфазная: суслики пребывают на поверхности утром и вечером, в то время как в середине дня они не покидают нор (Орлов, 1925; Огнев, 1947; Млекопитающие…, 1969; Россолимо и др., 2004).
В данной работе мы впервые предприняли инструментальное исследование суточной активности желтого суслика в дикой природе с использованием светочувствительных датчиков и акселерометров, целью которого было получить сведения о суточной динамике пребывания на поверхности земли и локомоторной активности сусликов в период подготовки к спячке.
МАТЕРИАЛЫ И МЕТОДЫ
Объект и место исследования. Мы проводили сбор данных в период подготовки сусликов к спячке в июне–июле 2021 г. в рамках долговременного исследования природного поселения желтого суслика (Spermophilus fulvus orlovii Ogn. 1937) в окрестностях с. Дьяковка Краснокутского района Саратовской области (50°43′88′′ N, 46°46′04′′ E). В данном поселении индивидуальное мечение сусликов проводится с 2001 года. Как следствие, возраст, происхождение, основные события годового цикла известны практически для всех особей.
Сусликов отлавливали с помощью сетчатых ловушек оригинальной конструкции и петель с радиопередатчиками, которые при срабатывании мгновенно передавали предупреждающий сигнал по радиоканалу. При отлове сусликов метили с помощью подкожных ветеринарных микрочипов и с помощью черного красителя (урзола) для визуального распознавания. Детальное описание отлова и мечения желтых сусликов можно найти в ранее опубликованных работах (Васильева и др., 2009; Vasilieva et al., 2014; Vasilieva, Tchabovsky, 2014, 2015).
Разработка и использование радиопередатчиков для желтого суслика. Желтый суслик – одиночный вид, который и сегодня является объектом охоты (Шилова и др., 2015). Возможно, это отчасти определяет чрезвычайную осторожность этих животных, которую еще в начале ХХ века особо отмечали исследователи (Материалы…, 1929). Отлов желтых сусликов крайне трудоемок: для некоторых особей установка практически невидимой петли из тонкого тросика с датчиком прикосновения служит поводом не выходить из норы на протяжении суток. Как следствие, большинство сусликов удается отловить не более двух раз за весь сезон активности (т.е., за год). При этом желтым сусликам свойственны резкие изменения массы тела на протяжении годового цикла: с момента выхода из спячки и до залегания зверьки увеличивают массу тела в 1.5–2 раза, а детеныши после первого выхода из нор и до момента залегания в спячку увеличивают массу в 4–5 раз и более (Васильева и др., 2009). Таким образом, использование ошейников в качестве физической основы для каких-либо устройств слежения и регистраторов для этого вида недопустимо: надетый на зверька ошейник может привести к удушью при быстром увеличении массы тела, а вовремя отловить животное и снять устройство может оказаться невозможным. Кроме того, ошейник в условиях колючей степной растительности может привести к нарушению целостности кожи, воспалению и раздражению при попадании колючих семян под ошейник (Williams et al., 2014). Чтобы избежать этих неблагоприятных сценариев, мы разработали безопасный способ крепления устройств слежения для желтого суслика. В ходе этой работы были впервые использованы радиопередатчики оригинальной конструкции, специально разработанные для желтого суслика в лаборатории популяционной экологии Института проблем экологии и эволюции им. А.Н. Северцова. Малогабаритные передатчики массой 12 г (0.8–2.5% от массы животного), были упакованы в термоусадочную пленку и закреплены на коже спины сусликов с помощью медицинского клея Perma-Type Surgical Cement (Perma Type Company, USA). После линьки радиопередатчики самопроизвольно отделялись от кожи зверьков, при этом ни у одной особи на месте крепления передатчиков на коже не было раздражения или повреждений. У перелинявших особей не было визуальных признаков того, что ранее на них был закреплен передатчик.
Радиопередатчики были снабжены датчиками освещенности, которые каждые 10 мин регистрировали, где находится зверек – в норе или на поверхности, также они записывали точное время входа и входа из норы. Кроме того, радиопередатчики были оснащены акселерометрами, которые каждые 10 мин регистрировали долю времени из предыдущих 10 мин, когда животное находилось в движении. Эти показания складывались из пятисекундных срезов: каждые 5 с передатчик сохранял “1”, если животное двигалось (движение регистрировалось при ускорении ≥0.19 g хотя бы по одной из трех осей акселерометра), и “0”, если оно было неподвижно (ускорение составляло <0.19 g), и “единицы” суммировались за каждые 10 мин.
Данные с радиопередатчиков передавались по радиоканалу на два радиоприемника (использовались диапазоны гражданской радиосвязи 434 МГц), расположенных на деревьях. За 2021 г. с помощью радиопередатчиков были получены данные (40 310 регистраций) от 4 самцов (1 взрослого и 3 детенышей) и 4 самок (1 взрослой и 3 детенышей) на протяжении периода с 21 июня по 30 июля.
Все манипуляции с животными прошли экспертизу и были одобрены Комиссией по биоэтике ИПЭЭ РАН (протоколы № 31, 44), используемые методы соответствовали рекомендациям “Руководства по обращению с животными при проведении поведенческих исследований и обучении” (ASAB/ABS, Guidelines for the treatment of animals in behavioural research and teaching (Buchanan et al., 2012)).
Анализ данных. Для оценки изменений в активности сусликов по мере приближения спячки мы использовали следующие переменные, полученные для каждой особи для каждого дня наблюдений:
1) время начала дневной активности (время первого выхода из норы утром);
2) продолжительность дневной активности: разность во времени от первого выхода из норы утром до последней регистрации особи на поверхности;
3) точное время пребывания на поверхности в течение дня: рассчитывалось как продолжительность дневной активности за вычетом суммарного времени, проведенного в норе в течение дня (от первого выхода из норы утром до последней регистрации на поверхности);
4) время конца дневной активности (время последней регистрации на поверхности вечером).
Для анализа суточной динамики наземной активности и подвижности мы использовали две переменные:
1) Пребывание в норе/на поверхности на протяжении суток: последовательность нулей и единиц, полученных с частотой раз в 10 мин на основе показаний датчика освещенности. Единицы соответствовали нахождению на поверхности, а нули – пребыванию в норе. Дополнительными единицами отмечались моменты каждого выхода из норы, поэтому фактически регистрации были несколько чаще, чем раз в 10 мин.
2) Уровень локомоторной активности на протяжении суток: для тех же моментов времени, что и показания наземной активности, отмечалась доля времени в движении от времени, прошедшего с момента предыдущей регистрации; эта доля варьировала от 0 (полная неподвижность) до 1 (непрерывное движение).
Дни поимки зверьков, дни залегания в спячку, а также дни гибели от нападения хищников (один молодой самец был убит орлом-карликом, другой – собакой) были исключены из анализа.
Анализ данных был проведен в среде R 4.1.0 (R Development Core Team, 2021). В первую очередь, мы описали общий паттерн дневной активности зверьков и проанализировали, как время начала, конца дневной активности и продолжительность дневной активности зверьков менялись в зависимости от календарной даты. Также мы проверили, насколько при пребывании в норе суслики находятся в состоянии покоя по сравнению с пребыванием на поверхности, чтобы оценить правомерность подхода, при котором животное на поверхности земли считается активным, а животное в норе – неактивным (Hut et al., 1999; Long et al., 2005). Для этого мы исследовали, как меняется уровень локомоторной активности сусликов при переходе между норой и поверхностью земли. Чтобы разделить эффекты суточного ритма и ухода в нору как такового, мы построили две модели: одну – для всех измерений в течение суток, и другую – только для измерений с момента первого выхода каждого зверька утром из норы до момента последнего ухода вечером в нору. Для решения всех этих задач мы построили линейные модели смешанных эффектов в пакете nlme (Pinheiro et al., 2022), при этом номер особи был включен во все модели как случайный фактор.
Чтобы визуализировать суточную динамику наземной активности зверьков (пребывания в норе/на поверхности земли) мы использовали пакет activity (Rowcliffe, Rowcliffe, 2022) в среде R. Для построения кривых были получены частотные распределения регистраций каждого зверька на поверхности земли (при этом, данные о том, в норе или на поверхности находилось животное, поступали с постоянной частотой на протяжении суток). Кроме того, мы использовали пакет overlap (Ridout, Linkie, 2009; Meredith et al., 2021) и тест Вальда (Wald test) из пакета activity для того, чтобы количественно сравнить суточную динамику наземной активности самцов и самок (это сравнение мы проводили только для молодых особей), а также взрослых и молодых особей. Чтобы визуализировать изменения в режиме наземной активности по мере приближения залегания в спячку, мы для тех особей, которые были прослежены до спячки (N = 4), сравнили суточную динамику активности в последние 7 дней перед спячкой (т.е., непосредственно перед залеганием) и в период 8–14 дней до залегания. Результаты статистических тестов мы считали достоверными при значении p < 0.05.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
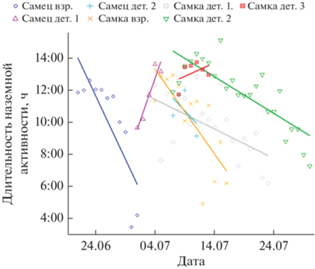
Общая характеристика суточной активности. Все особи желтого суслика были активны строго в дневное время, ни один суслик не покидал нору в темное время суток. В среднем, суслики выходили из нор в 6:16 ± 1:49 утра и уходили в норы в 17:07 ± 1:22 в то время, как восход солнца в эти дни был до 4:00, а заход около 20:00 (время везде указано по Московскому времени). Таким образом, в среднем продолжительность дневной наземной активности сусликов составила 10:51 ± 2:22 ч, из которых на поверхности зверьки проводили в среднем около 67 ± 13% времени, а остальную часть дня они находились в норах. Бюджет наземной активности сусликов зависел от календарной даты: чем больше была дата, тем позже суслики выходили из нор утром (модель смешанных эффектов, номер особи добавлен в данную и последующие модели как случайный фактор; B = 0.50 ± 0.14, t = 3.5, p = = 0.0007) и тем раньше они уходили в норы вечером (B = –0.71 ± 0.14, t = –5.1, p < 0.0001). Таким образом, продолжительность дневной наземной активности сокращалась по мере приближения спячки (B = –0.87 ± 0.13, t = –6.7, p < 0.0001, рис. 1). При этом точное время пребывания на поверхности земли (за вычетом времени, которое зверек проводил в норе в течение дня) сокращалось даже несколько быстрее, чем продолжительность наземной активности (B = –0.83 ± 0.09, t = –9.0, p < 0.0001).
Рис. 1.
Изменение продолжительности наземной активности (в часах) в зависимости от календарной даты у желтого суслика в период подготовки к спячке. Разными цветами обозначены разные особи. Длительность наземной активности достоверно снижалась (линейные модели смешанных эффектов, p < 0.05).
Соответствие бюджета наземной и локомоторной активности. Уровень локомоторной активности сильно зависел от того, где находился суслик – в норе или на поверхности: двигательная активность на поверхности была многократно выше, чем в норе (B = 1.46 ± 0.01, t = 99.7, p < 0.0001). В среднем, из каждого 10-минутного промежутка, проведенного на поверхности, суслик двигался 5.3 ± 2.9 мин (медиана = 5.3 мин/10 мин), в то время как в норе время в движении в среднем составляло 1.2 ± 1.7 мин/ 10 мин (медиана = 0.6 мин/10 мин). При этом ночью суслики в норе были практически неподвижны (время в движении составляло 0.1 ± ± 0.2 мин/10 мин, медиана = 0.05 мин/10 мин). При уходе в нору в дневное время суслики резко снижали двигательную активность по сравнению с уровнем подвижности при нахождении на поверхности земли (B = 1.14 ± 0.02, t = 48.3, p < 0.0001).Таким образом, бюджет наземной активности в данном исследовании можно использовать в качестве оценки бюджета двигательной активности.
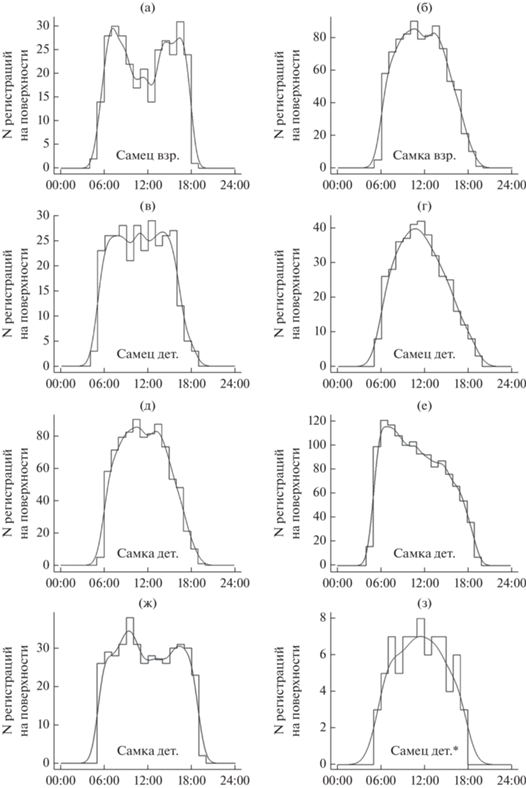
Суточная изменчивость наземной активности. С помощью пакета activity мы построили графики вероятности нахождения сусликов на поверхности земли в зависимости от времени суток (рис. 2). Во-первых, эти графики демонстрирую большую изменчивость бюджета активности между особями: у некоторых особей пик наземной активности приходился на вечер, у других – на утро, у некоторых – на середину дня. Лишь взрослый самец (рис. 2а) придерживался двухфазной наземной активности: в середине дня он находился преимущественно в норе. Остальные суслики, включая взрослую самку (рис. 2б), были активны на протяжении всего дня, и их активность можно назвать однофазной.
Рис. 2.
Суточная динамика пребывания на поверхности земли желтых сусликов в период подготовки к спячке. По оси ординат представлено число регистраций на поверхности земли на каждый час на протяжении суток. “Взр.” = взрослый зверек, “дет.” = детеныш. * График построен лишь по одному дню работы радиопередатчика (эта особь была убита собакой на третий день после поимки).
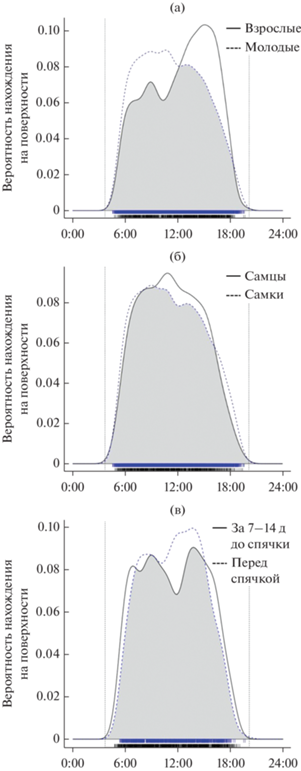
Анализ перекрывания сроков наземной активности у взрослых и молодых особей в пакете overlap показал, что молодые в течение дня активны несколько дольше, при этом они больше пребывают на поверхности утром по сравнению со взрослыми (тест Вальда, p = 0.005, рис. 3а). При сравнении бюджетов самцов и самок (только молодых особей) различий между ними мы не выявили (p = 0.2, рис. 3б). Наконец, мы сравнили бюджет наземной активности перед спячкой и за 7–14 дней до спячки: длительность наземной активности снижалась по мере приближения залегания в спячку (p = 0.047, рис. 3в), что согласуется с результатами линейных моделей.
Рис. 3.
Сравнение суточной динамики пребывания желтых сусликов на поверхности у взрослых и молодых особей (а), самцов и самок (б) а также в сроки 7–14 дней до спячки и в последнюю неделю перед спячкой (в). Различия достоверны (тест Вальда, p < 0.05) для а) и в); кривые соответствуют функциям плотности, построенным в пакете overlap; серым цветом показаны области перекрывания. В нижней части графиков черным и синим цветами отмечены точные моменты регистраций (синий цвет везде соответствует пунктирным линиям, черный – сплошным). Вертикальные пунктирные линии соответствуют среднему времени восхода и заката в сроки наблюдений.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Данная работа является первым инструментальным исследованием бюджета активности желтого суслика, в ходе которого были получены надежные данные о суточном цикле этого вида в период подготовки к спячке. Ранние работы, описывающие бюджет суточной активности желтого суслика, опирались на визуальные наблюдения за особями, которые не были помечены для индивидуального распознавания (Млекопитающие…, 1969); в некоторых работах исследователи закрывали входы в норы сусликов и судили о бюджете наземной активности зверьков по тому, когда суслики открывали их вновь (Материалы…, 1929). Эти методы могут быть недостаточно надежными для столь пугливых и осторожных животных, какими являются желтые суслики, кроме того, при таких подходах трудно оценить точное время всех заходов в норы и выходов из них. Наш подход исключает необходимость присутствия наблюдателя на поселении, ошибки наблюдателей и т.п., и дает высокую точность оценки времени входа/выхода. Разумеется, выборка в нашей работе крайне мала, и это исследование можно считать пилотным; однако мы можем хотя бы на качественном уровне сделать ряд выводов.
Наши данные подтверждают ранние наблюдения о том, что желтые суслики являются строго дневными животными. Ни одна особь не выходила из норы до восхода солнца или после заката, что согласуется с другими работами по желтому суслику, в которых упоминается, что суслики практически не бывают на поверхности до 6:00 и после 18:00 (Орлов, 1925; Материалы…, 1929; Млекопитающие…, 1969) и в целом соответствует данным по другим видам наземных беличьих (Шилова, 2000).
В других работах по наземным беличьим связь между наземной и двигательной активности подразумевается неявно, без инструментальной валидации (Hut et al., 1999; Long et al., 2005). В ходе данного исследования с помощью сочетания акселерометров и датчиков освещенности мы проверили, насколько для желтого суслика правомерен подобный подход. Мы показали, что практически вся двигательная активность сусликов на протяжении суток приходилась на то время, когда зверьки пребывали на поверхности земли. Ночью суслики в норах были практически полностью неподвижны; но и в дневное время при уходе в нору зверьки снижали двигательную активность в несколько раз, хотя можно было ожидать, что в период подготовки к спячке суслики будут интенсивно расчищать норы. Вероятно, наиболее активная роющая деятельность у желтых сусликов приходится на другую фазу годового цикла, которая не попала в период, который мы анализируем в данной работе. Таким образом можно утверждать, что в норе желтые суслики находятся в состоянии покоя, и пребывание зверька на поверхности земли может быть использовано в качестве индикатора двигательной активности. По крайней мере, эта закономерность свойственна желтым сусликам в последние недели перед уходом в спячку.
В нашем исследовании бросается в глаза высокая вариабельность паттернов активности между особями: максимум наземной активности мог приходиться на утро, вечер или день; он мог быть не один, как у взрослого самца (см. рис. 2). Эту особенность наблюдали и ранее: Е. И. Орлов (1925) писал о сусликах, обитавших в районе с. Дьяковка в 1924 г.: “у каждого из них замечаются излюбленные моменты выхода из нор”; сходным образом, у голохвостого суслика (Xerospermophilus tereticaudus) разные особи демонстрировали устойчивые различия во времени начала дневной активности (Drabek, 1973). Наших данных недостаточно для исследования индивидуальных особенностей бюджета наземной активности, но имеющиеся сведения допускают существование внутри популяции различных поведенческих стратегий распределения активности в течение суток, что требует дополнительных данных из природных популяций.
В ряде работ по желтому суслику исследователи отмечали, что в разгар лета наземная активность у этого вида, как правило, двухфазная: зверьки находятся на поверхности в утренние и вечерние часы, а в середине дня как взрослые, так и молодые зверьки уходят в норы (Орлов, 1925; Млекопитающие…, 1969). Например, на острове Барсакельмес (Казахстан) в июне перерыв в наземной активности желтых сусликов в течение дня продолжался с 10:00 до 16:00 (Млекопитающие…, 1969). Однако некоторые авторы указывают, что у желтого суслика может быть один пик наземной активности в течение суток (Орлов, 1925; Материалы…, 1929), что не вполне обычно для наземных беличьих.
Двухфазная наземная активность в середине лета, при которой днем зверьки отдыхают в норах, описана для многих других сусликов и сурков (напр., Bronson, 1962; Drabek, 1973; Шубин, 1988 ; Armitage, 1991; Katona et al., 2002), а также для других видов дневных грызунов (Bacigalupe et al., 2003). У дневных грызунов снижение наземной активности днем связывают с приспособленностью к жизни в местообитаниях с высокими дневными температурами воздуха; то есть, уход в нору в середине дня является у таких видов своего рода поведенческой терморегуляцией, защищающей зверьков от перегрева (Млекопитающие…, 1969; Loehr, Risser, 1977; Vispo, Bakken, 1993;Váczi et al., 2006). В течение срока наблюдений в нашем исследовании дневные температуры достигали +40°C градусов в тени (данные с регистратора температуры, размещенного на модельной территории). Тем не менее, в нашей выборке двухфазный паттерн бюджета активности был присущ только взрослому самцу, остальные особи, в том числе взрослая самка, демонстрировали однофазную наземную активность, в отличие от других видов наземных беличьих. Одна из причин подобного расхождения наших данных с другими работами может быть в том, что бюджет наземной активности у этого вида зависит от возраста: существуют свидетельства того, что детеныши пребывают на поверхности в течение всего дня, в то время как взрослым свойственна двухфазная наземная активность (Млекопитающие…, 1969). Согласно нашим данным, и молодые самцы, и самки дольше пребывали на поверхности земли на протяжении суток по сравнению со взрослыми зверьками. Можно предполагать, что экстремально длительная спячка задает жесткие временные рамки для детенышей, которые с момента выхода из нор в мае-июне к концу июля должны успеть в несколько раз увеличить массу тела, чтобы успешно перезимовать, а некоторые детеныши еще и успевают расселиться (Шилова и др., 2006; Васильева, 2011). В таких условиях молодые зверьки остаются на поверхности на протяжении дня максимально долго, в том числе в наиболее жаркие часы. Не вполне ожидаемым было отсутствие различий в бюджетах активности между молодыми самцами и самками, поскольку самцы у наземных беличьих являются расселяющимся полом, а самки склонны к филопатрии, и можно было предположить большую продолжительность наземной активности у самцов. Отчасти, это может объясняться недостаточным размером выборки; в то же время, вероятно, что молодые самцы и самки при одинаковых бюджетах наземной активности по-разному используют время, проведенное на поверхности, и по-разному используют пространство.
Снижение продолжительности пребывания на поверхности земли по мере приближения момента залегания в спячку не было очевидным образом связано с сокращением светового дня или с погодными условиями, поскольку эта закономерность прослеживалась у разных зверьков в разные календарные даты: как во второй половине июня, так и в конце июля. Данных в настоящем исследовании недостаточно для детального анализа этих взаимосвязей, поэтому мы можем обсуждать эти закономерности лишь на качественном уровне. Отметим, что у двух зверьков, у которых сокращения активности не было выявлено, был очень короткий период наблюдений, а в первые дни после поимки можно предполагать, наоборот, увеличение активности у этих пугливых животных. Снижение продолжительности наземной активности перед спячкой было описано, например, для колумбийского суслика Urocitellus columbianus (Betts, 1976).
Следует отметить, что не вполне понятно, как у желтого суслика и других дневных наземных беличьих возможна синхронизация бюджета активности с длиной светового дня. Такая синхронизация присуща млекопитающим в целом (Refinetti, 2019), однако наши данные и данные по близким видам сусликов свидетельствуют о том, что суслики никогда не находятся на поверхности земли в сумерках (Hut et al., 1999; Everts et al., 2004). Более того, использование датчиков освещенности позволяет утверждать, что они даже не приближаются к выходу из норы в эти периоды, чтобы определить моменты захода и восхода солнца и освещенность, при том, что такое поведение (“light sampling behaviour”) известно для некоторых видов млекопитающих (De Coursey, 1986). Известно, что у других видов наземных беличьих по мере приближения спячки в организме происходят существенные изменения в гормональном статусе, биохимических показателях крови и др. (напр., Boonstra et al., 2011; Wilsterman et al., 2015). Возможно, постепенные изменения в продолжительности наземной активности у желтых сусликов в большей степени могут быть связаны с физиологическими перестройками, обусловленными подготовкой к спячке, а не с изменениями внешних условий.
ЗАКЛЮЧЕНИЕ
Таким образом, мы впервые получили детальные данные о суточном бюджете наземной активности желтого суслика. Несмотря на ограниченный объем выборки, эти данные могут быть полезны при планировании исследований данного вида, в том числе, учетов численности, мониторинга популяций и др. Один из результатов данной работы – отсутствие выраженного снижения уровня наземной активности в середине дня – согласуется с представлениями о желтом суслике как о виде с “быстрым” жизненным циклом. Возможно, в условиях экстремально длительной зимней спячки и, соответственно, ограниченного времени на рост и набор веса, желтые суслики (в первую очередь, детеныши) максимально используют дневные часы для питания, подготовки нор к зиме и др., однако, это предположение требует дальнейших исследований в природе.
Список литературы
Васильева Н.А., Савинецкая Л.Е., Чабовский А.В. Крупный размер тела и короткий период наземной активности не препятствуют быстрому росту желтого суслика Spermophilus fulvus // Зоол. Журн. 2009. Т. 88. № 3. С. 339–343.
Васильева Н.А. Факторы, влияющие на репродуктивное поведение и успех размножения у желтого суслика (Spermophilus fulvus): Автореф. дис. … канд. биол. наук. М.: ИПЭЭ РАН, 2011. 24 с.
Васильева Н.А., Чабовский А.В. Принятие репродуктивных решений в контексте “быстрого” жизненного цикла (на примере желтого суслика Spermophilus fulvus) // Журн. Общ. Биол. 2017. Т. 78. № 1. С. 3–14.
Материалы к познанию фауны нижнего Поволжья. Вып. IV / Ред. Траут И.И., Орлов Е.И. Саратов: Изд. отдела применения Нилов. 1929. 122 с.
Млекопитающие Казахстана. Т. 1 / Ред. Слудский А.А. Алма-Ата: Наука, 1969. 455 с.
Огнев С.И. Звери СССР и прилежащих стран (звери Восточной Европы и Северной Азии). М.-Л.: АН СССР. 1947. 559 с.
Орлов Е.И. Желтый суслик. Биологические наблюдения // Вестн. Микробиол. и Эпидемиол. 1925. Т. 4. Вып. 1. С. 58–66.
Рашитов С.С. Влияние суточной активности сурков на накопление ими жира // Извест. Межд. Акад. Аграр. Образ. 2018. № 43. С. 175–179.
Россолимо О.Л., Павлинов И.Я., Крускоп С.В., Лисовский А.А., Спасская Н.Н., Борисенко А.В., Панютина А.А. Разнообразие млекопитающих. Ред. Россолимо О.Л. М.: КМК, 2004. 366 с.
Шилова С.А. Пространственная и социальная организация земляных белок (р. Spermophilus, Xerus, Cynomys) как модель эколого-этологических исследований // Усп. Совр. Биол. 2000. Т. 120. № 6. С. 559–572.
Шилова С.А. Земляные белки // Природа. 2004. № 3. С. 41–48.
Шилова С.А., Савинецкая Л.Е., Чабовский А.В. Долговременная и современная динамика популяции желтого суслика (Spermophilus fulvus, Rodentia, Sciuridae) в Приерусланских песках Заволжья // Зоол. журн. 2015. Т. 94. № 8. С. 944–954.
Шилова С.А., Чабовский А.В., Попов B.C. Особенности формирования поселений желтого суслика (Spermophilus fulvus Licht., 1823) при вселении на свободную территорию // Бюллетень МОИП. Отдел биологический. 2006. Т. 111. № 5. С. 71–75.
Шубин В.И. Особенности взаимоотношений в семейных группах байбака // Экология и поведение млекопитающих Казахстана // Тр. Ин-та зоологии АН Каз. ССР. 1988. Т. 44. С. 112–132.
Allen M.L., Peterson B., Krofel M. No respect for apex carnivores: distribution and activity patterns of honey badgers in the Serengeti // Mamm. Biol. 2018. V. 89. P. 90–94.
Armitage K.B. Individual differences in the behavior of juvenile yellow-bellied marmots // Behav. Ecol. Soc. 1986. V. 18. P. 419–424.
Armitage K.B. Social and population dynamics of yellow-bellied marmots: results from long-term research //Annu. Rev. Ecol. Syst.1991. V. 22. P. 379–407.
Ashby K.R. Patterns of daily activity in mammals // Mammal Review. 1972 V. 1. P. 171–185.
Bacigalupe L.D., Rezende E.L., Kenagy G.J., Bozinovic F. Activity and space use by degus: a trade-off between thermal conditions and food availability? // J. Mammal. 2003. V. 84. P. 311–318.
Bartness T.J., Albers H.E. Activity patterns and the biological clock in mammals // Activity patterns in small mammals / Springer, Berlin, Heidelberg. 2000. P. 23–47.
Betts B.J. Behaviour in a population of Columbian ground squirrels, Spermophilus columbianus columbianus // Anim. Behav. 1976. V. 24. P. 652–680.
Boonstra R., Bradley A.J., Delehanty B. Preparing for hibernation in ground squirrels: adrenal androgen production in summer linked to environmental severity in winter // Funct. Ecol. 2011. V. 25. P. 1348–1359.
Bronson F.H. Daily and seasonal activity patterns in woodchucks // J. Mammal. 1962. V. 43. P. 425–427.
Buchanan K., Burt de Perera T., Carere C., Carter T., Hailey A., Hubrecht R., Jennings D., Metcalfe N., Pitcher T., Peron F., Sneddon L., Sherwin C., Talling J., Thomas R., Thompson M. Guidelines for the treatment of animals in behavioural research and teaching // Anim. Behav. 2012. V. 83. P. 301–309.
Chmura H.E., Zhang V.Y., Wilbur S.M., Barnes B.M., Buck C.L., Williams C.T. Plasticity and repeatability of activity patterns in free-living Arctic ground squirrels //Anim. Behav. 2020. V. 169. P. 81–91.
Clutton-Brock T.H., Iason G.R., Albon S.D., Guinness F.E. Effects of lactation on feeding behaviour and habitat use in wild red deer hinds // J. Zool. 1982. V. 198. P. 227–236.
Daan S., Aschoff J. Circadian rhythms of locomotor activity in captive birds and mammals: their variations with season and latitude // Oecologia.1975. V. 18. P. 269–316.
De Coursey P.J. Light-sampling behavior in photoentrainment of a rodent circadian rhythm // J. Comp. Physiol. A. 1986. V. 159. P. 161–169.
Dobson F.S. Environmental influences on sciurid mating systems // The Biology of Ground-Dwelling Squirrels / Eds Murie J.O., Michener G.R. Lincoln, L.: Univ. Nebraska Press. 1984. P. 227–249.
Drabek C.M. Home range and daily activity of the round-tailed ground squirrel, Spermophilus tereticaudus neglectus //Am. Midl. Nat. 1973. V. 89. P. 287–293.
Everts L.G., Strijkstra A.M., Hut R.A., Hoffmann I.E., Millesi E. Seasonal variation in daily activity patterns of free-ranging European ground squirrels (Spermophilus citellus) // Chronobiol. Int. 2004. V. 21. P. 57–71.
Froy O. The relationship between nutrition and circadian rhythms in mammals //Front. Neuroendocrinol. 2007. V. 28. P. 61–71.
Heller H.C., Ruby N.F. Sleep and circadian rhythms in mammalian torpor // Annu. Rev. Physiol., 2004. V. 66. P. 275–289.
Hut R.A., van Oort B.E., Daan S. Natural entrainment without dawn and dusk: the case of the European ground squirrel (Spermophilus citellus) // J. Biol. Rhythms. 1999. V. 14. P. 290–299.
Katona K., Váczi O., Altbäcker V. Topographic distribution and daily activity of the European ground squirrel population in Bugacpuszta, Hungary // Acta Theriol. 2002. V. 47. P. 45–54.
Kenagy G.J., Hoyt D.F. Speed and time-energy budget for locomotion in golden-mantled ground squirrels // Ecology. 1989. V. 70. P. 1834–1839.
Loehr K.A., Risser A.C. Daily and seasonal activity patterns of the Belding ground squirrel in the Sierra Nevada // J. Mammal. 1977. V. 58. P. 445–448.
Long R.A., Hut R.A., Barnes B.M. Simultaneous collection of body temperature and activity data in burrowing mammals: a new technique // J. Wildl. Manag. 2007. V. 71. P. 1375–1379.
Long R.A., Martin T.J., Barnes B.M. Body temperature and activity patterns in free-living arctic ground squirrels // J. Mammal. 2005. V. 86. P. 314–322.
Malan A. Is the torpor-arousal cycle of hibernation controlled by a non-temperature-compensated circadian clock? // J. Biol. Rhythms. 2010. V. 25. P. 166–175.
Meredith M., Ridout M., Meredith M.M. Package ‘overlap’. Estimates of coefficient of overlapping for animal activity patterns. 2021. https://CRAN.R-project.org/package=overlap.
Pinheiro J., Bates D. R Core Team. nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1-159. 2022. https://CRAN.R-project.org/package=nlme.
Podolski I., Belotti E., Bufka L., Reulen H., Heurich M. Seasonal and daily activity patterns of free-living Eurasian lynx Lynx lynx in relation to availability of kills // Wildlife Biol. 2013. V. 19. P. 69–77.
R Core Team. R: a language and environment for statistical computing. Vienna (Austria): R Foundation for Statistical Computing. 2021. Available from: https://www.R-project.org/.
Refinetti R. Circadian physiology. CRC press. 2019. 174 p.
Ridout M.S., Linkie M. Estimating overlap of daily activity patterns from camera trap data // J. Agric. Biol. Environ. Stat. 2009. V. 14. P. 322–337.
Rowcliffe M., Rowcliffe M.M. Package ‘activity’. Animal activity statistics R Package Version 1.3.2.2022. https://CRAN.R-project.org/package=activity.
Schmidt K. Variation in daily activity of the free-living Eurasian lynx (Lynx lynx) in Białowieża Primeval Forest, Poland // J. Zool. 1999. V. 249. P. 417–425.
Váczi O., Koósz B., Altbäcker V. Modified ambient temperature perception affects daily activity patterns in the European ground squirrel (Spermophilus citellus). J. Mammal. 2006. V. 87. P. 54–59.
Vasilieva N.A., Pavlova E.V., Naidenko S.V., Tchabovsky A.V. Age of maturation and behavioral tactics in male yellow ground squirrel Spermophilus fulvus during mating season // Cur. Zool. 2014. V. 60. P. 700–711.
Vasilieva N.A., Tchabovsky A.V. Timing is the only thing: Reproduction in female yellow ground squirrels (Spermophilus fulvus) // Can. J. Zool. 2014. V. 92. P. 737–747.
Vasilieva N.A., Tchabovsky A.V. A shortage of males causes female reproductive failure in yellow ground squirrels // Science Advances. 2015. V. 1. № 9. e1500401.
Vieira E.M., Baumgarten L.C., Paise G., Becker R.G. Seasonal patterns and influence of temperature on the daily activity of the diurnal neotropical rodent Necromys lasiurus. Can. J. Zool. 2010. V. 88. P. 259–265.
Vispo C.R., Bakken G.S. The influence of thermal conditions on the surface activity of thirteen-lined ground squirrels. Ecology. 1993. V. 74. P. 377–389.
Waterman J.M. Male mating strategies in rodents // Rodent societies: an ecological and evolutionary perspective / Eds Sherman P.W., Wolff J.O. Chicago: Univ. Chicago Press. 2007. P. 27–41.
Williams C.T., Wilsterman K., Kelley A.D., Breton A.R., Stark H., Humphries M.M., McAdam A.G., Barnes B.M., Boutin S., Buck C.L. Light loggers reveal weather-driven changes in the daily activity patterns of arboreal and semifossorial rodents // J. Mammal. 2014. V. 95. P. 1230–1239.
Wilsterman K., Buck C.L., Barnes B.M., Williams C.T. Energy regulation in context: free-living female arctic ground squirrels modulate the relationship between thyroid hormones and activity among life history stages // Horm. Behav. 2015. V. 75. P. 111–119.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая