

Известия РАН. Серия биологическая, 2023, № 6, стр. 650-658
К экологии видов-двойников обыкновенной полевки – Microtus arvalis Pall. и Microtus rossiaemeridionalis Ognev на северной периферии ареала
1 Петрозаводский государственный университет
185910 Петрозаводск, просп. Ленина, 33, Россия
2 Карельский научный центр РАН
185000 Петрозаводск, ул. Пушкинская, 11, Россия
* E-mail: Ivanter@petrsu.ru
Поступила в редакцию 07.08.2022
После доработки 09.12.2022
Принята к публикации 11.06.2023
- EDN: NZFQVS
- DOI: 10.31857/S1026347022600881
Аннотация
На основании многолетних (1958–2018 гг.) полевых и стационарных исследований дана характеристика численности, демографической структуры популяции, хода репродукции, питания и других экологических особенностей, общих для видов-двойников обыкновенной полевки (Microtus arvalis sensu lato). Установлено, что в прошедшее шестидесятилетие северная граница их распространения сначала переместилась к северу и достигла 63°40′ с.ш., а затем также быстро отступила к югу. Причина последнего – резкое сокращение посевных площадей, прежде всего, зерновых, в связи с переориентацией сельского хозяйства с зернового на молочно-овощное. Это, а также замещение обыкновенных полевок аборигенными видами грызунов (экономкой, водяной, темной и рыжей полевками) вызвало резкое сокращение численности и сужение ареала видов в восточных районах Фенноскандии. В питании видов-двойников основное место занимает зеленая масса травянистых растений (показатель встречаемости 72%), меньшую – семена (25%) и совсем редкий компонент – кора деревьев (3%). В условиях северной периферии ареала размножение протекает лишь в течение одного, причем достаточно ограниченного, весенне-летнего сезона. Возрастной и половой состав населения неоднороден и меняется по сезонам.
Вопросу дифференциации хромосомных видов-двойников серых полевок группы arvalis, обитающих в Восточной Фенноскандии в условиях северной периферии ареала, до последнего времени практически не уделялось должного внимания. Тем не менее установлено, (Малыгин, 1983; Соколов и др., 1994; Громов, Ербаева, 1995; Мейер и др., 1996; Бобров и др., 2008; Хляп, Варшавский, 2010; Малыгин и др., 2019), что в северной части европейской области распространения, в том числе и в Карелии, встречаются, хотя и не часто, оба вида, причем считается, что преобладающим из них является обыкновенная полевка. Что же касается различий в образе жизни, то в целом, по мнению большинства исследователей, они вряд ли существенны (Малыгин, 1974; Жигарев, 1993, 2004; Доброхотов и др., 1985; Баскевич и др., 2009, 2012; Малыгин, Рябов, 2013, Жигарев и др., 2019), что и позволило нам приводить полученные по их экологии материалы совместно, не разделяя их по видам.
Обыкновенная и восточноевропейская полевки, обе обитатели нелесных биотопов (Башенина, 1962; Малыгин, 1974; Доброхотов и др., 1985; Жигарев, 1993, 2004, Малыгин, Рябов, 2013). Использование ими лесных местообитаний ограничено нарушенными участками, такими как лесопарки, сады, рекреационно-нарушенные леса (Мейер и др., 1996; Карасева и др., 1999; Ермолаева и др., 2000; Жигарев, 2004). По мнению В.М. Малыгина (1983), восточноевропейские полевки в большей степени, чем обыкновенные, занимают пограничные местообитания, сочетающие луга и поля с кустарниками и садами. В целом же в большинстве мест, при совместном обитании видов-двойников, обычно численно доминируют обыкновенные полевки, лишь в природных биотопах Москвы восточноевропейские полевки в этой паре видов преобладают (Жигарев и др., 2019). Большую склонность к обитанию Microtus rossiaemeridionalis на урбанизированных территориях отмечают многие исследователи (Карасева и др., 1999; Ермолаева и др., 2000; Жигарев, 2004; Жигарев и др., 2019). Имеются данные об определенной склонности восточноевропейских полевок к заселению хозяйственных построек человека (Жигарев, 1993). Кроме того, были обнаружены и некоторые отличия в предпочтении видами-двойниками различных типов антропогенных биотопов. Так, обыкновенные полевки чаще занимают влажные и мезофильные луга и поля, а восточноевропейские – более сухие участки. Отмечено также, что восточноевропейские полевки достигают большей численности на сильно нарушенных человеком территориях (Жигарев, 2004).
Такова общая ситуация с существованием видов-двойников обыкновенной полевки в различных частях их ареала. Этому соответствует и крайняя территориальная неравномерность их изученности. В итоге большинство публикаций, касающихся экологии этих видов, относится к центральным и южным районам Европейской России (Виноградов, 1934; Формозов, 1947; Огнев, 1950; Карасева, Кучерук, 1954; Башенина, 1962, 1968, 1977; Лапинь, 1063; Громов, Поляков, 1977; Мейер и др., 1996; Малыгин, 1983; Соколов и др., 1994; Жигарев, 2004; Баскевич и др., 2009), приграничные же их популяции, и в том числе занимающие северную периферию ареала, изучены совершенно недостаточно. Появляющиеся от случая к случаю и, как правило, фрагментарные исследования видов-двойников в этих условиях крайне немногочисленны и спорадичны (Исаков, 1939; Снигиревская, 1954; Марвин, 1959; Айрапетьян, 1970). Настоящее сообщение – достаточно скромная попытка частично восполнить этот существенный пробел.
МАТЕРИАЛЫ И МЕТОДЫ
Материалом для настоящей работы послужили проводившиеся в Карелии многолетние (1958–2018 гг.) сборы зверьков рассматриваемых видов Microtus arvalis и M. rossiaemeridionalis, отлавливаемые при учетах численности двумя основными методами: ловушко-линиями (плашки Геро) и ловчими 30-метровыми канавками. При этом общий объем проведенных нами учетов составил свыше 360 тыс. ловушко-суток и более 9 тыс. канавко-суток. В итоге на ловушко-линиях, относительно равномерно разбросанных по всей исследованной территории было поймано 33 обыкновенные полевки, а в ловчие канавки – 45. Таким образом, общее число добытых и исследованных нами при вскрытии особей рассматриваемых видов-двойников составило 78. К ним можно добавить 144 экз. недифференцированных по видам зверьков, добытых в антропогенном ландшафте в различных районах Карелии в период до 1960-х гг., сведения по которым сохранились в архивах Института биологии Карельского центра РАН (в картотеке и коллекционных журналах того времени).
Устройство нор изучали путем их раскапывания и зарисовки на планшетах, питание – посредством разбора поедей на кормовых столиках и при анализе содержимого желудков, возраст – по конфигурации черепа и стертости зубов, активность репродукции, зрелость и активность половой системы – по ее состоянию и характерным изменениям у вскрытых животных. Оценку динамики численности, полового и возрастного состава популяции, а также биотопического размещения животных, проводили по общепринятым методикам (Новиков, 1953; Карасева и др., 2008).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Численность и ее изменения
Проводившееся на Северо-западе России многолетние учеты численности мелких млекопитающих (1965–2018 гг.) показывают, что в изученном регионе оба вида обыкновенной полевки встречаются спорадически и в целом редки. В коллекционных сборах 1948–1972 гг., их насчитывалось всего 49 экз., в Карельской же части былого ареала численность вида и вовсе упала до нуля. Правда, затем, с 2001 по 2011 гг., представители этого вида снова стали ежегодно попадаться по несколько экземпляров в уловах наших южных стационаров – Приладожском (Карку, Питкярантский р-н Карелии) и Каскеснаволокском (Пряжинский). В первом из них после почти 40‑летнего перерыва зверьков этого вида впервые (6 экз.) отловили на линиях ловушек в 2001 г., а затем в ловчие канавки – в 2005 (1 экз.), 2009 (3) и 2011 (1), после чего они полностью исчезли. В Каскеснаволоке же с 2001 г. она по несколько экземпляров ловилась ежегодно, но с 2012 г. снова перестала появляться в уловах и отсутствует в них до сих пор, причем это касается обоих южных стационаров (Якимова, 2018). Показательны в этом отношении и учеты в окрестностях с. Кончезеро (Кондопожский р-н Средней Карелии). Если в 1948–1956 гг. обыкновенная полевка составляла здесь от 3 до 10% отловленных зверьков, то в 1966–1972 гг. она вообще не была здесь ни разу встречена. То же самое происходило и в Лахденпохском районе. В 1951 г. здесь отловили 22 экз. обыкновенны полевок, или 3% от общего числа отловленных зверьков, тогда как проведенные там же учеты в 1959 и 1961 гг. вообще не выявили присутствие этого вида. Причем, это связано с тем, что на Северо-Западе России происходит, связанное с переходом на молочно-овощное сельское хозяйство, резкое сокращение зерновых площадей, что вместе с вытеснением обыкновенной полевки аборигенными видами грызунов и вызвало резкое сокращение численности и сужение ее ареала.
В общем же, как показывают учеты, на таежном Северо-Западе России и, в частности, в Карелии численность видов-двойников обыкновенной полевки держится в последние десятилетия на крайне низком уровне: средний многолетний показатель учетов составил для них 0.06 экз. на 100 ловушко-суток (степень доминирования 0.05%) и 0.17 на 10 канавко-суток (0.2%). Недостаток подходящих местообитаний и основных кормов для обыкновенной полевки, связанный с переходом земледелия с зерновых на пропашные культуры, зарастанием полей сенокосных и выпасных лугов, исчезновением деревень и общим значительным сокращением сельхозугодий, а также вытеснение ее аборигенными видами грызунов (экономкой, водяной, темной и рыжей полевками) послужило для видов-двойников обыкновенной полевки главной причиной наблюдавшегося в последние годы сокращения численности и сужения ареалов на территории Карелии
По М.Я. Марвину (1959), волосяной покров обыкновенных полевок по сравнению с другими видами грызунов Карелии отличается наименьшей густотой. Это обстоятельство он считает главной причиной отсутствия представителей этих видов в северной половине республики.
Между тем, по мнению Н.В. Башениной (1962), проникновению обыкновенных полевок на Крайний Север препятствует весь комплекс экологических условий: заболачивание, малая площадь пашен, биоценотические конкурентные отношения с близкими видами и т.д. Это хорошо объясняет как процесс постепенного продвижения видов на север вслед за земледелием, так и отступление их в последние годы в связи с резким сокращением здесь посевных площадей под зерновые.
При стабильном чрезвычайно низком уровне численности обыкновенных полевок, характерном для всей территории Восточной Фенноскандии, как, впрочем, и для других периферических популяций этого, да и других видов наших мелких млекопитающих, уловить ее колебания ни по годам, ни по сезонам не представляется возможным. В других же регионах она демонстрирует вполне определенные периодически повторяющиеся подъемы и спады, позволяющие говорить о четкой их цикличности. Еще Н.В. Башениной (1962, 1977, 1994) было показано, что для рассматриваемых видов на всем протяжении их ареалов, скорее всего, можно с достаточной долей уверенности выделить определенную периодичность наступления пиков и депрессий численности с частотой их повторяемости через 3–5–8–12 лет. Правда, для всего ареала такой единой цикличности, наверное, и не существует, однако для отдельных, но достаточно больших территорий ее, как правило, удается вычленить. В частности, были установлены и впоследствии многократно подтверждены “большие волны” колебания численности мышевидных грызунов на обширных территориях с периодом 10–11 лет и “малые волны” с периодом 3–5 лет на более локальных территориях.
По расстоянию между пиками определен 3–4‑летний цикл у видов-двойников обыкновенной полевки а также темной и американских видов рода Microtus (Frank, 1957; Migula et al., 1975), что позволило авторам считать цикличность, в принципе, присущей всем этим грызунам (Krebs, Myers, 1974), однако для тех же видов описываются и “нециклические популяции” (Krebs, Myers 1974; Keller, 1981), правда, при наблюдениях не более 5 лет. Тем не менее, проведенное математическое моделирование многолетней динамики численности изучаемых видов в условиях Стрелецкой степи (Баскевич и др., 2012) показало, что в наблюдаемых там изменениях прослеживается цикличность близкая к трехлетней, причем наряду с этим имеются все основания предполагать колебания с интервалом 10–11 лет. При этом особенно важно, что в данном случае гипотеза цикличности не задавалась в модели заранее, а была получена в результате учета экологических факторов.
В итоге удалось установить, что в связи с существенными изменениями, которые вносит в динамику численности исследуемых видов антропогенный пресс, наблюдаются периодически повторяющиеся вспышки их численности. Так, в Польше установлены трехлетние циклы массового размножения грызунов с доминированием обыкновенной полевки (Пекарчик, 1983). На территории Молдавии в течение 1971–1986 гг. отмечены три высоких (1975, 1981, 1985) и четыре средних пика численности (Мунтяну и др., 1987). В Латвии массовые размножения (большие волны) повторяются через 9–12 лет (в среднем 11). Фазы с высоким уровнем численности обычно охватывали все районы республики и проявлялись в местах, где выращивались многолетние травы и озимые культуры (Тауриньш, 1964).
В течение 1978–2002 гг. сезонную и многолетнюю динамику численности обыкновенной полевки изучали в различных биотопах юго-западной и центральной Белоруссии (Блоцкая, Гайдук, 2004). В итоге удалось установить, что в связи с существенными изменениями, которые вносит в динамику численности этого вида антропогенный пресс, периодически повторяющиеся вспышки его численности в естественных биотопах и агроценозах полностью не реализуются.
В этом ключ к эффективному прогнозированию и регулированию численности грызунов. Без знания реальных популяционных процессов, их закономерностей и определяющих факторов, не возможно ни управление природными популяциями и контролирование их численности, ни сохранение уникального природного баланса, ни рациональное земледелие.
Биотопическое размещение
Согласно В.М. Малыгину с соавторами (2019), собственно инвазионный процесс, заключавшийся в ускоренном расселении видов далеко за пределы нативного ареала – у полевок группы “arvalis” повсеместно был связан с антропогенными факторами, и прежде всего с сельскохозяйственным освоением земель человеком. Будучи мезофильными видами, эти полевки изначально обитали во второстепенных по площади луговых участках лесостепей и северных степей, в основном расположенных вдоль рек и озер. В зоне же хвойных лесов подходящих для них местообитаний было крайне мало разве что мелкие разрозненные участки приречных и приручьевых лугов и и олуговевшие гари. В итоге изначально весь исконный ареал полевок группы “arvalis” представлял собой редкое “кружево”, и поселения этого вида нигде, тем более у границ ареала, не занимали фоновых биотопов (Тупикова и др., 2000; Бобров и др., 2008; Малыгин и др., 2019). Так постепенно новым местообитанием, значительно более благоприятным, чем исконные, оказались агроценозы, где рассматриваемые виды полевок достигали все более высоких показателей численности (Тупикова и др., 2000; Неронов и др., 2001), при этом площадь пригодных местообитаний росла по мере увеличения площадей пахотных земель. и снижалась соответственно сокращению посевов зерновых Все это и привело к становлению общей картины современного биотопического размещения видов-двойников обыкновенной полевки.
Как установлено нашими исследованиями, в настоящее время на таежном Северо-Западе России эти оба селятся преимущественно в открытых местообитаниях, включая культурные земли (сельскохозяйственные угодья и др.) – на возделываемых полях и среди посевов многолетних трав. Зимой же они встречаются в стогах сена и скирдах соломы, где обитают вместе с другими представителями рода Microtus – темной полевкой и экономкой, не избегая, впрочем, и соседства с рыжей полевкой. Очевидно, в этот период конкурентные отношения у этих видов выражены слабо. Близкие данные по особенностям современного биотопического размещению обыкновенных полевок получены и в других областях их ареала (Малыгин, 1974; Доброхотов и др., 1985; Соколов и др., 1994).
Устройство убежищ
Подземные убежища обыкновенные полевки устраивают обычно на небольшой глубине: чаще всего 10–30 см, а если глубже (например, в пахотном слое почвы), до 30 и даже 50 см, то норка кончается обычно глухой камерой и имеет один выход. Строение же гнездовых нор наиболее разнообразно (рис. 1, 2). Количество выходов бывает от 1 до нескольких десятков, но обычно не более 10. В наших условиях, как и вообще у северных границ ареала, где норная деятельность полевок наименее выражена, этот зверек довольствуется самыми примитивными по строению подземными жилищами с 1–2, реже 3–4 выходами. Общая длина норы редко превышает здесь 0.5–1, максимум – 1.5 м. Общая площадь раскопанных Н.В. Башениной (1962) гнездовых нор колебалась от 0.18 до 6 м2, в северных же районах, в том числе в Карелии, не превышала, по нашим данным, 0.5–1.5 м2. Расположенные на глубине до 60, но чаще в пределах 20–30 см, гнездовые камеры имеют обычно форму неправильного шара с диаметром не превышающим 9–17 (в среднем 14.5) см с отходящими от них на различной высоте от 1 до 3–4 ходов.
Рис. 1.
Защитно-кормовые (а) и защитные (б) норы обыкновенной полевки. 1 – подземные ходы с отверстиями; 2 – наземные тропинки; 3 – подземные ходы (нижние этажи); 4 – запасы.
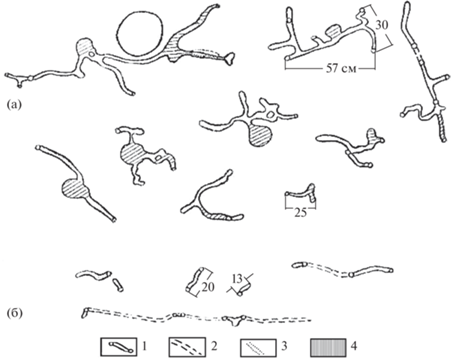
Рис. 2.
Примеры строения гнездовых нор обыкновенной полевки: (а) – план; (б) – разрез (цифры – глубина в см). 1 – подземные ходы; 2 – ходы “нижних этажей”; 3 – жилые гнезда; 4 – старые гнезда.
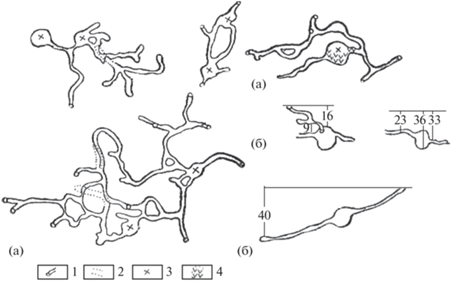
Гнездо устраивается из листьев и расщепленных вдоль на узкие полоски стеблей злаков. Изнутри оно обычно выстилается растительным пухом. Подснежные гнезда обыкновенных полевок размещаются на участках, где глубина снежного покрова превышает 40 см. Обычно они располагаются на межах, оврагах, стерне. Например, в Белоруссии (Блоцкая, Гайдук, 2004) плотность таких гнезд на 100 м2 в различные годы варьировала от 1 до 3. Подснежные гнезда отличались там от летних большей толщиной стенок и теплой выстилкой. В основании гнезда в большинстве случаев присутствует земля, а в качестве строительного материала, как и летом, используются сухие стебли и листья растений. Так как в подснежных гнездах часто находятся несколько зверьков, то такие коллективные жилища имеют, по сравнению с летними гнездами, гораздо большие размеры. Диаметр подснежных гнезд, по их данным, варьировал в пределах 10–22, в среднем 15.6 см, высота 7–14, в среднем 11.2, см, толщина дна – 3–7, в среднем 5.1, см. Температура же в гнездах, в которых находились детеныши, колебалась от 17 до 35°C (ср. = 28.4 ± 1.3°C (n = 12)).
Иногда обыкновенные полевки делают убежища в несвойственных им местах. Один из таких случаев описывает Н.В. Башенина (1962), обнаружившая на севере Вологодской области на окраине обследованной ею заброшенной пашни небольшое поселение этих полевок, устроивших норы в старых замшелых грудах валунов, заросших травой и кустарником. Сеть тропинок в траве неизменно приводила к центру заросшей полосы, где под кустами и дерновинами можно было найти в земле 2–3 отверстия, остальными входами служили щели между камнями.
Питание
Согласно проведенному нами исследованию содержимого 42 желудков, основную роль в питании обоих видов обыкновенных полевок играют зеленые части травянистых растений (показатель встречаемости 72%), меньшую – семена (25%) и совсем редкий компонент – кора деревьев (3%). В то же время проведенное нами вслед за М.Я. Марвиным (1959) экспериментальное скармливание карельским полевкам различных кормов в неволе дополняет приведенный им список из 71 вида растений, еще несколькими десятками кормовых объектов, в том числе обнаруженных в норах и кормовых камерах пищевых остатков. В числе предпочитаемых зверьками кормов можно назвать различные виды клевера, вику, щавель, одуванчик, мышиный горошек, чину, сныть, подорожник, тысячелистник, ячмень, пшеницу, рожь, овес, картофель и др.
Размножение и экологическая структура популяции
Известно (Башенина, 1962), что в пределах всего обширного ареала оба рассматриваемых вида отличается высоким репродуктивным потенциалом и способны даже к круглогодичному размножению. В Карелии же, в условиях северных пределов ареала, они размножаются в течение лишь одного, причем достаточно ограниченного весенне-летнего сезона. При этом сперматогенез и овогенез происходит у них по общей, типичной для всех мелких млекопитающих схеме. Тем не менее, данные по размножению обыкновенных полевок в Карелии весьма фрагментарны. Почти все отловленные нами самки (18 из 22) или размножались (были беременными, кормили выводок), или уже завершили размножение (темные пятна на рогах матки и рубцы в яичниках).
Как и в других частях ареала, в Карелии размножение зверьков рассматриваемых видов, в основном, определяется состоянием самок, течка которых подвержена влиянию многочисленных и разнообразных факторов среды и зависит от внутреннего состояния организма в большей степени, чем гон самцов. Средняя продолжительность цикла, как и у других мелких грызунов, 5–6 дней, однако продолжительность фаз может варьировать в зависимости от условий.
В Карелии, как и в других, более южных областях обитания, обыкновенные полевки могут, видимо, хотя и не регулярно, размножаться и зимой. Косвенным подтверждением этого является поимка 6.03.1981 г. в стоге сена на агробазе Института биологии Карельского филиала АН СССР трех самок с отчетливо выраженными плацентарными пятнами в рогах матки. Там же была добыта пара молодых самцов весом 12 и 16 г. В то же время зимнее размножение происходит здесь при хорошей обеспеченности кормом в скирдах, ометах и зернохранилищах
Приведенная в табл. 1 характерная для карельских полевок средняя величина выводка (6.7 ± 0.6) оказалась несколько (но все-таки статистически достоверно) выше средней видовой нормы, равной 5.0–6.0. Обыкновенные полевки Ленинградской обл. также отличаются повышенной плодовитостью: среднее число эмбрионов на 1 самку – 5.8 (Айрапетьянц, 1970). Это согласуется с обнаруженной наличием у них тенденции к увеличению плодовитости в зоне пессимума (Башенина, 1962).
Таблица 1.
Величины выводков молодых и взрослых обыкновенных полевок в разных регионах
| Место | Средняя величина выводка | Источник | |
|---|---|---|---|
| взрослые | молодые | ||
| Ленинградская обл. | 5.8 | 5.6 | Айрапетьянц, 1970 |
| Карелия | 6.9 | 5.3 | Наши данные |
| Вологодская обл. | 5.4 | 4.1 | Башенина, 1962 |
| Московская обл. | 5.6 | 4.6 | Башенина, 1962 |
| Брянская обл. | 6–7 | 4.9 | Башенина, 1962 |
| Татария | 5.4–6.3 | 4.4 | Назарова, 1958 |
| Башкирия | 6.2 | 5.3 | Снигиревская, 1954 |
| Среднее Поволжье | 4.5–5.6 | 4.7–5.0 | Башенина, 1968 |
| Черкасская обл. | 5.4 | 4.65 | Башенина, 1994 |
| Красноярский край | 4.0–6.1 | 3.5–5.7 | Башенина, 1994 |
| Сев. Осетия | 5.1–5.8 | 4.9–5.6 | Башенина, 1994 |
| Запорожская обл. | 5.6 | 4.8 | Наумов, 1948 |
| Белоруссия | 5.1 | 4.95 | Блоцкая, Гайдук, 2004 |
Возрастная структура карельских популяций обыкновенной полевки закономерно изменяется в течение года (табл. 2). Зимой в них присутствуют исключительно прибылые зверьки поздних выводков прошлого года, в Белоруссии же (Блоцкая, Гайдук, 2004), как и в большинстве других расположенных ближе к центру ареала регионов, еще и часть особей ранних выводков. Взрослые перезимовавшие полевки прошлого года рождения в уловах не попадаются. Продолжительность их жизни в природе составляет около 20 мес., так что две зимы зверьки этого вида в наши условиях не переживают. В конце марта – первой декаде апреля (у северных границ ареала, включая Карелию, это происходит почти на месяц позднее) обыкновенные полевки начинают размножаться, и во второй половине мая в уловах нашего региона начинают попадаться прибылые, которые весной составляют до 40% населения зверьков.
Таблица 2.
Возрастной и половой состав карельских популяций обыкновенной полевки
| Месяцы | Число взрослых | Число прибылых ранних выводков | Число прибылых поздних выводков | Общее число, n | Относительное число зверьков, % | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| взрослые | прибылые | |||||||||
| n | из них самцы, % | n | из них самцы, % | n | из них самцы, % | ранних выводков | поздних выводков | |||
| Май–июнь | 21 | 66.7 | 15 | 60.0 | – | – | 36 | 58.3 | 41.7 | – |
| Июль–август | 11 | 54.5 | 43 | 58.1 | 49 | 67.1 | 103 | 15.5 | 59.2 | 25.3 |
| Сентябрь–октябрь | 5 | 60.0 | 26 | 50.0 | 52 | 59.3 | 83 | 8.4 | 31.3 | 60.3 |
| Весь период | 37 | 62.1 | 84 | 57.1 | 101 | 57.4 | 222 | 19.8 | 45.9 | 34.3 |
В летний период к группам зимовавших и зверьков ранних выводков в популяцию включаются прибылые поздних выводков, причем их становится все больше и больше. В Белоруссии, и других зонах оптимума в летних уловах на их долю приходится до 60% , в Карелии же, где оборот популяции проходит гораздо быстрее, еще больше 64–75%. Поздней осенью зимовавшие особи составляют в Белоруссии не более 15%, а на северной периферии ареала, в том числе и в нашем регионе, их уже нет вовсе. Они присутствуют в уловах до августа, иногда задерживаясь до середины – конца сентября. В дальнейшем же они быстро стареют и полностью вымирают. Фактически та же судьба постигает и подавляющее большинство полевок ранних выводков. Все они, как правило, активно участвуют в размножении, принося от 1 до 3 выводков, однако к концу репродуктивного сезона растрачивают все свои внутренние резервы и к концу лета – началу осени полностью элиминируются из популяции. Особи же поздних выводков, напротив, осенью преобладают в популяции и практически составляют основу зимнего населения зверьков. Половое же созревание происходит у них лишь следующей весной, после успешной зимовки и ускоренного весеннего роста.
Тем не менее, в целом для репродукции изученных популяций характерен крайне низкий темп и неровный ритм репродукции, определяющий соответствующий “рваный” ритм изменений численности вида, что весьма типично для периферийных зон видовых ареалов этих грызунов.
ЗАКЛЮЧЕНИЕ
Выявленные в процессе исследований экологические особенности, характерные для видов-двойников обыкновенной полевки в специфических условиях северо-западных пределов ареала, отличаются значительной лабильностью и гармоничной согласованностью с эндо- и экзогенными факторами среды. Наряду с узкими специальными приспособлениями индивидуального ранга здесь успешно реализуются структурно-популяционные адаптивные комплексы, отличающиеся гибкостью, динамичностью и высокой скоростью компенсаторной перестройки. Более выражены и гораздо рельефнее проявляются вблизи северных границ распространения и специфические структурно-популяционные адаптации, направленные на максимально эффективное воспроизводство видового населения, способное обеспечить виду и его популяциям возможности преодолевать экстремальные условия экологического пессимума. Наиболее характерны в этих условиях частые¸ глубокие и длительные депрессии численности при относительно коротких и невысоких ее подъемах.
Тем не менее в начале сезона размножения численность молодых зверьков и полнота их участия в репродукции во многом определяется погодными условиями. К концу же сезона размножении большое значение приобретает ускоренное половое созревание и максимально полное участие в воспроизводстве популяции сеголеток разного времени рождения.
Работа проводилась в соответствии с Государственным заданием федерального исследовательского центра “Карельский научный центр Российской академии наук.”
Список литературы
Айрапетьянц А.Э. Насекомоядные и грызуны // Звери Ленинградской области. Л.: ЛГУ, 1970. С. 47–165.
Баскевич М.И., Потапов С.Г., Окулова Н.М., Сапельников С.Ф., Власов А.А. и др. К распространению и изменчивости видов двойников Мicrotus arvalis sensu lato (Rodentia, Arvicolinae) в Центральном Черноземье по хромосомным и молекулярно-генетическим данным // Зоологический журн. 2009. Т. 88. Вып. 4. С. 473–483.
Баскевич М.И., Окулова Н.М., Потапов С.Г., Миронова Т.А., Сапельников С.Ф. и др. Новые данные о распространении видов-двойников и гибридизации 46 хромосомнеых форм Microtus arvalis sensu lato (Rodentia, Arvicolinae) в Центральном Черноземье // Зоологический журн. 2012. Т. 91. Вып. 8. С. 994–1005.
Башенина Н.В. Экология обыкновенной полевки и некоторые черты ее географической изменчивости. М.: МГУ, 1962. 307 С.
Башенина Н.В. Материалы к экологии мелких млекопитающих зоны европейской тайги // Уч. зап. Пермск. гос. пед. ин-та. 1968. Т. 52.
Башенина Н.В. Пути адаптаций мышевидных грызунов. М.: Наука, 1977. 294 с.
Блоцкая Е.С., Гайдук В.Е. Популяционная экология мелких млекопитающих. Брест: Брест. гос. ун-та, 2004. 187 с.
Бобров В.В., Варшавский А.А., Хляп Л.А. Чужеродные виды млекопитающих в экосистемах России. М.: Товарищество научных изданий КМК, 2008. 232 с.
Виноградов Б.С. Материалы по динамике фауны мышевидны грызунов СССР Л., 1934. с.
Громов И.М., Ербаева М.А. Зайцеобразные и грызуны. Млекопитающие фауны России. СПб., 1995. 522 с.
Громов И.М., Поляков И.Я. Полевки Microtinae // Фауна СССР. Л.: Наука, 1977. Млекопитающие. Т. 3. Вып. 8. 504 с.
Доброхотов Б.П., Барановский П.М., Демидова Т.Н. Особенности стациального распределения видов-двойников Microtus arvalis и M. rossiaemeridionalis Rodentia, Microtinae) и их роль в природных очагах туляремии луго-полевогг типа // Зоологическвий журн. 1985. Т. 64. Вып. 2. С. 269–275.
Ермолаева Е.З., Карасева Е.В., Телицына А.Ю. Особенности обитания мелких млекопитающих в Ботаническом саду МГУ (Воробьевы горы) // Животные в городе. Материалы научно-практической конференции. М. 2000. С. 34–37.
Жигарев И.А. Изменения плотности населения мышевидных грызунов под влиянием рекреационного пресса на юге Подмосковья // Зоологический журн. 1993. Т. 72. Вып. 12. С. 117–137.
Жигарев И.А. Мелкие млекопитающие рекреационных и естественных лесов Подмосковья. М.: Изд-во Прометей, 2004. 232 с.
Жигарев И.А., Жигарев Д.И., Алпатов В.В., Лапковский В.В., Малыгин В.М., Симак С.В. Белковый электрофорез в полиакриламидном геле, как метод идентификации видов-двойников обыкновенной полевки (Microtus arvalis Pallas 1779 и M. rossiaemeridionalis Ognev 1924 (Rodentia, Cricetidae) // Известия РАН. Серия биологическая, 2019. № 4. С. 380–388.
Исаков Ю.А. Материалы по фауне млекопитающих Средней и Северной Карелии // Бюл. Моск. о-ва испыт. прир. Отд. биол. 1939. Т. 48. Вып. 2–3.
Караcева Е.В., Кучерук В.В. Изучение подвижности обыкновенных полевок с помощью мечения зверьков // Матер. экол. конф. Тез. докл. М. 54. С. 80–83.
Карасева Е.В., Телицына А.Ю., Жигальский О.А. Методы изучения грызунов в полевых условиях. М.: Из-во ЛКИ, 2008. 416 с.
Карасева Е.В., Телицина А.Ю., Самойлов Б.Л. Млекопитающие Москвы в прошлом и настоящем. М.: Наука, 1999. 245 с.
Лапинь И.М. Биология и паразитофауна мелких лесных млекопитающих Латвийской ССР. Рига, 1963. 135 с.
Малыгин В.М. Стациальное размещение видов-двойников обыкновенной полевки // Экология 1974. Т. 5. № 1. С. 89–91.
Малыгин В.М. Систематика обыкновенных полевок. М.: Наука, 1983. 208 с.
Малыгин В.М., Баскевич М.И., Хляп Л.А. Инвазии видов-двойников обыкновенной полевки // Российский журн. Биологических Инвазий 2019. № 4. С. 71–93.
Малыгин В.М., Рябов С.В. Распространение и биология видов-двойников обыкновенной полевки в зоомедицинском аспекте // Дезинфекционное дело. 2013. № 4. С. 27–35.
Марвин М.Я. Млекопитающие Карелии Петрозаводск. 1959. 238 с.
Мейер М.Н., Голенищев Н.М., Рабджабли С.И., Саблина О.Э. Серые полевки фауны России и сопредельных территорий // Труды Зоологического института РАН. Спб. 1996. С. 320
Мунтяну А.И., Савин А.И., Патрашку В.И., Сытник В.Л. Флуктуации численности опуляций обыкновенной полевки // Влияние антроп. трансф. ландшафта на население назем, позв. животных. Тез. Всес. совещ. М. 1987. Ч. 2. С. 26–27.
Назарова И.В. Эколого-морфологический очерк серых полевок Волжско-Камского края // Изв. Казанск. фил. АН СССР. Сер. биол. 1958. № 6. С. 27–41.
Наумов Н.П. Очерки сравнительной экологии мышевидных грызунов. М.; Л.: Изд-во АН СССР. 1948. 203 с.
Неронов В.Н., Хляп Л., Тупикова Н.В., Варшавский А.А. Изучение мирования сообщества грызунов на пахотных землях Северной Евразии // Экология, 2001. № 5. С. 355–362.
Новиков Г.А. Полевые исследования экологии наземных позвоночных животных Л.: Сов. наука. 1953. 602 с.
Огнев С.И. Звери СССР и прилежащих стран. 1950. Т. VII. Грызуны. М.; Л.: Изд-во АН СССР. 706 с.
Пекарчик К. Исследование по образованию прогнозов появления и развития вредителей сельскохозяйственных культур в Польше // Тр. ВИЗР, 1983. Вып. 18. С. 338–341.
Снигиревская Е.М. Экология и хозяйственное значение мышевидных грызунов в широколиственны лесах Жигулевской возвышенности. Автореф. дис. …. канд. биол. наук. Л., 1854. 24 с.
Соколов В.Е., Башенина Н.В., Малыгин В.М. и др. Обыкновенная полевка: виды-двойники. Microtus arvalis. Pallas. 1779, M. rossiaemeridionalis, Москва: Наука, 1994. 431 с.
Тауриньш Э.Я. Динамика размножения полевых мышевидных грызунов в Латвийской ССР с 1948 по 1962 гг. // Прогноз в защите растений от вредителей и болезней. Рига, 1964: Изд-во АН Латв. ССР. 239–261 с.
Тупикова Н.В., Хляп Л.А., Варшавский А.А., Грызуны полей Северо-Восточной Палеарктики // Зоологический журн. 2000. Т. 79. № 4. С 480–494.
Формозов А.Н. Очерк экологии мышевидных грызунов, носителей туляремии. М., 1947. 94 с.
Хляп Л.А., Варшавский А.А. Синантропные и агрофильные грызуны как чужеродные млекопитающие // Российский журн. биологических инвазий. 2010. № 3. С. 73–91.
Якимова А.Е. Результаты мониторинга мелких млекопитающих в Средней Карелии // Тр. КарНЦ РАН. Серия биогеография. 2018. № 1. С. 67–80.
Frank F. Tho causality of microtine cycles in. Germany. J. Wildl. Manag., 1957. 21 p.
Keller A. Determination des mammiferes de la uisse par leur pelage. IV. Crlcetidae et Muridae. Rev. uisse zool., 1981. V. 88. № 2. P. 463–473.
Krebs C.J., Myers J.H. Population cycles small mammals // Adv. Ecol. Res., V. 8. London-New York., 1974. P. 267–399.
Migula P., Gano B., Stepin Z., Bugdal U. Ocena zageszczenia i prizeplyw energii przez populoacje drobnych (Male Pleninu) // Prz. Zool. 1975. V. 19. № 4. P. 467–474.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия биологическая