

Известия РАН. Серия физическая, 2020, T. 84, № 11, стр. 1623-1627
Обнаружение с помощью 7-канального СКВИД-магнитометра рассеивания в коре мозга откликов на непрерывную ритмическую зрительную стимуляцию
В. Л. Введенский 1, *, Д. П. Бондарев 1, А. П. Гребенкин 1, С. П. Наурзаков 1, М. А. Поликарпов 1, В. Я. Панченко 1
1 Федеральное государственное бюджетное учреждение
“Национальный исследовательский центр “Курчатовский институт”
Москва, Россия
* E-mail: victorlvo@yandex.ru
Поступила в редакцию 18.06.2020
После доработки 10.07.2020
Принята к публикации 27.07.2020
Аннотация
С помощью 7-канального СКВИД-магнитометра измерены магнитные отклики мозга на ритмическую зрительную стимуляцию на частотах около 10 Гц. Несмотря на стационарность стимуляции, отклики мозга демонстрируют постоянную нестабильность амплитуды и пространственного распределения магнитного поля. Сигнал, поступающий от глаз, перенаправляется в разные участки коры, хотя стимуляция всегда одна и та же.
ВВЕДЕНИЕ
Исследование воздействия ритмической зрительной стимуляции на активность мозга имеет длительную историю, и вопрос представляется уже в значительной мере изученным [1]. Тем не менее, вплоть до настоящего времени ставятся многочисленные эксперименты, по результатам которых пытаются понять, как увязаны между собой собственная активность мозга и внешние воздействия, например, зрительные [2]. И хотя ритмическая зрительная стимуляция используется даже при прохождении врачебной комиссии на получение водительского удостоверения, до сих пор остается неясным, как же взаимодействуют собственная ритмика мозга и внешние стимулы. Исследователи надеются, что навязыванием альфа ритма (частотой около 10 Гц) с помощью повторяющихся вспышек можно оказать влияние на когнитивную деятельность мозга человека [3]. Тут неявно предполагается, что мозг откликается на внешний стимул путем резонансной активации собственных электрических осцилляторов в коре мозга. Однако не ясно, какой процесс наблюдается в каждом отдельном случае – наведенные колебания, или просто устойчивая реакция на стимул [2]. Несмотря на такую неясность, широко применяется метод “частотной метки”, при котором предъявляемое изображение, по какому-либо второстепенному параметру, например, виду шумового узора на объекте, модулируется с различными частотами в районе 10 Гц. В экспериментах на 306-канальном магнитометре путем усреднений по времени и сенсорам удается, хотя и неточно, определить области мозга, куда поступают сигналы на той или иной частоте и как они ведут себя во времени [4]. Знакомство с методикой магнитных измерений и процедурами обработки сигналов заставляет думать, что процессы в коре мозга организованы несколько иначе, чем та картина, которая возникает после усредняющих процедур. Основываясь на этом, мы провели измерения откликов мозга на ритмические световые вспышки с помощью 7-канального СКВИД-магнитометра.
МЕТОДИКА ЭКСПЕРИМЕНТА
Для описываемого исследования были выбраны четверо мужчин (19, 25, 65, 77 лет), имеющие частоту альфа ритма вблизи 10 Гц, и устойчивый отклик на оптическую стимуляцию при закрытых глазах. Испытуемые прошли предварительное клиническое ЭЭГ-обследование. Все испытуемые дали информированное согласие на участие в экспериментах. Исследование получило одобрение локального этического комитета по вопросам биомедицинских исследований НИЦ “Курчатовский институт”.
Магнитоэнцефалографические (МЭГ) измерения проводились в НИЦ “Курчатовский институт” внутри экранирующей комнаты с виброзащищенным фундаментом [5]. Стены комнаты толщиной 5 см сделаны из чистого алюминия и подавляют в 300 раз помехи на промышленной частоте 50 Гц. Использовался сверхпроводящий гелиевый 7-канальный градиентометр второго порядка [6]. Приемные витки измерительных катушек площадью 2 кв. см, расположенные по вершинам и в центре правильного шестиугольника радиусом 3 см, регистрировали нормальную компоненту магнитного поля, перпендикулярную плоскости петель. Высота приемных катушек (база) – 5 см, на них намотаны приемные и компенсирующие витки. Область измерения показана на рис. 1. Испытуемый лежал лицом вниз, упираясь лбом в регулируемый по высоте упор. Снизу, в полуметре от лица, устанавливался круглый светодиод диаметром 5 мм. Манипулятор с 6 степенями свободы подводил МЭГ криостат к затылку и обеспечивал точность позиционирования. Приоритетом при измерениях считались комфортные условия испытуемого. Связь была голосовой – испытуемый по команде открывал или закрывал глаза. Видеоконтроль позволял следить за ходом эксперимента.
Рис. 1.
Вид магнитометра расположенного над затылком испытуемого. На схеме цифрами обозначены положения измерительных катушек.
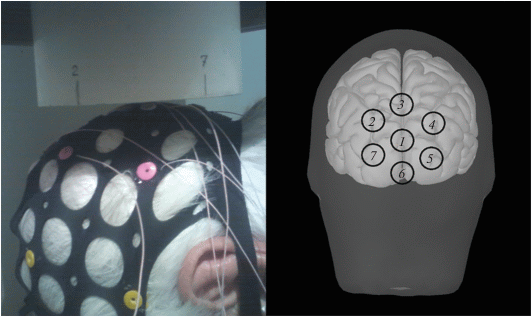
Вспышки светодиода формировались файлами мультимедийного проигрывателя персонального компьютера с использованием дополнительного усилителя мощности, если необходимо. Импульсы на несущей частоте 11 025 Гц были упакованы в запись в формате wav (waveform audio file format), образуя пачки со скважностью 1 Гц, повторяющиеся с частотами 10 и 13 Гц. Интенсивность света была на уровне индикаторов функционирования бытовых приборов, конкретный уровень подбирался, чтобы быть удобным для каждого испытуемого. Общая длительность стимуляции не превышала 5 мин.
Аналого-цифровое преобразование МЭГ-сигналов проводилось на частоте 1 кГц. МЭГ-данные были доступны для текущего наблюдения сигналов, фильтрации и демонстрации фурье-преобразования. Управление прибором и запись МЭГ проводилась электроникой российской фирмы “Криотон”, изготовившей магнитометр [6]. Как вспомогательный режим, совместно с МЭГ использовался электроэнцефалограф NVX-52 российской фирмы “Neurobotics” для синхронной записи 15 каналов ЭЭГ по схеме 10–20 с затылочной части головы, одного канала ЭКГ с центра грудины и одного канала регистрации управляющих импульсов для синхронизации МЭГ и ЭЭГ данных при последующей обработке. Полученные данные МЭГ и ЭЭГ доступны для всех традиционных математических методов обработки, в том числе и решения обратной задачи МЭГ программой MRIAN [7].
РЕЗУЛЬТАТЫ
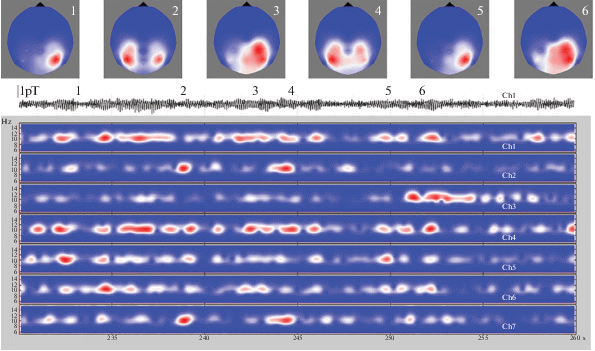
На рис. 2 показаны магнитные сигналы в семи каналах магнитометра в виде амплитудно-частотного рельефа, меняющегося со временем. Цветом отражена магнитуда Morlet-разложения в пакете BrainStorm [8]. МЭГ данные были предварительно обработаны многооконной полосовой фильтрацией для выделения откликов на частоту стимула и его целочисленных гармоник. Для 10-герцовой стимуляции задавался диапазон 5–15 Гц. Для сравнения, приведен и сам сигнал в канале № 1 (его положение показано на рис. 1), имеющий характерный вид спонтанного ритма. Принимаемый сигнал постоянно сильно меняется по амплитуде в каждом из каналов. В течение всей записи идет внешняя непрерывная стимуляция на частоте собственного ритма испытуемого и неустойчивость амплитуды говорит о том, что связь внешнего воздействия и генерируемых мозгом сигналов в текущем времени очень неустойчива. Это не добавочный шум. Возможно, что усредненные величины, замазав вариации, и дадут аналитическую связь между стимулом и откликом, но нас интересует динамика процесса.
Рис. 2.
Зависимость от времени магнитного сигнала в семи каналах магнитометра. Время указано в секундах. Текущая амплитуда задана цветом, по вертикали – частота. Красный цвет соответствует максимальной амплитуде. Идет непрерывная зрительная стимуляция на частоте 10 Гц, соответствующей собственному ритму испытуемого. Амплитуда сигнала сильно изменяется со временем во всех каналах. Для первого канала черным цветом показан вид самого сигнала, указан масштаб амплитуды. Для моментов времени, обозначенных цифрами 1–6, построены карты распределения амплитуды магнитного поля в затылочной части головы. Эти распределения сильно отличаются, хотя и встречаются похожие.
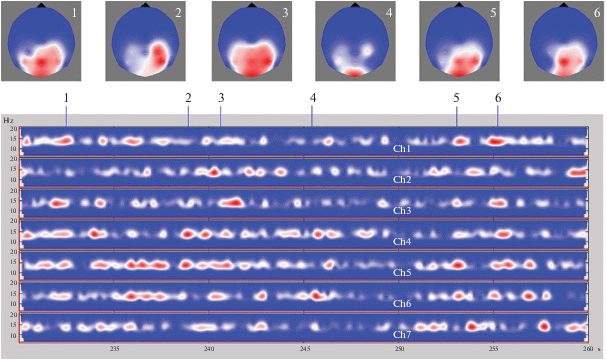
Постоянные вариации амплитуды сигнала на частоте 10 Гц могут быть следствием невыясненных до сих пор механизмов связи “резонансных” внешних стимулов с собственной ритмикой мозга, что отмечено во введении. Поэтому аналогичные измерения были проведены на частоте стимуляции 13 Гц, заметно отличающейся от частоты собственного ритма испытуемого. Результаты приведены на рис. 3. Мы видим ту же самую картину “рваного ритма”. Амплитуда принимаемого сигнала постоянно меняется от максимально возможной при естественной ритмике до практически нулевой, причем эти изменения идут почти независимо в разных каналах.
Рис. 3.
То же, что и на рис. 2, только зрительная стимуляция идет на частоте 13 Гц, отличающейся от собственного ритма.
С помощью пакета BrainStorm вычислялись карты распределения амплитуды магнитного поля в затылочной части головы. Оказалось, что карты поля сильно различаются в разные моменты времени. Бывают и похожие рельефы: например, показанные на рис. 2 моменты 1 и 5, или моменты 3 и 6. Иногда наблюдаются характерные распределения амплитуды, изображенные на картах 2 и 4 на рис. 2. Так выглядит магнитное поле от источника, расположенного прямо под центром системы катушек, то есть в первичной зрительной коре мозга. Считается, что именно сюда направляются сигналы от сетчатки глаза. Однако многочисленные другие встречающиеся варианты распределения магнитного поля указывают на электрическую активность ансамблей нейронов, расположенных вдали от первичной зрительной коры, причем, при отсутствии сравнимых по амплитуде сигналов, идущих из первичной зрительной коры. Можно провести различные усредняющие процедуры и определить зоны преимущественного воздействия внешней ритмической зрительной стимуляции на мозг, однако при усреднении динамика процессов, связывающих различные участки коры, будет утрачена. Мы же считаем существенным выделение характерных временных промежутков длительностью около 700 мс, которые дробят активность коры в условиях непрерывной стимуляции на периоды, в течение которых осуществляются разные режимы работы мозга.
Здесь приведены данные для одного испытуемого. У других испытуемых поведение аналогичное. Точно также выглядят сигналы, полученные на 306-канальном магнитометре в экспериментах с “частотной меткой” [4], но в том эксперименте решалась другая задача.
ОБСУЖДЕНИЕ
Мы видим, что в условиях непрерывного и стационарного внешнего воздействия на органы чувств (глаза) магнитные сигналы, генерируемые мозгом, ни в коей мере не стационарны и их источники не имеют четко сосредоточенного расположения в коре. В то же время, многочисленные исследования с помощью ЭЭГ безусловно указывают, что самые сильные отклики на зрительную стимуляцию регистрируются на затылочных электродах. При этом следует помнить, что усредняющие свойства покрывающих полушария мозга слоев (спинномозговая жидкость, череп, скальп) и изрезанный рельеф поверхности коры, позволяют с помощью ЭЭГ определять лишь очень размытую область в задней части головы, где располагаются активные популяции нейронов. Все они генерируют сигнал на затылочных электродах, причем вклады от разных групп сильно перемешиваются. Из магнитных же измерений видно, что реакция на вспышки света как бы постоянно “разбрасывается” по разным участкам этой достаточно большой части мозга, а в первичной зрительной коре, куда преимущественно поступают потоки нервных импульсов от глаз, реакция временами полностью отсутствует. То, куда направлены активирующие потоки импульсов, вероятно, зависит от текущих представлений внутренней системы мозга о сущности стимула. На этом основании она принимает решение.
Столь нестабильная реакция мозга на рутинно предлагаемые ему идентичные стимулы отнюдь не исключение, а скорее наоборот – правило. Это лучше проявляется именно в магнитных измерениях, как при моторных действиях, так и при восприятии повторяющихся слов [9, 10]. Что может быть причиной такой неустойчивости? Непрерывная оптическая стимуляция является просто некоторым устойчивым условием внешней среды, как температура воздуха, удобство позы или ожидание будущей команды. Нет никаких оснований считать, что внутренняя система мозга все время занята именно этими вспышками. То же самое относится к стандартным нейрофизиологическим экспериментам, в которых предлагаются повторяющиеся идентичные стимулы. Рассмотрение таких ситуаций не только академический вопрос – целое направление создания автономных агентов, принимающих самостоятельные решения, зависит от того, как будут организованы их системы внутреннего управления. Моделью для таких исследований служат простейшие животные [11]. Анализ их поведения приводит к выводу, что в каждый момент времени даже относительно простые организмы, такие как насекомые, используют сразу несколько режимов работы или наборов правил. Они конкурируют внутри их управляющей системы, и лишь победитель осуществляет собственную подпрограмму действий. В оригинале: “There is parallel activation of several rules at any given time, and these rules compete to determine behavior” [12]. Похоже, это относится и к человеку [13]. Нестабильность, которая постоянно наблюдается в периоды, когда не осуществляются никакие действия, однако человек готов выполнить то или иное задание, скорее всего, является следствием “конкурентной борьбы” различных, но вполне стереотипных режимов поведения. Число их невелико, вероятно, менее 10. Можно сказать, что в мозге непрерывно работает система принятия решений, поддерживающая готовность к выполнению задания одним из нескольких стандартных способов. Наиболее вероятным претендентом на роль системы, организующей внимание и готовящей выполнение задания, являются ансамбли корковых и таламических нейронов, синхронно колеблющихся в диапазоне альфа-ритма [14]. МЭГ дает возможность детально следить за поведением этой системы.
ЗАКЛЮЧЕНИЕ
Магнитные измерения позволяют вплотную подойти к задаче исследования системы принятия решений в мозге человека. Для этого есть все необходимые условия. На данный момент лишь магнитная энцефалография позволяет работать с характерными временами и пространственным разрешением, необходимыми для изучения системы принятия решений испытуемым, находящимся в рабочем контакте с исследователем.
Исследование частично поддержано РФФИ (проект № 18-00-00634 (К) (18-00-00575) Введенский В.Л.; проект № 18-00-00634 (К) (18-00-00513) Бондарев Д.П., Гребенкин А.П., Наурзаков С.П., Поликарпов М.А.).
Список литературы
Федотчев А.И. Резонансные явления в ЭЭГ человека и адаптация ЦНС к ритмической стимуляции. Дис. … д-ра. биол. наук. Москва: МГУ, 1997. 282 с.
Keitel C., Quigley C., Ruhnau P. // J. Neurosci. 2014. V. 34. № 31. Art. № 10137.
Mathewson K.E., Prudhomme C., Fabiani M. et al. // J. Cogn. Neurosci. 2012. V. 24. P. 2321.
Parkkonen L., Andersson J., Hämäläinen M., Hari R. // PNAS. 2018. V. 105. № 51. Art. № 20500.
Polikarpov M.A., Yurenya A.Y., Naurzakov S.P. et al. // J. Magn. Magn. Mater. 2017. V. 427. P. 139.
Maslennikov Y.V. // Phys. Proc. 2012. V. 36. P. 88.
Устинин М.Н., Сычев В.В., Линас Р.Р. // Мат. биол. и биоинф. 2013. № 8. С. 691.
Tadel F., Baillet S., Mosher J.C. et al. // Comput. Intel. Neurosci. 2011. Art. № 879716.
Введенский В.Л., Коршаков А.В. // Сб. науч. тр. XV Всерос. науч.-тех. конф. “Нейроинформатика-2013”. Часть 1. (Москва, 2013). С. 43.
Vvedensky V.L. // Neurosci. Lett. 2014. V. 569. P. 94.
Непомнящих В.А. // В кн. Синергетика и психология: Тексты. Выпуск 3: Когнитивные процессы. М.: Когито-Центр, 2004. С. 197.
Inglis I.R. // Behaviour. 2000. V. 137. № 12. P. 1567.
Vvedensky V.L., Prokofyev A.O. // Neur. Comput. 2016. V. 28. P. 286.
Bollimunta A., Mo J., Schroeder C.E., Ding M. // J. Neurosci. 2011. V. 31. № 13. P. 4935.
Дополнительные материалы отсутствуют.
Инструменты
Известия РАН. Серия физическая