

Журнал высшей нервной деятельности им. И.П. Павлова, 2023, T. 73, № 2, стр. 271-288
Изучение способности к исследовательской деятельности у низших обезьян
А. Е. Аникаев 1, *, В. Г. Чалян 1, Н. В. Мейшвили 1, Е. Н. Аникаева 1
1 ФГБНУ “Научно-исследовательский институт медицинской приматологии”
Сочи, Россия
* E-mail: mg_anykey@mail.ru
Поступила в редакцию 12.08.2022
После доработки 01.11.2022
Принята к публикации 09.12.2022
- EDN: IFOMSV
- DOI: 10.31857/S004446772302003X
Аннотация
Проведено изучение уровня исследовательской способности у четырех видов низших обезьян: зеленых мартышек (Chlorocebus aethiops), макаков яванских (Macaca fascicularis), макаков резусов (Macaca mulatta) и павианов гамадрилов (Papio hamadryas). Оценка производилась по таким показателям, как уровень исследовательской активности, динамика исследовательской активности, разнообразие исследовательской деятельности. Обнаруженные результаты указывают на более высокий уровень исследуемых параметров у макаков яванских и павианов гамадрилов в сравнении с макаками резусами и зелеными мартышками.
ВВЕДЕНИЕ
Исследовательское поведение является одним из очевидных проявлений когнитивных способностей животных, отражением степени их активности во взаимоотношениях с внешней средой. Зоопсихологи рассматривают исследовательскую активность как стремление животных “передвигаться и осматривать окружающую среду, даже когда они не испытывают ни голода, ни жажды, ни полового возбуждения” (Шовен, 1972). Исследовательская активность запускается в основном при появлении нового предмета или звука, изменения среды. Нужно отличать исследовательское поведение, которое запускается изменением среды или ее освоением, от инстинктивного, запускаемого ключевыми стимулами. Выраженное исследовательское поведение, несомненно, можно рассматривать в качестве одного из важнейших предикторов эволюционного успеха такой группы животных, как приматы, высокая приспособленность которых в значительной степени определяется когнитивными способностями. (Berlyne, 1978; Keller et al., 2012). Традиционно используемым подходом при изучении когнитивных способностей приматов является сравнительный подход (Schmitt et al., 2012; Petit et al., 2015; Joly et al., 2016). Использование сравнительного подхода позволяет проследить основные направления эволюционного развития мозга и когнитивных способностей приматов. Сравнительные исследования когнитивных способностей также позволяют оценить вклад взаимоотношений со средой и выраженности исследовательского поведения в особенности когнитивных способностей различных видов приматов. Известно, что существует две основные версии возникновения уникальных среди остального животного мира когнитивных способностей приматов – экологическая и социальная. Согласно первой – “экологической” теории когнитивные способности различных видов приматов определяются прежде всего сложностью их отношений со средой, степенью экологической пластичности, характером рациона, сложностью местообитания. Установлено, в частности, что виды приматов с преобладанием фруктоядности, а также виды, характеризующиеся выраженной всеядностью, отличаются сравнительно более высокими когнитивными показателями по целому ряду тестов, в частности в тестах на новые объекты (Bergman, Kitchen, 2008), по сравнению с более специализированными по характеру питания видами. Согласно второй – “социальной” точке зрения, уровень когнитивных способностей приматов является функцией особенностей социального устройства каждого вида и связан прежде всего с численностью особей в группах. Предполагается, что необходимость выстраивания отношений с каждой из многочисленных особей в больших группах определяет сравнительно высокий уровень когнитивных способностей у данных видов.
Целью нашего исследования является сравнительное изучение исследовательского поведения у макаков резусов (Macaca mulatta), макаков яванских (Macaca fascicularis), мартышек зеленых (гриветы) (Chlorocebus aethiops) и павианов гамадрилов (Papio hamadryas). Павианы, макаки и мартышки являются представителями различных родов одного и того же семейства мартышковых обезьян Cercopithecoidea – обитателей тропических регионов Старого Света. Все они характеризуются сходными физиологическими и биохимическими показателями, но имеют существенные различия в биологии, взаимоотношениях со средой, образе жизни и социальном устройстве. Можно предположить, что эти различия могут быть связаны с уровнем развития исследовательского поведения у представителей данных видов. Следует отметить несопоставимость количества существующих когнитивных исследований павианов, макаков и мартышек. Наиболее исследованными являются когнитивные способности макаков резусов и макаков яванских, что обусловлено широтой их использования в качестве лабораторных приматов. При этом когнитивные способности зеленых мартышек, также часто используемых в качестве лабораторных приматов, сравнительно мало исследованы. Существуют только единичные работы, посвященные исследованию когнитивных способностей павианов гамадрилов (Marsh et al., 2015).
Работы, посвященные изучению исследовательского поведения, как, впрочем, в целом в зоопсихологии и этологии, носят стихийный и хаотичный характер, что затрудняет системное изложение полученных в данной области результатов. Значительная часть работ посвящена изучению ориентировочно-исследовательского поведения у крыс и мышей (Belzung, 1999; Cowan, 1976). Достаточно широко представлены исследования игрового и исследовательского поведения у собак (Rossi et al., 2018; Siwak et al., 2001). Однако, несомненно, лучший объект для изучения исследовательского поведения представляют собой приматы, потому что у них хорошо развиты кисти рук. Всем известна привычка этих животных осматривать, облизывать и ощупывать со всех сторон все необычные предметы, которые попадают им в руки.
Существует ряд классических работ, выполненных в данном направлении. Так, Велькер (Welker, 1956b), работая с шимпанзе, установил, что при предъявлении им последовательно одного и того же объекта время, затрачиваемое на их осмотр, постепенно уменьшается. При этом он отмечал, что немаловажным фактором для шимпанзе является сложность предъявляемых им предметов. К подобному выводу пришел Войтонис (Войтонис, 1949), исследуя павианов и макаков. Он обнаружил, что предмет сначала привлекает животных, а затем интерес к нему постепенно ослабевает.
Также Велькер обнаружил зависимость уровня исследовательской активности от возраста, указав на различие в технике и продолжительности изучения новых объектов (Welker, 1956a). В других работах с молодыми шимпанзе (Menzel et al., 1961) было показано, что обезьяны более активно контактируют с новыми объектами, чем с уже известными объектами. На значение фактора “новизны” объекта указывают многие авторы (Loizos, 2017).
Ренш (Rensch, 1957) осуществил масштабное исследование по предпочтению цветов и форм объектов у трех видов обезьян: шимпанзе, зеленых мартышек и капуцина, – и пришел к выводу, что, в целом, предпочтение цветов по сравнению с серым, цветовых комбинаций и геометрических узоров у обезьян зависит от факторов, соответствующих эстетическим компонентам, действующим у людей.
В работе, посвященной изучению исследовательской деятельности макаков резусов (Harlow et al., 1956), было показано, что количество и эффективность манипуляций возрастают с возрастом и практикой. Исследователи предполагают, что манипуляторное поведение является самоподдерживающимся и не зависит от внутренних побуждений, таких как голод или жажда, или их систем стимулов, и не вытекает из них. При этом исследования японских обезьян в условиях естественного существования (Menzel, 1966) показали, что спонтанные манипуляции проявлялись менее часто и были менее стойкими, чем у лабораторных приматов. Подавляющее большинство действий осуществлялись молодыми особями. Доминирующие способы реакции на новые объекты носили осторожный характер.
Авторы исследования половых различий в реакции на детские игрушки у зеленых мартышек (Alexander, Hines, 2002) предполагают, что предпочтения к объектам, дифференцированным по половому признаку, возникли на раннем этапе эволюции человека, до появления отдельной линии гоминид. Это означает, что сексуально диморфные предпочтения в отношении характеристик (например, цвета, формы, движения) могли развиться в результате различного давления отбора, основанного на различных поведенческих ролях мужчин и женщин, и что эволюционные предпочтения в отношении характеристик объекта могут способствовать нынешним предпочтениям сексуально диморфных игрушек у детей. К схожим выводам пришли исследователи, изучавшие реакции на детские игрушки у макаков резусов (Hassett et al., 2008). Они отмечают сходство с человеческими данными и считают, что такие предпочтения могут развиваться без явной гендерной социализации. Авторы предлагают гипотезу о том, что предпочтения игрушек отражают гормонально обусловленные поведенческие и когнитивные предубеждения, которые социальные процессы превращают в половые различия, наблюдаемые у обезьян и людей.
Масштабное изучение исследовательского поведения макаков резусов разного пола и выращенных в разных условиях (нормальной социальной среде, в среде с различным уровнем социальной и пространственной депривации) (Sackett, 1972) показало следующее. Животные, выращенные в более богатой социальной и пространственной среде, демонстрировали более высокий уровень исследовательского поведения, однако влияние воспитания в условиях депривации на исследовательское поведение в значительной степени зависело от пола. После выращивания в условиях частичной или полной изоляции самки были намного более активными, чем самцы. При этом исследование влияния обогащения среды новыми объектами на поведение пожилых макаков резусов (Line et al., 1991) не показало никаких изменений в поведении обезьян. Различия в уровне исследовательской активности у особей разного пола и разной социальной структуры обнаруживаются и в других исследованиях (Santillán-Doherty et al., 2010).
Исследования распознавания визуальных объектов у макаков резусов и человека (Rajalingham et al., 2015) показывают, что обезьяны и люди статистически неотличимы в большом количестве задач распознавания визуальных объектов базового уровня, предполагая, что макаки резусы и люди могут иметь общее представление о нейронной “форме”, которое непосредственно лежит в основе восприятия объекта.
Также обнаруживается определённая взаимосвязь между уровнем исследовательской деятельности и темпераментом у макаков резусов (Coleman et al., 2005).
Немалый интерес представляет масштабное сравнительное исследование 74 видов обезьян на предмет разнообразия манипуляций с объектом (Tia et al., 2018). Обезьяны были разделены на три условные группы: 1) лемуры, мартышки, паукообразные обезьяны и гульманы; 2) обезьяны Старого Света, кроме гульманов; 3) капуцины и человекообразные обезьяны. В целом исследование показало преобладание количества и качества манипуляций у обезьян второй и третьей группы над обезьянами первой группы.
Различия в уровне исследовательской активности у разных видов приматов подтверждаются сравнительным исследованием трех видов приматов: галаго, лори и дурукули (Ehrlich, 1970). Галаго проводили больше всего времени в контакте с предметами. Дурукули и лори были менее восприимчивы к тестовым объектам, но, в то время как лори активно исследовали пространственные особенности новой среды, дурукули оставались неактивными. Для всех видов: наибольшая реакция на новые объекты произошла в течение первых пяти минут; начальные контакты объекта производились через обонятельные контакты, они были краткими и сопровождались более продолжительными контактами ртом и руками.
Обобщая представленную информацию, можно привести ряд вполне однозначных заключений. Исследовательское поведение свойственно различным видам приматов, но уровень исследовательской активности и разнообразие данной активности достаточно широко варьирует в зависимости от видовой принадлежности, пола, возраста, условий содержания и ряда других факторов.
Исходя из обнаруженных выше данных можно заключить, что изучение исследовательского поведения у различных видов животных и, в частности, у приматов представляло немалый интерес для ученых в начале второй половины прошлого столетия и постепенно шло на спад. Однако следует отметить, что сегодня интерес к данной проблеме возрождается, к сожалению, преимущественно среди зарубежных зоопсихологов, этологов и нейробиологов.
МЕТОДИКА
В качестве объекта исследования были использованы половозрелые самцы следующих четырех видов: зеленые мартышки (Chlorocebus aethiops) (n = 11), макаки резусы (Macaca mulatta) (n = 23), макаки яванские (Macaca fascicularis) (n = 14) и павианы гамадрилы (Papio hamadryas) (n = 57).
Все животные были рождены и содержались до начала эксперимента в вольерах ФГБНУ “НИИ Медицинской приматологии”, где используется методика содержания обезьян, предполагающая минимальное вмешательство персонала в их жизнь. Фактически, с точки зрения опыта контакта с людьми, содержащиеся в вольерах Адлерского центра обезьяны соответствуют диким обезьянам, привезенным из мест естественного обитания.
Исследование начиналось с отсаживания животных из родных групп и помещения их в клетки индивидуального содержания (размер клеток варьировал в зависимости от вида: 66 × 67 × 90 см для макаков и мартышек и 82 × 82 × 90 см для павианов), где первоначально они проходили этап предварительного привыкания к индивидуальному содержанию и постоянному присутствию и близости человека. Никакого дополнительного приручения и оценки состояния животных не осуществлялось. После двухнедельного этапа привыкания обезьян к условиям индивидуального содержания начиналось тестирование животных.
Для определения уровня способности к исследовательской деятельности была применена методика “Реакция на новый объект” (Аникаев и др., 2021). Суть методики заключается в следующем: животному предоставляется объект (объект помещался рядом с клеткой в зоне досягаемости), ключевой характеристикой которого является новизна. В данном исследовании в качестве объекта использовался пластиковый разноцветный куб (15 × 15 × 15 см). На взаимодействие с объектом животному отводится определенный промежуток времени, в данном случае 300 с. Наблюдения за ходом выполнения теста осуществляются с помощью видеокамеры. Фиксируется время и вариант контакта с объектом. В случае прекращения контакта с объектом фиксируется время и отмечается “оставил”.
Для определения уровня способности к исследовательской деятельности использовались следующие показатели.
Активность. Выражается суммой временных отрезков контактов животных с объектом.
Динамика активности. Оценивалась путем подсчета временных отрезков контактов животных с объектом в каждом из 10 интервалов по 30 с.
Разнообразие. Выражается в количестве вариантов контакта с объектом.
Характер разнообразия. Определялся по соотношению количества контактов каждой категории. Экспериментальным путем нами были выделены следующие виды контактов животных с объектом, разделенные на четыре категории: тактильные (касается, переворачивает, держит, затягивает, трясет, ковыряет, отталкивает, двигает, выбрасывает), оральные (грызет, облизывает), визуальные (рассматривает (фиксировались случаи, при которых животные направляли взгляд на объект без физического контакта с ним)) и обонятельные (обнюхивает).
Статистический анализ полученных данных осуществлялся путем вычисления стандартной ошибки средней (SEM), критерия Манна–Уитни (U), коэффициента корреляции Спирмена (R), стандартного отклонения (σ) и коэффициента прироста (Kпр) пакетами программного обеспечения Microsoft Offiсe Excel 2010 и Statistica 7.0.
Все эксперименты проводились в соответствии с международными правилами по использованию лабораторных животных (National Research Council, 2010). На проведение исследования было получено разрешение Комиссии по этике ФГБНУ “НИИ Медицинской приматологии”.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Исследовательская активность и разнообразие исследовательской деятельности
Полученные результаты (табл. 1) указывают на следующее. Наиболее высокий уровень активности и разнообразия исследовательской деятельности продемонстрировала группа макаков яванских, самые низкие показатели наблюдались в группе зеленых мартышек, при этом разница в уровне активности и разнообразия между ними достоверно составила соответственно 48.8% (критерий Манна–Уитни, U = 16.0, p ≤ 0.05) и 41.3% (критерий Манна–Уитни, U = 15.5, p ≤ 0.05). В группе павианов гамадрилов фактические значения уровня активности и разнообразия исследовательской деятельности ниже, чем у макаков яванских, соответственно на 16.8 и 5.3%, однако отсутствие статистически значимых различий (критерий Манна–Уитни, U = 111.0 и U = 325.0, p ≥ 0.05) позволяет предположить сопоставимость уровня исследуемых параметров у этих двух видов. При этом различия между павианами гамадрилами и зелеными мартышками достоверно составили 32.0% (критерий Манна–Уитни, U = = 111.5, p ≤ 0.05) по уровню активности и 35.9% (критерий Манна–Уитни, U = 75.0, p ≤ 0.05) по уровню разнообразия. Результаты, полученные в группе макаков резусов, указывают на их промежуточное положение как в уровне активности, так и в уровне разнообразия исследовательской деятельности. Так, уровень активности в исследуемой группе данного вида (достоверно во всех случаях) на 37.5% (критерий Манна–Уитни, U = 64.0, p ≤ 0.05) ниже, чем у макаков яванских, на 20.7% (критерий Манна–Уитни, U = 48.0, p ≤ 0.05) ниже, чем у павианов гамадрилов, но на 11.3% (критерий Манна–Уитни, U = 57.0, p ≤ 0.05) выше, чем у зеленых мартышек. При этом значения уровня разнообразия исследовательской деятельности в группе макаков резусов сопоставимы с таковыми у макаков яванских и павианов гамадрилов (15.3% (критерий Манна–Уитни, U = 107.5, p ≥ 0.05) – разница с макаками яванскими и 10.0% (критерий Манна–Уитни, U = 523.5, p ≥ 0.05) – разница с павианами гамадрилами), но достоверно выше – на 25.9% (критерий Манна–Уитни, U = 48.0, p ≤ 0.05), чем в группе зеленых мартышек.
Таблица 1.
Значения показателей уровня способности к исследовательской деятельности у изучаемых видов обезьян Table 1. Values of indicators of the level of ability for research activity in the studied species of monkeys
| Вид | Средний уровень активности, % | Средний уровень разнообразия, % |
|---|---|---|
| Зеленые мартышки | 10.9 ± 5.7 | 15.2 ± 3.1 |
| Макаки яванские | 59.7 ± 10.2 | 56.4 ± 7.4 |
| Макаки резусы | 22.2 ± 5.3 | 41.1 ± 5.5 |
| Павианы гамадрилы | 42.9 ± 4.5 | 51.1 ± 3.4 |
Также следует отметить, что для групп макаков яванских, макаков резусов и павианов гамадрилов обнаруживается высокий уровень корреляции между уровнем исследовательской активности и уровнем разнообразия исследовательской деятельности: R = 0.81, R = 0.88 и R = 0.68 соответственно. При этом зависимость между этими показателями в группе зеленых мартышек практически отсутствует (R = 0.05).
Динамика исследовательской активности
Анализ динамики исследовательской активности (рис. 1) показывает следующее. Во всех исследуемых группах животных наблюдается сходная динамика исследовательской активности: в первый интервал (30 секунд) обнаруживается максимально высокий для каждого исследуемого вида уровень исследовательской активности, затем происходит постепенный спад активности. Данный факт подтверждается значениями коэффициентов корреляции между динамическими рядами, полученными при анализе динамики активности исследуемых видов. Максимальные коэффициенты получены между динамическими рядами зеленых мартышек и макаков резусов (R = 0.96), зеленых мартышек и павианов гамадрилов (R = 0.90) и павианов гамадрилов и макаков резусов (R = 0.86). При этом между динамическими рядами макаков яванских и макаков резусов обнаружен самый низкий уровень корреляции (R = 0.62). Значения коэффициентов между динамическими рядами макаков яванских и павианов гамадрилов, а также макаков яванских и зеленых мартышек занимают промежуточное положение и соответственно составили 0.83 и 0.74. То есть динамика активности исследуемых групп зеленых мартышек, макаков резусов и павианов гамадрилов в большей степени совпадает между этими группами и в некоторой степени отличается от динамики активности макаков яванских. Для уточнения вышеуказанных данных был произведен сравнительный анализ значений, полученных с 1-го по 5-й интервал, и значений, полученных с 6-го по 10-й интервал.
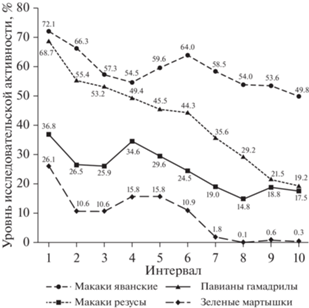
Так, среднее значение динамики активности с 1-го по 5-й интервал в группе макаков яванских составило 61.9 ± 3.2%, а с 6-го по 10-й интервал – 56.0 ± 2.4%, соответственно разница между средними составила 6.0% (критерий Манна–Уитни, U = 5.0, p ≥ 0.05). Среднее значение динамики активности с 1-го по 5-й интервал в группе павианов гамадрилов составило 54.4 ± 3.9%, а с 6-го по 10-й интервал – 30.0 ± 4.6%, соответственно разница между средними составила 24.5% (критерий Манна–Уитни, U = 0.0, p ≤ 0.05). Среднее значение динамики активности с 1-го по 5-й интервал в группе макаков резусов составило 30.7 ± 2.2%, а с 6-го по 10-й интервал – 18.9 ± ± 1.6%, соответственно разница между средними составила 11.8% (критерий Манна–Уитни, U = 0.0, p ≤ 0.05). Среднее значение динамики активности с 1-го по 5-й интервал в группе зеленых мартышек составило 15.8 ± ± 2.8%, а с 6-го по 10-й интервал – 2.7 ± 2.1%, соответственно разница между средними составила 13.0% (критерий Манна–Уитни, U = = 2.0, p ≤ 0.05).
Полученные результаты указывают на то, что, в отличие от исследуемой группы макаков яванских, в исследуемых группах зеленых мартышек, макаков резусов и павианов гамадрилов спад активности, выраженный в различии между средними значениями, полученными с 1-го по 5-й интервал и с 6-го по 10-й интервал, носит статистически значимый характер. То есть в исследуемой группе макаков яванских по сравнению с обезьянами других видов наблюдается самый незначительный спад исследовательской активности. Что, в свою очередь, подтверждается значениями коэффициентов темпа роста, которые составили: –4.0 для макаков яванских, –7.9 для макаков резусов, –13.2 для павианов гамадрилов и –39.1 для зеленых мартышек.
Таким образом, можно заключить, что на общем фоне спада исследовательской активности в исследуемых группах обезьян наименьшая интенсивность снижения наблюдается в группе макаков яванских, наибольшая – в группе зеленых мартышек, а макаки резусы и павианы гамадрилы занимают промежуточное положение.
Характер разнообразия исследовательской деятельности
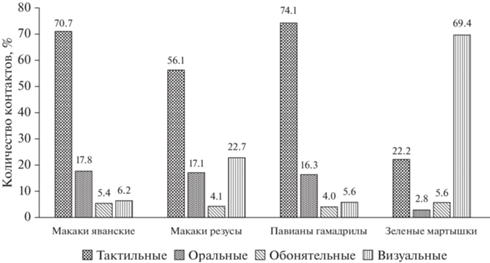
Анализ распределения категорий контактов животных с объектом (рис. 2) показал следующее. Наибольшее сходство в характере распределения наблюдается между данными, полученными при анализе показателей в группе макаков яванских и павианов гамадрилов (коэффициент корреляции Спирмена R = 1.00). Для данных групп обнаружено следующее распределение контактов разных категорий: преобладающее количество контактов относится к категории “тактильные”, на втором месте, но с более низкими значениями, – категория “оральные”, на третьем месте – “визуальные”, и самые низкое количество контактов – в категории “обонятельные”. Схожее распределение наблюдается в группе макаков резусов: также преобладают “тактильные” контакты, но на втором месте “визуальные”, а “оральные” – на третьем, при этом “обонятельные” также представлены в наименьшем количестве. То есть в целом в группе макаков резусов распределение сопоставимо с таковым у макаков яванских и павианов гамадрилов, что подтверждают коэффициенты корреляции, которые составили по 0.94. Распределение контактов разных категорий у зеленых мартышек существенно отличается от описанных выше: преобладающее количество контактов относится к категории “визуальные”, на втором месте, но с более низкими значениями, – категория “тактильные”, на третьем – “обонятельные”, и “оральные” – на четвертом месте. Корреляция между распределениями контактов разных категорий низкая обратная при сравнении с макаками яванскими (R = –0.15), также низкая обратная при сравнении с павианами гамадрилами (R = –0.13) и прямая слабая при сравнении с маками резусами (R = 0.19).
Рис. 2.
Распределение категорий контактов животных с объектом. Fig. 2. Distribution of categories of animal contact with the object.
Детальный внутривидовой анализ характера разнообразия исследовательской деятельности представлен в табл. 2. Так, в исследуемой группе макаков яванских обнаруживается следующее. Тактильные типы контактов с объектом достоверно преобладают над оральными, обонятельными и визуальными взаимодействиями. При этом между этими тремя видами контактов статистически значимые различия отсутствуют. То есть макаки яванские отдают явное предпочтение тактильному взаимодействию со средой.
Таблица 2.
Внутривидовой анализ характера разнообразия исследовательской деятельности Table 2. Intraspecific analysis of the nature of the diversity of research activities
| Вид | Сравниваемые категории контактов | Разница | Достоверность (критерий Манна–Уитни (U)) |
|---|---|---|---|
| Макаки яванские | Тактильные/оральные | 9.8 | 31.0* |
| Тактильные/обонятельные | 12.2 | 23.5* | |
| Тактильные/визуальные | 12.0 | 21.0* | |
| Оральные/обонятельные | 2.3 | 47.0 | |
| Оральные/визуальные | 2.2 | 50.5 | |
| Обонятельные/визуальные | 0.2 | 81.5 | |
| Макаки резусы | Тактильные/оральные | 4.6 | 7.5* |
| Тактильные/обонятельные | 6.1 | 7.5* | |
| Тактильные/визуальные | 3.9 | 16.5 | |
| Оральные/обонятельные | 1.5 | 18.0* | |
| Оральные/визуальные | 0.7 | 28.5 | |
| Обонятельные/визуальные | 2.2 | 31.0 | |
| Павианы гамадрилы | Тактильные/оральные | 6.9 | 558.0* |
| Тактильные/обонятельные | 8.4 | 274.0* | |
| Тактильные/визуальные | 8.2 | 308.5* | |
| Оральные/обонятельные | 1.5 | 1075.0* | |
| Оральные/визуальные | 1.3 | 1108.0* | |
| Обонятельные/визуальные | 0.2 | 1618.0 | |
| Зеленые мартышки | Тактильные/оральные | 0.6 | 43.0 |
| Тактильные/обонятельные | 0.5 | 47.5 | |
| Тактильные/визуальные | 1.5 | 24.5* | |
| Оральные/обонятельные | 0.1 | 55.0 | |
| Оральные/визуальные | 2.2 | 8.0* | |
| Обонятельные/визуальные | 2.1 | 10.5* |
В исследуемой группе макаков резусов картина не столь однозначная. Тактильные типы контактов также преобладают над другими типами взаимодействия с объектом, но с оральными и обонятельными полученные различия носят достоверный характер. Визуальные типы контактов преобладают над оральными и обонятельными, но полученные различия статистически незначимы. При этом оральные типы контактов достоверно преобладают над обонятельными. Таким образом, макаки резусы, как и макаки яванские, предпочитают тактильный вариант взаимодействия с объектом, но относительно чаще используют только визуальный контакт с объектом. И, так же как макаки яванские, практически не используют обонятельный тип контакта.
В исследуемой группе павианов гамадрилов получены следующие результаты. Тактильные типы контактов достоверно преобладают над тремя другими категориями взаимодействий с объектом. При этом оральные типы контактов также достоверно преобладают над обонятельными и визуальными. И только между обонятельными и визуальными контактами отсутствуют статистически значимые различия. То есть павианы гамадрилы, как и оба вида макаков, однозначно предпочитают тактильный тип взаимодействия с объектом, при этом на втором месте у данного вида – достоверно – оральные типы контактов. Визуальный и обонятельный контакт практически не используется.
В исследуемой группе зеленых мартышек наблюдается следующее. Над всеми типами контактов достоверно преобладают визуальные. На втором месте тактильные, но между этим типом контакта и оральными, а также обонятельными взаимодействиями, как и между двумя последними, достоверных различий не обнаружено. То есть зеленые мартышки предпочитали только рассматривать объект и практически не касались его.
Данные по анализу межвидовых различий в предпочтении определённых типов контактов с объектом (табл. 3) указывают на следующее. Макаки яванские демонстрируют достоверно большее количество тактильных взаимодействий, чем макаки резусы и зеленые мартышки, и недостоверно – чем павианы гамадрилы. Павианы гамадрилы демонстрируют достоверно большее количество тактильных контактов, чем зеленые мартышки, и недостоверно – чем макаки резусы. В свою очередь макаки резусы демонстрируют достоверно большее количество тактильных контактов, чем зеленые мартышки.
Таблица 3.
Межвидовой анализ характера разнообразия исследовательской деятельности Table 3. Cross-species analysis of the nature of the diversity of research activities
| Категории контактов | Сравниваемые виды | Разница | Достоверность (критерий Манна–Уитни (U)) |
|---|---|---|---|
| Тактильные | Макаки яванские/макаки резусы | 6.6 | 89.0* |
| Макаки яванские/павианы гамадрилы | 4.3 | 268.5 | |
| Макаки яванские/зеленые мартышки | 12.4 | 13.5* | |
| Макаки резусы/павианы гамадрилы | 2.3 | 507.5 | |
| Макаки резусы/зеленые мартышки | 5.8 | 54.0* | |
| Павианы гамадрилы/зеленые мартышки | 8.1 | 62.0* | |
| Оральные | Макаки яванские/макаки резусы | 1.3 | 113.0 |
| Макаки яванские/павианы гамадрилы | 1.4 | 145.5 | |
| Макаки яванские/зеленые мартышки | 3.2 | 24.0* | |
| Макаки резусы/павианы гамадрилы | 0.1 | 625.5 | |
| Макаки резусы/зеленые мартышки | 1.9 | 72.5* | |
| Павианы гамадрилы/зеленые мартышки | 1.9 | 143.5* | |
| Обонятельные | Макаки яванские/макаки резусы | 0.5 | 96.0 |
| Макаки яванские/павианы гамадрилы | 0.5 | 249.0 | |
| Макаки яванские/зеленые мартышки | 0.8 | 35.5* | |
| Макаки резусы/павианы гамадрилы | 0.0 | 604.0 | |
| Макаки резусы/зеленые мартышки | 0.3 | 109.0 | |
| Павианы гамадрилы/зеленые мартышки | 0.3 | 241.0 | |
| Визуальные | Макаки яванские/макаки резусы | 1.5 | 75.5* |
| Макаки яванские/павианы гамадрилы | 0.5 | 294.0 | |
| Макаки яванские/зеленые мартышки | 1.1 | 41.5 | |
| Макаки резусы/павианы гамадрилы | 2.0 | 238.0* | |
| Макаки резусы/зеленые мартышки | 0.4 | 105.5 | |
| Павианы гамадрилы/зеленые мартышки | 1.6 | 120.0* |
Достоверно меньшее количество оральных контактов, чем три других вида обезьян, демонстрируют зеленые мартышки. При этом между макаками яванскими, макаками резусами и павианами гамадрилами достоверные различия в количестве используемых оральных контактов отсутствуют.
По количеству обонятельных контактов преобладают макаки яванские, но достоверно – только над зелеными мартышками. Макаки резусы и павианы гамадрилы демонстрируют равное количество обонятельных контактов, большее (недостоверно), чем зеленые мартышки.
Самое большое количество визуальных контактов наблюдается в группе макаков резусов, достоверно больше, чем в группе павианов гамадрилов и макаков яванских. Зеленые мартышки демонстрируют (недостоверно) меньшее количество визуальных контактов, чем макаки резусы, но большее, чем макаки яванские и павианы гамадрилы, достоверно в отношении последних. При этом различия в количестве визуальных контактов, демонстрируемых макаками яванскими и павианами гамадрилами, носят статистически незначимый характер.
Индивидуальные различия
Распределение исследуемых обезьян в зависимости от уровня исследовательской активности представлено на рис. 3 (а). Во всех исследуемых группах наблюдается достаточно высокий уровень индивидуальных различий в уровне исследовательской активности.
Рис. 3.
Распределение исследуемых обезьян в зависимости от уровня исследовательской активности (а) и от уровня разнообразия исследовательской деятельности (б). Fig. 3. The distribution of the studied monkeys depending on the level of research activity (а) and on the level of diversity of research activity (б).
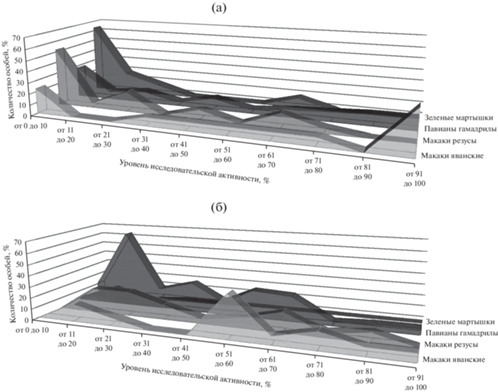
Наибольший разброс индивидуальных различий обнаруживается в группе макаков яванских: σ = 38.2, при этом максимальный уровень исследовательской активности составляет 100.0%, а минимальный 1.3%. Наибольшее количество особей макаков яванских относятся к категориям “от 91 до 100” активности (38.5%) и “от 0 до 10” активности (23.1%). Остальные особи равномерно (по 7.7%) распределились в интервале категорий “от 31 до 40” до “от 71 до 80”. То есть среди макаков яванских обнаружены как особи с очень высоким уровнем исследовательской активности, так и особи с крайне низким уровнем исследовательской активности, однако преимущественно распределение смещено в сторону высокого уровня исследовательской активности (медиана распределения M = 66.7%).
Исходя из значения стандартного отклонения σ = 33.9, следующие по степени индивидуальных различий в уровне исследовательской активности – павианы гамадрилы. Максимальный уровень исследовательской активности составляет 99.0%, минимальный – 0.0%. При этом в категории “от 0 до 10” самый высокий процент особей (28.1%). Остальные особи в относительно равном количестве (от 3.5% до 10.5%) распределились по другим категориям уровня активности (при этом представлены во всех категориях). То есть в группе павианов гамадрилов исследуемые животные распределяются относительно равномерно (медиана распределения M = 41.0%).
У макаков резусов стандартное отклонение составило σ = 25.4. При этом максимальный уровень исследовательской активности составляет 75.0%, минимальный – 0.3%. Преобладающее количество особей (52.2%) относятся к категории “от 0 до 10”. На втором месте (17.4%) категория “от 21 до 30” активности. Также макаки резусы в относительно равной степени (от 4.4 до 13.0%) представлены в категориях “от 11 до 20”, “от 41 до 50”, “от 61 до 70” и “от 71 до 80”. Таким образом, среди макаков резусов в основном обнаружены особи с крайне низким уровнем исследовательской активности, и в разной степени – средним уровнем исследовательской активности, но не обнаружены особи с максимальным уровнем исследовательской активности. Распределение выраженно смещено в сторону крайне низкого значения уровня исследовательской активности (медиана распределения M = 6.0%).
Самый низкий уровень индивидуальных различий (стандартное отклонение σ = 18.8) наблюдается в группе зеленых мартышек. При этом максимальный уровень исследовательской активности составляет 61.3%, минимальный – 0.0%. Максимальный процент (63.6%) особей представлен в категории “от 0 до 10”, на втором месте (18.2%) категория “от 11 до 20”, на третьем (9.1%) – “от 21 до 30”. Распределение еще более выражено, чем в группе макаков резусов, смещено в сторону крайне низкого значения уровня исследовательской активности (медиана распределения M = 0.7%).
Распределение исследуемых обезьян в зависимости от уровня разнообразия исследовательской деятельности представлено на рис. 3 (б). Индивидуальные различия в уровне разнообразия исследовательской деятельности менее выражены, чем индивидуальные различия в уровне исследовательской активности, однако также находятся на достаточно высоком уровне.
Относительно одинаковый уровень индивидуальных различий наблюдается в группе макаков яванских (стандартное отклонение σ = 27.8), в группе макаков резусов (стандартное отклонение σ = 26.5) и в группе павианов гамадрилов (стандартное отклонение σ = 25.4). При этом в группах макаков яванских и павианов гамадрилов есть представители с максимальным уровнем (100.0%) разнообразия исследовательской деятельности. Минимальный уровень данного показателя для макаков яванских составляет 11.1%, для павианов гамадрилов – 0.0%. При этом максимальный уровень разнообразия исследовательской деятельности в группе макаков резусов составил 88.9%, а минимальный уровень разнообразия исследовательской деятельности – 11.1%.
В группе макаков яванских самый высокий процент (38.5%) особей наблюдается в категории “от 51 до 60”. На втором месте (по 15.4%) категории “от 11 до 20” и “от 71 до 80”. И по 7.7% особей макаков яванских распределились по категориям “от 21 до 30”, “от 61 до 70”, “от 81 до 90” и “от 91 до 100”. То есть среди макаков яванских преимущественно обнаружены особи со средним уровнем разнообразия исследовательской деятельности, распределение в большей степени смещено в сторону высокого уровня исследовательской активности (медиана распределения M = 55.6%).
В группе павианов гамадрилов максимальный процент особей наблюдается в категориях “от 51 до 60” (24.6%) и “от 61 до 70” (22.8%) разнообразия деятельности. Остальные особи в разной степени (от 1.8 до 14.0% особей) представлены в других категориях (при этом представлены во всех категориях). Таким образом, среди павианов гамадрилов, так же как и в группе макаков яванских, преимущественно обнаружены особи со средним уровнем разнообразия исследовательской деятельности, однако в отличие от макаков яванских в группе павианов гамадрилов исследуемые животные распределяются более равномерно (медиана распределения M = 55.6%).
В группе макаков резусов максимальный процент (по 21.7%) особей наблюдается в категориях “от 11 до 20” и “от 21 до 30”. Остальные особи в разной степени (от 4.3 до 17.4% особей) представлены в категориях “от 31 до 40” до категории “от 81 до 90”. То есть среди макаков резусов преимущественно обнаружены особи с низким уровнем разнообразия исследовательской деятельности, распределение в большей степени смещено в сторону низкого уровня исследовательской активности (медиана распределения M = 33.3%).
Менее выраженный уровень индивидуальных различий наблюдается в группе зеленых мартышек (стандартное отклонение σ = 10.3). При этом максимальный уровень разнообразия исследовательской деятельности в группе зеленых мартышек составил 33.3%, а минимальный уровень разнообразия исследовательской деятельности – 0.0%.
В группе зеленых мартышек преобладающий процент (63.9%) особей обнаружен в категории “от 11 до 20”, остальные особи в относительно равной степени (от 9.1 до 18.2%) представлены в категориях “от 0 до 10”, “от 21 до 30” и “от 30 до 41”. Таким образом, среди зеленых мартышек в основном обнаружены особи с крайне низким уровнем разнообразия исследовательской деятельности. Распределение выраженно смещено в сторону низкого уровня исследовательской активности (медиана распределения M = 11.1%).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Анализ результатов проведенных исследований обнаруживает существенные межвидовые различия во всех использованных показателях исследовательской деятельности. Наихудшие результаты по всем показателям отмечены у зеленых мартышек. Обезьяны этого вида демонстрировали самый низкий уровень активности при исследовании новых объектов. В качестве основной формы исследовательской деятельности зеленые мартышки выбирали наиболее простую его форму – визуальное исследование. Частота использования различных форм исследовательской деятельности отрицательно коррелирует с показателями макаков яванских и павианов гамадрилов. При этом именно для зеленых мартышек был характерен минимальный разброс индивидуальных различий в показателях (в сторону низких значений).
Наиболее близкие к зеленым мартышкам результаты из 3 остальных исследованных видов обнаруживали макаки резусы. Исследовательская деятельность макаков резусов характеризовалась сравнительно невысоким уровнем активности, который, тем не менее, был выше, чем у зеленых мартышек. Следует отметить, что среди наиболее часто используемых форм исследовательской деятельности этих обезьян преобладали тактильные прикосновения и визуальное исследование. Имеется положительная, но невысокая корреляция между частотой различных форм исследовательской деятельности у макаков резусов и зеленых мартышек. При этом динамика спада исследовательской деятельности у макаков резусов значительно отличалась от таковой у зеленых мартышек и практически совпадала с динамикой спада активности у макаков яванских и павианов гамадрилов.
Представляется любопытным высокое совпадение большинства показателей исследовательской деятельности у макаков яванских и павианов гамадрилов. И у макаков яванских, и у павианов гамадрилов исследовательская деятельность максимально выражается тактильными прикосновениями к исследуемому предмету, а также оральными контактами, и в меньшей степени – визуальным исследованием и обнюхиванием. Имеется приближающаяся к единице положительная корреляция в частоте используемых обоими видами форм исследовательской деятельности. У макаков яванских и павианов гамадрилов обнаружены близкие и бóльшие по сравнению с макаками резусами и зелеными мартышками значения индивидуального разнообразия показателей исследовательской активности. При этом следует отметить, что именно макаки яванские обнаруживают самый высокий уровень исследовательской активности, а также самую слабую динамику спада этой активности. У павианов гамадрилов уровень исследовательской активности несколько ниже, чем у макаков яванских, но выше, чем у макаков резусов. Напротив, динамика спада исследовательской активности у павианов гамадрилов значительно выше, чем у макаков яванских, и практически совпадает с таковой у макаков резусов.
Таким образом, сравнение когнитивных способностей изучаемых видов приматов показывает, что по большинству критериев исследовательской деятельности 4 изучаемых вида разделились на 2 категории. К категории лидеров следует отнести макаков яванских и павианов гамадрилов, к категории аутсайдеров – макаков резусов и, особенно, зеленых мартышек. Макаки яванские и павианы гамадрилы демонстрировали высокий уровень исследовательской активности, у них была отмечена высокая корреляция используемых форм исследовательской деятельности, среди которых преобладали тактильные взаимодействия с исследуемыми объектами, а также сопоставимо низкие показатели индивидуального разнообразия исследовательской деятельности. Макаки резусы и зеленые мартышки также обнаруживают определенное сходство в исследовательской активности. Для обоих этих видов был характерен сравнительно низкий уровень исследовательской деятельности и низкий уровень показателей индивидуального разнообразия. Такое распределение в общем уровне исследовательской деятельности может быть объяснено с позиции общего уровня эволюционного развития данных видов животных. Согласно современным представлениям, зеленые мартышки занимают низшую ступень (из представленных видов), павианы гамадрилы высшую ступень, а макаки резусы и макаки яванские – промежуточное положение (Perelman P. et al., 2011). И противоречие в данном случае складывается в отношении макаков яванских, продемонстрировавших в нашем исследовании самые высокие значения исследовательской активности и разнообразия исследовательской деятельности.
Для объяснения обнаруженных различий между макаками яванскими и павианами гамадрилами, с одной стороны, и макаками резусами и зелеными мартышками, с другой, следует, по видимому, оценить 3 важнейшие характеристики изучаемых видов: 1) особенности строения мозга и его объем; 2) существующие “экологические” различия между этими видами; 3) особенности их социального устройства.
Проведенные ранее исследования показали, что когнитивные способности приматов в значительной степени связаны с абсолютными размерами мозга (Deaner, 2007). Имеются также сведения о существовании экологических коррелятов размеров неокортекса. Согласно гипотезе “экологического мозга” пространственные параметры, связанные со стратегией фуражирования и пищевыми ресурсами, определяют размер мозга и когнитивную сложность (Milton, 1988). В пользу этого предположения свидетельствует тот факт, что у дневных приматов размеры неокортекса больше по сравнению с приматами, ведущими ночной образ жизни, а также положительно коррелируют с таким, сугубо экологическим, показателем, как степень выраженности фруктоядности (Barton, 1996). Анализ такого близкого к исследовательской деятельности показателя, как манипуляционная активность, выполненный у 36 видов приматов, показал, что сложность этой формы поведения определяется 2 взаимосвязанными факторами: размерами мозга и степенью наземности (Helstab et al., 2016). Вместе с тем установлено, что у гаплориновых приматов размеры групп положительно коррелируют с объемом неокортекса, что рассматривается как аргумент в пользу “социальной” теории интеллекта (Dunbar, 1992). В соответствии с этой теорией объем мозга и когнитивная сложность напрямую зависят от социальной сложности. В частности, исследование показало, что у социально содержащихся в клетках и вольерах павианов анубисов, у тех особей, которые живут в составе более крупных групп, мозг более крупный и с большим количеством серого вещества, чем у особей из более мелких групп (Meguerditchian et al., 2021). Следует отметить, что существуют только немногочисленные исследования, направленные на выяснение возможности использования особенностей экологии приматов в качестве предиктора их когнитивных способностей (Schwartz, 2019). В частности, в исследовании при сравнении экологии и пространственной памяти двух видов южноамериканских приматов – Golden lion tamarin (Leontopithecus rosalia) и Wied’s marmoset (Callithrix kuhli) – было установлено, что у тамаринов площадь участка обитания больше и пространственная память лучше, чем у мармозеток (Platt et al., 1996).
Рассмотрение гипотезы о связи между структурой мозга и когнитивными способностями приматов применительно к 4 изучаемым нами видам обезьян свидетельствует о том, что павианы гамадрилы имеют в этом отношении очевидные преимущества перед 3 другими видами. Сравнение объема мозга 4 изучаемых видов приматов показывает, что павианы гамадрилы имеют сравнительно больший объем неокортекса, тогда как у макаков и мартышек объем неокортекса сопоставим. По всей видимости, данное обстоятельство можно рассматривать в качестве одного из объяснений обнаруженных преимуществ павианов гамадрилов при выполнении исследовательской деятельности.
Оценка экологии и социальной структуры 4 изучаемых видов обезьян показывает, что все они относятся к числу процветающих и хорошо приспособленных видов приматов. Зеленые мартышки (Chlorocebus spp.) живут в самых разных средах обитания, но предпочитают саванны и мозаику саванных лесов и не встречаются во влажных лесах Центральной и Западной Африки или в пустынях юго-западной Африки (Bi et al., 2009). Эти обезьяны не проводят большую часть времени на деревьях, день проводят в поисках пищи на земле, поднимаясь на деревья на ночь для сна (Cawthon Lang, 2012). Макаки резусы являются 2-м после человека видом приматов по размеру своего ареала, который занимает значительную часть Азии, от Афганистана до Индии и Таиланда. Как и большинство видов макак, ведут полудревесный и полуназемный образ жизни. Свое весьма разнообразное пропитание эти всеядные приматы разыскивают на деревьях и на земле (Seth, Seth, 1986). Макаки яванские живут в равнинных тропических лесах, как первичных, так и в нарушенных и вторичных, в том числе вблизи человеческих поселений, а также в мангровых лесах, умеют хорошо плавать. Этот вид чаще всего встречается на невысоких высотах, где он предпочитает прибрежные и мангровые леса, берега рек и болотные леса (Fooden, 1995). Павианы гамадрилы держатся преимущественно на земле, с большим искусством лазая по самым крутым обрывам и скалам.
Общим для всех 4 видов является их определенная “синантропность”, то есть способность существовать на территориях в непосредственном соседстве и зачастую во взаимодействии с человеком. Кроме того, можно утверждать о значительной экологической пластичности всех этих видов обезьян, позволяющей им жить и размножаться в различных экологических условиях, включая условия неволи либо содержания в иных географических широтах. Именно эта пластичность, по-видимому, обеспечила возможность использования макаков резусов, макаков яванских и зеленых мартышек в качестве главных лабораторных приматов.
К числу общих для изучаемых видов свойств следует отнести то, что все они являются так называемыми “генералистами” в питании. Спектр используемых ими кормов включает в себя корма как растительного, так и животного происхождения, с уклоном в преимущественную фруктоядность, и определяется обычно условиями местности. Так, основной рацион павианов гамадрилов, обитающих в Filoha (Эфиопия), почти круглогодично состоит из плодов дум-пальмы, а также листьев, цветов, почек и семян нескольких других видов растений (Swedell, 2002). При этом в состав рациона павианов гамадрилов входят насекомые и даже некоторые позвоночные животные, а именно: ящерицы, яйца, птицы (Swedell et al., 2008; Schrier et al., 2019). Павианы гамадрилы, несколько сотен которых почти 2 десятилетия свободно жили на территории Гумистинского и Туапсинского заказников, использовали в качестве основного источника корма плоды бука восточного и каштана посевного, а также плоды, почки, листья и семена нескольких десятков других местных дикорастущих растений и некоторые виды животных кормов (Чалян, Мейшвили, 2022). Возможность использования широкого спектра местных кормов была приобретена павианами в процессе освоения местности, на которой они были выпущены, что, несомненно, потребовало от них исходно высокого уровня исследовательской активности. Характерной особенностью павианов гамадрилов также являются очень большие размеры участка обитания, протяженности дневных переходов и численности особей в группах. Так, группа 1, состоявшая из 66 павианов гамадрилов (Эфиопия), занимала участок площадью 28 кв. км (Sigg, Stolba, 1981). Максимальная протяженность дневных переходов этих обезьян могла достигать 19.2 километра. Очень важной особенностью павианов гамадрилов, определяющей их уникальность в семействе приматов, является их социальное устройство и сложное социальное поведение. Группы павианов гамадрилов могут включать в себя до нескольких сотен особей и имеют несколько уровней организации. Для них характерна выраженная патрилинейность, в соответствии с которой обмен особями осуществляется путем переходов самок в гаремы неродственных самцов. При этом имеет место выраженное доминирование самцов над самками.
Макаки яванские, также показавшие в эксперименте высокий уровень исследовательской деятельности, как и павианы, демонстрируют высокую пластичность и способность жить в различных условиях – от мангровых зарослей до субальпийских лесов (Hadi et al., 2007). Экологическая пластичность макаков яванских непосредственно проявляется в пищевой экологии, численности групп и в размерах участков обитания. Для макаков яванских, живущих по соседству с человеком в подкармливаемых популяциях, характерна преимущественная фруктоядность, большая численность особей в группах и маленькие размеры участков обитания, тогда как у макаков яванских, обитающих во вторичных лесах и в мангровых зарослях, небольшая численность особей в группах, большие участки обитания (до 300 га (Fooden, 1995)), всеядность и сложное пищевое поведение (Hansen et al., 2020). Они используют любые возможные пищевые ресурсы, включая насекомых, земляных червей, яйца птиц и самих птиц, и могут применять различные приемы для их добывания (Kemp, Burnett, 2003). В отличие от павианов гамадрилов, у макаков яванских, как и у всех других видов макаков, имеет место матрилинейная структура групп и мультисамцовость. Молодые самцы могут покидать родную группу и присоединяться к другой группе, тогда как молодые самки остаются в составе своей матрилинии. Тем не менее система иерархических отношений у макаков яванских не такая жесткая, как у макаков резусов.
Характеризуя макаков резусов, следует прежде всего отметить, что, несмотря на огромный ареал, для представителей этого вида характерен ряд общих свойств, к числу которых следует отнести сравнительно небольшие размеры участка обитания и протяженности дневных переходов. Анализ участков обитания более 400 групп макаков резусов, живущих в различных биотопах, показал, что размер участка обитания каждой группы в большинстве случаев варьировал в пределах 0.5–3 кв. км (Kali, 2001). Для макаков резусов также характерна значительная всеядность при преимущественной фруктоядности. Рацион этих обезьян включает в себя плоды, семена, цветы, листья и насекомых, причем фрукты могут составлять до 74% их рациона (Sengupta, Radhaktishna, 2016). Характерной особенностью социального устройства макаков резусов является ярко выраженная матрилинейность, в соответствии с которой социальное устройство групп и преимущественное общение особей определяется родственными отношениями самок. Численность особей в группах макаков резусов сильно варьирует. Так, в Бангладеш в состав каждой из 210 исследованных групп входило от 10 до 78 особей (Hasan et al., 2013).
Зеленые мартышки (гриветы), как и павианы гамадрилы, являются африканскими животными. Они используют преимущественно растительные корма, включая фрукты, семена, цветы, почки и листья, изредка поедая случайным образом попадающихся беспозвоночных и даже позвоночных (яйца, птенцы, ящерицы) (Skinner, Smithers, 1990). Для гривет характерны сравнительно небольшие размеры участка обитания, так же как и небольшая численность особей в группах (от 6 до 20 особей) (Ejigu, Bekele, 2009). Как и у макаков резусов, у гривет очень строго выражена матрилинейность и строгая деспотическая система иерархических отношений.
Таким образом, рассмотрение экологии и особенностей социальной структуры изучаемых обезьян показывает, что имеются некоторые существенные различия, которые, по-видимому, можно рассматривать в качестве определяющих особенности исследовательского поведения каждого из видов. К ним следует отнести такие показатели, как размеры участка, пищевое разнообразие, численность особей в группах и жесткость социальных отношений в группах. Так, наиболее низкий уровень исследовательской активности демонстрируют зеленые мартышки и макаки резусы, живущие в сравнительно небольших группах, на сравнительно небольших участках обитания и имеющие очень жестко выраженную деспотическую систему отношений. При этом макаки резусы, с их широким разбросом источников корма, демонстрируют более высокий уровень исследовательской активности по сравнению с зелеными мартышками. Аналогичные преимущества в когнитивных способностях, связанных с разнообразием пищевых источников, обнаружены при сравнении реакции на новые объекты у павианов чакма (Papio ursinus) и гелад (Theropithecus gelada), близкого к павианам наземного вида обезьян (Bergman, Kitchen, 2009). Предполагается, что преимущественные способности павианов связаны с тем, что они обитают в различных биотопах и используют широкий спектр источников пищи, тогда как гелады питаются преимущественно травой (90% рациона). Обращает на себя внимание тот факт, что для макаков яванских, демонстрирующих в нашем исследовании высокие показатели исследовательского поведения, характерно сравнительно более высокое содержание животных кормов в естественном рационе (до 23%). Можно предположить, что способность к отлову беспозвоночных и мелких позвоночных, используемых обезьянами в пищу, может быть тесно связана с показателями исследовательского поведения, выступая в качестве предиктора эволюционного успеха особей данного вида. У павианов гамадрилов таким предиктором, определяющим необходимость эффективного использования исследовательского поведения, являются очень большие размеры участка обитания и широкий выбор пищевых источников. Другими словами, успешность выживания павианов тесно связана с необходимостью постоянного исследования посещаемой местности и находящихся на ней возможных источников пищи.
ЗАКЛЮЧЕНИЕ
Наиболее высокий уровень исследовательской активности и разнообразия исследовательской деятельности обнаружен у макаков яванских и павианов гамадрилов.
Самый низкий уровень исследовательской активности и разнообразия исследовательской деятельности обнаружен у зеленых мартышек.
Макаки резусы по уровню исследовательской активности и уровню разнообразия исследовательской деятельности занимают промежуточное положение.
Для макаков яванских, макаков резусов и павианов гамадрилов обнаружена высокая корреляция между уровнем исследовательской активности и уровнем разнообразия исследовательской деятельности.
В исследуемых группах макаков яванских, павианов гамадрилов и макаков резусов преобладают “тактильные” типы взаимодействия с объектом, у зеленых мартышек – “визуальные”.
Во всех исследуемых группах наблюдается высокая степень индивидуальных различий как по уровню исследовательской активности, так и по уровню разнообразия исследовательской деятельности.
Список литературы
Аникаев А.Е., Мейшвили Н.В., Чалян В.Г., Аникаева Е.Н. Исследование различий между показателями обучения и исследовательской деятельности у половозрелых самцов макаков-резусов (Macaca mulatta) и павианов гамадрилов (Papio hamadryas). Бюллетень экспериментальной биологии и медицины. 2021. 172 (9): 380–384.
Войтонис Н.Ю. Предистория интеллекта: (К проблеме антропогенеза). Изд-во АН СССР. 1949.
Чалян В., Мейшвили Н. Павианы гамадрилы в лесах Черноморского побережья Кавказа. Litres, 2022.
Шовен Р. Поведение животных. М.: Мир, 1972.
Alexander G.M., Hines M. Sex differences in response to children’s toys in nonhuman primates (Cercopithecus aethiops sabaeus). Evolution and Human Behavior. 2002. 23 (6): 467–479.
Barton R.A. Neocortex size and behavioural ecology in primates. Proceedings of the Royal Society of London. Series B: Biological Sciences. 1996. 263 (1367): 173–177.
Belzung C. Measuring rodent exploratory behavior // Techniques in the behavioral and neural sciences. Elsevier. 1999. 13: 738–749.
Bergman T.J., Kitchen D.M. Comparing responses to novel objects in wild baboons (Papio ursinus) and geladas (Theropithecus gelada). Animal cognition. 2009. 12 (1): 63–73.
Berlyne D.E. Curiosity and learning. Motivation and emotion. 1978. 2 (2): 97–175.
Bi S.G., Bené J.K., Bitty E.A., Koné I., Zinner D. Distribution of the green monkey (Chlorocebus sabaeus) in the coastal zone of Côte d’Ivoire. Primate Conservation. 2009. 24(1): 91–97.
Cawthon Lang K.A. Primate Factsheets: Vervet (Chlorocebus) Taxonomy, Morphology, & Ecology. Primate Factsheets Retrieved. 2006. 7 (2012): 2012.
Coleman K., Tully L.A., McMillan J.L. Temperament correlates with training success in adult rhesus macaques. American J. Primatology: Official J. American Society of Primatologists. 2005. 65 (1): 63–71.
Deaner R.O., Isler K., Burkart J., van Schaik C. Overall Brain size and Not Encephalization Quotient, Best Predicts Cognitive Ability across Nonhuman Primates. Brain, Behavior and Evolution. 2007. 70 (2): 115–124.
Dunbar R.I. Neocortex size as a constraint on group size in Primates. J. Human Evolution. 1992. 22 (6): 469–493.
Ejigu D., Bekele A. Population structure, feeding ecology and human-grivet monkeys’ conflict at Bahir Dar University main campus, Bahir Dar. Ethiopian Journal Biological Science. 2010. 9 (1): 35–47.
Ehrlich A. Response to novel objects in three lower primates: greater galago, slow loris, and owl monkey. Behaviour. 1970. 37 (1–2): 55–63.
Fooden J. Systematic review of Southwest Asian longtail macaques, Macaca fasicularis (Rafles, 1821). Fieldiana Zool., new series. 1995. 81: 2–3.
Hadi I., Suryobroto B., Perwitasari–Farajallah D. Food preference of semi-provisioned macaques based on feeding duration and foraging party size. HAYATI J. Biosciences. 2007. 14 (1): 13–17.
Hansen M.F., Ellegaard S., Maeller M.M., van Beest F.M., Fuentes A., Nawangsari V.A., Groendahl C., Frederiksen M.L., Stelvig M. Comparative home range size and habitat selection in provisioned and non-provisioned long-tailed macaques (Macaca fascicularis) in Baluran National Park, East Java, Indonesia. Contributions to Zoology. 2020. 89 (4): 393–411.
Harlow H.F., Blazek N.C., McClearn G.E. Manipulatory motivation in the infant rhesus monkey. J. Comparative and Physiological Psychology. 1956. 49 (5): 444.
Hasan Md.K., Aziz M.A., Alam S.M.R., Kawamoto Y., Jones-Engel L., Kyes R.C., Akhtar Sh., Begum S., Feeroz M. Distribution of Rhesus Macaques (Macaca mulatta) in Bangladesh: inter-population variation in group size and composition. Primate Conservation. 2013. 26 (1): 125–132.
Hassett J.M., Siebert E.R., Wallen K. Sex differences in rhesus monkey toy preferences parallel those of children. Hormones and behavior. 2008. 54 (3): 359–364.
Joly M., Micheletta J., De Marco A., Langermanns J.A., Sterck E.H.M., Waller B.M. Comparative physical and social cognitive skills in macaque species with different degrees of social tolerance. Proceedings of the Royal Society B: Biological Sciences. 2017. 284 (1862): 20162738
Helstab S.A., Kozonen Z.K., Koski S.E., Van Schaik C., Isler K. Manipulation complexity in primates coevolved with brain size and terrestriality. Scientific reports. 2016. 6 (1): 1–9.
Kali G. Ecology and behavour of Rhesus monkeys (Macaca mulatta) in Baikunthapur forest division, Jalpaiguri, West Bengal, India: University of North Bengal. 2001.
Keller H., Schneider K., Henderson B. (ed.). Curiosity and exploration. Springer Science & Business Media, 2012.
Kemp N.J., Burnett J.B. A biodiversity risk assessment and recommendations for risk management of long-tailed macaques (Macaca fascicularis) in New Guinea. Indo-Pacific Conservation Alliance. 2003.
Line S.W., Morgan K.N., Markowitz H. Simple toys do not alter the behavior of aged rhesus monkeys. Zoo Biology. 1991. 10 (6): 473–484.
Loizos C. Play behaviour in higher primates: a review. Primate ethology. 2017. 176–218.
Meguerditchian A., Marie D., Margiotoudi K., Roth M., Nazarian B., Anton J.L., Claidière N. Baboons (Papio anubis) living in larger social groups have bigger brains. Evolution and Human Behavior. 2021. 42 (1): 30–34.
Marsh H.L., Vining A.Q., Levendoski E.K., Judge P.G. Inference by exclusion in lion-tailed macaques (Macaca silenus), a hamadryas baboon (Papio hamadryas), capuchins (Sapajus apella), and squirrel monkeys (Saimiri sciureus). J. Comparative Psychology. 2015. 129 (3): 256.
Milton K. Foraging behavior and the evolution of primate intelligence. In: Byrne R.W., Whitten A. (eds). Machiavellian intelligence: social expertise and the evolution of intellect in monkeys, apes, and humans. Oxford University Press. Oxford. 1988.
Menzel E.W. Responsiveness to objects in free-ranging Japanese monkeys. Behaviour. 1966. 26 (1–2): 130–150.
Menzel Jr E.W., Davenport Jr R.K., Rogers C.M. Some aspects of behavior toward novelty in young chimpanzees. J. Comparative and Physiological Psychology. 1961. 54 (1): 16.
National Research Council. Guide for the care and use of laboratory animals. National Academies Press. 2010.
Petit O., Dufour V., Herrenschmidt M., De Marco A., Sterck E.H., Call J. Inferences about food location in three cercopithecinae species: an insight into the socioecological cognition of primates. Animal Cognition. 2015. 18(4): 821–830.
Platt M.L., Brannon E.M., Briese T.L., French J.A. Differences in feeding ecology predict differences in performance between golden lion tamarins (Leontopithecus rosalia) and Wied’s marmosets (Callithrix kuhli) on spatial and visual memory tasks. Animal Learning & Behavior. 1996. 24(4): 384–393.
Perelman P., Johnson W.E., Roos C., Seuánez H.N., Horvath J.E. A molecular phylogeny of living primates. PLoS Genet. 2011. 7(3): e1001342.
Rajalingham R., Schmidt K., DiCarlo J.J. Comparison of object recognition behavior in human and monkey. J. Neuroscience. 2015. 35(35): 12127–12136.
Rensch B. Ästhetische Faktoren bei Farb-und Formbevorzugungen von Affen. Zeitschrift für Tierpsychologie. 1957. 14(1): 71–99.
Rossi A., Parada F.J., Stewart R., Barwell C., Demas G., Allen C. Hormonal Correlates of Exploratory and Play-Soliciting Behavior in Domestic Dogs. Frontiers in psychology. 2018. 9: 1559.
Sackett G.P. Exploratory behavior of rhesus monkeys as a function of rearing experiences and sex. Developmental Psychology. 1972. 6 (2): 260.
Santillán-Doherty A.M., Cortés-Sotres J., Arenas-Rosas R.V., Márquez-Arias A., Cruz C., Medellín A., Díaz J.L. Novelty-seeking temperament in captive stumptail macaques (Macaca arctoides) and spider monkeys (Ateles geoffroyi). J. Comparative Psychology. 2010. 124 (2): 211.
Schmitt V., Pankau B., Fischer J. Old world monkeys compare to apes in the primate cognition test battery. PloS one. 2012. 7 (4): e32024.
Schreier A.L., Schlaht R.M., Swedell L. Meat eating in wild hamadryas baboons: Opportunistic trade-offs between insects and vertebrates. American J. Primatology. 2019. 81 (7): e23029.
Schwartz B.L. Using Natural Ecology to Predict Higher Cognition in Human and Non-human Primates. Animal Behavior and Cognition. 2019. 6 (4): 344–354.
Sengupta A., Radhakrishna S. Influence of fruit availability on fruit consumption in a generalist primate, the rhesus macaque Macaca mulatta. International J. Primatology. 2016. 37(6): 703–717.
Seth P.K., Seth S. Ecology and behaviour of rhesus monkeys in India. Primate ecology and conservation. 1986. 2: 89–103.
Sigg H., Stolba A. Home range and daily march in a hamadryas baboon troop. Folia primatologica. 1981. 36 (1–2): 40–75.
Siwak C.T., Tapp P.D., Milgram N.W. Effect of age and level of cognitive function on spontaneous and exploratory behaviors in the beagle dog. Learning & Memory. 2001. 8 (6): 317–325.
Skinner J.D., Chimimba C.T. The mammals of the southern African sub-region. Cambridge University Press. 2005.
Swedell L. Ranging behavior, group size and behavioral flexibility in Ethiopian hamadryas baboons (Papio hamadryas hamadryas). Folia Primatologica. 2002. 73 (2–3): 95–103.
Swedell L., Hailemeskel G., Schreier A. Composition and seasonality of diet in wild hamadryas baboons: preliminary findings from Filoha. Folia Primatologica. 2008. 79 (6): 476–490.
Tia B., Viaro R., Fadiga L. Tool-use training temporarily enhances cognitive performance in long-tailed macaques (Macaca fascicularis). Animal cognition. 2018. 21 (3): 365–378.
Welker W.I. Effects of age and experience on play and exploration of young chimpanzees. J. comparative and physiological psychology. 1956a. 49 (3): 223.
Welker W.I. Variability of play and exploratory behavior in chimpanzees. J. Comparative and Physiological Psychology. 1956b. 49 (2): 181.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова