

Журнал высшей нервной деятельности им. И.П. Павлова, 2023, T. 73, № 2, стр. 256-270
Слуховое последействие: влияние неподвижного адаптера на восприятие движущегося стимула
Л. Б. Шестопалова 1, *, **, Д. А. Саликова 1, Е. А. Петропавловская 1
1 ФГБУН Институт физиологии им. И.П. Павлова РАН
Санкт-Петербург, Россия
* E-mail: shestopalovalb@infran.ru
** E-mail: shestolido@mail.ru
Поступила в редакцию 12.07.2022
После доработки 31.10.2022
Принята к публикации 31.10.2022
- EDN: IONKFD
- DOI: 10.31857/S0044467723020107
Аннотация
Воспринимаемые траектории движения дихотических звуковых стимулов с различными пространственными характеристиками были исследованы в тишине и после прослушивания неподвижных адаптеров. Латерализованный адаптер не влиял на воспринимаемое положение ближних (ипсилатеральных) точек траекторий, но “отталкивал” от себя стимулы на противоположной стороне акустического пространства. После воздействия центрального адаптера наблюдалось “отталкивание” латеральных точек траекторий от него, независимо от направления движения стимула. Воздействие неподвижных адаптеров на воспринимаемые траектории движущихся сигналов лучше всего объясняется трехканальной моделью нейронального кодирования слухового пространства.
Явление сенсорного последействия (after-effect) состоит в изменении восприятия тестового стимула после длительного предъявления другого постоянного или повторяющегося стимула. Последействие наблюдается в разных сенсорных модальностях, а также между модальностями. В целом эффекты последействия являются универсальными для всех сенсорных систем, участвующих в ориентации человека в пространстве (Андреева, 2014, 2015, 2016). Наиболее подробно явление последействия изучено для зрительных стимулов. Известными примерами зрительного последействия являются “иллюзия наклона” и “иллюзия водопада”. Первый представляет собой статический эффект последействия, при котором ориентация объектов меняется в зависимости от ориентации предшествующих стимулов (He, McLeod, 2001), а второй – динамический эффект последействия движения, в результате которого неподвижные объекты воспринимаются как движущиеся “навстречу водопаду” (Wade, 1994). Слуховое последействие описывается как аналогичное зрительному: после прослушивания неподвижных сигналов воспринимаемое положение тестовых стимулов смещается дальше от сигналов, вызвавших последействие (статический эффект: Carlile et al., 2001; Phillips, Hall, 2005), а после прослушивания движущихся стимулов неподвижный стимул воспринимается как смещающийся во встречном направлении (динамический эффект: см. обзоры Андреева, 2015, 2017).
Механизмы последействия чаще всего интерпретируют в рамках понятия нейрональной адаптации. Избирательная к признакам звукового стимула адаптация (далее – избирательная адаптация11) была впервые обнаружена на уровне слуховой коры (Ulanovsky et al., 2003), а впоследствии выявлена также на уровне задних холмов и в таламусе, преимущественно за пределами лемнисковых отделов (см. обзор Pérez-González, Malmierca, 2014). Считается, что длительное воздействие адаптирующего стимула (адаптера) приводит к снижению чувствительности нейронов-детекторов, специфичных к его признакам (Barlow, Hill, 1963; Movshon, Lennie, 1979; Maffei et al., 1973; Barlow, 1990). Тем самым избирательная адаптация приводит к смещению паттернов последующих ответов этих детекторов по направлению от паттернов активации, исходно вызываемых адаптером (Clifford et al., 2000; Gutschalk et al., 2008).
Явление избирательной адаптации традиционно используется для экспериментального изучения рецептивных полей нейронов, обеспечивающих пространственный слух. Вплоть до настоящего времени ведется дискуссия относительно двух основных моделей латерализации звука. Популяционная модель (population coding model) предполагает упорядоченное представление акустического пространства в структурах слухового пути, расположенных выше уровня медиальной верхней оливы, при узкой настройке нейронных популяций на определенные величины межушных различий по времени (ΔT) (Joris et al.,1998). Первоначальное подтверждение популяционной модели было получено в электрофизиологическом исследовании среднемозгового центра слуховой системы амбарной совы (Knudsen, Konishi, 1978). В более позднем психофизическом эксперименте по локализации источника звука человеком в свободном звуковом поле наблюдались смещения воспринимаемого положения источника звука в направлении от адаптера, которые также интерпретировались авторами в рамках популяционной теории (Carlile et al., 2001).
Согласно другой модели, нейрональное кодирование азимутального положения звуковых стимулов основывается на активности двух противоположных популяций, имеющих широкую настройку на левое и правое акустическое полупространство и перекрывающихся в области средней линии головы. Эта теория получила название “полусферной модели” (hemifield model). Она адекватно описывает результаты электрофизиологических исследований на животных (Phillips, Irvine, 1981; Jenkins, Masterton, 1982; Jenkins, Merzenich, 1984; Phillips, Brugge, 1985; McAlpine et al., 2001; Stecker, Middlebrooks, 2003) и психофизических исследований на человеке (Boehnke, Phillips, 1999; Phillips et al., 2003). Высказывалось также предположение о существовании третьего “центрального” канала, охватывающего популяцию нейронов, настроенных на малые величины межушных задержек (Phillips, Hall, 2005; Dingle et al., 2012, 2013).
Аргументы в пользу одной из этих моделей можно получить в эксперименте с избирательной адаптацией, в ходе которого задание, данное слушателям, состоит не в различении левосторонних и правосторонних источников звука, а в точной локализации набора стимулов, имеющих разное азимутальное положение. Согласно популяционной модели, максимальное изменение воспринимаемого положения тестового стимула в результате адаптации будет наблюдаться в случае близких значений межушных различий у стимула и адаптера. Согласно полусферной модели, изменения воспринимаемого положения будут распространяться на всю акустическую полусферу, в которой расположен адаптер, и могут отчасти захватывать противоположную полусферу (Boehnke, Phillips, 1999; Vigneault-MacLean et al., 2007).
В представленной работе избирательная адаптация исследована в условиях дихотической стимуляции. Использовались три положения неподвижного адаптера (у левого или правого уха либо у средней линии головы), а тестовые сигналы перемещались от средней линии к уху либо в обратном направлении. Испытуемые определяли положение начальных и конечных точек траекторий движения тестовых стимулов в тишине и в условиях адаптации. В представленном исследовании были поставлены следующие задачи: 1) выяснить, как влияет неподвижный адаптер на воспринимаемые траектории движущихся стимулов; 2) определить, какая из трех моделей в наибольшей степени соответствует полученным данным. Если полусферная модель латерализации верна, то эффект последействия приведет к изменению воспринимаемого положения не только ближайших к адаптеру участков траекторий, но и их отдаленных концов. Аргументом в пользу трехканальной модели может стать различие эффекта последействия для центрального и латерального адаптеров. Если верна популяционная модель, изменения затронут только ближайшие к адаптеру части траекторий.
МЕТОДИКА
Этические нормы. Все исследования проведены в соответствии с принципами биомедицинской этики, сформулированными в Хельсинкской декларации 1964 г. и ее последующих обновлениях, и одобрены Комиссией по этике ФГБУН Института физиологии им. И.П. Павлова РАН (Протокол № 22-02). Каждый участник исследования представил добровольное письменное информированное согласие, подписанное им после разъяснения ему потенциальных рисков и преимуществ, а также характера предстоящего исследования.
Условия эксперимента и испытуемые. В экспериментах приняли участие две группы испытуемых: контрольная и основная. Обе группы состояли из 18 праворуких испытуемых в возрасте от 18 до 45 лет с нормальным слухом (по данным тональной аудиометрии) и без истории неврологических заболеваний (по отчетам испытуемых). Основная группа включала 6 мужчин и 12 женщин, средний возраст 27.8 ± 1.4 лет; 13 слушателей были наивными испытуемыми, и 5 имели опыт участия в слуховых экспериментах. Контрольная группа включала 6 мужчин и 12 женщин, средний возраст 24.8 ± 4.7 лет; один слушатель имел опыт участия в слуховых экспериментах.
Во время экспериментов испытуемые располагались в кресле внутри экранированной звукоизолированной камеры, и им дихотическим способом предъявлялись звуковые сигналы. Как основной, так и контрольный эксперимент проводились за 2–3 визита испытуемых. Время регистрации в каждый экспериментальный день составляло в среднем около двух часов; по просьбе испытуемых им предоставлялись перерывы.
На предварительном этапе каждого эксперимента у всех испытуемых измеряли монауральные пороги слышимости с использованием шумовых посылок полосой 200–10 000 Гц и длительностью 700 мс. В дальнейшем интенсивность тех же звуковых сигналов устанавливалась на уровне 45 дБ над порогом слышимости испытуемого на каждом ухе. При необходимости интенсивность сигнала, подаваемого на одно из ушей, изменяли таким образом, чтобы звуковой образ находился по средней линии головы испытуемого. Обычно вводимая коррекция не превышала 3 дБ. Далее в эксперименте интенсивность всех звуковых стимулов устанавливалась относительно скорректированных значений порогов.
Структура стимуляции. Исследование проведено в рамках проекта по исследованию одновременной слуховой маскировки, что определило структуру стимуляции в условиях основного эксперимента: движущиеся тестовые сигналы предъявлялись либо в тишине, либо на фоне непрерывно звучащего неподвижного сигнала (рис. 1 (а)). В контрольных условиях испытуемым предъявлялись только тестовые сигналы в тишине (рис. 1 (б)).
Рис. 1.
Методика эксперимента. (а): структура эпохи стимуляции. Серая и черная линии – стимулы с медленным и быстрым движением. Верхний ряд – структура тестового сигнала, нижний ряд – структура адаптера. (б): структура серии в основном и контрольном эксперименте. Цифры 1–4 – разные траектории. Серые прямоугольники – эпохи с неподвижным фоновым сигналом, белые – тестовые сигналы в тишине. (в): пространственное положение стимулов. Дуги со стрелками – расчетные траектории тестовых сигналов. Числами от –90 до 90 обозначены угловые расстояния относительно средней линии головы (в градусах). Положение адаптеров показано звездами. Fig. 1. Schematic of the sound stimuli. (a): temporal pattern of a stimulation epoch. Gray and black lines show the slow and fast stimulus motion. Top and bottom panels show the test stimulus and adapter. (б): structure of stimulus block in the after-effect conditions and controls. Numbers 1–4 indicate different sound trajectories. Gray and white rectangles represent epochs with and without background noise, respectively. (в): spatial position of the stimuli. Arrowed arcs show calculated trajectories of the test stimuli. The angular distance relative to the head midline is indicated by numbers from –90 to 90 (in degrees). Location of adapters is indicated by stars.
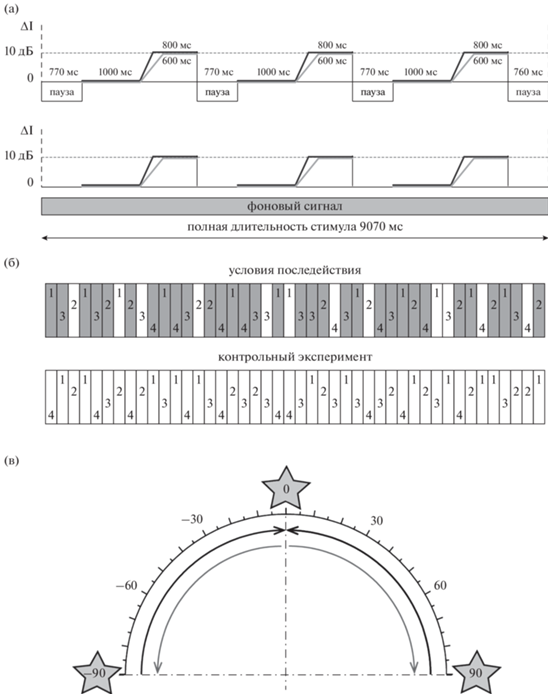
Результаты сравнения реакций на стимул на фоне неподвижного сигнала с реакциями на стимул в тишине (эффект маскировки) не входят в представленную статью и публикуются отдельно. В настоящей работе анализируется эффект последействия, под которым подразумевается влияние комплексных стимулов “фон + сигнал” на восприятие тестовых сигналов в тишине. Оценка эффекта последействия осуществляется путем сравнения реакций на сигнал в тишине из основного эксперимента, где они чередовались с адаптирующим звуком, с реакциями на тот же сигнал в контрольных условиях, где эпох с адаптером не было.
Стимулы. Подробное описание синтеза сигналов изложено в нашей предыдущей работе (Шестопалова и др., 2022). Исходными сигналами служили два отрезка белого шума, синтезированные с частотой дискретизации 96 кГц и фильтрованные в полосе 200–10 000 Гц. Шумовые посылки, использованные для создания движущегося и неподвижного сигналов, были синтезированы независимо, что обеспечивало отсутствие корреляции между ними. Сигналы преобразовывались в аналоговую форму при помощи многоканальной аудиоплаты Gina24 (Echo Audio, США) и предъявлялись дихотически с помощью звукоизлучателей Etymotic ER-2 (Etymotic Research Inc., США).
Полная длительность каждой эпохи стимуляции была одинаковой и составляла 9070 мс, что соответствует длительности неподвижного фонового сигнала. Фронты длительностью 10 мс, сглаженные косинусоидальной функцией, были наложены в начале и в конце фонового неподвижного сигнала, а также в начале и в конце каждого из тестовых сигналов. Длительность сигналов приведена без учета фронтов. Пространственное положение всех стимулов задавалось величиной межушных различий по интенсивности (ΔI).
В эксперименте использовались три типа фоновых неподвижных сигналов, различавшихся только пространственным положением: центральный (ΔI = 0 дБ), левый (ΔI = –10 дБ), и правый (+10 дБ). Они вызывали ощущение неподвижного звукового образа, расположенного либо около средней линии головы, либо вблизи левого или правого уха, что теоретически соответствует угловому положению –90 град или +90 град в случае полной латерализации (рис. 1 (в), звезды).
Тестовые сигналы состояли из трех фрагментов, следующих друг за другом без паузы: 1) начального участка длительностью 1000 мс с постоянной величиной ΔI, равной 0 или ±10 дБ; 2) участка движения длительностью 200 мс или 400 мс с линейным изменением ΔI (от 0 до ±10 дБ и от ±10 до 0 дБ); 3) конечного участка длительностью 800 мс или 600 мс соответственно, с постоянной конечной величиной ΔI, равной ±10 или 0 дБ. Полная длительность тестового сигнала составляла 2000 мс. Данные стимулы вызывали сначала ощущение неподвижного звукового образа по средней линии головы, а затем его плавного движения влево или вправо от средней линии головы, либо сначала ощущение неподвижного звукового образа около одного из ушей, а затем его движения к центру. Расчетные угловые скорости движения стимулов на втором участке, вычисленные на основе соотношения “10 дБ = 90 град”, составили 450 град/с (быстрый) и 225 град/с (медленный) для длительностей участка движения 200 мс или 400 мс соответственно. В течение эпохи, длящейся 9070 мс (с фоном или без него), один и тот же тестовый стимул повторялся трижды. Ответ о воспринимаемом положении сигнала в начале и в конце его звучания регистрировался один раз в конце эпохи, после того как испытуемый прослушал все три повтора стимула.
Интенсивность тестовых сигналов (без фона) устанавливалась на уровне 45 дБ над порогом слышимости испытуемого. Интенсивность фонового сигнала превосходила тестовый стимул на 5 дБ. В условиях маскировки суммирование тестового сигнала (45 дБ) и фона (50 дБ) приводило к увеличению общей интенсивности стимулов на 6 дБ.
Процедура эксперимента. Тип экспериментальной серии определялся пространственным положением фонового сигнала (контроль, левый, центральный, правый) и скоростью движения тестового стимула (медленный, быстрый), т.е. следовал принципу “одна серия – один фон” и “одна серия – одна скорость”. Вид тестового стимула определялся направлением и скоростью движения звукового образа в его среднем фрагменте (рис. 1 (а)). Направления движения соответствовали 4 траекториям: от центра к левому уху или обратно, от центра к правому уху или обратно. В основном эксперименте все серии состояли из 8 видов стимулов: 4 траектории стимулов без фона (в тишине) и те же 4 траектории в сочетании с фоновым сигналом одной локализации. В пределах одной серии эпохи стимуляции чередовались в квазислучайном порядке, причем эпохи без фона (условия последействия) повторялись по 4 раза, а эпохи с фоновым сигналом (адаптером) – по 12 раз. Таким образом, каждая серия основного эксперимента содержала по 48 эпох, в которых на протяжении 9070 мс звучал один и тот же неподвижный фоновый сигнал, и 16 эпох, в которых тестовые стимулы звучали в тишине (рис. 1 (б)).
Суммарная длительность эпох, содержащих адаптер, в каждой серии составляла около 7 минут. Эпохи с тестовыми стимулами в тишине в сумме продолжались около 2.5 минут. Интервал между эпохами, не содержавший звуковой стимуляции, зависел от темпа ответов испытуемого. В зависимости от этого интервала общая длительность серии могла составлять от 12 до 15 минут. Соответственно, звучание адаптера занимало от 45 до 60% времени в течение серии. Однако, в отличие от классической парадигмы адаптации, серии с каждым положением фонового сигнала и каждой скоростью движения стимулов чередовались в квазислучайном порядке в разные экспериментальные дни.
Слушатели должны были определять пространственное положение тестового звукового сигнала и в конце каждой эпохи показывать начало и конец его траектории касанием пера на рабочей поверхности графического планшета Genius G-pen 450. Для этого на рабочей поверхности была изображена дуга, на которую испытуемые должны были спроецировать воспринимаемое положение звуковых сигналов. Интервал от ответа испытуемого до начала следующей эпохи подбирался индивидуально и составлял 1–1.5 с. Положение звукового образа оценивали в градусах относительно средней линии головы.
В контрольном эксперименте 4 вида тестовых стимулов в каждой серии повторялись по 16 раз в квазислучайном порядке, а эпохи с неподвижным фоновым сигналом не предъявлялись. Каждому испытуемому было предъявлено по 4 серии с движущимися стимулами (2 с быстрым движением, 2 с медленным). Порядок следования серий был рандомизирован.
Таким образом, в пределах одного типа серий основного эксперимента пространственное положение фонового сигнала не изменялось, а направление движения тестового стимула было рандомизировано, как на фоне неподвижного сигнала, так и в тишине. Поэтому в контексте эффекта последействия повторяющимся адаптирующим фактором в комплексе “фон + сигнал” было только положение фонового сигнала. Следовательно, при исследовании эффекта последействия и лежащих в его основе процессов адаптации мы можем рассматривать положение фонового сигнала в качестве основной характеристики адаптера и вводить его положение как фактор в статистический анализ. В дальнейшем изложении адаптирующим стимулом (адаптером) будем называть левый, центральный или правый неподвижный сигнал, а тестовым сигналом – сигнал, предъявленный в тишине.
Анализ данных. Воспринимаемое положение начальных и конечных точек траектории движения регистрировали в градусах относительно средней линии головы. Для каждого типа стимулов в основном эксперименте усреднение проводилось по 24 ответам для каждого испытуемого и по 432 ответам по группе, а в контрольном эксперименте – по 16 и по 288 ответам соответственно.
Статистическую оценку полученных результатов проводили по отдельности для центральных и латеральных точек с помощью четырехфакторного дисперсионного анализа (ANOVA) с факторами “Скорость” (Медленно, Быстро), “Адаптер” (контроль, левый, центральный, правый), “Сторона” (слева, справа)22, “Точка” (начальная, конечная). Фактор “Адаптер” включал как данные контрольных измерений (в отсутствие адаптера), так и данные основного эксперимента (три положения адаптера). Взаимодействие “Адаптер*Сторона” характеризует как зависимость эффекта последействия от взаимного расположения тестового и адаптирующего стимулов в основном эксперименте, так и изменения воспринимаемых траекторий в результате последействия по сравнению с контрольными условиями. По фактору “Точка” исследовали эффект последействия для начальных и конечных точек траекторий. Важно подчеркнуть, что и для центральных, и для латеральных точек фактор “Точка” содержал оба уровня (начальная, конечная), так как движение могло быть направлено как от центра к уху, так и от уха к центру. При попарных сравнениях применяли поправку Бонферрони. Все сравнения проводили с уровнем значимости p < 0.05.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Все испытуемые основной и контрольной групп успешно определяли положение начальных и конечных точек траекторий движения стимулов. Эффект последействия оценивался прежде всего по изменениям воспринимаемой траектории стимулов в результате появления на слуховой сцене неподвижного адаптера. Длины воспринимаемых траекторий движения стимулов в отсутствие адаптера (контрольные условия) составили в левой полусфере 70 ± 2 град при движении от центра и 69 ± 2 град при движении к центру, а в правой полусфере 68 ± 2 град при движении в обоих направлениях. Изменения воспринимаемого положения и длины траекторий в условиях адаптации показаны на рис. 2.
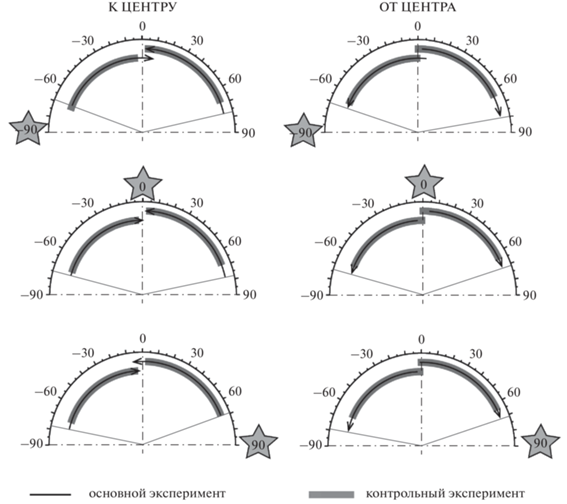
Рис. 2.
Воспринимаемые траектории движения тестовых сигналов в контрольных условиях и при действии адаптеров разной локализации. Тонкими серыми линиями показано угловое положение латеральных точек в условиях адаптации. Условные обозначения – как на рис. 1 (в). Звездами показано расчетное положение адаптеров. Fig. 2. Perceived trajectories of the test stimuli in the control conditions and after adaptation. Thin gray lines show angular positions of the lateral points in the after-effect conditions. Other designations as in fig. 1 (в). Stars indicate the calculated positions of adapters.
Эффект последействия для латеральных точек (как начальных, так и конечных) анализировали с помощью дисперсионного анализа (ANOVA) с факторами “Скорость” (Медленно, Быстро), “Адаптер” (контроль, левый, центральный, правый), “Сторона” (слева, справа), “Точка” (начальная, конечная). Выявлен единственный главный эффект Адаптера (F(3, 544) = 7.88, p < 0.001) и взаимодействие Адаптер*Сторона (F(3, 544) = = 9.30, p < 0.001). Все остальные эффекты и взаимодействия оказались незначимы (p > > 0.05). Результаты попарных сравнений по фактору “Адаптер” для стимулов, звучащих слева и справа, показаны на рис. 3 пунктирными линиями. В случае левосторонних сигналов появление контралатерального адаптера (справа) приводило к сдвигу левой латеральной точки к уху (p < 0.01). Появление центрального адаптера производило аналогичный эффект, но он был статистически незначим (p > 0.05), а ипсилатеральный адаптер не влиял на воспринимаемое положение левой латеральной точки (p > 0.05). В случае правосторонних сигналов появление контралатерального или центрального адаптера приводило к сдвигу правой латеральной точки к уху по сравнению с контролем (p < 0.001 для обоих), а ипсилатеральный не влиял на положение латеральной точки (p > 0.05).
Рис. 3.
Влияние последействия на положение латеральных точек траекторий. Латеральные точки показаны серыми кружками на схеме головы. По оси ординат – азимутальное смещение от средней линии (в градусах). Темные столбики – стимулы слева от средней линии, светлые столбики – стимулы справа. Вертикальными черточками показаны стандартные ошибки среднего. Сплошными и пунктирными линиями показаны значимые попарные сравнения (подробности в тексте). Fig. 3. After-effect induced by stationary adapters on the lateral points of trajectories. Lateral points are shown by gray circles in the head scheme. Y-axis measures the azimuthal displacement from the head midline (degrees). The dark bars show the left-side stimuli, the light bars show the right-side stimuli. The error bars indicate the standard error of means. Solid and dotted lines indicate significant ANOVA post-hocs (see details in the text).
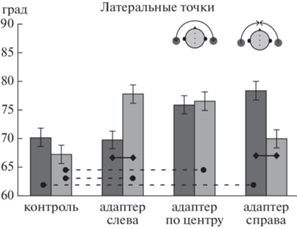
Для левого и правого адаптеров были обнаружены значимые попарные сравнения по фактору “Сторона” (сплошные линии на рис. 3), что подтверждает асимметрию воспринимаемого положения точек траектории, расположенных ипси- и контралатерально относительно адаптера. Значимые различия были связаны с тем, что на стороне, контралатеральной адаптеру, латеральная точка была расположена ближе к уху, чем на ипсилатеральной (p < 0.001 для левого и правого адаптеров). Для центрального адаптера и для контроля различий положения левой и правой латеральной точки не обнаружено (p > 0.05).
Эффект последействия для центральных точек (как начальных, так и конечных) также анализировали с помощью дисперсионного анализа (ANOVA) с факторами “Скорость” (Медленно, Быстро), “Адаптер” (контроль, левый, центральный, правый), “Сторона” (слева, справа), “Точка” (начальная, конечная). Обнаружен единственный главный эффект Адаптера (F(3, 544) = 17.29, p < 0.001), а также взаимодействия “Адаптер*Сторона” (F(3, 544) = 4.93, p < 0.01) и “Адаптер*Точка*Сторона” (F(3, 544) = 4.82, p < 0.01). Все остальные эффекты и взаимодействия не достигли уровня значимости (p > 0.05). Результаты попарных сравнений по фактору “Адаптер” для стимулов, звучащих слева и справа, показаны на рис. 4 (пунктир) отдельно для начальных и для конечных точек, поскольку фактор “Точка” входил в значимое взаимодействие.
Рис. 4.
Влияние последействия на положение центральных точек траекторий при движении стимулов от центра и к центру. Центральные точки показаны серыми кружками на схеме головы. Вверху – воспринимаемое положение начальных точек, внизу – конечных точек. Остальные обозначения – как на рис. 3. Fig. 4. After-effect induced by stationary adapters on the central points of trajectories. Central points are shown by gray circles in the head scheme. Top panel: the perceived position of the starting points for motion from center. Bottom panel: the perceived position of the end points for motion to center. Other designations as in fig. 3.
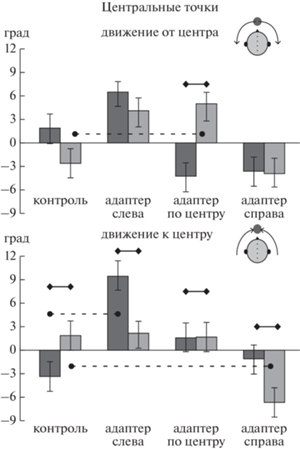
Вовлеченность фактора “Точка” во взаимодействие означает, что взаимодействие “Адаптер*Сторона” реализовывалось по-разному для центробежных и центростремительных стимулов. Сравнения по фактору “Адаптер” показали следующее. При движении от центра вправо в отсутствие адаптера траектория начиналась из-за средней линии (т.е. из левой полусферы), а при появлении центрального адаптера начало траектории смещалось по ходу движения стимула (различия положения начальных точек 7 град, p < 0.05). При движении от центра влево наблюдались аналогичные тенденции, но они не достигли уровня значимости (p > 0.05).
При движении справа к центру в отсутствие адаптера траектория не доходила до центра, и конечная точка оставалась в правой полусфере. При появлении ипсилатерального адаптера (справа) траектория заканчивалась в левой полусфере (различия положений концов 9 град, p < 0.01). Аналогично, при движении слева к центру в отсутствие адаптера траектория не доходила до центра, а введение ипсилатерального адаптера (слева) приводило к смещению конца траектории в правую полусферу (различия положений концов 13 град, p < 0.001).
Сравнения по фактору “Сторона” для каждого адаптера в отдельности (сплошные линии на рис. 4) показали, что при движении от центра влево и вправо только при действии центрального адаптера начальные точки обеих траекторий сдвинуты от центра по ходу движения (p < 0.001). Для остальных адаптеров и для контроля различий нет (p > 0.05). При движении слева и справа к центру при центральном адаптере (ближайшем к концу траекторий) различий нет (p > 0.05). В отсутствие адаптера траектории не доходят до центра (различия положений концов 5 град, p < 0.05). При действии обоих ипсилатеральных стимулам адаптеров траектории заканчивались в противоположной полусфере (различия положений концов 7 град, p < 0.01 для левого адаптера и 5 град, p < 0.05 для правого адаптера).
Сравнение длин траекторий в целом (с учетом как центральных, так и латеральных концов) с контрольными условиями показало, что при действии адаптера происходило удлинение траекторий за счет отдаленных от адаптера областей пространства. Максимальное удлинение наблюдалось при движении к центру (7–9 град).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Исследовали влияние неподвижных адаптирующих стимулов разной локализации на восприятие траекторий движения тестовых стимулов. Слушатели отмечали воспринимаемое положение начальных и конечных точек траекторий с помощью графического планшета. Положение латеральных точек (расположенных около левого или правого уха) и центральных (около средней линии головы) анализировали по отдельности. Эффект последействия проявлялся при сравнении воспринимаемых траекторий движения тестовых сигналов при наличии и при отсутствии адаптирующего стимула, т.е. при сравнении данных основного и контрольного эксперимента.
Длины воспринимаемых траекторий в контрольных условиях не достигали расчетных 90 град, так как межушные различия ΔI = = ±10 дБ не вызывали ощущения полной латерализации звукового образа. Неполная латерализация – достаточно распространенное явление при дихотическом предъявлении звуковых стимулов, даже в тех случаях, когда величина межушных различий превышает физиологически возможную. Так, при локализации щелчков (Варягина, Радионова, 2004) траектории движения слитного звукового образа у большинства испытуемых не достигали полной латерализации даже при больших значениях межушных различий (ΔI = ±13 дБ). Степень латерализации звукового образа, созданного в настоящем исследовании за счет межушных различий ΔI = ±10 дБ, была эквивалентна описанным в литературе данным по латерализации при межушных различиях по времени ΔТ = ± 800 мкс (Петропавловская и др., 2011).
Влияние длительности и порядка следования стимулов
Интересными представляются вопросы о продолжительности адаптирующего стимула и структуре последовательности звуковых стимулов, необходимой для изменения воспринимаемой траектории. Из литературы известно, что в работах по изучению эффекта последействия методами психофизики минимальное время непрерывного звучания адаптирующего стимула в начале серии составляло 30 секунд (табл. 1). В нашей работе время непрерывного звучания адаптера было меньше почти в три раза. Такая структура стимуляции, тем не менее, позволила получить устойчивый эффект последействия. В применявшейся нами последовательности количество эпох с адаптером почти втрое превышало количество эпох, в которых предъявлялся тестовый сигнал. Несколько эпох с адаптером могли следовать друг за другом с промежутком порядка 3–4 секунд от конца одного стимула до начала следующего, в зависимости от скорости ответов испытуемых (рис. 1 (б)). Суммарное время звучания адаптера в течение серии составило около 7 минут при общей длительности серии 12–15 минут (45–60% времени). Таким образом, для возникновения эффекта последействия не требуется длительное непрерывное предъявление адаптирующего стимула или фиксированная структура последовательности.
Таблица 1.
Продолжительность воздействия адаптера в психофизических исследованиях эффекта последействия Table 1. Adapter duration in the psychophysical studies of auditory after-effect
| Публикация | Структура стимуляции |
|---|---|
| Grantham D.W., Wightman F.L., 1979 | В течение 8 минут, каждые 10 секунд адаптирующий стимул прерывался на 1 секунду тестовым стимулом |
| Grantham D.W., 1989 | В начале серии адаптер звучал 30 секунд, далее повторяясь по 10 секунд после тестового стимула |
| Grantham D.W., 1992 | В начале серии адаптер звучал 30 секунд, далее повторяясь по 2 секунды после тестового стимула, длительность которого варьировалась |
| Phillips D.P., Hall S.E., 2005; Vigneault-MacLean B.K. et al., 2007; Dingle R.N. et al., 2012 |
В начале серии адаптер звучал 35 секунд, далее повторяясь по 5 секунд |
| Carlile S. et al., 2010 | В начале серии адаптер звучал 4 минуты, далее повторяясь по 15 секунд |
| Представленная работа | Адаптер звучал интервалами по 9 секунд, в случайном порядке чередуясь с тестовыми стимулами |
Нужно также отметить, что настоящее исследование выявило существенное влияние адаптирующих стимулов на воспринимаемое положение тестовых, несмотря на то, что применявшаяся парадигма стимуляции не является оптимальной для исследования эффекта последействия. Действительно, динамические стимулы, траектории которых располагались как справа, так и слева, предъявлялись не только после неподвижных адаптирующих, но и одновременно с ними, в соответствии с парадигмой одновременной маскировки. Однако в этих эпохах интенсивность динамических стимулов была на 5 дБ ниже, чем фонового неподвижного сигнала. Это позволяет предположить, что во время действия адаптера были преимущественно активированы те нейронные популяции, которые настроены на сторону, где звучал неподвижный сигнал. Динамические стимулы могли вовлекать в реакцию и те нейроны, которые настроены на противоположную сторону пространства, но в меньшей степени (из-за разницы в интенсивности). Важно также, что движущиеся стимулы, присутствовавшие в эпохах с адаптером, предъявлялись в случайном порядке, а неподвижный адаптер не изменялся на протяжении всей серии, поэтому их воздействие могло лишь незначительно ослаблять эффект последействия неподвижного стимула.
Движущиеся фрагменты стимулов повторялись по три раза в каждой эпохе (с адаптером и без него). Известно, что при достаточно длительном предъявлении или частом повторении движущихся стимулов может возникать эффект последействия движения (Андреева, 2015, 2017). В нашем исследовании вклад эффекта последействия движения можно полагать пренебрежимо малым по двум причинам: во-первых, из-за малой длительности движущихся фрагментов (200 мс или 400 мс) и, во-вторых, из-за случайного чередования стимулов, движущихся в разных направлениях, между эпохами. На этом основании мы считаем, что движущийся фрагмент стимулов не оказывал влияния на основные выводы нашей работы, связанные с последействием неподвижных адаптеров.
Таким образом, можно заключить, что эффект последействия может возникать при предъявлении адаптирующих и тестовых стимулов в случайном порядке. Вероятно, ключевым фактором для формирования эффекта последействия является соотношение времени воздействия адаптирующих и тестовых стимулов в серии.
Интеграция и разделение пространственных признаков
На локализацию траекторий комплексных стимулов также могут воздействовать эффекты “притягивания” (pulling) и “отталкивания” (pushing), неоднократно описанные в исследованиях, посвященных обработке пространственной информации при анализе слуховой сцены и разделении звуковых потоков (см. обзор Lee et al., 2009). Известно, что пространственные признаки отдельных элементов, входящих в состав комплексных звуковых стимулов, вносят существенный вклад в определение перцептивных границ между слуховыми объектами. Воспринимаемое положение одновременно или последовательно предъявляемых звуковых стимулов отличается от того, где бы они слышались, если бы предъявлялись по отдельности. “Притягивание” (уменьшение расстояния) свидетельствует об интеграции пространственных признаков и наблюдается между сигналами, которые по совокупности других характеристик (спектральных и временных) могут относиться к одному объекту. “Отталкивание” (увеличение воспринимаемого углового расстояния) между звуковыми стимулами считается признаком того, что слуховая система относит их к разным объектам. Влияние “отталкивания” уменьшается по мере разнесения конкурирующих стимулов во времени. Здесь важно отметить, что тестовые стимулы в основном и контрольном эксперименте были одинаковыми, и влияние процессов “притягивания” и “отталкивания” внутри тестового стимула в этих двух условиях совпадали, а в основном эксперименте к ним добавлялось взаимодействие с адаптирующим стимулом.
Влияние последействия на положение латеральных точек. Для латеральных точек (как и для центральных, см. ниже) обнаружено взаимодействие факторов Адаптер и Сторона звучания, указывающее на изменения воспринимаемых траекторий при появлении на слуховой сцене адаптирующего стимула. В случае адаптирующего воздействия на латеральную область траектории (около уха) это взаимодействие проявлялось одинаково для начальных и конечных латеральных точек. Таким образом, в латеральной области акустического пространства направление движения тестового стимула не влияло на проявление эффекта последействия.
Действие центрального адаптера на латеральные точки приводило к их смещению в сторону уха с обеих сторон, хотя этот эффект был статистически значим только справа. Явление “отталкивания” между адаптером и тестовыми стимулами свидетельствует о том, что эти два стимула интерпретировались слуховой системой как отдельные объекты. Возможное функциональное значение “отталкивания” – усиление обнаруженных различий по пространственному признаку, что должно способствовать выделению перцептивных границ и лучшему различению отдельных объектов слуховой сцены.
До некоторой степени удивительным представляется тот факт, что при появлении неподвижного адаптера около одного уха ближайшая к нему латеральная точка траектории не смещалась по сравнению с контролем. В то же время дальняя от адаптера латеральная точка (на противоположной стороне) смещалась в сторону другого уха, то есть дальше от адаптера (рис. 2, верхняя и нижняя панели). Важно также подчеркнуть, что эффект последействия зависел от взаимного положения тестового и адаптирующего стимулов. Эта зависимость показана на рис. 3 сплошными линиями. Латеральная точка траектории сигнала, контралатерального адаптеру, была сдвинута дальше к уху, чем ипсилатерального. Иными словами, наблюдалось расширение противоположной стороны субъективного акустического пространства по сравнению с контролем. Последействие латеральных адаптеров сказывалось главным образом в отдаленной от них части пространства.
Влияние последействия на положение центральных точек. В отношении воспринимаемого положения центральных точек факторы Адаптер и Сторона звучания оказались вовлечены в трехфакторное взаимодействие с фактором Точка, поэтому эффект последействия анализировали отдельно для начальных и конечных центральных точек. Иными словами, отдельно анализировали движение тестового стимула от центра и к центру (рис. 4). При движении от центра к уху влияние последействия на начальные точки наблюдалось только при действии центрального адаптера, то есть расположенного ближе всего к началу траекторий. Положение начальных точек было смещено по ходу движения.
При движении тестового сигнала от уха к центру эффект последействия имел место только для ипсилатерального адаптера, то есть расположенного вблизи начала траектории, как и в предыдущем случае. В контрольных условиях (без адаптера) траектория сигнала не доходила до центра, а при действии ипсилатерального адаптера конец траектории “проходил дальше центра” и смещался в противоположную полусферу. Различия положений концов траекторий при действии адаптера и без него составили 9 и 13 град при движении справа и слева соответственно. Тем самым продемонстрирован эффект последействия на удаленных от адаптера участках траекторий.
Зависимость воспринимаемого положения центральных точек от направления движения свидетельствует о том, что движущийся фрагмент в середине стимула оказывал воздействие на восприятие положения начала и конца траектории, соответствующих стационарным участкам. При движении тестовых сигналов от центра к уху положение начальных точек смещалось по ходу движения стимула, а на воспринимаемое положение конечных точек при движении от уха центральный адаптер не влиял. Можно предположить, что при локализации начальной точки направление “отталкивания” от адаптера совпадало с направлением движения, что привело к смещению воспринимаемого положения.
При локализации конечной точки действие этих двух факторов было противонаправленным и смещение не наблюдалось. Это свидетельствует о взаимодействии эффектов, действующих в разных временных интервалах – интеграции информации в пределах одного объекта (тестового стимула) и эффекта последействия, являющегося результатом влияния одного объекта (адаптера) на другой (тестовый стимул), отставленный во времени.
Модели пространственного восприятия
Обнаруженные в настоящем исследовании закономерности можно интерпретировать с точки зрения моделей пространственного восприятия. Как было сказано выше, в литературе описаны две принципиально различные концепции: популяционная модель, предполагающая узкую пространственную настройку рецептивных полей, и полусферная модель, основанная на понятии широких рецептивных полей. Частным случаем полусферной модели является трехканальная модель, допускающая существование отдельного нейронального ансамбля, настроенного на фронтальный акустический сектор. Можно утверждать со всей определенностью, что полученные нами результаты не могут быть описаны популяционной моделью, поскольку последействие латеральных адаптеров изменяло траектории не в ближайшей, а в удаленной от адаптера области пространства. Более сложным представляется вопрос о совместимости наших данных с полусферной либо с трехканальной моделью.
Полусферная модель восприятия движения звука предполагает зависимость ответа на движущийся стимул от его направления. Электрофизиологические исследования на животных и на человеке показали, что нейрональный ответ более выражен, когда источник звука приближается к предпочитаемой данной популяцией нейронов стороне пространства, чем когда он удаляется от нее (Magezi, Krumbholz, 2010; см. также обзор Salminen et al., 2012). Однако наш эксперимент не выявил связи между воспринимаемым положением латеральных точек и направлением движения.
С другой стороны, можно рассмотреть полученные результаты без учета направления. В рамках полусферной модели локализация источника звука определяется совместным действием нейронов, настроенных на левую и правую стороны пространства. Рассмотрим для примера случай левостороннего адаптера. Нейроны, настроенные на левую сторону, ответили бы сильнее на сигналы, движущиеся слева, чем справа, но при появлении левостороннего адаптера они станут подвержены избирательной адаптации и естественная асимметрия ответа этой нейронной популяции уменьшится. Нейроны, настроенные на правую сторону, ответят сильнее на сигналы, движущиеся справа, чем слева. При появлении левостороннего (контралатерального их предпочитаемой стороне) адаптера они в меньшей степени подвержены адаптации, поэтому последействие слабо повлияет на правостороннюю асимметрию их реакции. В итоге при левом адаптере больший вклад в результирующий ответ внесут правонастроенные нейроны, что может выражаться в улучшении восприятия правосторонних сигналов или смещении воспринимаемого положения любых сигналов вправо. Аналогично, в случае правостороннего адаптера преимущество получает левая сторона. Наши результаты находятся в соответствии с этой логикой: обнаружено расширение субъективного акустического пространства на противоположной адаптеру стороне.
Исследования методом МЭГ показали, что центральный адаптер вызывал практически такой же эффект, как и контралатеральный, по отношению к сигналу (Salminen et al., 2009), поэтому в рамках полусферной модели центральный и контралатеральный адаптеры относят к одной категории (Salminen et al., 2012). Сравним их действие в нашем эксперименте. И центральный, и контралатеральный адаптеры вызывали смещение латеральных точек к уху. Значит, по действию на латеральные точки они схожи. Действие на центральные точки надо рассматривать отдельно для начальных и конечных точек. Положение начальных точек было смещено по ходу движения только в случае центрального адаптера (латеральные адаптеры не производили значимого эффекта). При движении тестового сигнала от уха к центру, наоборот, эффект последействия проявлялся только в случае ипсилатерального адаптера, то есть расположенного вблизи начала траектории. Следовательно, центральный адаптер действовал на начальные центральные точки, а латеральный адаптер – на конечные центральные точки ипсилатеральных ему траекторий. В этом их важное отличие.
Трехканальная модель предполагает, что центральный адаптер приводит к сдвигу воспринимаемого положения неподвижных источников звука к периферии (Dingle et al., 2010, 2012, 2013). В рамках этой модели локализация точки источника звука определяется совместным действием не двух, а трех групп нейронов, настроенных на центральную, левую и правую части пространства. Рассмотрим случай центрального адаптера. Нейроны, настроенные на центральный сектор, ответили бы одинаково на сигналы, движущиеся в левой и в правой полусферах; при появлении центрального адаптера они станут подвержены избирательной адаптации и их ответ симметрично уменьшится, траектории отодвинутся от центра, что и показано на рис. 2.
Таким образом, принимая во внимание существенное различие эффектов центрального и латеральных адаптеров, мы можем сделать вывод, что полученные нами данные лучше всего описываются трехканальной моделью локализации.
Роль избирательной адаптации в слуховой системе не сводится только к предотвращению избыточной стимуляции тех или иных нейронных популяций. Избирательная адаптация может также влиять на чувствительность нейронов к стимульному контексту, что и получает отражение в эффекте последействия. Современный подход к интерпретации последействия ставит этот эффект в один ряд с потенциалом негативности рассогласования (Pérez-González, Malmierca, 2014; Malmierca, Auksztulewicz, 2021). Считается, что оба феномена неразрывно связаны с процессами предсказательного кодирования, поскольку отражают способность слуховой системы обнаруживать акустические закономерности и обеспечивать выраженную реакцию на стимулы, отличающиеся от текущего контекста. Полученные в настоящем исследовании свидетельства субъективного расширения противоположной от адаптера стороны акустического пространства вполне согласуются с такой интерпретацией избирательной адаптации.
ВЫВОДЫ
1. Последействие неподвижного адаптера, расположенного около уха, сказывалось главным образом в отдаленной части акустического пространства. Оно приводило к сдвигу воспринимаемого положения центральных и контралатеральных относительно адаптера точек траекторий в противоположную от адаптера сторону по сравнению с контролем, и не влияло на ипсилатеральные точки.
2. При действии адаптера, расположенного в центральном секторе слухового пространства, эффект последействия наблюдался также вблизи от адаптера, но только при движении тестовых сигналов от центра к уху: положение начальных точек было смещено от адаптера по ходу движения стимула. На воспринимаемое положение конечных точек при движении от уха центральный адаптер не влиял. Латеральные точки траекторий были смещены от адаптера, независимо от направления движения стимула.
3. Эффект последействия неподвижных звуковых стимулов на воспринимаемые траектории движущихся сигналов лучше всего описывается трехканальной моделью нейронального кодирования слухового пространства.
Список литературы
Андреева И.Г. Последействие движения как универсальное явление для сенсорных систем, участвующих в ориентации в пространстве. I. Зрительное последействие. Журн. эвол. биохим. и физиол. 2014. 50: 413–419.
Андреева И.Г. Последействие движения как универсальное явление для сенсорных систем, участвующих в ориентации в пространстве. II. Слуховое последействие. Журн. эвол. биохим. и физиол. 2015. 51: 145–153.
Андреева И.Г. Последействие движения как универсальное явление для сенсорных систем, участвующих в ориентации в пространстве. III. Последействие, возникающее при адаптации к движению в соматосенсорной и вестибулярной системах. Журн. эвол. биохим. и физиол. 2016. 52: 307–315.
Андреева И.Г. Сенсорное последействие движения. Сенсорные системы. 2017. 31: 279–290.
Варягина О.В., Радионова Е.А. Индивидуальные особенности испытуемых при латерализации неподвижного и движущегося звуковых образов (виртуальная реальность: частные проявления). Журн. эвол. биохим. и физиол. 2004. 40 (5): 441–449.
Петропавловская Е.А., Шестопалова Л.Б., Вайтулевич С.Ф. Предсказательная способность слуховой системы при плавном движении и скачкообразном перемещении звуковых образов малой длительности. Журн. высшей нервной деятельности им. И.П. Павлова, 2011. 61 (3): 293–305.
Шестопалова Л.Б., Петропавловская Е.А., Саликова Д.А., Семенова В.В., Никитин Н.И. Слуховые вызванные потенциалы человека в условиях пространственной маскировки. Физиология человека. 2022 (в печати).
Barlow H.B. A theory about the functional role and synaptic mechanisms of visual after-effects, in Vision: Coding and Efficiency. Ed. Blakemore C. Cambridge University Press. 1990. 363–375 p.
Barlow H.B., Hill R.M. Evidence for a physiological explanation of the waterfall phenomenon and figural after-effects. Nature. 1963. 28: 1345–1347.
Boehnke S.E., Phillips D.P. Azimuthal tuning of human perceptual channels for sound location. J. Acoust. Soc. Am. 1999. 106: 1948–1955.
Carlile S., Hyams S., Delaney S. Systematic distortions of auditory space perception following prolonged exposure to broadband noise. J. Acoust. Soc. Am. 2001. 110: 416–424.
Clifford C.W., Wenderoth P., Spehar B. A functional angle on some after-effects in cortical vision. Proc. Biol. Sci. 2000. 267: 1705–1710.
Dingle R.N., Hall S.E., Phillips D.P. A midline azimuthal channel in human spatial hearing. Hear. Res. 2010. 268: 67–74.
Dingle R.N., Hall S.E., Phillips D.P. The three-channel model of sound localization mechanisms: interaural level differences, J. Acoust. Soc. Am. 2012. 131 (5): 4023–4029. https://doi.org/10.1121/1.3701877
Dingle R.N., Hall S.E., Phillips D.P. The three-channel model of sound localization mechanisms: Interaural time differences. J. Acoust. Soc. Am. 2013. 133 (1): 417–424. https://doi.org/10.1121/1.4768799
Gutschalk A., Micheyl C., Oxenham A.J. The pulse-train auditory aftereffect and the perception of rapid amplitude modulations. J. Acoust. Soc. Am. 2008. 123 (2). https://doi.org/10.1121/1.2828057
Grantham D.W., Wightman F.L. Auditory motion after-effects. Perception & Psychophysics. 1979. 26 (5): 403–408.
Grantham D.W. Motion after-effects with horizontally moving sources in the free field. Perception & Psychophysics. 1989. 45 (2): 129–136.
Grantham D.W. Adaptation to auditory motion in the horizontal plane: Effect of prior exposure to motion on motion detectability. Perception & Psychophysics 1992. 52 (2): 144–150.
He S., MacLeod D.I. Orientation-selective adaptation and tilt after-effect from invisible patterns. Nature. 411: 473–476.
Jenkins W.M., Masterton R.B. Sound localization: effects of unilateral lesions in central auditory pathways. J. Neurophysiol. 1982. 47: 987–1016.
Jenkins W.M., Merzenich M.M. Role of cat primary auditory cortex for sound localization behavior. J. Neurophysiol. 1984. 52: 819–847.
Joris X., Smith P.H., Yin T.C. Coincidence detection in the auditory system: 50 years after Jeffress. Neuron. 1998. 21: 1235–1238.
Knudsen E.I., Konishi M. Space and frequency are represented separately in the auditory midbrain of the owl. J. Neurophysiol. 1978. 41: 870–884.
Lee A.K., Deane-Pratt A., Shinn-Cunningham B.G. Localization interference between components in an auditory scene. J. Acoust. Soc. Am. 2009. 126: 2543–2555. https://doi.org/10.1121/1.3238240
McAlpine D., Jiang D., Palmer A.R. A neural code for low-frequency sound localization in mammals. Nat. Neurosc. 2001. 4: 396–401.
Maffei L., Fiorentini A., Bisti S. Neural correlates of perceptual adaptation to gratings. Science. 1973. 182: 1036–1038.
Magezi D.A., Krumbholz K. Evidence for opponent-channel coding of interaural time differences in human auditory cortex. J Neurophysiol. 104: 1997–2007.
Malmierca M.S., Auksztulewicz R. Stimulus-specific adaptation, MMN and predictive coding. Hearing Research. 2021. 399. https://doi.org/10.1016/j.heares.2020.108076
Movshon J.A., Lennie P. Pattern-selective adaptation in visual cortical neurons. 1979. Nature. 278: 850–852.
Pérez-González D., Malmierca M.S. Adaptation in the auditory system: an overview. Frontiers in Integrative Neuroscience. 2014. 8: 19. https://doi.org/10.3389/fnint.2014.00019
Phillips D.P., Brugge J.F. Progress in neurophysiology of sound localization. Annu. Rev. Psychol. 1985. 36: 245–274.
Phillips D.P., Hall S.E. Psychophysical evidence for adaptation of central auditory processors for interaural differences in time and level. Hearing Research. 2005. 202: 188–199. https://doi.org/10.1016/j.heares.2004.11.001
Phillips D.P., Irvine D.R.F. Responses of neurons in physiologically defined area AI of cat cerebral cortex: sensitivity to interaural intensity differences. Hear. Res. 1981. 4: 99–307.
Phillips D.P., Vigneault-McLean B.K., Boehnke S.E., Hall S.E. Acoustic hemifields in the spatial release from masking of speech by noise. J. Am. Acad. Audiol. 2003. 14: 518–524.
Salminen N.H., May P.J., Alku P., Tiitinen H. A population rate code of auditory space in the human cortex. PLoS One. 2009. 4:e7600.
Salminen N.H., Tiitinen H., May P.J. Auditory Spatial Processing in the Human Cortex. The Neuroscientist. 2012. 18 (6): 602–612. https://doi.org/10.1177/1073858411434209
Stecker G.C., Middlebrooks J.C. Distributed coding of sound locations in the auditory cortex. Biol. Cybern. 2003. 89: 341–349.
Vigneault-McLean B.K, Hall S.E., Phillips D.P. The effects of lateralized adaptors on lateral position judgments of tones within and across frequency channels. Hear. Res. 2007. 24: 93–100.
Ulanovsky N., Las L., Nelken I. Processing of low-probability sounds by cortical neurons. Nat. Neurosci. 2003. 6: 391–398.https://doi.org/10.1038/nn1032
Wade N.J. A selective history of the study of visual motion after-effects. Perception. 1994. 23: 1111–1134.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова