

Журнал высшей нервной деятельности им. И.П. Павлова, 2023, T. 73, № 5, стр. 637-650
Половые различия селективного слухового внимания при дихотическом прослушивании различной степени сложности: данные фмрт-исследования
Л. А. Майорова 1, 2, *, А. Б. Кушнир 1
1 ФГБУН Институт высшей нервной деятельности и нейрофизиологии Российской академии наук
Москва, Россия
2 Федеральный научно-клинический центр реаниматологии и реабилитологии
Москва, Россия
* E-mail: larimayor@gmail.com
Поступила в редакцию 29.04.2023
После доработки 12.06.2023
Принята к публикации 03.07.2023
- EDN: LKHFVY
- DOI: 10.31857/S0044467723050076
Аннотация
Группа из 20 здоровых испытуемых (11 мужчин и 9 женщин) при проведении фМРТ выполняла задание на селективное слуховое внимание в парадигме дихотического прослушивания с различным уровнем перцептивной нагрузки. Анализ паттернов активации головного мозга, измеренной с помощью фМРТ во время селективного прослушивания слитной речи с дистракторами различной силы, выявил статистически значимые половые различия в топографии корковой активности. Эффекты “женщины > мужчины” преобладали в области левой верхней височной извилины, а также левых пре- и постцентральной извилин; в то время как наиболее выраженные эффекты “мужчины > женщины” выявлены в области левого островка, скорлупы и лобной покрышки. Статистически значимые эффекты также получены при сопоставлении паттернов активации по уровню сложности задания: показано, что маскировка голосом диктора женского пола вызывает бо́льшую активацию дополнительных областей высокого уровня обработки информации. Полученные данные свидетельствуют о наличии полового диморфизма организации системы селективного слухового внимания.
ВВЕДЕНИЕ
Обширная литература документирует влияние пола на анатомию, химию и функции мозга (Cahill, 2014), хотя до недавнего времени пол не был предметом систематических нейрофизиологических исследований. Высокая статистическая значимость влияния пола на организацию многих видов поведения, лежащих вне прямых половых функций, убедительно продемонстрирована в обзоре (Voyer et al., 2017), в котором обобщены результаты большого числа нейробиологических и биомедицинских исследований. Однако нейробиологические основы этих различий недостаточно хорошо изучены. Применительно к задачам, связанным с селективным слуховым вниманием, пол слушателя также может играть определенную роль. Многочисленные поведенческие и нейрофизиологические данные свидетельствуют о половых различиях в когнитивных задачах, особенно связанных с функцией внимания (Bosco et al., 2004; Gur et al., 1999, 2000; Hyde, 2016; Ingalhalikar et al., 2014; Михайлова и др., 2022). Поведенческие (Evans, Hampson, 2015; Hansen, 2011) и нейровизуализационные исследования с использованием различных методов (Neuhaus et al., 2009; Ramos-Loyo et al., 2016; Steffensen et al., 2008; Weiss et al., 2003) выявили половые различия в задачах с различными требованиями к вниманию. И, наконец, известно, что на протяжении всего развития и в зрелом возрасте женщины в среднем превосходят мужчин в вербальных способностях.
Направленное внимание – это способность удерживать внимание на значимых для задачи стимулах в присутствии отвлекающих факторов. Предполагается, что направленное внимание представляет собой гибкую мультимодальную систему. Разделение значимой и незначимой информации в ней может происходить на разных этапах переработки. Оно может происходить на раннем этапе, если между значимой и незначимой информацией существуют сенсорные отличия. Если таковых нет, разделение может происходить позднее, но при этом потребовать вовлечения большего количества ресурсов (Olguin et al., 2018). В случае слухового восприятия на ранней стадии переработки звуковые потоки могут быть быстро разделены за счет различия громкости или частоты основного тона. На более позднем этапе – за счет синтаксической и семантической информации (Bronkhorst, 2015).
Один из методов изучения селективного слухового внимания предполагает погружение испытуемого в парадигму дихотического прослушивания. Первым использовал метод дихотического прослушивания в области когнитивных исследований К. Черри более полувека назад (Cherry, 1953; Cherry, Taylor, 1954). Участникам его исследования предлагалось прослушать одновременно два потока слуховых сообщений через наушники и повторить одно сообщение, передаваемое в одно ухо, игнорируя другое сообщение. Он обнаружил, что испытуемые способны успешно следить за целевым сообщением, когда оно определено уникальной отличительной особенностью: местоположением (т.е. когда, например, дано указание “следить за голосом, подаваемым в правое ухо”), тембром голоса (т.е. “следить за голосом женщины и игнорировать голос мужчины”).
Существует ряд моделей или теорий селективного внимания, объясняющих механизм отбора целевой и подавления нецелевой информации. Наиболее состоятельной из них является теория перцептивной нагрузки (Lavie, 2005). Согласно этой теории ресурсы внимания человека, во-первых, ограничены; во-вторых, при обработке любой поступающей сенсорной информации происходит полное их включение в процесс. Следовательно, в условиях, когда основная задача не является чрезмерно сложной, вполне могут оказаться свободные ресурсы внимания для обработки других стимулов (таких как нерелевантный отвлекающий поток в дихотической задаче прослушивания). Лави утверждала, что при таких условиях “низкой нагрузки” может наблюдаться поздний отбор целевой информации. Тем не менее, если нагрузка основной задачи увеличивается (например, с увеличением сложности или скорости сообщения), тогда будет выделяться больше ресурсов на ее обработку, и, следовательно, меньше остаточных ресурсов внимания будет доступно для обработки других слуховых раздражителей. В условиях такой “высокой нагрузки” отбор будет происходить на относительно ранней стадии обработки информации (Lavie, 2005).
Степень когнитивной нагрузки возможно регулировать с помощью изменения сложности маскирующего сигнала. В работе Аллен с соавт. (Allen et al., 2011) было показано, что восприятие целевого стимула улучшалось, если маскирующая речь предъявлялась через предварительно обозначенный динамик, то есть тогда, когда слушателю заранее сообщали пространственные параметры маскера. Предварительный опыт общения с человеком также влиял на успешность выделения целевой слуховой информации. Целевой голос легче распознавался испытуемыми, если в качестве маскирующей речи использовался голос их собственного супруга по сравнению с голосами незнакомых им людей того же возраста и пола (Johnsrude et al., 2013). Известно, что эффективность распознавания при прослушивании снижается при наличии схожих речевых характеристик у маскирующего и целевого сигнала. В частности, пол диктора оказывает влияние на сложность маскировки. При равной громкости маскирующей и целевой фраз последняя наиболее понятна в случае их произнесения говорящими разного пола; если же пол дикторов одинаковый, то распознаваемость падает почти на 30%, а если обе фразы произносил один и тот же говорящий, то снижается еще примерно на ту же величину (Brungart et al., 2001). Активация нейрональных структур также зависит от сложности маскировки: в случае, если дикторы различны по полу, то наблюдается прирост активации (по сравнению с контрольными условиями) в зоне Вернике (22-е поле Бродмана), а если диктор был один и тот же, то наблюдается дополнительная активация в лобной ассоциативной коре: билатеральные прецентральные извилины, средние лобные извилины, поясные извилины, лобные покрышки (поля Бродмана 6, 9/46, 32, 13/47 соотв.), что свидетельствует о необходимости углубленной семантической, синтаксической и просодической обработки, а также усиленного избирательного внимания при увеличении сложности восприятия (Brungart et al., 2001; Nakai et al., 2005).
Целью настоящей работы был поиск половых различий паттернов активности головного мозга (по данным фМРТ) при дихотическом прослушивании с различным уровнем перцептивной нагрузки.
МЕТОДИКА
Испытуемые. В эксперименте приняли участие 20 здоровых испытуемых (9 мужчин, 11 женщин) без признаков заболеваний центральной или периферической нервной системы. Средний возраст испытуемых составил 34.25 ± 1.51 года (здесь и далее в качестве разброса значений приведена стандартная ошибка среднего). Все испытуемые дали письменное согласие на участие в исследовании с регистрацией функциональной МРТ (фМРТ). Исследование было одобрено этическим комитетом ИВНД и НФ РАН.
Оценка слуха, внимания, рабочей памяти, ведущей руки. Перед началом эксперимента все испытуемые прошли нейропсихологическое тестирование для оценки объема кратковременного и долговременного запоминания, а также коэффициента запоминания по методике “Заучивание 10 слов” (Лурия, 1962), функции внимания с помощью буквенных таблиц Шульте (Римский, Римская, 1995). Оценивали среднее время выполнения задачи по 4 таблицам. Определение ведущей руки выполнялось при помощи сенсибилизированного опросника рукости у подростков и взрослых (Чуприков, 1985). Для оценки ведущего уха испытуемым предлагали расслышать звукозапись, проигрываемую на мобильном телефоне на минимальной громкости. Отмечали, к какому уху испытуемый в первую очередь подносит устройство. Для оценки слуха у испытуемых использовался медицинский камертон.
Стимулы. В качестве стимулов использовали фрагменты аудиозаписи из произведений А. Куприна “Гранатовый браслет” и “Молох”, произносимые 3 дикторами (2 мужчин, 1 женщина). Отрывки текста внутри эксперимента не повторялись. Целевые стимулы были представлены фрагментами, записанными одним и тем же диктором мужского пола. Маскирующими стимулами (дистракторами) были записи отрывков из тех же произведений, произносимых голосом того же диктора, либо другого диктора мужского пола, либо диктора женского пола. Стимулы предъявлялись с помощью наушников и микрофона (OptoActive, Ltd., Израиль) с системой активного шумоподавления.
Условия и процедура фМРТ-исследования. В ходе исследования испытуемым необходимо было выполнить задачу направленного внимания в процессе дихотического прослушивания описанных выше фрагментов слитной речи. Для предъявления заданий использовали блочный дизайн эксперимента. В каждом блоке, длительностью 38–45 секунд, в каждое ухо предъявлялся для прослушивания единый по смыслу фрагмент текста. Использовали 4 типа блоков в зависимости от типа дистрактора: (1) идентичный поток в оба канала (S0); (2) диктор другого пола (S1); (3) диктор того же пола (S2); (4) тот же диктор (S3). На рис. 1 представлены спектрограммы предъявленных стимулов (на отрезке 35 секунд). Целевой стимул предъявлялся по каждому каналу с равной вероятностью. В начале каждого блока по предварительной команде “левое” или “правое” испытуемый переводил внимание в нужную сторону и старался воспринимать только указанный канал информации, игнорируя второй. После каждого блока имела место пауза длительностью 20 секунд, во время которой испытуемый повторял 4 последних слова целевого сообщения. Всего в ходе исследования было предъявлено 40 блоков в псевдослучайном порядке. Общая длительность исследования составила 44 минуты. Испытуемые были проинструктированы лежать расслабленно, не двигаться.
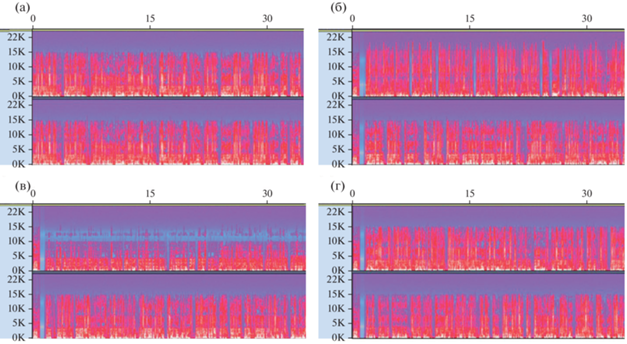
Рис. 1.
Пример спектрограмм стимульного материала (на отрезке 35 секунд) различного уровня сложности. На (а) – идентичный поток в оба канала (S0); (б) – маскировка голосом диктора противоположного пола (женский) (S1); (в) – маскировка голосом диктора того же пола (мужской) (S2); (г) – маскировка голосом того же диктора (S3). Ось х – время (с), ось y – частота (кГц). Fig. 1. Example spectrograms of stimulus material (on a segment of 35 seconds) of different levels of complexity. On (a) – identical flow to both channels (S0); (б) – masking with the voice of the speaker of the opposite sex (female) (S1); (в) – masking with the voice of the speaker of the same sex (male) (S2); (г) – masking with the voice of the same speaker (S3). The x-axis is time (s), the y-axis is frequency (kHz).
Перед фМРТ-сканированием испытуемые прослушали две тестовые звукозаписи. Используя специально разработанный опросник, отмечали понимание смысла и запоминание деталей целевого сообщения, подаваемого в правое (первая звукозапись) и левое (вторая звукозапись) ухо.
По окончании сканирования испытуемые заполняли специально разработанную анкету на понимание смысла целевого стимула (об опроснике не предупреждали заранее, для уменьшения влияния процессов запоминания). В опроснике два вопроса были по тексту канала целевого стимула, еще два – по тексту канала дистрактора, и один – по стимулу нулевой сложности (один канал в обоих наушниках). Отмечалось также, к какому наушнику (левому или правому) было легче прислушиваться и диктора какого пола было легче игнорировать, причем было предложено 4 варианта ответа (кроме основного выбора, были “одинаково легко” и “одинаково сложно”).
Регистрация фМРТ и статистический анализ данных. Функциональные и анатомические изображения были получены на томографе 3.0 T Siemens Verio (Siemens, Ltd., Германия) с двенадцатиканальной головной катушкой. Для сбора функциональных данных использовалась последовательность со следующими характеристиками: TR – 1000 мс, задержка – 0 мс, TE – 35 мс, толщина среза – 3 мм, матрица – 64 × 64, размер вокселя в плоскости – 1 × 1 мм. Исследование включало сбор 1240 измерений (объемов) для каждого испытуемого. Общая продолжительность функционального сканирования составила 21 минуту. Для получения анатомического изображения в сагиттальной плоскости использовалась последовательность Т1 MP RAGE (TR – 1900 мс, TE – 3.4 мс, 174 среза, толщина среза – 1 мм, матрица – 256 × 256, размер вокселя – 1 × 1 × 1 мм. Участников проинструктировали расслабиться и лежать неподвижно.
Данные обрабатывались с помощью пакета статистической обработки SPM12 (http://www.fil.ion.ucl.ac.uk/spm/) на платформе MATLAB (версия 2019b; MathWorks). Процедура предварительной обработки включала выравнивание функциональных изображений (коррекция движения), корегистрацию, сегментацию структурных данных, нормализацию в стандартное стереотаксическое пространство Монреальского неврологического института (MNI) и пространственное сглаживание с помощью гауссова ядра с полной шириной на половине максимума 8 мм. Статистические параметрические карты для фМРТ строились с использованием общей линейной модели (“Welcome.Trust.Centre.ForNeuroimaging:http://Www.Fil.Ion.Ucl.Ac.Uk/-Spm,” n.d.).
Для каждого испытуемого мы рассчитали 3 контраста (индивидуальные разностные карты), соответствующие 3 типам заданий, по сравнению с базовым уровнем активации (недихотическим прослушиванием) (контрасты S1-S0, S2-S0, S3-S0). При этом вклад артефактов движения учитывали путем введения соответствующих параметров (6 параметров) в модель. Далее индивидуальные разностные карты анализировали с применением смешанного дизайна дисперсионного анализа (RM ANOVA 2 × 3) с учетом фактора межгрупповой вариабельности Пол и фактора внутригрупповой вариабельности Сложность (3 уровня сложности). Post-hoc анализ проводился с помощью попарного t-теста. Множественный регрессионный анализ использовался для выявления взаимосвязи между паттернами активации и эффективностью воспроизведения текста, а также с результатами нейропсихологического тестирования. Результаты представлены с порогом статистической значимости p-value для групповой вероятности ошибки (FWE – family-wise error) < 0.05 на уровне кластера (на уровне вокселя p < 0.001). При анализе связи между паттернами активации и данными опросников вводилась поправка на множественные корреляции. Принимались результаты с pFWE < 0.003. Анатомические области мозга, включающие активированные кластеры, были определены с помощью атласа Neuromorphometrics, встроенного в SPM12. Области мозга, включающие менее 5% активированных кластеров, не приводятся в таблицах.
При оценке данных нейропсихологических тестов и опросников использовали t-тест для определения межгрупповых различий. Анализ проводили с использованием программного обеспечения STATISTICA 12 (StatSoft).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Результаты оценки слуха, внимания, рабочей памяти, ведущей руки. Опрос и инструментальная оценка показали, что все участники исследования не имели нарушений слуха. У 12 испытуемых ведущим было правое ухо, у 6 – левое. У двоих оценить ведущее ухо не представилось возможным. Согласно данным сенсибилизированного опросника рукости все испытуемые, принявшие участие в исследовании, были правшами. По данным анализа параметров рабочей памяти и внимания по всей группе установлено, что средний объем кратковременного запоминания составил 6.5 ± 0.39 слов, объем долговременного запоминания – 8.45 ± 0.35 слов, коэффициент запоминания – 0.83 ± 0.02, а время выполнения задачи на внимание – 35.98 ± 2.16 с. Показаны значимые межгрупповые различия. У мужчин по сравнению с женщинами отмечены более высокие показатели коэффициента запоминания (T(18) = 3.1, p < 0.01, 0.89 ± 0.03 – мужчины, 0.78 ± 0.02 – женщины) и времени выполнения задачи на внимание (T(18) = 2.33, p < 0.05, 40.51 ± 3.19 – мужчины, 31.45 ± 2.24 – женщины). По результатам анкетного опроса на понимание текста различия не были выявлены.
Результаты анализа фМРТ-исследования. В ходе 2-факторного дисперсионного анализа выявлен значимый эффект фактора Пол (pcorr < < 0.05) и фактора Сложность (рис. 2, табл. 1). Значимый эффект фактора Пол (F(1,34) > 32.61, pcorr < 0.05) был обнаружен в области височной доли, пре- и постцентральной извилин, затылочной области, а также в области левой лобной доли. Значимый эффект фактора Сложность (F(1,34) = 27.38, pcorr < 0.05) был обнаружен в правых пре- и постцентральной извилинах.
Рис. 2.
Результаты дисперсионного анализа (RM ANOVA 2 × 3) групповых активационных карт с факторами Пол и Сложность. Обозначения: R – правое полушарие, L – левое полушарие. Fig. 2. Results of analysis of variance (RM ANOVA 2 × 3) of group activation maps with factors Gender and Complexity. Notes: R – right hemisphere, L – left hemisphere.
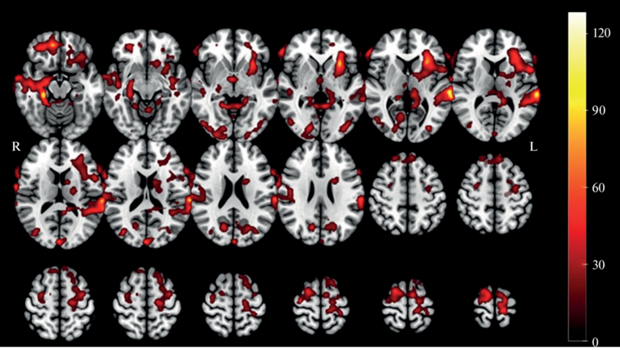
Таблица 1.
RM ANOVA активационных карт при дихотическом прослушивании различного уровня сложности Table 1. RM ANOVA activation maps in dichotic listening at different levels of complexity
| Структуры мозга, входящие в состав кластера | Полушарие | Объем кластера (воксели) |
Координаты локального максимума |
PFWE-corr | F(1,34) | Направление эффекта |
||
|---|---|---|---|---|---|---|---|---|
| x | y | z | ||||||
| Эффект фактора “Пол” | ||||||||
| Верхняя височная извилина Височная площадка Средняя лобная извилина |
ЛП | 2050 | –66 | –28 | 5 | <0.0001 | 133.80 | Ж > М |
| Прецентральная извилина Прецентральная извилина, средний сегмент |
ЛП | 828 | –9 | –25 | 77 | <0.0001 | 126.61 | Ж > М |
| Островок Скорлупа Лобная покрышка |
ЛП | 1440 | –30 | 17 | –1 | <0.0001 | 104.53 | М > Ж |
| Левый клин Правый клин Левый затылочный полюс |
ЛП, ПП | 79 | 3 | –94 | 17 | 0.03 | 81.57 | Ж > М |
| Покрышечная часть нижней лобной извилины Прецентральная извилина |
ЛП | 81 | –60 | 17 | 29 | 0.03 | 48.35 | Ж > М |
| Медиальная затылочная и звилина Верхняя затылочная извилина |
ПП | 319 | 36 | –76 | 17 | <0.0001 | 42.41 | М > Ж |
| Клин Предклинье |
ПП | 112 | 15 | –70 | 23 | 0.007 | 41.61 | М > Ж |
| Нижняя затылочная извилина | ЛП | 92 | –42 | –79 | –1 | 0.02 | 39.88 | М > Ж |
| Предклинье, Клин, Верхняя затылочная извилина |
ЛП | 206 | –24 | –70 | 23 | 0.0002 | 32.61 | М > Ж |
| Эффект фактора “Сложность” | ||||||||
| Прецентральная извилина Постценральная извилина |
ПП | 97 | 48 | –7 | 41 | 0.01 | 27.38 | (S1-S0) > (S2-S0), (S1-S0) > (S3-S0) |
| Взаимодействие факторов “Пол” × “Сложность” | ||||||||
| Нет значимого эффекта | ||||||||
Обозначения: ЛП – левое полушарие, ПП – правое полушарие; M – мужчины, Ж – женщины; S – сложность (1 – маскировка голосом диктора противоположного пола (женский); 2 – маскировка голосом диктора одноименного пола (мужской); 3 – маскировка голосом того же диктора). Объем кластера приведен в вокселях. Координаты локального максимума приведены для MNI-пространства.
ЛП – left hemisphere, ПП – right hemisphere; M – male, Ж – female; S – complexity (1 – masking with the voice of the speaker of the opposite sex (female); 2 – masking with the voice of the speaker of the same sex (male); 3 – masking with the voice of the same speaker). Cluster volume is given in voxels. Coordinates of local maximum are given for MNI-space.
Статистически значимого взаимодействия факторов обнаружено не было.
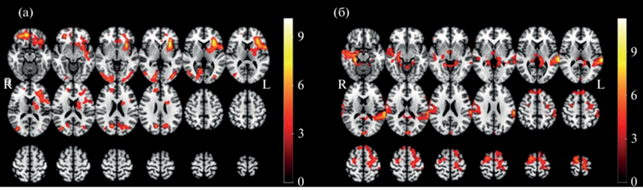
На рис. 3 приведены карты, полученные при post-hoc анализе для двух вариантов контраста: “мужчины > женщины” и “женщины>мужчины”. Как видно, зоны значимого контраста “мужчины > женщины” (T(34) > 5.09, pcorr < 0.05) локализованы в передней инсуле, скорлупе, фронтальной покрышечной области, в затылочной коре, клине и предклинье, надкраевой и ангулярной коре (рис. 3 (а)). Напротив, зоны значимого контраста “женщины > мужчины” (T(34) > 6.95, pсorr < 0.05) локализованы преимущественно в височных отделах коры, а также в прецентральной извилине (рис. 3 (б)).
Рис. 3.
Результат post-hoc анализа для фактора Пол. На (а) – контраст “мужчины > женщины”, (б) – контраст “женщины > мужчины”. Обозначения: R – правое полушарие, L – левое полушарие. Fig. 3. Results of the post-hoc analysis for the Gender factor. On (a) – the “male > female” contrast, (б) – the “female > male” contrast. Notes: R – right hemisphere, L – left hemisphere.
Список структур, для которых обнаружены значимые различия, приведен в табл. 1. Представлен состав кластеров, размеры, их полушарная принадлежность, координаты их локального максимума, направленность половых различий и их статистическая достоверность. Как видно на рис. 2, половые различия больше выражены в левом полушарии.
Для фактора Сложность обнаружен значимый эффект в области пре- и постцентральной извилины справа. В ходе дальнейшего анализа было выявлено, что активация структур головного мозга при предъявлении маскировки голосом диктора противоположного пола (самая легкая задача, за исключением недихотического прослушивания) (контраст S1-S0) выше в пре- и постцентральной извилинах и теменной коре относительно активации в ответ на предъявление маскировки тем же диктором (самая сложная задача) (контраст S3-S0) (T(34) > 4.58, pсorr < 0.05) (табл. 2). Также эта активация оказалась выше по сравнению с активацией на промежуточное по сложности задание (маскировка диктором того же пола) (контраст S2-S0) в области пре- и постцентральной извилин и средней лобной извилины справа и задних (теменная и затылочная) областей коры справа (T(34) > 4.12, pсorr < 0.05) (табл. 2).
Таблица 2.
T-тест активационных карт на сложность маскировки Table 2. T-test of activation maps for masking complexity
| Структуры мозга, входящие в состав кластера | Полушарие | Объем кластера (воксели) |
Координаты локального максимума |
PFWEcorr | T(34) -value | ||
|---|---|---|---|---|---|---|---|
| x | y | z | |||||
| Маскировка диктором другого пола > Маскировка тем же диктором | |||||||
| Прецентральная извилина Постценральная извилина |
ПП | 146 | 48 | –7 | 41 | 0.05 | 5.23 |
| Верхняя теменная долька Угловая извилина |
ЛП | 90 | –24 | –67 | 56 | 0.03 | 4.58 |
| Маскировка диктором другого пола > Маскировка диктором того же пола | |||||||
| Прецентральная извилина Постценральная извилина |
ПП | 88 | 48 | –4 | 44 | 0.03 | 5.87 |
| Прецентральная извилина Средняя лобная извилина |
ПП | 147 | 33 | –1 | 41 | 0.004 | 5.30 |
| Верхняя теменная долька | ЛП | 191 | –21 | –67 | 56 | 0.001 | 4.49 |
| Верхняя затылочная извилина Клин |
ЛП | 89 | –15 | –82 | 20 | 0.03 | 4.12 |
Сравнение активационных карт на 3 вида задач относительно недихотического прослушивания представлено на рис. 4.
Рис. 4.
Сравнение групповых активационных карт на 3 вида заданий: (а) – контраст “маскировка голосом диктора противоположного пола (женский) > недихотическое прослушивание” (S1-S0); (б) – “маскировка голосом диктора одноименного пола (мужской) > недихотическое прослушивание” (S2-S0); (в) – “маскировка голосом того же диктора > недихотическое прослушивание” (S3-S0). Обозначения: R – правое полушарие, L – левое полушарие. Fig. 4. Comparison of group activation maps for 3 types of tasks: (a) – contrast “masking with the voice of the speaker of the opposite sex (female) > non-dichotic listening” (S1-S0); (б) – “masking with the voice of the speaker of the same sex (male) > non-dichotic listening” (S2-S0); (в) – “masking with the voice of the same speaker > non-dichotic listening” (S3-S0). Notes: R – right hemisphere, L – left hemisphere.
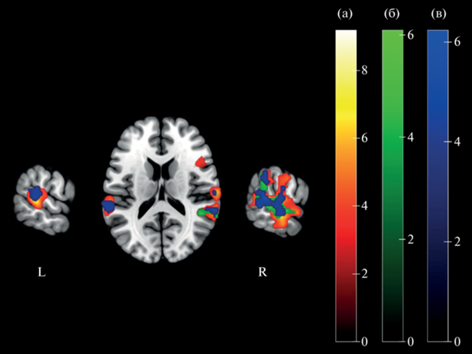
Как видно на рисунке, наибольшая по объему активация наблюдалась на задание с маскировкой голосом диктора противоположного пола (женским голосом) (контраст S1-S0). Эта активация наблюдалась с двух сторон в области верхней височной и теменно-височной коры больше справа (на рисунке представлена красным цветом). Активация на задание с маскировкой голосом диктора одноименного пола (контраст S2-S0) наблюдалась в тех же областях, но только справа (на рисунке представлена зеленым цветом). Активация на задание с маскировкой голосом того же диктора (контраст S3-S0), так же как и в первом случае, была двусторонней, хотя значительно меньшей по объему (на рисунке представлена синим). Во всех случаях преобладала правосторонняя активация, включавшая область верхней височной извилины и теменно-височного сочленения.
Анализ связи между паттернами активации и данными опросников (множественная регрессия). Корреляционный анализ активационных карт и данных опросников на предмет понимания текста не выявил статистически значимых взаимодействий.
Групповой анализ связи корковых активаций и данных нейропсихологического тестирования рабочей памяти выявил отрицательную корреляцию между объемом кратковременной рабочей памяти и BOLD-ответом в областях: правой височно-теменной коры [48 -52 11], размер кластера – 57, (T(6) = 15.64, pFWEcorr = 0.005); правой надкраевой извилины [48 -40 35], размер кластера – 90, (T(6) = 14.09, pFWEcorr = 0.000); левой и правой язычной извилины, левой шпорной коры [-3 -64 2], размер кластера – 92, (T(6) = 13.88, pFWEcorr = 0.000); левого клина и предклинья, левой верхней затылочной извилины [-18 -76 23], размер кластера – 63, (T(6) = = 12.69, pFWEcorr = 0.003), – для контраста (S1-S0) на задание с маскировкой голосом диктора противоположного пола (женским голосом). Для контраста (S2-S0) на задание с маскировкой голосом диктора одноименного пола подобная связь выявлена в области правого клина и предклинья [12 –70 35], размер кластера – 39, (T(6) =14.35, pFWEcorr = 0.045). Для объема долговременной памяти была получена положительная корреляция с BOLD-ответом (задание с маскировкой голосом диктора противоположного пола) в области правых пре- и постцентральной извилин [42 -10 53], размер кластера – 39, (T(6) = 10.61, pFWEcorr = 0.030).
Положительная корреляция времени выполнения задачи на внимание с BOLD-ответом в области правой постцентральной и верхней теменной области [24 -34 53], размер кластера – 61, (T(6) = 11.29, pFWEcorr = 0.003) наблюдалась для контраста S1-S0. Для контраста S3-S0 аналогичная корреляция выявлена в области правых надкраевой и угловой извилин [57 -49 32], размер кластера – 59, (T(6) = 10.75, pFWEcorr = 0.002). Для контраста S2-S0 такого взаимодействия выявлено не было.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В настоящей работе проведена оценка половых различий системы слухового селективного внимания мозга человека путем сравнения пространственной локализации зон активации у мужчин и женщин (по данным фМРТ) в ответ на прослушивание слитной речи с различным уровнем перцептивной нагрузки. Оценивали влияние половых различий, а также сложность задания в зависимости от типа дистрактора (диктор другого пола, диктор того же пола, тот же диктор). В качестве базового уровня активации использовали простое прослушивание аналогичного текста (идентичный поток в оба канала).
Половые различия. Половые различия активационных паттернов при решении задач на селективное слуховое внимание при прослушивании слитной речи наблюдались в широком ряде областей правого и левого больших полушарий головного мозга.
В группе женщин относительно группы мужчин активация преобладала в области левой верхней височной извилины, левых пре- и постцентральной извилин, левой нижней и средней лобной извилин, левом затылочном полюсе и области клина с двух сторон. В группе мужчин относительно женщин преобладала активация в области левого островка, скорлупы и лобной покрышки, левых и правых затылочных областей, клина и предклинья.
В исследовании (Falkenberg et al., 2011) было показано, что процессы когнитивного контроля в слуховой модальности задействуют две нейрональные сети мозга: лобно-теменную сеть, включающую премоторную область, переднюю поясную кору, нижнее лобное соединение, инсулу и нижнюю теменную дольку; сеть, включающую верхнюю височную и постцентральную извилины. Причем первая сеть была задействована в условиях с высокими требованиями к когнитивному контролю и, таким образом, рассматривалась как сеть когнитивного контроля “сверху вниз”. Вторая сеть была задействована в условиях с низкими требованиями к когнитивному контролю.
В нашей работе в группе женщин наибольший объем преобладающей активации наблюдался в височной области. В группе мужчин наибольшая разница по объему активации имела место в островковой, покрышечной и области скорлупы, т.е. в узлах сети “сверху вниз”. В совокупности с данными (Falkenberg et al., 2011) возможно предположить, что в целом экспериментальная задача на дихотическое прослушивание слитной речи для женщин была более легкой, чем для мужчин.
Отдельно стоит отметить, что у женщин выявлен более высокий уровень активации левых пре- и постцентральной извилин (сенсомоторной коры). Эти данные подтверждают немногочисленные свидетельства, что эта область, являясь генератором мю-ритма, и будучи частью дорсального пути, обеспечивает динамическую сенсомоторную когнитивную поддержку (например, внимание и рабочую память) для слухового восприятия (Liu et al., 2017; New et al., 2015; Poeppel, Hickok, 2004) и демонстрирует половой диморфизм в задачах слежения за речевым потоком (Thornton et al., 2019).
Полученные в настоящей работе результаты также свидетельствуют о большей (левостронней) латерализации процесса у женщин. В литературе имеются сведения о половых различиях при прослушивании слитной речи, в том числе предъявленной дихотически, касающиеся и латерализации (Ruytjens et al., 2007; Voyer, 2011). Например, в работе Кансаку с соавт. (Kansaku et al., 2000) при прослушивании слитной речи была выявлена бόльшая активация верхней и средней височных извилин в группе мужчин, в то время как у женщин наблюдалась более низкая билатеральная активация данной области. В другой же работе (Kaiser et al., 2007) отмечается большая левосторонняя активация нижней лобной извилины (44-е поле Бродмана) в группе женщин, у мужчин была отмечена распределенная билатеральная активация в области 45-го поля Бродмана.
Сложность задания. Паттерны активации менялись в зависимости от пола маскирующего диктора и, соответственно, сложности задания. Так, при предъявлении маскировки голосом диктора женского пола возрастал уровень метаболической активности в правых пре- и постцентральной извилинах и левой теменной коре относительно задания с маскировкой голосом того же диктора. Также метаболическая активность коры головного мозга при предъявлении маскировки голосом диктора женского пола превосходила таковую при предъявлении маскировки голосом диктора одноименного пола в областях правых пре- и постцентральной извилин, правой средней лобной извилины, левой верхней теменной дольки, левой верхней затылочной извилины и клина.
Известно, что эффективность распознавания речи снижается при наличии схожих речевых характеристик у маскирующего и целевого сигнала. При равной громкости маскирующей и целевой фразы последняя наиболее понятна в случае их произнесения говорящими разного пола, что сопровождается увеличением уровня метаболической активности в зоне Вернике (Brungart et al., 2001; Nakai et al., 2005). В случае, если диктор и для целевого, и для маскирующего потока один и тот же, наблюдается дополнительная активация в лобной ассоциативной коре: билатеральные прецентральные извилины, средние лобные извилины, поясные извилины, лобные покрышки (поля Бродмана 6, 9/46, 32, 13/47 соотв.), что, по мнению авторов, свидетельствует о необходимости углубленной семантической, синтаксической и просодической обработки, а также усиленного избирательного внимания при увеличении сложности восприятия (Nakai et al., 2005).
В настоящем исследовании вовлечение структур ассоциативной коры наблюдалось в противоположном случае – при предъявлении наиболее простого, согласно представлениям об энергетической маскировке, задания (маскировка женским голосом). Причем наибольшие отличия этой активации наблюдались от активации на задание с маскировкой голосом диктора того же пола, наименьшие – для активации в ответ на задание с маскировкой голосом того же диктора. То есть возрастание сложности распознавания и соответствующее подключение дополнительных областей высокого уровня обработки информации можно представить следующим образом: задание с маскировкой диктором того же пола → задание с маскировкой тем же диктором → задание с маскировкой диктором противоположного пола (женским голосом).
Выявленную сложность и “ресурсозатратность” задания с маскировкой женским голосом можно объяснить эволюционными причинами. Так, в исследовании с использованием модели “коктейльной вечеринки” (“cocktail party”) было установлено, что младенцы способны эффективнее прислушиваться к женскому голосу, когда в качестве маскировки используется мужской (Newman, Jusczyk, 1996).
Корреляция с нейропсихологическими данными. Корреляционный анализ BOLD-ответа и данных нейропсихологического тестирования выявил связь с объемом кратковременной и долговременной рабочей памяти, а также со временем выполнения задачи на внимание. В первом случае корреляция была отрицательной, в остальных двух – положительной.
Известно, что индивидуальные различия объема рабочей памяти коррелируют с измеримыми различиями в способности человека избирательно фокусировать слуховое внимание на определенном слуховом потоке (Conway et al., 2001). Эти данные, полученные впервые Конвеем с соавт. (2001), в настоящее время подтверждены рядом других исследований (Beaman, 2004; Berti, Schröger, 2003; Dalton et al., 2009; Muller-Gass, Schröger, 2007). Вопрос, почему именно слуховое селективное внимание снижается, когда ресурсы рабочей памяти (по какой-либо причине) ограничены, еще не решен до конца. Есть мнение (De Fockert et al., 2001), что способность участника помнить, что является его текущим заданием, напрямую влияет на эффективность концентрации внимания на определенном потоке (Lavie, De Fockert, 2005).
В настоящем исследовании было установлено, что чем меньше объем кратковременной памяти, тем больше активировались зоны правой височно-теменной коры, правой надкраевой извилины, левой и правой язычной извилины, левой шпорной коры, левого клина и предклинья, левой верхней затылочной извилины. А больший объем долговременной рабочей памяти был связан с большей активацией в области правых пре- и постцентральной извилин при выполнении задания дихотического прослушивания.
Полученные нами результаты соответствуют данным других нейровизуализационных исследований. Так, например, (Todd et al., 2005) показали, что загрузка буфера рабочей памяти приводит к подавлению нейрональной активности в правой височно-теменной области. Правая височно-теменная область, как известно, составляет основную структуру в вентральной сети внимания (Corbetta, Shulman, 2002). Считается, что эта область играет решающую роль в переориентировании внимания на окружающие события, независимо от того, актуальны они в настоящее время или нет (Berti, Schröger, 2003).
Время выполнения задачи на внимание было прямо связано с увеличенным BOLD-ответом в области правой постцентральной и верхней теменной области и правых надкраевой и угловой извилин. То есть, чем больше времени было затрачено на выполнение задачи на внимание, тем выше активация в правой теменно-височной области при выполнении задания на дихотическое прослушивание слитной речи. Правая теменно-височная область, как уже говорилось выше, играет ключевую роль в задачах избирательного слежения за потоком. Так, точность восстановления речевой огибающей на основе ЭЭГ, полученной для входящего слитного речевого потока, положительно коррелирует с амплитудой устойчивых BOLD-ответов в правом височно-теменном сочленении. Эта область имеет (связанную с задачей избирательного прослушивания) функциональную связь со вторичной слуховой корой и областями лобно-теменной сети внимания, включая внутритеменную борозду и нижнюю лобную извилину (Puschmann et al., 2017).
ЗАКЛЮЧЕНИЕ
В ходе настоящей работы были установлены паттерны половых различий активации головного мозга, характерные для нагрузки на систему слухового внимания различной силы, а также выявлена их связь с параметрами кратковременной и долговременной памяти и внимания. Так, у женщин по сравнению с мужчинами преобладала активация в области левой верхней височной извилины, левых пре- и постцентральной извилин, левой нижней и средней лобной извилин, левом затылочном полюсе и области клина с двух сторон. В то время как у мужчин по сравнению с женщинами преобладала активация в области левого островка, скорлупы и лобной покрышки, левых и правых затылочных областей, клина и предклинья. Показано, что пол диктора оказывает значимое влияние на сложность задания при дихотическом прослушивании слитной речи. Показано, что при маскировке голосом диктора женского пола требуется подключение дополнительных областей высокого уровня обработки информации по сравнению с другими типами маскировки. Относительно задания с маскировкой голосом того же диктора наблюдалось увеличение уровня метаболической активности в правых пре- и постцентральной извилинах и левой теменной коре. А по сравнению с маскировкой голосом диктора одноименного пола – в областях правых пре- и постцентральной извилин, правой средней лобной извилины, левой верхней теменной дольки, левой верхней затылочной извилины и клина.
Список литературы
Лурия А.Р. Высшие корковые функции и их нарушение при локальных поражениях мозга. 1962.
Михайлова Е.С., Майорова Л.А., Герасименко Н.Ю., Кушнир А.Б., Салтыков К.А. Половые различия в рабочей памяти на простые зрительные признаки. Анализ связанных с событием потенциалов в пространстве сенсоров и дипольных источников. Журн. высш. нерв. деят. им. И.П. Павлова. 2022. 72 (6): 836–850.
Римский С., Римская Р. Альманах психологических тестов. 1995.
Чуприков А.П. Сенсибилизированный опросник для определения рукости для подростков и взрослых. Леворукость, антропоизометрия и латеральная адаптация. Справочные и аннотированные материалы к I Всесоюзной междисциплинарной школе-семинару “Охрана здоровья леворуких детей”. 1985. 128.
Allen K., Alais D., Shinn-Cunningham B., Carlile S. Masker location uncertainty reveals evidence for suppression of maskers in two-talker contexts. J. Acoust. Soc. Am. 2011. 130 (4): 2043–2053.
Beaman C.P. The irrelevant sound phenomenon revisited: What role for working memory capacity? J. Exp. Psychol. Learn. Mem. Cogn. 2004. 30 (5): 1106–1118.
Berti S., Schröger E. Working memory controls involuntary attention switching: Evidence from an auditory distraction paradigm. Eur. J. Neurosci. 2003. 17 (5): 1119–1122.
Bosco A., Longoni A.M., Vecchi T. Gender effects in spatial orientation: Cognitive profiles and mental strategies. Appl. Cogn. Psychol. 2004. 18 (5): 519–532.
Bronkhorst A.W. The cocktail-party problem revisited: early processing and selection of multi-talker speech. Attention, Perception, Psychophys. 2015. 77 (5): 1465–1487.
Brungart D.S., Simpson B.D., Ericson M.A., Scott K.R. Informational and energetic masking effects in the perception of multiple simultaneous talkers. J. Acoust. Soc. Am. 2001. 110 (5 Pt 1): 2527–2538.
Cahill L. Fundamental sex difference in human brain architecture. Proc. Natl. Acad. Sci. U. S. A. 2014. 111 (2): 577–578.
Cherry E.C. Some experiments on the recognition of speech, with one and with two ears. J. Acoust. Soc. Am. 1953. 25 (5): 975–979.
Cherry E.C., Taylor W.K. Some Further experiments upon the recognition of speech, with one and with two ears. J. Acoust. Soc. Am. 1954. 26 (4): 554–559.
Conway A.R.A., Cowan N., Bunting M.F. The cocktail party phenomenon revisited: The importance of working memory capacity. Psychon. Bull. Rev. 2001. 8 (2): 331–335.
Corbetta M., Shulman G.L. Control of goal-directed and stimulus-driven attention in the brain. Nat. Rev. Neurosci. 2002. 3 (3): 201–215.
Dalton P., Santangelo V., Spence C. The role of working memory in auditory selective attention. Q. J. Exp. Psychol. 2009. 62 (11): 2126–2132.
Evans K.L., Hampson E. Sex differences on prefrontally-dependent cognitive tasks. Brain Cogn. 2015. 93: 42–53.
Falkenberg L.E., Specht K., Westerhausen R. Attention and cognitive control networks assessed in a dichotic listening fMRI study. Brain Cogn. 2011. 76 (2): 276–285.
De Fockert J.W., Rees G., Frith C.D., Lavie N. The role of working memory in visual selective attention. Science. 2001. 291 (5509): 1803–1806.
Gur R.C., Alsop D., Glahn D., Petty R., Swanson C.L., Maldjian J.A., Turetsky B.I., Detre J.A., Gee J., Gur R.E. An fMRI study of sex differences in regional activation to a verbal and a spatial task. Brain Lang. 2000. 74 (2): 157–170.
Gur R.C., Turetsky B.I., Matsui M., Yan M., Bilker W., Hughett P., Gur R.E. Sex differences in brain gray and white matter in healthy young adults: Correlations with cognitive performance. J. Neurosci. 1999. 19 (10): 4065–4072.
Hansen S. Inhibitory control and empathy-related personality traits: Sex-linked associations. Brain Cogn. 2011. 76 (3): 364–368.
Hyde J.S. Sex and cognition: Gender and cognitive functions. Curr. Opin. Neurobiol. 2016. 38: 53–56.
Ingalhalikar M., Smith A., Parker D., Satterthwaite T.D., Elliott M.A., Ruparel K., Hakonarson H., Gur R.E., Gur R.C., Verma R. Sex differences in the structural connectome of the human brain. Proc. Natl. Acad. Sci. U. S. A. 2014. 111 (2): 823–828.
Johnsrude I.S., Mackey A., Hakyemez H., Alexander E., Trang H.P., Carlyon R.P. Swinging at a cocktail party: voice familiarity aids speech perception in the presence of a competing voice. Psychol. Sci. 2013. 24 (10): 1995–2004.
Kaiser A., Kuenzli E., Zappatore D., Nitsch C. On females’ lateral and males’ bilateral activation during language production: a fMRI study. Int. J. Psychophysiol. Off. J. Int. Organ. Psychophysiol. 2007. 63 (2): 192–198.
Kansaku K., Yamaura A., Kitazawa S. Sex differences in lateralization revealed in the posterior language areas. Cereb. Cortex. 2000. 10 (9): 866–872.
Lavie N. Distracted and confused?: selective attention under load. Trends Cogn. Sci. 2005. 9 (2): 75–82.
Lavie N., De Fockert J. The role of working memory in attentional capture. Psychon. Bull. Rev. 2005. 12 (4): 669–674.
Liu X., Li J., Gao J., Zhou Z., Meng F., Pan G., Luo B. Association of medial prefrontal cortex connectivity with consciousness level and its outcome in patients with acquired brain injury. J. Clin. Neurosci. 2017. 42: 160–166.
Muller-Gass A., Schröger E. Perceptual and cognitive task difficulty has differential effects on auditory distraction. Brain Res. 2007. 1136 (1): 169–177.
Nakai T., Kato C., Matsuo K. An FMRI study to investigate auditory attention: a model of the cocktail party phenomenon. Magn. Reson. Med. Sci. MRMS an Off. J. Japan Soc. Magn. Reson. Med. 2005. 4 (2): 75–82.
Neuhaus A.H., Opgen-Rhein C., Urbanek C., Gross M., Hahn E., Ta T.M.T., Koehler S., Dettling M. Spatiotemporal mapping of sex differences during attentional processing. Hum. Brain Mapp. 2009. 30 (9): 2997–3008.
New A.B., Robin D.A., Parkinson A.L., Eickhoff C.R., Reetz K., Hoffstaedter F., Mathys C., Sudmeyer M., Grefkes C., Larson C.R., Ramig L.O., Fox P.T., Eickhoff S.B. The intrinsic resting state voice network in Parkinson’s disease. Hum. Brain Mapp. 2015. 36 (5): 1951–1962.
Newman R.S., Jusczyk P.W. The cocktail party effect in infants. Percept. Psychophys. 1996. 58 (8): 1145–1156.
Olguin A., Bekinschtein T.A., Bozic M. Neural Encoding of Attended Continuous Speech under Different Types of Interference. J. Cogn. Neurosci. 2018. 30 (11): 1606–1619.
Poeppel D., Hickok G. Dorsal and ventral streams: a framework for understanding aspects of the functional anatomy of language. Cognition. 2004. 92 (1–2): 1–12.
Puschmann S., Steinkamp S., Gillich I., Mirkovic B., Debener S., Thiel C.M. The right temporoparietal junction supports speech tracking during selective listening: Evidence from concurrent EEG-fMRI. J. Neurosci. 2017. 37 (47): 11505–11516.
Ramos-Loyo J., Angulo-Chavira A., Llamas-Alonso L.A., González-Garrido A.A. Sex differences in emotional contexts modulation on response inhibition. Neuropsychologia. 2016. 91: 290–298.
Ruytjens L., Georgiadis J.R., Holstege G., Wit H.P., Albers F.W.J., Willemsen A.T.M. Functional sex differences in human primary auditory cortex. Eur. J. Nucl. Med. Mol. Imaging. 2007. 34 (12): 2073–2081.
Steffensen S.C., Ohran A.J., Shipp D.N., Hales K., Stobbs S.H., Fleming D.E. Gender-selective effects of the P300 and N400 components of the visual evoked potential. Vision Res. 2008. 48 (7): 917–925.
Thornton D., Harkrider A.W., Jenson D.E., Saltuklaroglu T. Sex differences in early sensorimotor processing for speech discrimination. Sci. Rep. 2019. 9 (1).
Todd J.J., Fougnie D., Marois R. Visual short-term memory load suppresses temporo-parietal junction activity and induces inattentional blindness. Psychol. Sci. 2005. 16 (12): 965–972.
Voyer D. Sex differences in dichotic listening. Brain Cogn. 2011. 76 (2): 245–255.
Voyer D., Voyer S.D., Saint-Aubin J. Sex differences in visual-spatial working memory: A meta-analysis. Psychon. Bull. Rev. 2017. 24 (2): 307–334.
Weiss E.M., Kemmler G., Deisenhammer E.A., Fleischhacker W.W., Delazer M. Sex differences in cognitive functions. Pers. Individ. Dif. 2003. 35 (4): 863–875.
Дополнительные материалы отсутствуют.
Инструменты
Журнал высшей нервной деятельности им. И.П. Павлова