

Лесоведение, 2019, № 4, стр. 304-310
Применение электрометрического метода для диагностики влияния нефтешлама на жизнедеятельность березы повислой
Ю. А. Курило a, *, А. И. Григорьев a
a Омский государственный педагогический университет
644099 Омск, наб. Тухачевского, 14, Россия
* E-mail: curilo.yu@yandex.ru
Поступила в редакцию 17.02.2018
После доработки 16.04.2018
Принята к публикации 03.04.2019
Аннотация
Исследование электрического сопротивления (импеданса) древесных растений электрометрическим методом при воздействии неблагоприятных факторов, в том числе нефтешлама, служит одним из адекватных методических подходов к оценке состояния лесных экосистем. На протяжении всего периода исследования рассмотрена зависимость между морфологическими признаками (диаметр, высота ствола) и электрическим сопротивлением в условиях одноразового нефтешламового загрязнения. При этом изучалось сезонное изменение уровня электрического сопротивления березы повислой. Сезонный ход электрического сопротивления у березы повислой представляется в виде синусоиды с максимумом в осеннее-зимнее время, снижением в период выхода из состояния покоя, незначительным повышением в летний период и постепенным увеличением в сентябре, что соответствует периоду листопада, т.е. интенсивной подготовки к зимовке. Продукты распада нефтешлама, попадая в почву, приводят к изменению электрического сопротивления прикамбиального комплекса тканей у модельных деревьев березы, тогда как в контрольной группе деревьев оно относительно стабильно и существенно ниже, чем у модельных деревьев, что приводит к подавлению обменных процессов в организме древесного растения. Электрическое сопротивление прикамбиального комплекса тканей ствола деревьев березы отражает уровень корне-листовой связи их жизнедеятельности. В связи с этим электрометрический метод эффективно использовать в качестве биофизического показателя для оценки состояния уровня обменных процессов у древесных растений.
Исследование электрического сопротивления (импеданса) (ЭС) древесных растений при действии неблагоприятных факторов, в том числе нефтешламового загрязнения, служит одним из адекватных методических подходов к оценке состояния лесных экосистем (Григорьев, 2008). Анализ работ, посвященных диагностике заболеваний древесных растений, показал, что наиболее удобным в экспериментальном отношении и объективно отражающим особенности роста и развития древесных растений является прикамбиальный комплекс тканей (ПКТ) ствола, анатомо-морфологические свойства древесины с годичными кольцами последних лет, камбиального слоя луба могут быть оценены косвенно через их ЭС. Так как у растений ведущую роль в восприятии многих биотических и абиотических факторов, а также формировании клеточного ответа – сопротивления – на их воздействие играют плазматические мембраны, в которых возникает комплекс взаимосвязанных структурно-функциональных изменений, в их число входит и реакция мембранного потенциала. Электрический заряд на клеточной мембране является существенным фактором, влияющим на распределение заряженных частиц ионов, регулирующих обменные процессы. Известно, что электрические параметры клеток тесно связаны с функциональными особенностями тканей. Наличие поляризационной емкости является характерным показателем жизнеспособности живых клеток. Способность поляризовать ток и обусловленные этим зависимость электропроводности от частоты, сдвиг фаз, электрическое сопротивление являются характерной особенностью живых нормально функционирующих клеток. Б.Н. Тарусов (1938) предположил, что по электрическим параметрам клеток возможно определять жизнеспособность тканей, поскольку всякое воздействие, приводящее к их повреждению или заболеванию, вызывает уменьшение абсолютной величины электрического сопротивления. Этот метод определения жизнеспособности тканей основан на способности живых клеток поляризовать электрический ток. Возникающий под действием тока заряд клеток и тканей изменяет поляризационную емкость, величина которой падает при снижении их жизнеспособности или отмирании. Интенсивность и характер обмена веществ в ткани выражается в величине сопротивления, так как они обусловливают концентрацию и распределение ионов в тканях (Каширо и др., 1988), в свою очередь, отметили, что электрическую модель клетки можно представить как последовательное соединение активных сопротивлений.
Вообще изучение состояния древесных растений в техногенных условиях весьма актуально и использование современных экспресс-методов диагностики состояния обосновано. В настоящее время разработано довольно много косвенных методов оценки устойчивости растений к различным стресс-факторам. Среди них наиболее перспективными представляются электрофизиологические методы, поскольку они непосредственно регистрируют характеристики клеточных мембран, тесно связанные с общей устойчивостью растений к стрессам (Кушнеренко и др., 1986; Fromm, Eschrich, 1993; Fromm, Spanswick, 2006; Krol, 2006; Davies, 2004; Зубкова и др., 2011; Катичева и др., 2013). К электрофизиологическим инструментальным методам относятся: диагностика по электрическому сопротивлению прикамбиального комплекса тканей (Положенцев и др., 1970), диагностика по электрическим потенциалам (Коловский, 1978, 1980), экспресс-метод (Голодрига и др., 1972), термоэкспресс – метод (Карасев и др., 2017). Маторкиным А.А. и Карасевой М.А. (2007) предложен метод экспресс-оценки состояния деревьев по импедансу ПКТ и температуре стволов. Нами ранее изучены биоэлектрические реакции древесных растений на различные стресс-факторы, в частности электрическое сопротивление ПКТ березы повислой (Курило, Григорьев, 2010, 2015).
В данной работе представлены результаты восьми полевых сезонов по изучению изменения уровня электрического сопротивления ПКТ березы повислой, произрастающей в условиях одноразового нефтешламового загрязнения.
ОБЪЕКТЫ И МЕТОДИКА
Объектом исследования послужили деревья березы повислой, произрастающие в естественных условиях на территории бассейна буферного пруда АО “Газпромнефть – “ОНПЗ” (г. Омск).
Был заложен опыт 1 мая 2010 г. на участке размером 10 × 35 м с внесением нефтешлама в приствольные круги диаметром 2.0 м в пределах проекции кроны одиночно стоящих модельных деревьев березы. В эксперименте было три варианта по три модельных дерева: первый вариант – контроль (группа “А”), второй вариант – с внесением нефтешлама на половину приствольного круга с дозой 3.70 кг/м2 (асимметричное нефтешламовое загрязнение) (группа “Б”), третий вариант – с внесением нефтешлама на всей площади приствольного круга с дозой 3.70 кг/м2 (симметричное нефтешламовое загрязнение) (группа “В”). Таксационная характеристика модельных деревьев березы повислой по вариантам опыта приведена в таблице 1.
Таблица 1.
Таксационная характеристика модельных деревьев березы повислой по вариантам опыта
| Вариант опыта | Амплитуда по диаметру стволов, см | Амплитуда по высоте деревьев, м |
|---|---|---|
| “А” | 5.0–7.7 | 13.0–18.0 |
| “Б” | 7.7–10.4 | 18.3–23.6 |
| “В” | 10.4 –13.1 | 23.6–29.0 |
Измерения проводили мультиметром MY-6. Измерительные иглы вводили под углом 90° относительно оси ствола, на высоте 130 см от уровня почвы (Шеверножук, 1968). При регистрации электрического сопротивления оценивали распространение сигнала с двух противоположных сторон ствола – восточной и западной.
За период исследования проведено 2014 измерений.
Результаты исследований обработаны стандартными методами вариационной статистики, корреляционного, регрессионного и дисперсионного однофакторного анализов с использованием пакета “Statistica 10”. Сравнения вариантов опыта проводились по критериям Стьюдента и Фишера.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
На протяжении всего исследовании рассмотрена зависимость между морфологическими признаками (диаметр, высота ствола) и ЭС у березы повислой в условиях нефтешламового загрязнения, сезонное изменение уровня ЭС на почвах, загрязненных нефтешламом.
В группе с асимметричным нефтешламовым загрязнением годичный прирост по диаметру составил ≈0.3 см при таксационном диаметре 11.1 см (1.30 м от поверхности почвы), за то же время в контрольной группе прирост составил ≈1.3 см. В группе с симметричным нефтешламовым загрязнением годичный прирост по диаметру был равен ≈0.7 см.
Рис. 1.
Схема расположения модельных деревьев березы на ключевом участке (“Буферный пруд” АО “Газпромнефть – ОНПЗ”)
Исследуемая площадка “Буферный пруд” АО “Газпром – ОНПЗ”
А1–А3, естественно произрастающие модельные деревья березы повислой – контрольная группа;
Б1–Б3, экспериментальная группа с внесением продуктов нефтешлама на половину приствольного круга;
В1–В3, экспериментальная группа с внесением нефтешлама по всей площади приствольного круга.
Рис. 2.
Сезонная динамика ЭС ПКТ березы повислой (“Буферный пруд” АО Газпромнефть – ОНПЗ”, 2010–2017 гг.)
*1 – Контрольная группа “А”, естественно произрастающие модельные деревья березы повислой;
2 – Экспериментальная группа “В” с внесением нефтешлама по всей площади приствольного круга;
3 – Экспериментальная группа “Б” с внесением продуктов нефтешлама на половину приствольного круга.
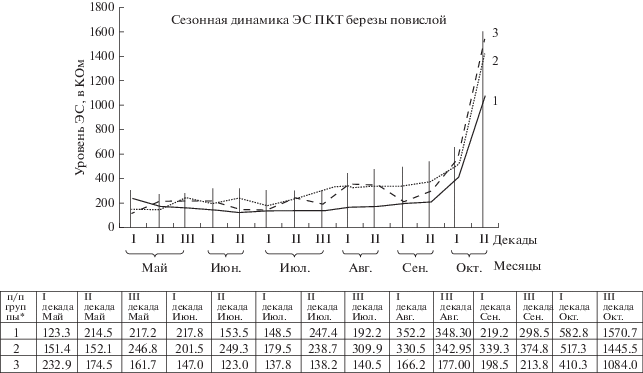
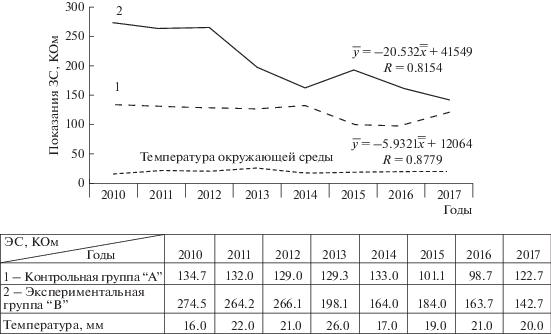
Рис. 3.
Динамика электрического сопротивления в период с 2010 г. по 2017 г.
1 – Контрольная группа “А”, естественно произрастающие модельные деревья березы повислой;
2 – Экспериментальная группа “В” с внесением нефтешлама по всей площади приствольного круга.
Зависимость прироста по диаметру и ЭС при нефтешламовом загрязнении была установлена достоверно высокая (${{\overline{\overline t} }_{{{\text{ф а к т }}}}}$ > ${{\overline{\overline t} }_{{0.05}}}$), отрицательная во всех группах (рассматривались три группы – малый, средний и большой диаметры): $\overline{\overline r} $ = 0.84 ± 0.03. Это свидетельствует о том, что деревья с минимальным диаметром имеют более высокое ЭС. На основании данного факта можно заключить, что они являются более чувствительными к нефтешламовому загрязнению.
Нами были проведены исследования по определению сезонной динамики развития ЭС у березы повислой в условиях нефтешламового воздействия.
Сезонная динамика электрического сопротивления деревьев березы повислой характеризуется рядом особенностей, связанных с прохождением морфо-физиологических фаз древесными растениями.
Измерения ЭС начали фиксировать с момента сокодвижения (первая декада апреля). В период роста растения, набухания почек наблюдается снижение измеряемых величин (конец апреля–начало мая), которое продолжается до появления листьев. Далее отмечается увеличение сопротивления в третьей декаде мая–первой декаде июня. А в июне–июле сильных скачков ЭС не наблюдалось. Стабильная картина нарушается снижением уровня ЭС с третьей декады июля по вторую декаду августа. Со второй–третьей декады августа ЭС растет. Далее с завершением вегетационного периода, что соответствует третьей декаде сентября, происходит увеличение ЭС; высокие значения сохраняются в период завершения сбрасывания (октябрь) листьев и зимой (декабрь).
Одновременно в тканях возрастает относительное содержание прочно связанной воды, параллельно снижению общей оводненности, а количество свободной воды, необходимой для протекания активных жизненных процессов, резко уменьшается. Таким образом, если подготовка к зимнему покою, повышение морозостойкости растения сопровождаются снижением активности обменных процессов (включая ионный обмен), то этим можно объяснить наблюдающееся повышение комплексного электросопротивления растительных тканей.
По результатам наших исследований сезонный ход электрического сопротивления у березы повислой в данных условиях представляется в виде синусоиды с максимумом в осеннее-зимнее время, снижением в период выхода из состояния покоя, незначительным повышением в летний период и постепенным увеличением в сентябре, что соответствует периоду листопада, т.е. интенсивной подготовки к зимовке.
Наши результаты подтверждаются также данными, опубликованными в работе (Fissenko S., Fissenko M., 2016) по сезонной динамике электрического сопротивления, полученные для березы маньчжурской и сосны обыкновенной. Причем, авторы отмечают, что ПКТ сосны обыкновенной в четыре раза выше ПКТ березы маньчжурской. Затем разница между ними стала уменьшаться и к концу вегетации почти ликвидировалась. Ранее Ю.П. Каширо с соавт. (1988) отметили, что в конце сезона вегетации при снижении температуры воздуха и почвы в растениях протекают процессы обезвоживания и в связи с этим электрическое сопротивление тканей увеличивается. В ряде исследований указывается на характер изменчивости величины импеданса в связи с состоянием дерева, разных этапов вегетационного периода и фенологического состояния (Грязькин и др., 2012).
Полученные нами экспериментальные данные по электрическому сопротивлению ПКТ стволов березы повислой за период 2010–2017 гг. показали, что в контрольной группе (группа “А”), произрастающей в естественных условиях, среднее значение уровня электрического сопротивления в течение исследуемого периода изменялось в пределах 28.7%. В экспериментальной группе (группа “В”) изменение уровня электрического сопротивления происходит по нисходящей линии зависимости и составило 44.3%.
ЗАКЛЮЧЕНИЕ
Продукты нефтешлама, попадая в почву, в экспериментальных условиях (деревья группы “В”) приводят во все годы к повышению ЭС ПКТ у модельных деревьев березы, тогда как в контрольной группе деревьев оно является относительно стабильным и существенно ниже, чем у модельных деревьев в группе “В”. Это обуславливает подавление обменных процессов в целом на древесном растении и более раннее наступление осеннего пожелтения листьев.
Следует особо отметить, что в течение трех лет после внесения нефтешлама величина ЭС устойчиво удерживалась стабильно на высоком уровне. Это позволяет судить о продолжении проявления сильного влияния нефтешламного загрязнения на состояние ПКТ у модельных деревьев березы, которое в конце третьего сезона составило всего 3% и статически недостоверно.
На четвертый год уже наблюдалось резкое снижение (на 25.55%) величины ЭС ПКТ у модельных деревьев березы. В пятый, шестой, седьмой годы эта тенденция сохранилась, снижение составило соответственно – 36.61%, 32.97% и 40.38%.
Даже на восьмой год после внесения нефтешлама наблюдаем существенное различие ЭС ПКТ в сравнении с контролем (40.67%), что указывает на отрицательное влияние нефтешлама в почве на жизнедеятельность деревьев.
Таким образом, при проведении полевых электрометрических исследований древесных растений, необходимо учитывать физиологические особенности растений, гидротермический режим воздушной среды и условия их произрастания. Поскольку биоэлектрическое сопротивление отражает уровень обменных процессов в жизнедеятельности древесных растений, возможно его эффективно и оперативно использовать в качестве биофизического показателя для диагностики состояния древесных растений в целом. В частности, биоиндикацию нефтешламового загрязнения почвенного покрова можно проводить путем оценки состояния деревьев по увеличению показателя ЭС ПКТ ствола, отражающего снижение обменных процессов и в целом нарушение корне-листовой связи.
Список литературы
Голодрига П.Я., Осипов А.В. Экспресс-метод и приборы для диагностики морозоустойчивости растений // Физиология и биохимия культурных растений. 1972. Т. 4. Вып. 6. С. 650–655.
Григорьев А.И. Эколого-физиологические основы адаптации древесных растений в лесостепи Западной Сибири. Омск: Изд-во ОмГПУ, 2008. 195 с.
Грязькин А.В. Герасюта С.М., Бернацкий Д.П., Трубачева Т.А., Ковалев Н.В. Изменчивость величины импеданса древесных пород // Известия Санкт-Петербургской лесотехнической академии. 2012. № 198. С. 11–17.
Зубкова Т.А. Мартынова Н.А., Белоусов В.М. Электрическое сопротивление структурных элементов биогеоценозов // Известия Иркутского государственного университета. Серия “Наука о Земле”. 2011. Т. 4. № 2. С. 82–89.
Карасев В.Н., Карасева М.А., Романов Е.М., Мухортов Д.И. Термоэкспресс-метод ранней диагностики физиологического состояния сосны обыкновенной // Экология. 2017. № 2. С. 92–99.
Катичева Л.А., Суворова Л.М., Шерстенева О.Н., Бушуева А.В., Глинская Е.В., Воденеев В.А. Изменение электрического сопротивления плазмалеммы клеток высшего растения при генерации вариабельного потенциала // Вестник Нижегородского университета им. Н.И. Лобачевского. 2013. № 3 (1). С. 151–154.
Каширо Ю.П., Хасанов Н.X., Дорожимн Е.М. Электрическое сопротивление тканей прикамбиального комплекса ствола у сосны обыкновенной и его диагностические возможности // Леса Урала и хозяйства в них. Свердловск: УРГУ, 1988. Вып. 14. С. 183–188.
Коловский Р.А. Биоэлектрическая реакция корней сеянцев кедра на ионный состав среды // Физиолого-биохимические механизмы роста хвойных. Новосибирск: “Наука” Сибирское отделение. 1978. С. 89–95.
Коловский Р.А. Биоэлектрические потенциалы древесных растений. – Новосибирск: “Наука” Сибирское отделение. 1980. 176 с.
Курило Ю.А., Григорьев А.И. Электрическое сопротивление как показатель устойчивости древесных растений в условиях нефтяного загрязнения // Проблемы региональной экологии. 2010. № 5. С. 111–116.
Курило Ю.А., Григорьев А.И. Изучение электрического сопротивления древесных растений в условиях нефтяного загрязнения почвы (на примере березы повислой) // Современные проблемы науки и образования. 2015. № 3; URL: http://www.science-education.ru/123-17465 (дата обращения: 18.08.2018).
Кушнеренко М.Д., Курчатова Г.П., Штефырца А.А., Печерская О.Н., Киевцова Е.В., Баштовая С.И. Комплексный метод определения жаро- и засухоустойчивости растений прибором “ТУРГОРОМЕР-1” (Т-1) // Экспресс-методы диагностики жаро- засухоустойчивости и сроков налива растений. Кишинев: “ШТИЧИЦА”, 1986. 36 с.
Маторкин А.А., Карасева М.А. Информативность импеданса прикамбиального комплекса тканей деревьев хвойных пород при диагностиках жизнедеятельности // Современная физиология растений: от молекул до экосистемы. 2007. Ч. 2. С. 265–266.
Положенцев П.А., Золотов Л.А. Динамика электрического сопротивления тканей луба сосны как индикатор изменения их физиологического состояния // Физиология растений. 1970. Т. 17. Вып. 4. С. 830–835.
Тарусов Б.Н. Электропроводность как метод определения жизнеспособности ткани // Архив ботанических наук. 1938. Т. 52. Вып. 2. С. 5–16.
Шеверножук Р.Г. Биоэлектрическая активность ели в насаждениях, методика ее измерения // Лесной журнал. 1968. № 4. С. 36–40.
Davies E. New Functions for Electrical Signals in Plants // New Phytologist. 2004. V. 161. P. 607–610.
Demidchik V., Maathuis F. Physiological roles of nonselective cation channels in plants: from salt stress to signaling and development // New Phytologist. 2007. V. 175. P. 387–404.
Fissenko S., Fissenko M. Vegetation variations of electric resistance of trees // International Scientific Ravine № 18 (28): XXVI International Scientific and Practical Conference. Boston, USA – 08 November, 2016. P. 39–43.
Fromm J., Eschrich W. Electric Signals Released from Roots of Willow (Salix viminalis L.) Change Transpiration and Photosynthesis // Plant Physiology. 1993. V. 141. P. 673–680.
Fromm J., Spanswick R. Characteristics of Action Potentials in Willow (Salix viminalis L.) // Plant Physiology. 2006. V. 163. P. 369–381.
Krol E., Dziubiñska H., Stolarz M., Trebacz K. Effects of ion Channel Inhibitors on Cold – and Electrically-Induced Action Potentials in Dionaea muscipula // Biological Plant. 2006. V. 50. P. 411–416.
Ksenzhek O., Petrova S., Kolodyazhny M. Electrical properties of plant tissues. Resistanceofamaizeleaf // Plant Physiology. 2004. № 30 (3–4). P. 61–67.
Дополнительные материалы отсутствуют.