

Лесоведение, 2019, № 5, стр. 449-464
Характеристика и перспективы сохранения сосновых лесов Московской области
Т. В. Черненькова a, b, М. Ю. Пузаченко a, b, Н. Г. Беляева a, b, И. П. Котлов b, О. В. Морозова a, b
a Институт географии РАН
119017 Москва, Старомонетный пер., 29, Россия
b Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский просп., 33, Россия
Поступила в редакцию 30.01.2019
После доработки 30.03.2019
Принята к публикации 05.06.2019
Аннотация
Изучены особенности структуры и состава лесов, сформированных с участием сосны (Pinus sylvestris), в центральной части Русской равнины (центральный, северный и западный секторы Московской области). По предварительной оценке площадь сосновых и сосново-еловых насаждений в составе лесопокрытой площади исследуемой территории составляет 16%. В работе поставлены следующие вопросы: есть ли возможность сохранения коренных сосновых сообществ в Московском регионе, и в каких ландшафтных условиях это наиболее вероятно? С целью оценки перспектив возобновления сосны в сообществах разного типа выполнен анализ динамики ее ценопопуляций. Для изучения распределения типов лесных сообществ в пределах исследуемой территории применен подход, интегрирующий наземную информацию и пространственно-распределенные данные (данные дистанционного зондирования Земли и цифровые модели рельефа). Выявлена связь сосновых сообществ разного типа с параметрами рельефа (высотой, уклонами, различными кривизнами и освещенностью), а также приуроченность к различным физико-географическим провинциям. Оценены некоторые ландшафтно-экологические метрики пространственной структуры групп ассоциаций сосновых лесов. Полученные данные уточняют существующие представления о фитоценотической структуре сообществ с участием сосны, состав которых отражает сукцессионную стадию (1), зональные черты растительности исследуемого региона (2) и приуроченность к определенным ландшафтным элементам (3). Делается вывод, что направленность восстановительных сукцессий на водораздельных поверхностях, сопровождающаяся активной демутацией еловых, в отдельных случаях – хвойно-широколиственных лесов, исключает существование сосновых и сосново-еловых сообществ в Московской области через несколько десятков лет. Небольшая доля сосновых насаждений сохранится по крутым склонам рек.
Сосна обыкновенная (Pinus sylvestris) относится к одной из основных лесообразующих пород на территории европейской части России. Существенная площадь сосновых лесов принадлежит бореальной зоне; в зоне смешанных лесов и южнее, в лесостепной и степной, они встречаются в виде изолированных массивов (Bohn, 2004; Рысин, Савельева, 2008; Кучеров, 2018). Экологический ареал сосны широк – она произрастает на почвах разного механического состава и режима увлажнения. При высокой устойчивости к некоторым стрессовым абиотическим условиям (дефициту питательных веществ, засухе, избыточному увлажнению, пирогенному фактору) на ювенильной стадии сосна не выдерживает конкуренцию со стороны мохового покрова и с трудом преодолевает слой подстилки; подрост сосны плохо развивается в условиях затенения под пологом ели, лиственных пород и кустарникового яруса (Рысин, 1964; Рязапов, Кабанов 2010).
Вопрос смены сосны елью имеет почти вековую историю и широко дискутировался в кругу лесоводов и фитоценологов. При этом большинством авторов было признано, что за исключением местообитаний с верховым типом заболачивания на торфяных почвах, а также на мощных песчаных отложениях ель однозначно вытеснит сосну. В последнем случае продолжительное существование сосняков на сухих и бедных песчаных почвах поддерживается периодически возникающими пожарами. Это дало право некоторым ученым (Морозов, 1924; Самбук, 1932; и др.) относить данный тип сосняков к коренным, что позднее получило частичное подтверждение в соответствии с теорией циклически-эрозионно-пирогенной эволюции (Санников, 1992).
Человек заселил Русскую равнину с конца оледенения (Бадер, 1936; Король, 1994–1997; Харитонов, 2010), следовательно, пожары в хвойных лесах с этого периода могли возникать не только от атмосферных электрических разрядов, но и вследствие деятельности человека. В современных условиях усиленного противопожарного контроля лесного фонда, особенно в лесах высокой природоохранной значимости, а также наличия лесокультурной практики естественные циклы аутогенных сукцессий нарушены. Таким образом, в условиях изменения естественной динамики проблема сохранения и воспроизводства современных сосновых лесов с учетом зональной, региональной специфики, а также эдафической приуроченности становится еще более актуальной. Целью данной статьи является изучение особенностей структуры и состава лесов, сформированных с участием сосны в основном древесном пологе, в центральной части Русской равнины (центральный, северный и западный секторы Московской области).
В работе дается оценка возможности сохранения сосновых лесов в Московском регионе в зависимости от ландшафтных условий. Использованы современные цифровые технологии, на необходимость применения которых в последние годы указывал в своих работах академик А.С. Исаев (Исаев, Черненькова, 2015).
ОБЪЕКТЫ И МЕТОДИКА
Московская область расположена в центральной части Восточно-Европейской равнины. В соответствии со схемой геоботанического районирования, основная часть Московской области находится в пределах зоны хвойно-широколиственных лесов, на юге региона проходит граница с широколиственной зоной (Петров, 1968; Грибова и др., 1980). Леса Московского региона отличает ряд особенностей: существенная рекреационная нагрузка вследствие высокой плотности населения; ограниченное целевое назначение лесов – в соответствии с Лесным кодексом Российской Федерации леса относятся к группе защитных; неудовлетворительное санитарное состояние и нестабильная экологическая обстановка, вызванные ограничением в проведении рубок лесных насаждений; наличие большого объема спелых и перестойных мелколиственных насаждений. Состав лесов Московской области и г. Москвы на землях лесного фонда и на землях обороны и безопасности представлен по официальным данным (Лесной план…, 2018) наполовину мелколиственными породами (березой – 41%, осиной – 9%, ольхой серой – 3%, ольхой черной – 2%), елью – 23%, сосной – 20% и широколиственными породами – 2%.
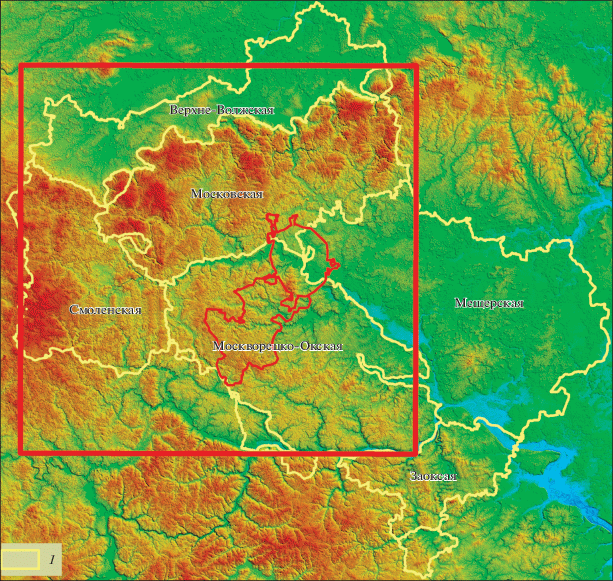
Исследуемая территория занимает бóльшую часть Московской области (центральный, северный и западный секторы) и так называемой “Новой Москвы”; ее площадь составляет 40 532 км2 (рис. 1). Особенности генезиса, геолого-морфологических и структурных характеристик в пределах изучаемой территории позволили выделить 5 крупных физико-географические провинций (ФГП): (1) Верхне-Волжскую, (2) Московскую, (3) Смоленскую, (4) Москворецко-Окскую и (4) Мещерскую (ее западная часть) (Анненская и др., 1997) (рис. 1).
Рис. 1.
Район исследований
1 – физико-географические провинции; приведены по (Анненская и др., 1997).
Сообщества с участием сосны рассматривались в составе общего ценотического разнообразия лесов территории. При классификации сообществ использован эколого-фитоценотический подход (Chernen’kova, Morozova, 2017). Выявлен полный видовой состав кустарникового, травяно-кустарничкового и мохового ярусов с оценкой проективного покрытия в процентах22. Принадлежность к культурам оценивалась на основе планов лесных насаждений (масштаб 1 : 25 000) с проверкой на местности. Для изучения организации сосняков разного типа и ценопопуляционной динамики основных видов лесообразователей (ели, сосны, дуба, липы, клена) с выявлением перспективы возобновления сосны выполнен детальный анализ состава всех ярусов сообществ33 по показателю активности видов (А) (Малышев, 1973)44.
Проанализировано 231 полное геоботаническое описание в чистых сосновых насаждениях и сообществах, где полог сосны в составе верхнего древесного яруса составлял не менее 40%. Для хранения и анализа материалов полевых геоботанических описаний использовалась созданная база данных “FORDIV” (в СУБД Access) (Свидетельство о гос. регистрации № 2 014 620 979).
Статистические методы использовали по трем направлениям: 1) при классификации для выявления особенностей флористического состава выделенных синтаксонов и их характеристик; 2) при оценке связи выделенных групп сообществ с факторами среды (в том числе пространственными переменными). Расчеты реализованы в пакетах прикладных программ (PС-ORD 5.0, STATISTICA 10, IBM SPSS Statistics); 3) при анализе пространственной структуры территориальных единиц лесного покрова в программе Fragstats.
Для оценки точности выделенных единиц применен пошаговый дискриминантный анализ. Индикаторная значимость видов оценена по методу IndVal (Dufrene, Legendre, 1997), виды со значением IndVal > 30 рассматривали как дифференцирующие. Для интерпретации экологического содержания групп применены методы непрямой ординации – неметрическое многомерное шкалирование (NMS ординация) в пакете PC-ORD 5.0 (McCune, Mefford, 2006) с использованием трансформированных (корень квадратный) данных. В качестве меры близости использовано расстояние Брея–Кёртиса (Legendre P., Legendre L., 1998; McCune, Grace, 2002). Значения факторов и экологическая характеристика сообществ оценены по шкалам Д.Н. Цыганова (1983): увлажнения (HD), обеспеченности почв азотом (NT), кислотности почв (RC), трофности (TR), освещенности (LC). Анализ различий выборок групп сообществ по факторам и по положению центроидов групп по осям ординации выполнен с помощью непараметрического критерия Манна–Уитни (Mann–Whitney U-test). Корреляция ординационных осей с экологическими факторами оценена с помощью коэффициентов ранговой корреляции Спирмена. При ординации учитывался состав подчиненных ярусов – травяно-кустарничкового и мохово-лишайникового.
Для пространственной оценки современного состава лесного покрова применен подход, интегрирующий наземную и дистанционную информацию. Использованы данные со спутников серии Landsat (сенсоры TM, ETM+, OLI и TIRS) за последние 10 лет в составе семи безоблачных сцен с марта по октябрь. Для более полного использования дистанционной информации рассчитывали индексы (NDVI, VI, NDWI и др.), являющиеся разностями и нормализованными разностями спектральных каналов съемки (Jackson, Huete, 1991). В качестве цифровой модели рельефа (ЦМР) применялись данные SRTM. Разрешение ЦМР соответствует разрешению данных дистанционного зондирования Земли (ДДЗ). На основе ЦМР рассчитаны морфометрические характеристики рельефа – уклоны, кривизны, освещенность, выпуклость и др. Для рельефа территории с помощью спектрального анализа определены иерархические уровни его организации (Котлов, Пузаченко, 2006).
Анализ закономерностей пространственного распределения выделенных синтаксонов и их пространственное отображение заключались в интерполяции классов растительности на верхние масштабные уровни путем их сопоставления с ДДЗ и ЦМР на основе обучающей выборки (Пузаченко, 2004; Пузаченко и др., 2014). Группа ассоциаций являлась базовой картографируемой единицей и использовалась в качестве группирующей переменной или обучающей выборки при проведении многомерного анализа мультиспектральных снимков. Выполнена также интерполяция отдельных характеристик растительного покрова, полученных из наземных описаний сообществ (распределение отдельных доминирующих видов по их проективному покрытию), в частности, показателя сомкнутости крон сосны. Более детально используемый подход освещен в ранних публикациях (Черненькова и др., 2015; Пузаченко, Черненькова, 2016).
Для оценки геометрических особенностей контуров и их взаимного расположения использован подход, основанный на измерении ландшафтно-экологических метрик. Ранее применимость пространственных метрик была показана для контуров почвенных (Фридланд, 1972), ландшафтных (Викторов, 1986) и геоботанических выделов (Холод, 2017). Расчет выполнен в Fragstats (McGarigal et al., 2012). В качестве базового объекта анализа взяты выделы сосновых лесов, соответствующие выделенным синтаксонам в ранге групп ассоциаций.
Метрики площади достаточно просты – площадь и доля площади группы сообществ в разрезе провинций, а также средняя площадь выделов каждой группы сообществ. Средняя площадь выдела является одним из показателей фрагментированности.
Метрика формы (SI) является индикатором сложности контуров выделов группы.
где pij – периметр выдела (м); aij – площадь выдела (м2). Метрика варьирует от 1 (квадрат) до бесконечности. В отличие от простого соотношения периметра и площади, этот показатель нечувствителен к размерам выделов.
Индекс площади экологического ядра (CAI) основан на подверженности отдельного выдела воздействию соседствующих типов ландшафта (например, загрязнение участка леса, прилегающего к дороге, или его гидроморфизация вблизи водного объекта). На основе эмпирического предположения о дистанции, на которую распространяется воздействие, индекс показывает, какая доля выдела (ядро) не подвержена воздействиям экосистем–соседей. Для вычисления метрики заранее вводится матрица парных дистанций воздействия ландшафтов друг на друга. Дистанции варьируют от 0 для морфологически гомогенных экосистем (различные типы леса) до 200 м для гетерогенных сочетаний с антропогенными территориями.
где $a_{{ij}}^{c}$ – площадь выдела, скорректированная на дистанцию воздействия соседних выделов.
Метрика контрастности границ (ECON) вычисляется по принципу, сходному с индексом площади ядра. Вместо площади за основу берется периметр выдела. Значения контрастности (неоднородности) в матрице варьируют от 0 до 1.
где pijk – длина (м) границы выдела ij, соседнего с выделом класса k; dik – неоднородность (вес контрастности границы) между выделами классов i и k, pij – периметр выдела ij.
Метрика агрегированности (ENN) – евклидова дистанция между ближайшими друг к другу выделами одной группы.
где hij – дистанция (м) до ближайшего выдела такого же класса, основанная на дистанции между границами и рассчитанная от ядра к ядру.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В составе сосновых и сосново-еловых сообществ выделены 11 групп ассоциаций, различающихся по составу растительности основных ярусов (табл. 1). Точность классификации по результатам дискриминантного анализа составила 84.5%. При этом с меньшей точностью (<60%) дискриминировались группы ассоциаций с неоднородным составом древесного (например, сосново-еловая мелкотравная) или подчиненных ярусов (например, сосновая с липой, дубом и лещиной мелкотравно-широкотравная), в значительной доле представленные культурами (около 80%). Общая доля лесных культур по доступным нам данным составила треть всех описаний.
Таблица 1.
Анализируемые группы сообществ с оценкой точности классификации по результатам дискриминантного анализа
| Группа сообществ | Группа ассоциаций | Число описаний | Доля правильной классификации, % | |
|---|---|---|---|---|
| от видового состава | от данных ДДЗ и ЦМР | |||
| Кустарничково-мелкотравно-зеленомошная | 1. Сосново-еловая кустарничково-мелкотравно-зеленомошная | 16 | 81.3 | 43.8 |
| 2. Сосновая с елью и березой кустарничково-мелкотравно-зеленомошная | 16 | 81.3 | 68.8 | |
| Мелкотравная | 3. Сосново-еловая мелкотравная | 11 | 54.6 | 63.6 |
| 4. Сосновая с елью и березой мелкотравная | 17 | 94.1 | 52.9 | |
| Мелкотравно-широкотравная | 5. Сосново-еловая мелкотравно-широкотравная | 37 | 83.8 | 30 |
| 6. Сосновая с елью и березой мелкотравно-широкотравная | 12 | 91.7 | 57.1 | |
| 7. Сосновая с липой, дубом и лещиной мелкотравно-широкотравная | 14 | 57.1 | 54.5 | |
| Широкотравная | 8. Сосново-еловая широкотравная | 55 | 78.2 | 46.5 |
| 9. Сосновая с липой, дубом и лещиной широкотравная | 34 | 85.3 | 55.9 | |
| Разнотравная | 10. Сосновая с елью и березой разнотравная | 12 | 100 | 75 |
| Кустарничково-сфагновая | 11. Сосновая с елью и березой кустарничково-сфагновая | 7 | 100 | 100 |
Дальнейший анализ состава и структуры выделенных групп осуществляли после их объединения в 6 групп сообществ55 по преобладающей экобиоморфе66 растительности наземных ярусов: кустарничково-мелкотравно-зеленомошные (1); мелкотравные (2); мелкотравно-широкотравные (3); широкотравные (4); разнотравные (5); кустарничково-сфагновые (6).
Группу сосновых сообществ кустарничково-мелкотравно-зеленомошных дифференцируют бореальные виды травяно-кустарничкового яруса Vaccinium myrtillus и Melampyrum pratense и зеленые мхи (Hylocomium splendens, Dicranum polysetum), которые имеют значения IndVal > 50 (табл. 2). Наличие молодых елей в травяно-кустарничковом ярусе указывает на активную демутацию ели.
Таблица 2.
Индексы индикаторного значения (IndVal) видов в группах сосновых сообществ
| Группа сообществ | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Кустарничково-мелкотравно-зеленомошная | Мелкотравная | Мелкотравно-широкотравная | Широкотравная | Разнотравная | Кустарничково-сфагновая | ||||||
| вид | IV | вид | IV | вид | IV | вид | IV | вид | IV | вид | IV |
| Pleurozium schreberi | 76 | Mycelis muralis | 43.2 | Corylus avellana B | 37.5 | Athyrium filix-femina | 47.5 | Trifolium medium | 66.7 | Eriophorum vaginatum | 71.4 |
| Vaccinium myrtillus | 58.5 | Oxalis acetosella | 41.9 | Dryopteris carthusiana | 35.6 | Ranunculus cassubicus | 36.4 | Calamagrostis arundinacea | 60.5 | Sphagnum magellanicum | 71.4 |
| Hylocomium splendens | 51.8 | Circaea alpina | 30.9 | Paris quadrifolia | 32.2 | Galeobdolon luteum | 31.5 | Agrimonia eupatoria | 58.3 | Ledum palustre | 57.1 |
| Dicranum polysetum | 50.2 | Sorbus aucuparia B | 30 | Viburnum opulus С | 30.8 | Knautia arvensis | 58.3 | Vaccinium uliginosum | 57.1 | ||
| Picea abies С | 31.8 | Leucanthemum vulgare | 55 | Sphagnum angustifolium | 57.1 | ||||||
| Melampyrum pratense | 31.7 | Veronica officinalis | 51.4 | Vaccinium vitis-idaea | 54.9 | ||||||
| Clinopodium vulgare | 45.8 | Carex globularis | 53.8 | ||||||||
| Carex pallescens | 44.1 | Oxycoccus palustris | 42.9 | ||||||||
| Vicia cracca | 41.7 | Polytrichum strictum | 42.9 | ||||||||
| Campanula persicifolia | 40.6 | Aulacomnium palustre | 40.7 | ||||||||
| Fragaria vesca | 40.2 | Betula pubescens B | 37.9 | ||||||||
| Pinus sylvestris С | 36.5 | ||||||||||
| Lathyrus vernus | 34.9 | ||||||||||
| Melica nutans | 33.7 | ||||||||||
| Antennaria dioica | 33.3 | ||||||||||
| Astragalus glycyphyllos | 33.3 | ||||||||||
| Viola hirta | 33.3 | ||||||||||
| Chamaenerion angustifolium | 32.4 | ||||||||||
В мелкотравной и мелкотравно-широкотравной группах сообществ77 в составе подчиненных ярусов наблюдается содоминирование типичных представителей таежного мелкотравья (Rubus saxatilis, Gymnocarpium dryopteris, Oxalis acetosella), зеленых мхов (Hylocomium splendens, Pleurozium schreberi) и видов широколиственных и широколиственно-хвойных лесов (Circaea alpina, Paris quadrifolia). Присутствие лещины в качестве индикационного вида в группе сосняков с участием ели, березы, липы, дуба мелкотравно-широкотравных достаточно показательно и отличает данную группу от мелкотравной, в древесном пологе которой не отмечены широколиственные породы, а только ель и береза помимо сосны. Такая картина в целом является, с одной стороны, характерной для группы сложных сосняков Московского региона (Рысин, 1964), с другой стороны, свидетельствует о “неморализации” средневозрастных насаждений, в том числе искусственного происхождения, при переходе на стадию более “зрелых сообществ” (Пестерова, 2013). В составе мелкотравно-широкотравной группы сообществ в целом отмечена наибольшая доля культур сосны (66%).
Широкотравные сосновые и сосново-еловые леса дифференцируют в основном виды неморального широкотравья, а в верхнем ярусе отмечены широколиственные виды деревьев (табл. 2). Доля насаждений искусственного происхождения, главным образом в составе сосново-еловых широкотравных сообществ, также высока (65%).
Индикаторные виды группы разнотравных сосняков с участием ели и березы в древесном пологе включают лугово-опушечные виды (Trifolium medium, Agrimonia eupatoria и др.), а также Calamagrostis arundinacea. Данные сообщества произрастают на террасах и крутых склонах рек на легких песчаных почвах с хорошим дренажом, в том числе антропогенно нарушенных местообитаний. В составе травяно-кустарничкового яруса в качестве индикаторного вида присутствует Pinus sylvestris (табл. 2).
Сосняки с елью и березой кустарничково-сфагновые отмечены в понижениях заболоченных низменностей; индикаторные виды – Vaccinium vitis-idaea, V. uliginosum, Ledum palustre, Eriophorum vaginatum и Carex globularis, в моховом покрове – Sphagnum angustifolium и S. magellanicum (табл. 2).
Расположение шести групп сообществ в ординационных осях (рис. 2) демонстрирует их распределение в пространстве экологических факторов: 1) хорошо дифференцируются группы сосняков кустарничково-мелкотравно-зеленомошных, широкотравных, разнотравных и кустарничково-сфагновых; 2) мелкотравно-широкотравная и мелкотравная группы частично перекрываются, что указывает на сходный флористический состав сообществ этих двух групп, представленный близкими по экологии видами. Положения центроидов шести групп по осям ординации статистически значимо различаются для всех пар групп сообществ хотя бы по одной оси (согласно результатам теста Манна–Уитни, при p < 0.05). Исключение составила пара кустарничково-мелкотравно-зеленомошных и кустарничково-сфагновых групп, – положение их центроидов различается только в трехмерном пространстве. Все векторы экологических факторов имеют статистически значимую связь с обеими осями ординации, за исключением увлажненности почв (табл. 3). Однако коэффициенты корреляции невысоки и превышают 0.5 только в случае трофности и богатства почв азотом по отношению к первой оси.
Рис. 2.
Распределение сообществ с участием сосны в осях NMDS ординации.
На рис. 2–4 обозначения векторов экологических факторов: HD – влажность почвы, NT – обеспеченность почв азотом, RC – кислотность почвы, TR – трофность почвы, LC –освещенность. Группы сообществ: 1 – кустарничково-мелкотравно-зеленомошная; 2 – мелкотравная, 3 – мелкотравно-широкотравная; 4 – широкотравная; 5 – разнотравная; 6 – кустарничково-сфагновая.
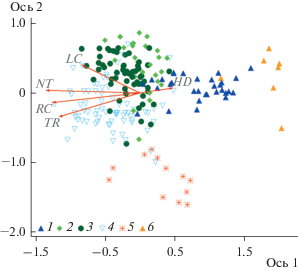
Таблица 3.
Корреляция Спирмена между значениями осей NMS ординации и значениями экологических факторов
| HD | TR | NT | RC | LC | |
|---|---|---|---|---|---|
| Ось 1 | 0.219000 | 0.532710 | 0.540015 | 0.443258 | 0.169022 |
| Ось 2 | 0.077736 | 0.433672 | 0.392263 | 0.287883 | 0.398358 |
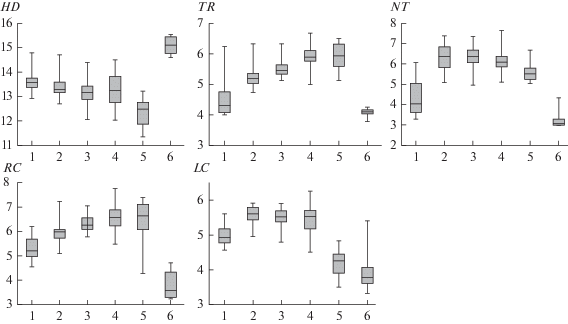
Влажность почв (HD) наибольшая в местообитаниях кустарничково-сфагновых лесов, наименьшая – в местообитаниях разнотравных лесов (рис. 3). Мелкотравная, мелкотравно-широкотравная и широкотравная группы статистически значимо не различаются между собой по этому экологическому режиму. Остальные пары демонстрируют значимые различия. Наибольшая трофность (TR) характерна для широкотравных и разнотравных лесов, причем они статистически значимо не различаются между собой по этому экологическому режиму, наименьшая – для местообитаний кустарничково-сфагновых лесов. Все пары, за исключением первой, статистически значимо различаются по трофности почв. Наиболее богатые азотом почвы (NT) отмечены в местообитаниях мелкотравной, мелкотравно-широкотравной и широкотравной групп лесных сообществ. Наиболее бедные азотом почвы характерны для кустарничково-сфагновых лесных сообществ. По данному экологическому режиму статистически значимо различаются все пары, за исключением мелкотравной и мелкотравно-широкотравной, мелкотравной и широкотравной групп. Наибольшие значения кислотности почв (RC) (наименьшие балльные оценки) отмечены для кустарничково-сфагновых лесных сообществ, наименьшие (самые высоки баллы по шкале) – для местообитаний мелкотравно-широкотравных, широкотравных и разнотравных групп лесов. По кислотности почв все пары значимо различаются между собой, за исключением мелкотравной-широкотравной и разнотравной, широкотравной и разнотравной. Самыми освещенными (LC) (наименьшие балльные оценки) являются подчиненные ярусы разнотравных и кустарничково-сфагновых лесов, причем статистически значимо они не различаются по этому режиму. Наиболее затенены подчиненные ярусы мелкотравной, мелкотравно-широкотравной и широкотравной групп лесов, и эти пары также статистически значимо не различаются по освещенности.
Рис. 3.
Варьирование экологических характеристик местообитаний лесных сообществ.
На боксплотах отражены медианы значений, интерквартильные расстояния (25–75% варьирования) и минимум и максимум значений. Обозначения см. на рис. 2.
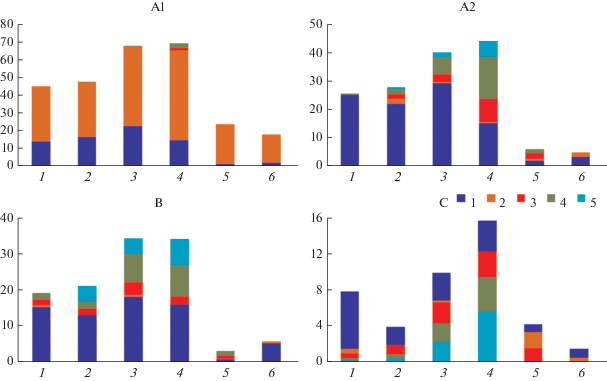
Для оценки устойчивости сообществ и перспективы сохранения сосновых насаждений прослежено наличие сосны и соотношение основных древесных видов (ели, сосны, дуба, липы и клена) в различных ярусах выделенных групп сообществ. Как видно из рис. 4, присутствие сосны в подчиненных ярусах (А2 и С) отмечается не во всех группах сообществ: в ярусе А2 – в мелкотравной и кустарничково-сфагновой группах, в ярусе С – в кустарничково-мелкотравно-зеленомошной, разнотравной и кустарничково-сфагновой группах; в ярусе В подрост сосны отсутствует во всех группах. В ярусе А2 в трех группах сообществ преобладает ель, в широкотравной группе – широколиственные породы, в ярусе В ситуация аналогичная, а в ярусе С ель “активна” лишь в кустарничково-мелкотравно-зеленомошных сообществах, тогда как в остальных группах преобладают широколиственные древесные виды либо подрост сосны.
Рис. 4.
Изменение активности древесных видов в разных ярусах сосняков. А1 – первый подъярус древостоя; А2 – второй подъярус древостоя, B – кустарниковый ярус, С – травяно-кустарничковый ярус. Обозначения групп см. на рис. 2.
Для оценки пространственного распределения сообществ с участием сосны выполнена совместная обработка наземных полевых данных и пространственной информации – ДДЗ и ЦМР. Относительное качество дискриминации выделенных синтаксонов от данных ДДЗ и ЦМР равно 64.6% (табл. 1). Распространение сообществ по-разному зависит от переменных ДДЗ и ЦМР. С меньшей точностью от ДДЗ (30%) классифицируется сосново-еловая мелкотравно-широкотравная группа сообществ, характеризующаяся смешанным составом как древесного, так и подчиненных ярусов, т.е. для данной группы отсутствует четкий спектральный отклик, что может указывать на производный характер данных сообществ, их искусственное происхождение в нехарактерных для естественных сосновых насаждений ландшафтных условиях (отсутствие связи с морфометрическими данными рельефа). Лучше дискриминируются группы, ассоциированные с характерными для коренных сосняков местообитаниями (кустарничково-сфагновая и разнотравная), а также более однородные по составу древостоя группы.
Дискриминантный анализ позволяет выполнить оценку отдельных характеристик растительного покрова, полученных из наземных описаний сообществ. Точность дискриминационной модели распределения сосновых сообществ по показателю сомкнутости крон деревьев верхнего подъяруса (А1) составила 69.6%. При этом наблюдается достаточно высокая величина качества дискриминации только по показателям рельефа – 56.1%, что доказывает определенную зависимость распространении сосняков от ландшафтных условий.
Анализ пространственного распределения выделенных групп сообществ с участием сосны для исследуемой территории (рис. 5) выявил, что наибольшая встречаемость сосновых массивов отмечается в северной части региона по северным отрогам Клинско-Дмитровской гряды и на юге по крутым берегам р. Оки. Характерно, что на относительно небольшом географическом пространстве в пределах исследуемой территории Московского региона в субширотном направлении можно отследить зональные особенности распределения сосновых насаждений. Так, леса кустарничково-мелкотравно-зеленомошной группы преобладают в северо-западном секторе (рис. 5а), а леса широкотравной группы сообществ увеличивают площадь покрытия в юго-восточном секторе исследуемой территории (рис. 5г). Площади распределения сосновых сообществ разного типа по ФГП (табл. 4) подтверждают визуальную оценку их распределения по карто-схемам. Площадь сосняков кустарничково-мелкотравно-зеленомошной группы в Верхне-Волжской ФГП на порядок превышает, а широкотравной, соответственно, на порядок меньше представленности их в остальных ФГП исследуемой территории. Интразональные группы сообществ (сосняки разнотравные и кустарничково-сфагновые) распределены в исследованном регионе более-менее равномерно (рис. 5д, 5е), что подтверждается равномерным соотношением их площадей в разных ФГП (табл. 4). Сосняки разнотравные имеет отчетливую ленточную конфигурацию полигонов, свидетельствующую о приуроченности к морфоскульптурным формам рельефа – речным террасам.
Рис. 5.
Распределение групп сообществ с участием сосны для исследуемой территории Московской области
Группы сообществ: а – кустарничково-мелкотравно-зеленомошная; б – мелкотравная, в – мелкотравно-широкотравная; г – широкотравная; д – разнотравная; е – кустарничково-сфагновая.
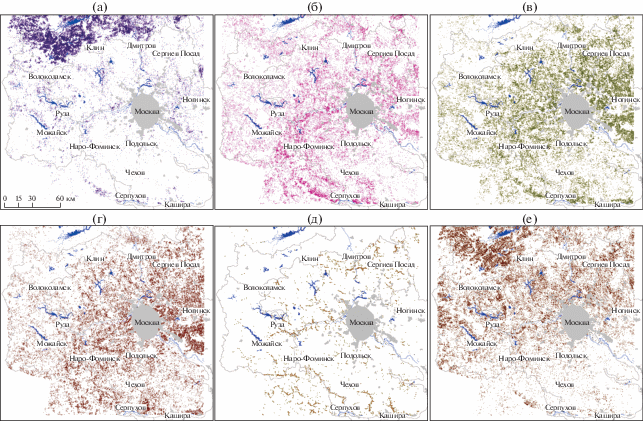
Таблица 4.
Ландшафтно-экологические метрики для различных групп сообществ сосняков в пределах физико-географических провинций Московской области
| ФГП | Группа сообществ | Площадь, тыс. га | Доля в провинции, % | Средняя площадь выдела, га | Индекс формы | Индекс ядра, % | Дистанция, м | Индекс контраста |
|---|---|---|---|---|---|---|---|---|
| 1 | 1 | 60.5 | 13.20 | 5.64 | 1.20 | 90.08 | 184.27 | 13.03 |
| 2 | 1.9 | 0.41 | 0.54 | 1.03 | 95.35 | 390.39 | 4.99 | |
| 3 | 1.8 | 0.40 | 0.68 | 1.06 | 94.73 | 364.60 | 4.30 | |
| 4 | 2.7 | 0.59 | 0.64 | 1.05 | 97.94 | 322.56 | 5.91 | |
| 5 | 0.1 | 0.02 | 2.32 | 1.11 | 93.50 | 2187.43 | 12.90 | |
| 6 | 8.5 | 1.85 | 1.17 | 1.09 | 96.14 | 253.28 | 13.33 | |
| 2 | 1 | 3.2 | 0.51 | 0.79 | 1.07 | 90.84 | 344.92 | 7.01 |
| 2 | 4.5 | 0.70 | 0.73 | 1.04 | 91.90 | 369.86 | 7.09 | |
| 3 | 6.6 | 1.04 | 0.86 | 1.07 | 89.89 | 289.54 | 5.67 | |
| 4 | 6.1 | 0.95 | 0.78 | 1.07 | 96.15 | 298.23 | 5.64 | |
| 5 | 1.3 | 0.20 | 3.03 | 1.16 | 86.14 | 858.51 | 18.42 | |
| 6 | 6.0 | 0.93 | 0.76 | 1.06 | 94.88 | 287.80 | 12.61 | |
| 3 | 1 | 5.0 | 0.53 | 0.69 | 1.05 | 70.50 | 349.53 | 20.59 |
| 2 | 7.8 | 0.83 | 0.67 | 1.05 | 86.61 | 321.41 | 8.71 | |
| 3 | 30.1 | 3.19 | 1.34 | 1.12 | 90.14 | 208.11 | 5.59 | |
| 4 | 16.7 | 1.77 | 0.98 | 1.09 | 94.61 | 244.38 | 5.45 | |
| 5 | 3.4 | 0.36 | 3.16 | 1.15 | 86.36 | 621.20 | 17.56 | |
| 6 | 14.2 | 1.51 | 0.83 | 1.07 | 86.88 | 258.44 | 24.66 | |
| 4 | 1 | 2.3 | 0.25 | 0.63 | 1.04 | 55.59 | 462.63 | 31.42 |
| 2 | 8.6 | 0.91 | 0.77 | 1.06 | 81.96 | 309.04 | 12.77 | |
| 3 | 16.7 | 1.77 | 1.20 | 1.10 | 91.46 | 251.95 | 6.71 | |
| 4 | 26.3 | 2.78 | 1.48 | 1.11 | 93.67 | 232.85 | 7.29 | |
| 5 | 2.6 | 0.28 | 2.68 | 1.15 | 86.14 | 607.42 | 19.68 | |
| 6 | 3.2 | 0.34 | 0.61 | 1.04 | 77.63 | 427.72 | 27.47 | |
| 5 | 1 | 3.3 | 0.94 | 0.59 | 1.05 | 61.11 | 272.91 | 25.18 |
| 2 | 3.6 | 1.04 | 0.63 | 1.05 | 82.38 | 285.69 | 11.62 | |
| 3 | 14.6 | 4.16 | 1.38 | 1.14 | 89.60 | 184.58 | 5.88 | |
| 4 | 22.6 | 6.47 | 1.82 | 1.16 | 91.24 | 176.67 | 6.88 | |
| 5 | 0.1 | 0.04 | 1.27 | 1.12 | 84.78 | 2056.33 | 19.57 | |
| 6 | 2.2 | 0.62 | 0.58 | 1.04 | 77.43 | 335.27 | 26.73 |
Примечание. Названия ФГП: 1 – Верхне-Волжская, 2 – Московская, 3 – Смоленская, 4 – Москворецко-Окская, 5 – Мещерская. Группы сообществ: 1 – кустарничково-мелкотравно-зеленомошная; 2 – мелкотравная; 3 – мелкотравно-широкотравная; 4 – широкотравная; 5 – разнотравная; 6 – кустарничково-сфагновая. Жирным шрифтом выделены высокие значения, подчеркиванием – низкие.
Для более детального изучения пространственных особенностей распределения сообществ выполнен дисперсионный анализ связи сосновых насаждений с разными ландшафтными условиями на разных иерархических уровнях организации рельефа. Получено, что группы ассоциации в наибольшей степени достоверно связаны с высотами для различных иерархических уровней рельефа, уклонами, различными кривизнами и освещенностью для уровней в диапазоне 300–1140 м. Наиболее выпуклые формы рельефа характерны для сосняков с липой, дубом и лещиной широкотравных, наиболее вогнутые – для сосняков кустарничково-сфагновых. Распространение сосняков кустарничково-мелкотравно-зеленомошных связано с местообитаниями, характеризующимися очень небольшими показателями высоты рельефа и его форм для иерархического уровня организации рельефа с линейным размером 19980 м, но одновременно распространение широкотравных и кустарничково-мелкотравно-зеленомошных – с большими высотами для уровня 300 м и со средними высотами для уровней 540 и 2220 м. Уклоны более 6° отмечаются только для сосняков разнотравных. Освещенность при азимуте 90° и 180°, определяющая экспозиции, в наибольшей степени выражена для сосняков разнотравных, имеющих наибольшие уклоны рельефа и преобладание юго-западной ориентации склонов. Наименьшая освещенность с востока и юга отмечается для сосново-еловых мелкотравных и широкотравных лесов, занимающих склоны северо-западной ориентации.
Оценка неоднородности пространственной структуры контуров групп сообществ основана на расчете и анализе пространственных ландшафтно-экологических метрик – площади, формы, площади экологического ядра, агрегированности и контрастности (табл. 4). Для групп сообществ сосновых лесов метрики рассчитаны в разрезе ФГП. В пределах исследуемой территории в Верхне-Волжской и Мещерской провинциях площадь сосновых лесов наибольшая (16.45 и 13.27%, соответственно), Смоленской – наименьшая (4.33%), в Московской и Москворецко-Окской – 8.19 и 6.32%. Проанализированы две площадные характеристики – доля группы сообществ в общей площади провинции и средняя площадь выдела. Во всех 5 провинциях сосновые леса характеризуются незначительной средней площадью выдела (1.31 га). На фоне других групп выделяются сосняки разнотравные, которые имеют средние площади выше среднего 1.27–3.16 га, однако при этом доля этой группы в общей площади провинции минимальна (0.02–0.36%) (табл. 4). Это говорит об очаговом (островном) характере их распространения, связанном с приуроченностью к речным террасам.
В Верхне-Волжской провинции доминирует кустарничково-мелкотравно-зеленомошная группа – 13.2%, при этом для нее характерен наибольший средний размер выдела – 5.64 га. Доминирующие группы в Смоленской провинции не выделяются. В Московской, Москворецко-Окской и Мещерской провинциях с небольшими отличиями доминируют широкотравные и широкотравно-мелкотравные группы (табл. 4).
Группы, доминирующие в своей провинции, обычно имеют большее значение метрики формы (и соответственно более сложную форму, что косвенно индицирует меньшую степень воздействия антропогенных процессов, нередко “спрямляющих” границы). Исключение – разнотравные сосняки, при крупных размерах выделов сохраняющие естественную сложную форму контуров.
Индекс площади ядра для всех групп сосновых лесов Верхне-Волжской и Смоленской провинций высокий (93.13% в среднем), что говорит о соседстве сосновых лесов с лесами других типов, и, следовательно, низкой степени антропогенной нарушенности. В Московской, Москворецко-Окской и Мещерской провинциях высокий индекс площади ядра имеют группы, преобладающие по площади (мелкотравные, широкотравно-мелкотравные и широкотравные – по 83.65, 90.4 и 93.18%, соответственно) и сосняки разнотравные (85.76%) (табл. 4). Для двух других групп наблюдается существенное снижение площади экологического ядра вследствие соседства с гетерогенными типами растительности и наземного покрова. Вкупе с небольшими средними площадями выделов это говорит о “рисковом” состоянии этих групп.
Агрегированность (дистанция) варьирует для сосновых лесов в целом от 176 до 2187 м (458 м в среднем). Для упомянутых выше в контексте изолированности сосняков разнотравных она составляет в среднем 1266 м для всех провинций. Группы-доминанты Московской, Москворецко-Окской и Мещерской провинций характеризуются меньшими дистанциями между выделами (264 м) нежели субдоминирующие кустарничково-мелкотравно-зеленомошная и кустарничково-сфагновая (317 м) в целом по группам (табл. 4).
Метрика контрастности границ в целом низкая для выделов групп-доминантов (в среднем 6.9), что коррелирует с выводом о соседстве сосновых лесов с другими типами леса. Для сосняков разнотравных она составляет 17.63, для групп-субдоминантов – 11.46 в пределах Верхне-Волжской и Смоленской провинций и 26.0 в пределах Московской, Москворецко-Окской и Мещерской (табл. 4).
Заключение. Считается, что сосново-еловые леса с участием широколиственных пород являются коренным типом растительности в определенных ландшафтных условиях геоморфологического района Клинско-Дмитровской гряды (Огуреева и др., 1996). Полученные данные и разработанная классификация уточняют существующие представления о типологическом разнообразии сообществ с участием сосны в Московской области. Отсутствие возобновления сосны в сообществах автоморфных местообитаний свидетельствует о производном происхождении сосняков после пожаров и на месте вырубок, а также в составе искусственных насаждений. Наш вывод о производном характере всех сосняков Московского региона, за исключение сосняков верховых болот согласуется с мнением других исследователей (Маслов, 2000, 2002; Рысин, Савельева, 2002).
Всего в ранге групп ассоциаций выделено 11 синтаксонов, объединенных по преобладающей экобиоморфе растительности подчиненных ярусов в 6 групп сообществ: кустарничково-мелкотравно-зеленомошные, мелкотравные, мелкотравно-широкотравные, широкотравные, разнотравные, кустарничково-сфагновые. Анализ состава выделенных групп сообществ, с одной стороны, отражает экологическую специфичность их набора видов, с другой стороны, – переходный сукцессионный статус в направлении к более “зрелой” стадии, в частности, для сообществ искусственного происхождения. В так называемых “сложных” сосняках с участием липы и дуба мелкотравно-широкотравных доля культур (по материалам геоботанических описаний) – около 80%. На водораздельных поверхностях восстановительные сукцессии идут по двум направлениям: в одном случае восстановление сопровождается активной демутацией ели; в другом при повышенном богатстве почв – широколиственными породами, исключающими существование сосновых и сосново-еловых сообществ через несколько десятков лет. Небольшая доля сосновых насаждений сохранится по крутым склонам рек в силу благоприятных для сосны экологических условий (легкие песчаные почвы с хорошим дренажем) при постоянно поддерживаемом рекреационном воздействии, а также в гидроморфных условиях.
Организация сосновых лесов Московской области существенно различаются по 5 физико-географическим провинциям в связи с зональными различиями, а также вследствие различной степени антропогенной трансформации. Ландшафтно-экологические метрики являются надежными индикаторами как зональных различий групп сообществ, так и степени фрагментации, которая более ярко проявляется в центральной и юго-восточной частях территории исследования. Сообщества, отнесенные к кустарничково-мелкотравно-зеленомошной и кустарничково-сфагновой группам в пределах Московской, Москворецко-Окской и Мещерской провинций в наибольшей степени подвержены фрагментации. Площади выделов минимальны, дистанции между выделами сравнительно с другими группами велики. Выделы, как правило, окружены гетерогенными по отношению к сосновому лесу типами ландшафтов. Глубина проникновения краевых эффектов сокращает площадь экологического ядра до 70%. Наиболее уязвимы во всех провинциях сосняки разнотравные. Несмотря на то, что отдельные массивы сохраняют площадь выше среднего – 2.49 га, изолированность выделов достигает почти 2200 м. В таких условиях выделы лишаются возможности заноса семязачатков и приближаются к состоянию “островов”.
Полученные данные дополняют существующие представления о фитоценотической структуре и пространственном распределении сообществ с участием сосны, состав которых отражает сукцессионную стадию (1), зональные черты растительности исследуемого региона (2) и приуроченность к определенным ландшафтным элементам (3). В субширотном направлении отмечено разное соотношение сообществ с преобладанием видов растений бореального, субнеморального и неморального спектра в наземном ярусе.
Благодарим О.А. Пестерову, М.В. Архипову, Н.Г. Кадетова, С.Ю. Попова, Е.В. Тихонову, принимавших участие в сборе материала, а также Е.А. Игнатову за определение видов мохообразных.
Список литературы
Анненская Г.Н., Жучкова В.К., Калинина В.Р., Мамай И.И., Низовцев В.А., Хрусталева М.А., Цесельчук Ю.Н. Ландшафты Московской области и их современное состояние. Смоленск: Изд-во Смоленского гуманитарного университета. 1997. 296 с.
Бадер О.Н. Новая палеоантропологическая находка под Москвой // Антропологический журнал. № 4. 1936. С. 471–475.
Быков Б.А. Экологический словарь. Алма-Ата: Наука, 1983. 21 с.
Викторов А.С. Рисунок ландшафта. М.: Мысль, 1986. 179 с.
Грибова С.А., Исаченко Т.И., Лавренко Е.М. Растительность Европейской части СССР. Л.: Наука. Ленинградское отделение, 1980. 429 с.
Заугольнова Л.Б., Морозова О.В. Типология и классификация лесов Европейской России: методические подходы и возможности их реализации // Лесоведение. 2006. № 1. С. 1−15.
Исаев А.С., Черненькова Т.В. Уникальность современного этапа дистанционного зондирования лесов России // Сибирский лесной журн. 2015. № 5. С. 26–41.
Король Г.Г. Археологическая карта России. Московская область. Балашихинский, Домодедовский, Красногорский, Ленинский, Люберецкий, Мытищинский, Одинцовский, Подольский, Пушкинский, Химкинский, Щелковский районы. 1994. Т. 1. 319 с.
Король Г.Г. Археологическая карта России. Московская область. Ногинский, Орехово-Зуевский, Павлово-Посадский, Раменский, Сергиево-Посадский, Шатурский районы. 1996. Т. 3.267 с.
Король Г.Г. Археологическая карта России. Московская область. Волоколамский, Дмитровский, Истринский, Клинский, Лотошинский, Можайский, Наро-Фоминский, Рузский, Солнечногорский, Талдомский, Шаховской районы. 1995. Т. 2. 237 с.
Король Г.Г. Археологическая карта России. Московская область. Воскресенский, Егорьевский, Зарайский, Каширский, Коломенский, Луховицкий, Озерский, Серебряно-Прудский, Серпуховский, Ступинский, Чеховский районы. 1997. Т. 4. 350 с.
Котлов И.П., Пузаченко Ю.Г. Структура рельефа Русской равнины как ландшафтообразующего фактора // Ландшафтное планирование, труды конференции, Москва, 22–25 августа 2006 г., М.: Изд-во Географического факультета МГУ, 2006. С. 166–171.
Кучеров И.Б. Эколого-ценотическое разнообразие светлохвойных лесов средней и северной тайги Европейской России // Автореф. дисс. … д-ра биол. наук (03.02.08). С-Пб.: Ботанический институт им. В.Л. Комарова, 2018. 46 с.
Лесной план Московской области. Красногорск, 2018. Кн. 1. 83 с. http://klh.mosreg.ru/download/document/ 374116, 03.09.2018.
Малышев Л.И. Флористическое районирование на основе количественных признаков // Ботанический журнал. 1973. Т. 58. № 11. С. 1581–1588.
Маслов А.А. Динамика сосняков и ельников на территории заповедных лесных участков // Динамика хвойных лесов Подмосковья. М.: Наука, 2000. С. 67–85.
Маслов А.А. Сукцессионная динамика древостоя и нижних ярусов в послепожарном 100-летнем сосняке лишайниково-зеленомошном // Лесоведение. 2002. № 2. С. 23–29.
Морозов Г.Ф. Учение о лесе. М.; Пг.: Государственное издательство, 1924. 404 с.
Огуреева Г.Н., Микляева И.М., Суслова Е.Г., Швергунова Л.В. Растительность Московской области. Пояснительный текст и легенда к карте М 1 : 200 000. М.: МГУ. 1996. 45 с.
Пестерова О.А. Эколого-фитоценотические особенности и роль лесных культур в формировании биоразнообразия лесов юго-западного Подмосковья // Автореф. дисс. … канд. биол. наук (03.02.08). С-Пб.: Ботанический институт им. В.Л. Комарова, 2013. 25 с.
Петров В.В. Новая схема геоботанического районирования Московской области // Вестник Московского государственного университета. Серия 6: Биология, почвоведение. 1968. № 5. С. 44–49.
Порфирьев В.С. О применении понятий серии и цикла при изучении хвойно-широколиственных лесов // Бюллетень МОИП. Отделение биологическое. 1960. Т. 65. Вып. 3. С. 93−102.
Пузаченко Ю.Г. Математические методы в экологических и географических исследованиях. М.: Академия, 2004. 408 с.
Пузаченко Ю.Г., Сандлерский Р.Б., Кренке А.Н., Пузаченко М.Ю. Мультиспектральная дистанционная информация в исследовании лесов // Лесоведение. 2014. № 5. С. 13–29.
Пузаченко М.Ю., Черненькова Т.В. Определение факторов пространственного варьирования растительного покрова с использованием ДДЗ, ЦМР и полевых данных на примере центральной части Мурманской области // Современные проблемы дистанционного зондирования Земли из космоса. 2016. Т. 13. № 5. С. 167–191.
Рысин Л.П. Возобновление сосны в сложных борах с подлеском из лещины // Лесное хозяйство. 1964. № 10. С. 12–15.
Рысин Л.П., Савельева Л.И. Сосновые леса России. М.: Товарищество научных изданий КМК, 2008. 289 с.
Рысин Л.П., Савельева Л.И. Еловые леса России. М.: Наука, 2002. 335 с.
Рязапов Р.И., Кабанов С.В. Возрастная структура ценопопуляций Pinus sylvestris L. малонарушенных сосняков южной части Приволжской возвышенности // Вестник Саратовского госагроуниверситета им. Н.И. Вавилова. 2010. № 8. С. 27–35.
Сабуров Д.Н. Леса Пинеги. Л.: Наука, 1972. 173 с.
Савельева Л.И. Типы хвойных лесов Подмосковья // Динамика хвойных лесов Подмосковья. М.: Наука, 2000. С. 33–66.
Самбук Ф.В. Печорские леса // Труды Ботанического Музея АН СССР. Л.: 1932. Вып. 24. С. 63–250.
Санников С.Н. Экология и география естественного возобновления сосны обыкновенной. М.: Наука, 1992. 264 с.
Фридланд В.М. Структура почвенного покрова. М.: Мысль, 1972. 423 с.
Харитонов М.В. Находки ископаемых гоминид на территории Восточной Европы и сопредельных регионов Азии. [электронный ресурс] 14.04.2019,http://antropogenez.ru/article/157/, 14.04.2019.
Холод С.С. Структура растительного покрова острова Врангеля. Автореф. дисс. … д-ра биол. наук (03.02.08). С-Пб.: Ботанический институт им. В.Л. Комарова, 2017. 43 с.
Цыганов Д.Н. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов. М.: Наука, 1983. 196 с.
Черепанов С.К. Сосудистые растения России и сопредельных государств. СПб.: Мир и семья, 1995. 992 с.
Черненькова Т.В., Морозова О.В., Пузаченко М.Ю., Попов С.Ю., Беляева Н.Г. Состав и структура еловых лесов юго-западного Подмосковья // Лесоведение. 2015. № 5. С. 323–338.
Экобиоморфа // Большая советская энциклопедия: В 30 томах. / Гл. ред. А.М. Прохоров. 3-е изд. М.: Советская энциклопедия, 1978. Т. 29. 639 p.
Bohn U., Gollub G., Hettwer C., Neuhäuslová Z., Raus T., Weber H. Karte der natürlichen Vegetation Europas. Map of the natural vegetation of Europe (Maßstab/Scale 1 : 2 500 000), Bonn: Bundesamt für Naturschutz, Federal Agency for Nature Conservation, 2004. 512 p.
Chernen’kova T.V., Morozova O.V. Classification and mapping of coenotic diversity of forests, Contemporary problems of ecology, 2017. V. 10. № 7. P. 738–747.
Dufrêne M., Legendre P. Species assemblages and indicator species: the need for a flexible asymmetrical approach, Ecological Monographs, 1997. V. 67. № 3. P. 345–366.
Ignatov M.S., Afonina O.M., Ignatova E.A., Abolina A., Akatova T.V., Baisheva E.Z., Bardunov L.V., Baryakina E.A., Belkina O.A., Bezgodov A.G., Boychuk M.A., Cherdantseva V.Y., Czernyadjeva I.V., Doroshina G.Y., Dyachenko A.P., Fedosov V.E., Goldberg I.L., Ivanova E.I., Jukoniene I., Kannukene L., Kazanovsky S.G., Kharzinov Z.K., Kurbatova L.E., Maksimov A.I., Mamatkulov U.K., Manakyan V.A., Maslovsky O.M., Napreenko M.G., Otnyukova T.N., Partyka L.Y., Pisarenko O.Y., Popova N.N., Rykovsky G.F., Tubanova D.Y., Zheleznova G.V., Zolotov V.I. Check-list of mosses of East Europe and North Asia. Arctoa, 2006. V. 15. P. 1–130.
Jackson R.D., Huete A.R. Interpreting vegetation indices // Preventive Veterinary Medicine. 1991. V. 11. № 3–4. P. 185–200.
Legendre P., Legendre L. Numerical Ecology. Amsterdam: Elsevier science B.V. 1998. 853 p.
McCune B., Grace J.B. Analysis of ecological communities, Gleneden Beach: MjM Software Design, 2002, 304 p.
McCune B., Mefford M.J. PC-ORD. Multivariate analysis of Ecological Data. Version 5. Gleneden Beach: MjM Software, 2011.
McGarigal K., Cushman S.A., Ene E. FRAGSTATS v4: Spatial Pattern Analysis Program for Categorical and Continuous Maps. Computer software program produced by the authors at the University of Massachusetts. 2012. Amherst. http://www.umass.edu/landeco/research/fragstats/fragstats. html, 14. 04. 2019.
Дополнительные материалы отсутствуют.
Инструменты
Лесоведение