

Лесоведение, 2019, № 5, стр. 423-434
Первичная продукция сосняков Республики Коми
А. Ф. Осипов a, *, К. С. Бобкова a
a Институт биологии Коми научного центра УрО РАН
167982 г. Сыктывкар, ул. Коммунистическая, 28, Россия
* E-mail: osipov@ib.komisc.ru
Поступила в редакцию 12.02.2019
После доработки 27.03.2019
Принята к публикации 05.06.2019
Аннотация
Накопление углерода органического вещества лесными фитоценозами в процессе фотосинтеза является их важнейшей функцией, обеспечивающей смягчение изменения климата на планете. В литературе приводятся данные, что территория Российской Федерации является крупным стоком углерода атмосферы. Однако его оценки, как и величины нетто-продукции (NPP) лесных экосистем, изменяются в широких пределах. В работе приводятся материалы, характеризующие нетто-продукцию углерода фитомассы в сосняках разных типов условий произрастания в северной и средней тайге Республики Коми. Исследования выполнены на лесных стационарах в приспевающих, спелых и перестойных сосняках зеленомошной, лишайниковой и сфагновой групп типов. Связывание углерода древостоем оценено по биологической продуктивности, с использованием модельных деревьев, при анализе которых выведены регрессионные уравнения зависимости прироста отдельных органов от диаметра ствола на высоте 1.3 м. Обнаружена статистически значимая взаимосвязь между этими параметрами, характеризующаяся высокими показателями аппроксимации тренда, варьирующими от 0.44 до 0.99 (при 95%-м уровне значимости). Рассчитано, что в условиях европейского Северо-Востока России NPP углерода сосновых фитоценозов изменяется в пределах 1.9–4.5 т С га–1 год–1. Выявлено, что на долю древостоя приходится 32–73% от общей нетто-продукции. С увеличением увлажнения почвы и при продвижении на север в общей продукции углерода возрастает участие растений напочвенного покрова. Отмечено, что значительную часть NPP углерода древесного яруса сосняков формируют фотосинтетический аппарат и древесина ствола. На основании полученных данных выведены конверсионные отношения между NPP и запасом древесины/углерода в древостое для сосняков северной и средней тайги.
Леса на территории Российской Федерации занимают более 20% от общей площади лесных массивов планеты (Food…, 2015), поэтому они имеют огромное экологическое, экономическое и социальное значение планетарного масштаба, играя важную роль в регулировании биосферных циклов (Исаев, Коровин, 2013). Принятие Рамочной конвенции ООН об изменении климата в 1992 г. послужило толчком к развитию исследований роли лесов Российской Федерации в глобальном углеродном цикле, которые возглавил Александр Сергеевич Исаев. Под его руководством были разработаны методы оценки и опубликованы первые сведения по запасам и нетто-продукции углерода (NPP) в лесных экосистемах России (Исаев и др., 1993; Isaev et al., 1995).
Важнейшей функцией лесных экосистем является продукция растительного органического вещества, которая в условиях изменяющегося климата рассматривается как экосистемная услуга по смягчению его последствий (Лукина и др., 2015). Кроме того, определение депонирования углерода фитомассы в насаждениях актуально для решения задач выращивания лесных сообществ оптимального состава, структуры и характеристики круговорота углерода в биосфере (Исаев и др., 2007). В работе S. Schaphoff с соавт. (2016) приведен анализ литературы, в котором содержится несколько оценок NPP на территории России. Согласно этим данным показатели NPP, полученные разными методами, варьируют от 397 до 2610 млн т в год, т.е. разнятся в 6.6 раза. По мнению ряда авторов (Luo et al., 2015; Metsaranta et al., 2018) накопление экспериментальных данных по продуктивности лесов разных условий произрастания имеют решающее значение для разработки экосистемных моделей, используемых при прогнозировании роли лесов в глобальном углеродном цикле и снижении неопределенностей при его оценке. Применительно к NPP это могут быть сведения по соотношению между надземной фитомассой (или объемом древесины ствола) и NPP, между которыми ранее была отмечена положительная взаимосвязь (Замолодчиков, Уткин, 2000; Keeling, Philips, 2007).
Европейский Северо-Восток России – это один из регионов, где сосредоточены крупнейшие лесные массивы, выполняющие важные средообразующие функции. Сосновые леса в этом регионе имеют давнюю историю лесопользования, в результате которой в возрастной структуре суммарная доля молодняков, средневозрастных и приспевающих насаждений достигает 55%. Однако спелые и перестойные древостои, роль которых в цикле углерода до сих пор остается дискуссионной, занимают значительные площади. По мнению многих авторов, в бореальном поясе хвойные леса на поздних стадиях развития продолжают активно связывать диоксид углерода атмосферы в процессе фотосинтеза (Luyssaert et al., 2008; Hardiman et al., 2013; Vedrova et al., 2018; и др.). Сведения по NPP сосняков европейского Северо-Востока России приведены в основном для среднетаежных насаждений (Бобкова, Осипов, 2012; Кутявин и др., 2016; Кутявин, 2018), а для сосновых насаждений северной тайги эти сведения единичны (Бобкова, 2005). Следует отметить, что к настоящему времени недостаточно данных, характеризующих изменчивость нетто-продукции сосняков, особенно растений нижних ярусов, в зависимости от лесорастительных условий не только в этом регионе, но и в России в целом. Цель работы − определить нетто-продукцию углерода фитомассы в сосняках разных типов условий произрастания в северной и средней тайге Республики Коми.
ОБЪЕКТЫ и МЕТОДИКА
Исследования выполнены в сосновых фитоценозах на Зеленоборском (северная тайга, 64°28′ с.ш., 55°22′ в.д.) и Чернамском (средняя тайга, 62°01′ с.ш., 50°28′ в.д.) лесных стационарах Института биологии Коми НЦ УрО РАН. Климат района исследований умеренно-континентальный. С продвижением на север среднегодовая температура воздуха снижается от +0.5 до –1.9°С, тогда как количество осадков в рассматриваемых подзонах сопоставимо (629 и 646 мм в год); примерно половина из них выпадает в течение вегетационного периода.
Работа проводилась на семи постоянных пробных площадях, заложенных в сосновых насаждениях разных типов, пройденных пожарами. Краткая характеристика древостоев исследованных фитоценозов приведена в табл. 1. Древостои среднетаежных сосняков практически чистые по составу, с небольшой примесью березы и ели, высокополнотные. Насаждения сосняков бруснично-лишайникового и чернично-сфагнового находятся в стадии приспевания, о чем свидетельствует значительная плотность деревьев в древостое, тогда как сосняк черничный вступает в стадию спелости (Осипов, Бобкова, 2016). В условиях северной тайги в составе древостоев сосновых фитоценозов на автоморфных почвах встречается лиственница (до 2 единиц). Береза и ель отмечены во всех исследованных насаждениях. Северотаежные сосняки характеризуются средней полнотой, которая уменьшается с увеличением увлажнения почв. Исследованные сосновые сообщества произрастают на подзолистых песчаных почвах, подстилаемых суглинками.
Таблица 1.
Краткая таксационная характеристика древостоев сосняков
| Тип леса* | Состав** | Густота, экз. га–1 | Полнота | Запас древесины, м3 га–1 | Средние | Запасы углерода, т га–1 | Почва | ||
|---|---|---|---|---|---|---|---|---|---|
| диаметр, см | высота, м | древостой | живой напочвенный покров | ||||||
| Северная тайга | |||||||||
| Чернично-зеленомошный (90) | 7С2Лц1Б+Е | 1400 | 0.6 | 153 | 13.5 | 13.4 | 54.08*** | 8.39*** | Глее-подзолистая, профильно-глеевая |
| Голубично-зеленомошный (90) | 9С1Лц+Б,Е | 1080 | 0.6 | 125 | 14.5 | 13.1 | 47.61*** | 6.28*** | Контактно-глеевый подзол |
| Черничный (100) | 6С2Лц1Е1Б | 1540 | 0.7 | 178 | 19 | 17 | 62.01*** | 5.61*** | Глее-подзолистая, профильно-глеевая |
| Осоково-сфагновый (180) | 10С+Б | 530 | 0.3 | 61 | 13 | 8.6 | 23.45*** | 5.87*** | Торфяно-глеевая иллювиально-гумусово-железистая |
| Средняя тайга | |||||||||
| Бруснично-лишайниковый (90) | 10С ед. Б | 2533 | 1.0 | 246 | 12 | 14 | 82.95***** | 1.02***** | Подзол иллювиально-железистый |
| Черничный (95) | 9C1Б+Е | 1195 | 1.0 | 220 | 19.1 | 15 | 66.41**** | 2.84**** | Торфянисто-подзолисто-глееватая иллювиально-железистая |
| Чернично-сфагновый (90) | 10С ед.Е | 2266 | 0.9 | 139 | 12 | 11 | 53.91**** | 2.74**** | Торфянисто-подзолисто-глееватая гумусовая |
* В скобках указан возраст, лет. **Рассчитан по вкладу пород в общие запасы древесины: С – сосна (Pinus sylvestris L.), Лц – лиственница (Larix sibirica Ledeb.), Б – береза (Betula pubescens Ehrh.), Е – ель (Picea obovata Ledeb.); *** Рассчитано по данным, приведенным в работе (Бобкова, 1987); **** Опубликовано в (Осипов, Бобкова, 2016); ***** Собственные неопубликованные данные.
Для оценки нетто-продукции древостоев применяли метод модельных деревьев (Усольцев, 2007). В сосняках на Зеленоборском лесном стационаре проанализировано 21 модельное дерево сосны, 6 лиственниц, 5 берез, 3 ели из разных ступеней толщины, а в сосняках на Чернамском лесном стационаре 16 деревьев сосны разных ступеней толщины. Для оценки продукции ели и березы в сосняке черничном отобрано по одному среднему дереву соответствующих пород. Прирост хвои оценивали как средний по вкладу хвои четырех лет в общую массу. Текущий прирост стволовой древесины определяли как среднее за последние пять лет по спилам древесины ствола, отобранным на уровне корневой шейки, высотах 1 и 1.3 м, далее через каждые 2 м до верхушки. Анализ радиального прироста древесины ствола проводили по четырем радиусам с использованием Lintab 5 и программы Tsap Win. По этим данным строили ход роста дерева по высоте и диаметру. Прирост ветвей определяли по средней ветви, отобранной с середины кроны каждого дерева, как сумму среднего прироста главной ветви I-го порядка и всех ветвей II-го порядка и по формуле:
где I – прирост средней ветви (единицы массы), MbrI – масса ветви I-го порядка (единицы массы), MbrII – масса ветви II-го порядка (единицы массы), А – возраст ветви (лет).
На основе полученных данных по продукции массы отдельных фракций модельных деревьев выводили регрессионные уравнения зависимости их прироста от диаметра ствола на высоте 1.3 м, для оценки формирования органического вещества по данным перечета деревьев на постоянных пробных площадях. Характеристики степенных уравнений приведены в табл. 2. Прирост коры сосны в насаждениях средней тайги принимали равным ее опаду, который определялся при помощи опадоуловителей, а в сосняках северной тайги − по соотношению между массой коры и древесины модельных деревьев. Прирост корней деревьев определяли по формуле (Андреева и др., 2002):
Таблица 2.
Показатели степенных (I = aDb) уравнений взаимосвязи прироста массы отдельных фракций деревьев (I, кг) с диаметром (D, см) ствола на высоте 1.3 м
| Фракция | Коэффициенты | SEE | ||
|---|---|---|---|---|
| а | b | детерминации (R2) | ||
| Северная тайга | ||||
| Сосна (зеленомошная группа типов леса): | ||||
| хвоя | 0.0052 | 1.971 | 0.91 | 0.45 |
| ветви | 0.0022 | 1.9176 | 0.86 | 0.14 |
| древесина | 0.0251 | 1.544 | 0.81 | 0.62 |
| кора | 0.001 | 1.77 | 0.77 | 0.06 |
| Сосна (сфагновый тип леса): | ||||
| хвоя | 0.0026 | 2.2877 | 0.93 | 0.09 |
| ветви | 0.0021 | 1.823 | 0.84 | 0.06 |
| древесина | 0.013 | 1.4431 | 0.87 | 0.06 |
| кора | 0.0009 | 1.5331 | 0.71 | 0.27 |
| Лиственница (зеленомошная группа типов леса) | ||||
| хвоя | 0.002 | 2.4308 | 0.91 | 0.31 |
| древесина | 0.0051 | 2.0606 | 0.96 | 0.19 |
| ветви | 0.0011 | 2.1357 | 0.93 | 0.11 |
| кора | 0.0006 | 1.7866 | 0.56 | 0.04 |
| Береза (зеленомошная группа типов) | ||||
| листья | 0.0347 | 1.3883 | 0.98 | 0.04 |
| древесина | 0.0038 | 2.0267 | 0.63 | 0.14 |
| кора | 0.0002 | 2.1299 | 0.73 | 0.01 |
| ветви | 0.0003 | 2.4745 | 0.63 | 0.04 |
| Средняя тайга | ||||
| Сосна (бруснично-лишайниковый сосняк) | ||||
| хвоя | 0.001 | 2.4222 | 0.95 | 0.22 |
| ветви | 0.0008 | 2.2734 | 0.78 | 0.20 |
| древесина | 0.0013 | 1.7991 | 0.80 | 0.06 |
| Сосна (зеленомошная группа типов леса): | ||||
| хвоя | 0.002 | 2.07 | 0.95 | 0.18 |
| ветви | 0.004 | 1.65 | 0.65 | 0.23 |
| древесина | 0.009 | 1.74 | 0.96 | 0.05 |
| Сосна (чернично-сфагновый тип леса): | ||||
| хвоя | 0.0016 | 2.2767 | 0.85 | 0.17 |
| ветви | 0.0052 | 1.7887 | 0.44 | 0.58 |
| древесина | 0.0049 | 2.2412 | 0.71 | 0.73 |
где, Пкр – прирост корней (единицы массы); nс – прирост стволов и ветвей (единицы массы); k – доля корней от суммарной массы ствола, ветвей и корней, %; с – доля ствола и ветвей от суммарной массы ствола, ветвей и корней, %.
Прирост отдельных фракций деревьев ели в сосняке чернично-сфагновом рассчитывали по уравнениям, выведенным для среднетаежного ельника чернично-сфагнового (Кузнецов, 2010).
Формирование органического вещества надземными органами растений напочвенного покрова оценивали по модельным экземплярам надземных частей кустарничков, отобранных в 20–100-кратной повторности (в зависимости от его встречаемости) в августе. У них отделяли побеги текущего года и по соотношению с оставшейся частью рассчитывали долю побегов первого года. В дальнейшем эти данные переводились на массу каждого вида растений напочвенного покрова в насаждениях. Для среднетаежного чернично-сфагнового и сосновых насаждений северной тайги ежегодный прирост растений напочвенного покрова получены расчетным путем. Так, продукцию черники принимали равной 17%, брусники − 16, зеленых мхов − 20, сфагновых − 15 (Казимиров и др., 1977), а подземных органов трав и кустарничков – 25% от их массы (Эколого-физиологические…, 1993).
Статистическая обработка полученных данных выполнена с использованием Microsoft Excel. Рассчитывались средние значения и стандартная ошибка среднего. Регрессионный анализ выполнен при 95%-ом уровне значимости. Для регрессионных уравнений рассчитывался коэффициент детерминации и стандартная ошибка уравнения регрессии (SEE).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
NPP углерода в древостоях. Для определения NPP углерода в древостоях сосняков мы применяли степенные (y = a Db) уравнения зависимости прироста массы отдельных фракций от диаметра ствола на высоте 1.3 м. Тип уравнения был выбран исходя из анализа характера кривой, которая не должна пересекать ось абсцисс и давать отрицательные значения продукции. Как представлено в табл. 2, полученные уравнения имеют достаточно высокие показатели аппроксимации тренда, которые варьируют от 0.44 до 0.99.
На современном этапе исследований для определения нетто-продукции углерода лесных насаждений достаточно широко применяется метод микровихревых пульсаций (Metsaranta et al., 2018 и др.), а в перспективе можно рассматривать конверсию данных фитомассы, полученных при аэрофотосъемке с использованием LiDAR (Phua et al., 2017). Однако эти методы дорогостоящие и имеют определенные ограничения. Поэтому при определении бюджета углерода в лесных насаждениях по биологической продуктивности применение регрессионных уравнений для насаждений сходных лесорастительных условий и возраста до сих пор является оправданным. Помимо диаметра в качестве входящих параметров для регрессионных уравнений при характеристике прироста отдельных фракций деревьев сосны возможно использование других показателей, таких как высота деревьев, протяженность и площадь кроны (Vanninen, Mäkelä, 2000), с которыми также отмечаются тесные корреляционные связи. Эти показатели часто имеют положительную взаимосвязь с диаметром ствола, однако их измерение более трудоемко и менее точно.
Прирост органического вещества в древостоях приспевающих и спелых сосняков разных типов в условиях северной тайги изменяется от 0.7 до 2.4, средней – от 1.4 до 1.8 т С га–1 год–1 (рис. 1). В чистых по составу древостоях среднетаежных сосняков практически вся продукция растительного органического вещества формируется деревьями сосны. Суммарное участие березы и ели в накоплении органического вещества не превышает 1–2%. В смешанных по составу древостоях сосняков северной тайги на автоморфных почвах вклад деревьев сосны в накопление их годичной продукции варьирует от 52 до 77%, с более высоким значением в сосняке голубично-зеленомошном, произрастающем на относительно влажном контактно-глеевом подзоле. В сосняке осоково-сфагновом на гидроморфной почве участие деревьев сосны в NPP достигает 90%. Береза встречается в составе древостоев всех исследованных насаждений, а ее доля в нетто-продукции углерода изменяется в довольно широких пределах − от 2 до 21%. В северотаежных сосняках зеленомошной группы типов, развитых на автоморфных почвах, значительное количество углерода (15–19%) депонируют деревья лиственницы, тогда как участие ели в накоплении углерода этих сообществ изменяется от 1 до 19%.
Рис. 1.
Нетто-продукция углерода фитомассы древостоев и вклад в нее отдельных органов древесных растений в сосняках.
A – черничный; B – чернично-зеленомошный; C – голубично-зеленомошный; D – осоково-сфагновый; E – бруснично-лишайниковый; F – черничный; G – чернично-сфагновый.
1 – хвоя; 2 – ветви; 3 – древесина ствола; 4 – кора ствола; 5 – корни.
Указана общая нетто-продукция углерода древостоя, т га–1 год–1.
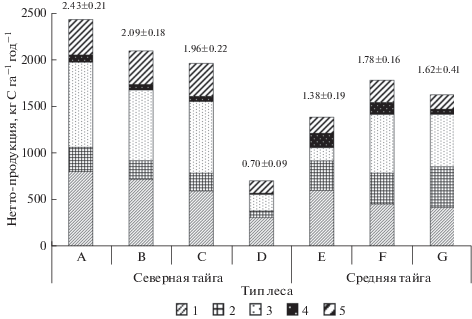
В общем приросте углерода органического вещества древостоев северотаежных сосняков значительная доля приходится на хвою/листья (30–44%) и древесину ствола (25–39%). Участие корней в депонировании углерода составляет 16–19%, ветвей − 9–11, коры – 2–3%. В среднетаежном сосняке бруснично-лишайниковом в NPP высока доля участия хвои (43%) и ветвей (23%) сосны, а вклад древесины ствола, коры и корней составляет 10, 11 и 12%, соответственно. В среднетаежных сосняках черничном и чернично-сфагновом на торфянисто-подзолистых почвах отмечаются некоторые сходные тенденции. Чуть более трети (35%) NPP формирует древесина ствола, в хвое ежегодно накапливается 25% от общей продукции. Следует отметить значимую роль ветвей (19–27%) в депонировании углерода, что свидетельствует об активном освоении кронами деревьев окружающего пространства древесным ярусом. Вклад корней и коры составляет 10–13 и 3–7%, соответственно.
Известно, что продуктивность древостоев снижается при продвижении к северу, хотя для исследованных фитоценозов зеленомошного типа эта закономерность не проявляется. Как было отмечено выше, в накоплении углерода в древостоях северотаежных сосняков значительна роль лиственницы и березы, участвующих в его составе. Величины, характеризующие темп связывания углерода только деревьями сосны, показывают, что депонирование углерода в сосняках средней тайги в 1.2–1.5 раза выше, чем в северной.
Отмечено, что во всех насаждениях исследованных сосняков более высокая скорость накопления углерода древостоя характерна для древесины ствола и ассимиляционного аппарата. Схожая закономерность была ранее выявлена для сосняков Карелии (Казимиров и др., 1977) и Финляндии (Vanninen, Mäkelä, 2000). Авторы отмечают, что распределение продукции фитомассы по отдельным органам обусловлено приоритетом и необходимостью древесных растений в осуществлении процессов жизнедеятельности на отдельных этапах развития. Так, небольшая продолжительность жизни и большой опад органов ассимиляционного аппарата требуют регулярного его обновления за счет прироста. Данные как наших (Бобкова, 2005; Бобкова, Осипов, 2012; и др.), так и других (Казимиров и др., 1977; Никонов, 1987; Bhatti, Jassal, 2014) исследований показывают, что более половины массы надземного древесного опада в сосняках приходится на хвою. Высокая доля ветвей в приросте древостоев среднетаежных сосняков, отличающихся большей полнотой по сравнению с северотаежными, свидетельствует о необходимости освоения окружающего пространства в условиях высокой конкуренции за свет между деревьями. Вместе с тем, спелые сосняки в условиях севера таежной зоны отличаются относительно высокими темпами накопления углерода органического вещества стволовой древесины, что характеризует активное депонирование углерода древостоями в этом возрасте.
Прирост растений напочвенного покрова. Растения напочвенного покрова в сосняках северной тайги ежегодно формируют 1.4–1.6, средней – 0.5–0.8 т С/га в год (рис. 2). Более половины общей продукции углерода растениями этого яруса фитоценоза концентрируется в подземных органах, участие которых выше в сообществах на полугидроморфных почвах. В надземной части растений напочвенного покрова закономерностей во вкладе его отдельных компонентов в нетто-продукцию не выявлено, а их участие варьирует в широких пределах. Основными поглотителями углерода в этом ярусе являются виды-доминанты. Кустарнички в зеленомошных сосняках представлены черникой, голубикой, брусникой, травянистые растения – осокой, ожикой, иван-чаем. В сфагновых типах сообществ встречаются влаголюбивые растения – багульник, кассандра, подбел. Сплошной моховой покров в сосняках зеленомошной группы типов образуют Pleurozium schreberi, Hylocomium splendens, единично встречаются Polytrichum juniperinum и P. commune, в сфагновой группе типов − Sphagnum warnstorfii, S. angustifolium, S. medium. Лишайниковый покров в сосняке бруснично-лишайниковом образован представителями рода Cladonia и Cladina, которые также образуют небольшие пятна в сосняках зеленомошной группы типов.
Рис. 2.
Нетто-продукция углерода фитомассы растений напочвенного покрова сосняков.
A – черничный; B – чернично-зеленомошный; C – голубично-зеленомошный; D – осоково-сфагновый; E – бруснично-лишайниковый; F – черничный; G – чернично-сфагновый.
1 – кустарнички; 2 – травы; 3 – мхи и лишайники; 4 – корни трав и кустарничков.
Указана нетто-продукция растений напочвенного покрова в фитоценозе, т С га–1 год–1 (вверху) и доля участия отдельных компонентов, %.
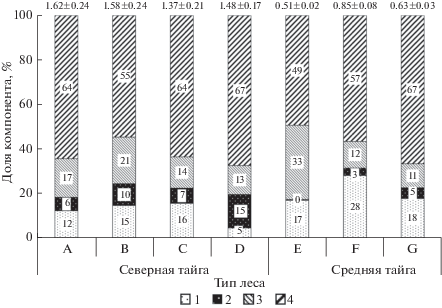
Общая нетто-продукция углерода. Суммарная текущая продукция углерода фитомассы в северотаежных сосняках экологического ряда изменяется от 2.2 до 4.1, среднетаежных – от 1.9 до 2.6 т га–1 год–1 (табл. 3). По отношению к общей массе фитоценозов он колеблется от 2 до 7%. Древостои сосняков на автоморфных почвах в условиях северной тайги формируют 57–60% от общего количества углерода фитомассы, тогда как в заболоченном сосняке на гидроморфной почве доля древостоя уменьшается до 32%. В среднетаежных сосняках вклад древостоев в нетто-продукцию углерода выше по сравнению с северотаежными и составляет 68–73%. Ранее нами (Осипов, Бобкова, 2016) было отмечено, что соотношение между приростом напочвенного покрова и древостоя изменяется по мере возрастного развития насаждения. Усиление средообразующей роли древостоя и увеличение его полноты при переходе из средневозрастного в приспевающий и спелый уменьшает поступление солнечной радиации к почве, что приводит к сокращению проективного покрытия растительности нижних ярусов и снижению темпов накопления органического вещества. Меньшая полнота древостоев северотаежных сосняков способствует пропусканию света сквозь полог деревьев. Согласно Э.П. Галенко (1983), в северотаежных сосняках под полог древостоя поступает 30–60% фотосинтетически активной радиации, тогда как в среднетаежных – 12–36%. В результате фитомасса растений напочвенного покрова в сообществах северотаежных сосняков несколько выше, чем в среднетаежных (табл. 1).
Таблица 3.
Общая продукция углерода фитомассы и коэффициенты, характеризующие отношение нетто-продукции (NPP, т га–1 год–1) с запасами древесины (М, м3 га–1) и углеродом фитомассы древостоя (Ph, т га–1) в сосняках
| Сообщество | Нетто-продукция, т га–1 год–1 | Отношение* | |||
|---|---|---|---|---|---|
| древостоя | напочвенного покрова | общая | общая продукция | древостой | |
| Северная тайга | |||||
| Чернично-зеленомошный | 2.43 | 1.62 | 4.05 | $\frac{{0.0265}}{{0.0749}}$ | $\frac{{0.0159}}{{0.0449}}$ |
| Голубично-зеленомошный | 2.09 | 1.58 | 3.67 | $\frac{{0.0294}}{{0.0771}}$ | $\frac{{0.0167}}{{0.0439}}$ |
| Черничный | 1.96 | 1.37 | 3.33 | $\frac{{0.0187}}{{0.0537}}$ | $\frac{{0.0110}}{{0.0316}}$ |
| Осоково-сфагновый | 0.70 | 1.48 | 2.18 | $\frac{{0.0357}}{{0.0930}}$ | $\frac{{0.0115}}{{0.0299}}$ |
| Средняя тайга | |||||
| Бруснично-лишайниковый | 1.38 | 0.51 | 1.89 | $\frac{{0.0077}}{{0.0228}}$ | $\frac{{0.0056}}{{0.0166}}$ |
| Черничный | 1.78 | 0.85 | 2.63 | $\frac{{0.0120}}{{0.0396}}$ | $\frac{{0.0081}}{{0.0268}}$ |
| Чернично-сфагновый | 1.62 | 0.63 | 2.25 | $\frac{{0.0162}}{{0.0417}}$ | $\frac{{0.0117}}{{0.0301}}$ |
Расчеты депонирования углерода фитомассы в сосняках, проведенные на основе результатов исследований прироста органического вещества в сосняках европейского Севера (Казимиров и др., 1977; Никонов, 1987), показали, что полученные нами данные NPP сопоставимы с результатами для Кольского полуострова, но ниже, чем в Карелии. Так, формирование углерода органического вещества северотаежными сосняками Кольского полуострова изменяется в пределах 2.0–3.5 т С га–1 год–1, среднетаежными в условиях Карелии – от 4.2 до 9.6 т С га–1 год–1.
На соотношение участия отдельных компонентов фитоценоза в накоплении углерода органического вещества в сосняках значимое влияние оказывают условия произрастания. Показано (Казимиров и др., 1977; Nilsson, Wardle, 2005), что в низкопродуктивных сообществах продукция растений напочвенного покрова и древостоя незначительно отличаются друг от друга, тогда как в более продуктивных насаждениях нетто-продукция древесного яруса ~ в 1.5 раза выше. Приведенные данные свидетельствуют, что влажность почв сосновых лесов является фактором, определяющим вклад растений отдельных ярусов фитоценоза в формирование органического вещества. В настоящей и более ранних работах (Кутявин и др., 2016) показано, что в среднетаежных сосняках лишайниковых, произрастающих на бедных песчаных почвах, вклад растений напочвенного покрова составляет 24–27%. Более развитый кустарничковый и моховой покров в сосняках на болотно-подзолистых почвах способствует увеличению доли участия в продукции растений нижних ярусов до 32–49%. Несмотря на небольшую разницу вклада отдельных компонентов фитоценоза в NPP в относительных величинах, она становится более выраженной в абсолютных значениях (табл. 3).
Отношение первичной продукции к запасу древесины и углерода в древостоях. Как показано в табл. 3, отношение между общим приростом углерода фитомассы и запасом стволовой древесины изменяется от 0.0077 до 0.0357 т м–3 га–1, а отношение между NPP и углеродом фитомассы древостоя – от 0.0228 до 0.0930 т т–1 га–1. Среднее значение этих коэффициентов составляет 0.0276 ± 0.004 и 0.0119 ± 0.002 т м–3 га–1 и 0.0747 ± 0.008 и 0.0347 ± ± 0.006 т т–1 га–1 для сосняков северной и средней тайги, соответственно. Более высокие показатели, отмеченные для северотаежных сосняков, объясняются, во-первых, сложным породным составом древостоев, что способствует более интенсивному накоплению в них органического вещества; во-вторых, относительно высокими показателями формирования фитомассы растениями напочвенного покрова.
Коэффициенты, характеризующие отношение между NPP древостоя и запасом древесины/углерода фитомассы в нем, представляют больший интерес. Это связано с тем, что углерод, поглощенный древостоем, в первую очередь рассматривается при оценках его депонирования лесными насаждениями из-за меньшей скорости оборота, по сравнению с растениями напочвенного покрова. Рассчитанная средняя величина для конверсии данных по запасам древесины древостоя в его нетто-продукцию в исследованных нами насаждениях составляет 0.0115 ± 0.0015 т м–3 га–1, для перевода запасов углерода в древостое в его прирост – 0.0320 ± 0.0037 т т–1 га–1с более высокими значениями в северотаежных сосняках.
Следует отметить, что полученные нами коэффициенты, отражающие взаимосвязь нетто-продукции и запасов древесины в древостое, несколько ниже приведенных в работе (Замолодчиков, Уткин, 2000). Авторы отмечают, что в использованной ими базе данных по биологической продуктивности лесов большая часть результатов приведена для чистых по составу, высокобонитетных древостоев. Большинство из исследованных нами древостоев смешанные по составу, невысокого класса бонитета, что, вероятно, и послужило причиной более низких показателей конверсионных отношений.
В анализе литературы, приведенном в работе (Keeling, Phillips, 2007), показано, что теоретическая взаимосвязь углерода фитомассы и нетто-продукции углерода обусловлена несколькими причинам: во-первых, фитомасса является результатом нетто-продукции, во-вторых, оба показателя лимитируются общими почвенно-экологическими факторами, такими как режим температуры и влажности, содержание питательных элементов. На наш взгляд выведение конверсионных коэффициентов между запасами углерода фитомассы древостоя и нетто-продукции более целесообразно по сравнению с конверсионным коэффициентом нетто-продукция:запас древесины древостоя. Это связано с активным развитием методов дистанционного зондирования углерода фитомассы лесов (Исаев и др., 2014; Щепащенко и др., 2017), по которым в дальнейшем возможен расчет нетто-продукции, лесных экосистем на основе коэффициентов, зависящих от возраста и породного состава насаждений.
Заключение. На европейском Северо-Востоке России в условиях северной и средней тайги приспевающие и спелые сосновые фитоценозы депонируют 1.9–4.1 т С га–1 год–1. В общей продукции углерода в сосняках на долю древостоя приходится 32–73%. Характерно, что по мере продвижения на север в пределах типа леса и с повышением влажности в пределах растительных подзон значительно уменьшается относительная масса продукции углерода древостоя и повышается доля растений напочвенного покрова. В целом в сосновых сообществах в годичном депонировании углерода фитомассы довольно высока значимость травяно-кустарничкового и мохово-лишайникового ярусов растительности. Следует отметить, что в структуре годичного прироста углерода фитомассы в сосновых фитоценозах преобладают надземные части, в которых существенную долю составляют ассимиляционные органы. С усилением заболоченности почв в накоплении углерода увеличивается доля фотосинтетического аппарата и корней, уменьшается участие многолетних надземных органов. Выведенные уравнения взаимосвязи продукции углерода отдельными органами древесных растений от диаметра ствола могут быть использованы для оценки бюджета углерода в сосновых экосистемах Севера.
Список литературы
Андреева Е.Н., Баккал И.Ю., Горшков В.В., Лянгузова И.В., Мазная Е.А., Нешатаев В.Ю., Нешатаева В.Ю., Ставрова Н.И., Ярмишко В.Т., Ярмишко М.А. Методы изучения лесных сообществ. СПб.: НИИХимии СПбГУ, 2002. 240 с.
Бобкова К.С. Биологическая продуктивность хвойных лесов европейского Северо-Востока. Л.: Наука, 1987. 156 с.
Бобкова К.С. Биологическая продуктивность и компоненты баланса углерода в молодняках сосны // Лесоведение. 2005. № 6. С. 30–37.
Бобкова К.С., Осипов А.Ф. Круговорот углерода в системе “фитоценоз−почва” в чернично-сфагновых сосняках средней тайги Республики Коми // Лесоведение. 2012. № 2. С. 11–18. КРУГОВОРОТ УГЛЕРОДА
Галенко Э.П. Фитоклимат и энергетические факторы продуктивности хвойного леса Европейского Севера. Л.: Наука, 1983. 129 с.
Замолодчиков Д.Г., Уткин А.И. Система конверсионных отношений для расчета чистой первичной продукции лесных экосистем по запасам насаждений // Лесоведение. 2000. № 6. С. 54–63.
Исаев А.С., Коровин Г.Н. Лес как национальное достояние России //Лесоведение. 2013. № 5. С. 5–12.
Исаев А.С., Барталев С.А., Лупян Е.А., Лукина Н.В. Спутниковое зондирование Земли – уникальный инструмент мониторинга лесов России // Вестник Российской академии наук. 2014. Т. 84. № 12. С. 1073–1079.
Исаев А.С., Овчинникова Т.М., Суховольский В.Г. Распределение фитомассы деревьев и насаждений по фракциям: модель конкуренции // Проблемы экологического мониторинга и моделирования экосистем. СПб.: Гидрометеоиздат, 2007. Т. 21. С. 232–250.
Исаев А.С., Коровин Г.Н., Уткин А.И., Пряжников А.А., Замолодчиков Д.Г. Оценка запасов и годичного депонирования углерода в фитомассе лесных экосистем России // Лесоведение. 1993. № 5. С. 3−10.
Казимиров Н.И., Волков А.Д., Зябченко С.С., Иванчиков А.А., Морозова Р.М. Обмен веществ и энергии в сосновых лесах Европейского Севера. Л.: Наука, 1977. 304 с.
Кутявин И.Н. Сосновые леса северного Приуралья: строение, рост, продуктивность. Сыктывкар: Изд-во ИБ Коми НЦ УрО РАН, 2018. 176 с.
Кутявин И.Н., Торлопова Н.В., Осипов А.Ф., Кузьмина Е.С., Бобкова К.С. Биологическая продуктивность коренного среднетаежного сосняка бруснично-лишайникового (средняя Печора) // Растительные ресурсы. 2016. № 4. С. 484–500.
Кузнецов М.А. Динамика содержания органического углерода в заболоченных ельниках средней тайги. Автореф. дисс. к.б.н. Сыктывкар: Институт биологии Коми НЦ УрО РАН, 2010. 20 с.
Лукина Н.В., Исаев А.С., Крышень А.М., Онучин А.А., Сирин А.А., Гагарин Ю.Н., Барталев С.А. Приоритетные направления развития лесной науки как основы устойчивого управления лесами // Лесоведение. 2015. № 4. С. 243–254.
Никонов В.В. Почвообразование на северном пределе сосновых биогеоценозов. Л.: Наука, 1987. 142 с.
Осипов А.Ф., Бобкова К.С. Биологическая продуктивность и фиксация углерода среднетаежными сосняками при переходе из средневозрастных в спелые // Лесоведение. 2016. № 5. С. 346–354.
Усольцев В.А. Биологическая продуктивность лесов Северной Евразии: методы, база данных и ее приложения. Екатеринбург: Изд-во УрО РАН, 2007. 637 с
Щепащенко Д.Г., Швиденко А.З., Пергер К., Дресел К., Фриц Ш., Лакида П.И., Мухортова Л.В., Усольцев В.А., Бобкова К.С., Осипов А.Ф., Мартыненко О.В., Карминов В.Н., Онтиков П.В., Щепащенко М.В., Кракснер Ф. Изучение фитомассы лесов: текущее состояние и перспективы // Сибирский лесной журнал. 2017. № 4. С. 3−11.
Эколого-физиологические основы продуктивности сосновых лесов европейского Северо-Востока / Под ред. К.С. Бобковой. Сыктывкар: Коми НЦ УрО РАН,1993. 176 с.
Bhatti J.S., Jassal R.S. Long term aboveground litterfall production in boreal jack pine (Pinus banksiana) and black spruce (Picea mariana) stands along the Boreal Forest Transect Case Study in western central Canada // Écoscience. 2014. V. 21 (3−4). P. 301−314.
FAO, 2015. http://www.fao.org/3/a-az316e.pdf, (14 May 2019).
Hardiman B.S., Gough C.M., Halperin A., Hofmeister K.L., Nave L.E., Bohrer G., Curtis P.S. Maintaining high rates of carbon storage in old forests: A mechanism linking canopy structure to forest function // Forest Ecology and Management. 2013. V. 298. P. 111–119
Isaev A., Korovin G., Zamolodchikov D., Utkin A., Pryaznikov A. Carbon stock and deposition in phytomass of the Russian forests // Water, Air and Soil Pollution. 1995. V. 82. P. 247–256.
Keeling H.C., Phillips O.L. The global relationship between forest productivity and biomass // Global Ecology and Biogeography. 2007. V. 16. Is. 5. P. 618–631.
Luo Y., Keenan T. F., Smith M. Predictability of the terrestrial carbon cycle // Global Change Biology. 2015. V. 21. Is. 5. P. 1737–1751.
Luyssaert S., Schulze E.–D, Börner A., Knohl A., Hessenmöller D., Law B.E., Ciais P., Grace J. Old-growth forests as global carbon sinks // Nature. 2008. V. 455. P. 213–215.
Metsaranta J.M., Trofymow J.A., Black T.A., Jassal R.S. Long-term time series of annual ecosystem production (1985–2010) derived from tree rings in Douglas-fir stands on Vancouver Island, Canada using a hybrid biometric-modelling approach // Forest Ecology and Management. 2018. Vol. 429. P. 57–68.
Nilsson M.-C., Wardle D.A. Understory vegetation as a forest ecosystem driver: evidence from the northern Swedish boreal forest // Frontiers in Ecology and the Environment. 2005. V. 3(8). P. 421–428.
Phua M.-H., Johari S.A., Wong O.C., Ioki K., Mahali M., Nilus R., Coomes D.A., Maycock C.R., Hashim M. Synergistic use of Landsat 8 OLI image and airborne LiDAR data for above-ground biomass estimation in tropical lowland rainforests // Forest Ecology and Management. 2017. V. 406. P. 163–171
Schaphoff S., Reyer C.P.O., Schepaschenko D., Gerten D., Shvidenko A. Tamm Review: Observed and projected climate change impacts on Russia’s forests and its carbon balance // Forest Ecology and Management. 2016. V. 361. P. 432–444.
Vanninen P., Mäkelä A. Needle and stem wood production in Scots pine (Pinus sylvestris) trees of different age, size and competitive status // Tree Physiology. 2000. V. 20. Is. 8. P. 527–533
Vedrova E.F., Mukhortova L.V., Trefilova O.V. Contribution of old growth forests to carbon budget of the boreal zone in Central Siberia // Biology Bulletin. 2018. V. 45. Is. 3. P. 288–297.
Дополнительные материалы отсутствуют.