

Лесоведение, 2020, № 1, стр. 64-75
Биологическая продуктивность гидрофильных сероольшаников Ярославской области
Я. И. Гульбе a, *, Т. А. Гульбе a, А. Я. Гульбе a, Л. С. Ермолова a
a Институт лесоведения РАН
143030 Одинцовский г. о., Московская обл., с. Успенское, ул. Советская, 21, Россия
* E-mail: goulbe@ilan.ras.ru
Поступила в редакцию 26.12.2018
После доработки 08.04.2019
Принята к публикации 08.10.2019
Аннотация
Определена фитомасса и годичная продукция ольхи серой (Alnus incana (L.) Moench) по фракциям в 12-, 13-, 22-, 31-, 34- и 41-летнем гидрофильных (таволговых) сероольшаниках. В 41-летнем древостое гидрофильного сероольшаника надземная фитомасса достигает 141.46 т га–1, доля стволов в ней составляет 85.2% (120.53 т га–1), скелета крон – 12.1% (17.15 т га–1), листьев – 2.6% (3.64 т га–1), генеративных органов – 0.1% (0.14 т га–1). Установлено, что гидрофильные сероольшаники по запасу древесины, массе стволов и скелета крон ольхи серой сопоставимы с неморально-кисличными сероольшаниками близкого возраста. В ходе их роста и развития выявлен инвариантный период, в течение которого величина годичной продукции надземной части древостоя мало зависит от возраста и близка к максимальной для данных условий произрастания (8–9 т га–1 год–1) с относительно стабильным распределением по фракциям (доля стволов составляет 47.0%, скелета крон – 19.7%, листьев – 33.3%). С использованием инвариантного периода в качестве базового проведен сравнительный анализ продукционного потенциала древостоев гидрофильных и неморально-кисличных сероольшаников. По величине годичной продукции древостоев гидрофильные сероольшаники уступают неморально-кисличным. По сравнению с неморально-кисличными сероольшаниками они характеризуются меньшими значениями годичной продукции стволов, которые не компенсируются незначительным увеличением годичной продукции листьев при близких значениях годичной продукции скелета крон, а также более низким коэффициентом продуктивности ассимиляционного аппарата.
В результате длительного антропогенного воздействия породный состав южнотаежных лесов Русской равнины претерпел существенные изменения: на больших площадях хвойные леса сменились мягколиственными. С увеличением доли мягколиственных пород потребовалась переоценка их биосферной роли и хозяйственного значения, и в том числе ольхи серой. Обладая мощным возобновительным потенциалом, ольха серая в подзоне южной тайги активно участвует в демутации лесной растительности на вырубках и вышедших из-под сельскохозяйственного пользования землях, заселяет эродированные и скарифицированные участки. Сероольшаники благодаря быстрому росту, интенсивному круговороту веществ, фиксации атмосферного азота привлекают все большее внимание и как объект плантационного выращивания для получения древесной массы и энергетического сырья, и как сообщества, имеющие важное средообразующее значение. Выполнение сероольшаниками и хозяйственных, и экологических функций в значительной степени связано с их биологической продуктивностью. Для научного обоснования системы лесохозяйственных мероприятий в этих насаждениях возникла необходимость оценки и анализа продуцирования и накопления ими органического вещества.
Несмотря на довольно широкую экологическую амплитуду, ольха серая образует сомкнутые фитоценозы лесного облика в сравнительно узких эдафических пределах. Их рамки более сжаты, чем те, которые характерны для осинников и, тем более, березняков (Юркевич и др., 1963; Ярмишко, 1976). В Ярославской области сероольшаники представлены в основном двумя примерно равными по площади группами типов леса: неморально-кисличной и гидрофильной (Лесной план …, 2011). Первичная биологическая продуктивность неморально-кисличных сероольшаников, расположенных на суходоле, была определена нами ранее (Уткин и др., 1980, 1982; Гульбе, 2012, 2017). Гидрофильные или таволговые сероольшаники типичны для более влажных местообитаний. Они располагаются в нижних частях пологих склонов в поймах и излучинах ручьев и речек, формируются на дерново-подзолисто-глеевой и перегнойно-глеевой почве и характеризуются переменным режимом увлажнения проточными водами (Осипов и др., 1977; Борисова и др., 2012). Приуроченные к низинным местоположениям гидрофильные сероольшаники имеют большое почвозащитное и водоохранное значение. В Европе прибрежные сероольшаники расцениваются как экосистемы, способные аккумулировать и трансформировать потоки загрязняющих веществ с расположенных рядом интенсивно эксплуатируемых территорий. Такие насаждения ольхи обычно используются в качестве буферной зоны для защиты водных объектов от загрязнения (Mander et al., 1995; Mander et al., 1997; Lohmus et al., 1996). Как в России, так и в странах Восточной Европы и Скандинавии биологическая продуктивность гидрофильных (таволговых) сероольшаников практически не изучалась (Усольцев, 2010; Aosaar et al., 2012). Задачей проводимых исследований стало определение биологической продуктивности гидрофильных (таволговых) сероольшаников с учетом возрастной динамики и сравнительная оценка продукционного потенциала ольхи серой в различных почвенно-гидрологических условиях.
ОБЪЕКТЫ И МЕТОДИКА
Исследования проводились в Ярославской области на базе стационара “Городище” Северной лесной опытной станции Института лесоведения РАН (57°42′ с.ш., 38°34′ в.д.). Объектами исследований служили 6 участков чистых сомкнутых 12-, 13-, 22-, 31-, 34- и 41-летних гидрофильных сероольшаников. Почвы рассматриваемых насаждений дерново-грунтово-глееватые, легкосуглинистые, среднегумусные на покровных отложениях, подстилаемые аллювиальными отложениями с повышенным увлажнением. В пределах участков были заложены пробные площади, на которых проведено геоботаническое обследование, выполнена инструментальная таксация древостоев и по модельным деревьям определены их запас, прирост по объему, фитомасса и годичная продукция.
Травяной покров гидрофильных (таволговых) сероольшаников высокий, густой с общим покрытием 75–95%, обычно формирует 2 (3) подъяруса. Верхний представлен видами крупнотравья: Filipendula ulmaria (L.) Maxim., Urtica dioica L., Angelica sylvestris L., Athyrium filix femina (L.) Roth, Cirsium oleracium (L.) Scop., Lysimachia vulgaris L., Rubus idaeus L. Второй подъярус формируют A-egopodium podagraria L., Equisetum pratense Ehrh, Geum rivale L., в молодых фитоценозах присутствует Impatiens noli-tangere L. При более сложной структуре травяного покрова образуется 3-й подъярус из Cardamina impatiens L., Chrisosplenium alternifolium L., Ranunculus repens L., Stellaria nemorum L. Для этого типа сероольшаников характерно частое присутствие Solanum dulcamara L., а после 20-летнего возраста – Humulus lupulus L.
Подлесок в сероольшаниках этого типа сомкнутого полога не образует, хотя общее число его видов достигает 10. На всех стадиях их развития в подлеске в значительном количестве присутствуют черемуха (Padus avium Mill.), рябина (Sorbus aucuparia L.), ивы (Salix sp.), смородина красная (Ribes rubrum L.) и черная (R. nigrum L.). С 15-летнего возраста в состав подлеска обычно внедряются жимолость (Lonicera xyilosteum L.), крушина (Frangula alnus Mill.), калина (Viburnum opulus L.) (Ермолова и др., 2013). В обследованных сероольшаниках подлесок высотой более 1.3 м, представленный зачастую черемухой, рябиной, ивами, реже крушиной, калиной, имеет густоту от 0.30 до 1.20 тыс. экз. га–1 и лишь в 31-летнем насаждении − 4.93 тыс. экз. га–1. Средний диаметр подлеска в сероольшаниках до 30 лет не превышает 1.5 см, в более старых варьирует в пределах 2.0–2.6 см.
В подросте встречаются ольха серая, береза, ель. Густота подроста высотой более 1.3 м в большинстве случаев не превышает 1.50 тыс. экз. га–1. Исключение составляют 22- и 41-летний сероольшаники, где подрост ольхи вегетативного происхождения (в виде корневых отпрысков) достигает 2.90 и 6.00 тыс. экз. га–1, соответственно. Немногочисленный подрост ели отмечен в двух сероольшаниках: в 13-летнем насаждении он составляет 0.10 тыс. экз. га–1, в 34-летнем – 0.82 тыс. экз. га–1. Незначительное количество и биометрические показатели подроста и подлеска в гидрофильных сероольшаниках свидетельствуют, что вклад этих компонентов в фитомассу и годичную продукцию рассматриваемых фитоценозов незначителен.
Модельные деревья для определения запаса, прироста по объему, фитомассы и годичной продукции древостоя (по 10–12 экз. на пробной площади) брали в августе методом пропорционально-ступенчатого представительства (по ступеням толщины). При обработке модельных деревьев в кроне выделялись следующие фракции: ветви (сегменты скелетных осей крон 2 лет и старше), побеги текущего года (включая вершинный побег), листья, генеративные органы, отмершие ветви. Ствол разделяли на секции (0.5–1.0 м в зависимости от возраста деревьев), которые взвешивали индивидуально. Из сегментов ствола выпиливали диски для анализа хода роста по диаметру и отбирали образцы для определения соотношения древесины и коры. Образцы фракций кроны и ствола для определения доли абсолютно сухого вещества высушивались в сушильном шкафу до постоянного веса при температуре 105°.
Данные о фитомассе и годичной продукции модельных деревьев выравнивались методом наименьших квадратов по регрессионным моделям. В качестве независимой переменной использовался диаметр дерева на высоте груди (d1.3). При выравнивании использовались уравнения:
(1)
$P{{h}_{i}}\left( {{{P}_{i}}} \right) = {{\left( {a{\text{ }} + {\text{ }}b{{d}_{{1.3}}}} \right)}^{2}},$(4)
$P{{h}_{i}}\left( {{{P}_{i}}} \right) = \exp \left( {a + b{{{\left( {{{d}_{{1.3}}}} \right)}}^{{--1}}}} \right),$Уравнения, описывающие зависимость массы и годичной продукции деревьев ольхи серой от их диаметра, были выбраны с учетом величины коэффициентов детерминации (R2). Для всех фракций фитомассы и годичной продукции отмечена высокая степень выравнивания эмпирических данных используемыми аллометрическими уравнениями. При аппроксимации массы древесины стволов R2 составил 0.996–0.976; коры стволов – 0.989–0.913; ветвей – 0.961–0.922; побегов – 0.970–0.762; листьев – 0.981–0.814. При аппроксимации годичной продукции древесины стволов R2 равнялся 0.993–0.904; коры стволов – 0.990–0.909; ветвей – 0.985–0.907.
Выровненные значения фитомассы и годичной продукции деревьев по ступеням диаметра умножались на число деревьев этой ступени на пробной площади. Полученные значения надземной фитомассы и годичной продукции древостоя на пробной площади пересчитывались на единицу площади (1 га).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
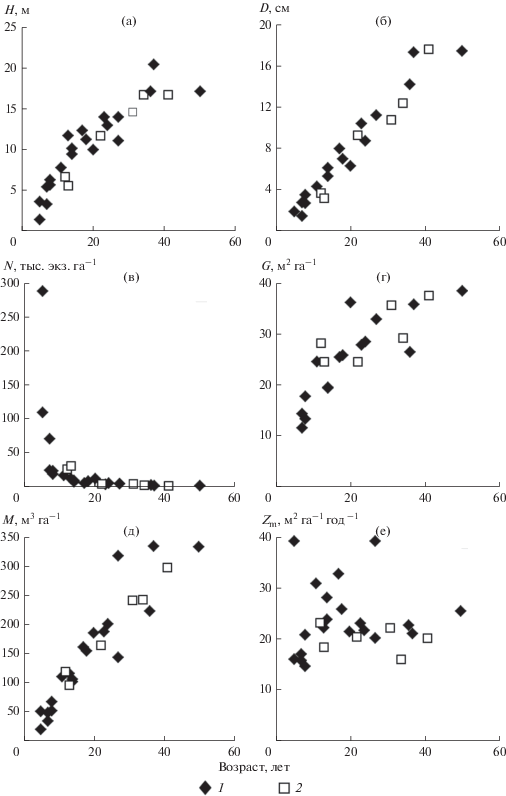
Таксационная характеристика древостоев ольхи серой на пробных площадях приведена в табл. 1. Все они одновозрастные и относятся к одному естественному ряду. Возраст обследованных древостоев (от 12 лет до 41 г.) позволяет проследить динамику таксационных показателей, фитомассы и годичной продукции древостоев гидрофильных сероольшаников от момента завершения формирования ими сомкнутого полога до возраста спелости. Число обследованных объектов обеспечивает получение достаточно полной и объективной картины изменения с возрастом морфоструктурных и биопродукционных показателей древостоев ольхи в этих условиях произрастания и позволяет сравнить их с аналогичными характеристиками древостоев ольхи в условиях неморально-кисличной группы типов леса.
Таблица 1.
Таксационная характеристика древостоев ольхи серой
| № № | Состав | A, лет | H, м | D, см | N, тыс. экз. га–1 | G, м2 га–1 | M, м3 га–1 | ZM, м3 га–1 год–1 |
|---|---|---|---|---|---|---|---|---|
| 1 | 10Олс | 12 | 6.7 | 3.7 | 25.50 | 28.2 | 119 | 10.1 |
| 2 | 10Олс,едБ,Ив | 13 | 5.6 | 3.2 | 30.3 | 24.6 | 96 | 8.1 |
| 3 | 10Олс,едБ,Ив | 22 | 6.3 | 9.3 | 3.66 | 24.6 | 164 | 9.1 |
| 4 | 10Олс | 31 | 14.6 | 10.8 | 3.93 | 35.7 | 241 | 10.1 |
| 5 | 10Олс,едБ,Ив | 34 | 16.7 | 12.4 | 2.43 | 29.2 | 242 | 7.2 |
| 6 | 9Олс1Ос + Б,едИв | 41 | 16.7 | 17.6 | 1.55 | 37.6 | 297 | 9.0 |
Примечание. Состав: Олс – ольха серая, Б – береза, Ив – ива, Ос – осина. Здесь и на рис. 1 A, H, D – средние возраст, высота и диаметр древостоев соответственно; N – число растущих деревьев; G, M – сумма площадей сечений стволов, запас стволовой древесины соответственно; ZM – текущий среднепериодический прирост стволов.
В рассматриваемый возрастной период происходит интенсивное естественное изреживание древостоев: если в 12-летнем сероольшанике густота составляет 25.5 тыс. экз. га–1, то в 41‑летнем она в 16 раз меньше (1.55 тыс. экз. га–1). С увеличением возраста и изреживанием древостоев возрастают их средний диаметр, средняя высота, сумма площадей сечений стволов и запас: так, в 12-летнем древостое они равны 3.7 см, 6.7 м, 28.2 м2 га–1 и 119 м3 га–1, а в 41-летнем – 17.6 см, 16.7 м, 37.6 м2 га–1 и 297 м3 га–1, соответственно. В отличие от этих таксационных показателей текущий среднепериодический прирост не зависит от возраста древостоев и составляет 7.2–10.1 м3 га–1 год–1.
Таксационные показатели древостоев гидрофильных серооольшаников, за исключением прироста по объему, в большинстве случаев близки к аналогичным показателям неморально-кисличных сероольшаников региона (Гульбе, 2012, 2017) (рис. 1). Несколько меньшие по сравнению с неморально-кисличными сероольшаниками значения средней высоты и диаметра в 12- и 13-летнем сероольшаниках компенсируются большими значениями густоты и, как следствие, суммы площадей сечений стволов. В результате эти древостои по запасу стволовой древесины практически не уступают неморально-кисличным сероольшаникам близкого возраста. Величина текущего среднепериодического прироста по объему ольхи серой в этой группе типов леса, как правило, меньше и тяготеет в основном к нижней границе области рассеивания этого показателя в неморально-кисличных сероольшаниках.
Рис. 1.
Возрастная динамика таксационных показателей древостоев неморально-кисличных (1) и гидрофильных (2) сероольшаников: средней высоты (а), среднего диаметра (б), числа стволов (в), суммы площадей сечений стволов (г), запаса (д), среднепериодического прироста стволов (е).
Масса надземной части древостоев ольхи увеличивается с 51 т га–1 в 12-летнем древостое до 141 т га–1 в 41-летнем (табл. 2). Накопление органического вещества происходит преимущественно в стволах и скелете крон деревьев. В течение рассматриваемого периода масса стволов линейно увеличивается с 43 до 121 т га–1; при этом масса древесины стволов возрастает в 2.9 раза (с 36 до 104 т га–1), коры – в 2.4 раза (с 7 до 17 т га–1). Доля древесины в массе стволов несколько возрастает: с 84% в 12-летнем древостое до 86–88% в более старых сероольшаниках. Масса скелета крон увеличивается по слабо выраженной параболической кривой с 4.5 до 17 т га–1, в основном за счет многолетних сегментов скелета крон (ветвей), масса которых возрастает почти в 4 раза (с 3.8 до 16.4 т га–1). Масса побегов текущего года в рассматриваемый период не связана с возрастом и составляет 0.5–0.8 т га–1. В результате доля побегов в скелете крон сокращается с 14–15% в 12–13 лет до 5% в 41-летнем древостое. Масса листьев, как и масса побегов, мало зависит от возраста и варьирует в пределах 2.0–3.6 т га–1. Типичной для древостоев ольхи серой в этих условиях следует, видимо, считать массу листьев 2.9–3.6 т га–1. Более низкие значения этого показатели (2.0 т га–1 в 34-летнем сероольшанике и 2.4 т га–1 в 22-летнем) связаны, видимо, с погодными условиями в год наблюдений.
Таблица 2.
Фитомасса надземной части древостоев гидрофильных сероольшаников
| № № | Возраст, лет |
Масса | Всего | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| стволов | крон | древес- ных фракций |
||||||||||
| древесины | коры | итого | ветвей | побегов текущего года |
скелета крон |
листьев | генера- тивных органов |
итого | ||||
| 1 | 12 | $\frac{{36.27}}{{71.5}}$ | $\frac{{6.97}}{{13.7}}$ | $\frac{{43.24}}{{85.2}}$ | $\frac{{3.85}}{{7.6}}$ | $\frac{{0.65}}{{1.3}}$ | $\frac{{4.5}}{{8.9}}$ | $\frac{{2.97}}{{5.9}}$ | $\frac{0}{0}$ | $\frac{{7.47}}{{14.8}}$ | $\frac{{47.74}}{{94.1}}$ | $\frac{{50.71}}{{100.0}}$ |
| 2 | 13 | $\frac{{28.34}}{{66.3}}$ | $\frac{{7.06}}{{16.5}}$ | $\frac{{35.4}}{{82.8}}$ | $\frac{{3.79}}{{8.9}}$ | $\frac{{0.66}}{{1.6}}$ | $\frac{{4.45}}{{10.5}}$ | $\frac{{2.88}}{{6.7}}$ | $\frac{0}{0}$ | $\frac{{7.33}}{{17.2}}$ | $\frac{{39.85}}{{93.3}}$ | $\frac{{42.73}}{{100.0}}$ |
| 3 | 22 | $\frac{{56.12}}{{76.5}}$ | $\frac{{7.68}}{{10.5}}$ | $\frac{{63.80}}{{87.0}}$ | $\frac{{6.17}}{{8.4}}$ | $\frac{{0.49}}{{0.7}}$ | $\frac{{6.66}}{{9.1}}$ | $\frac{{2.41}}{{3.3}}$ | $\frac{{0.45}}{{0.6}}$ | $\frac{{9.52}}{{13.0}}$ | $\frac{{70.46}}{{96.1}}$ | $\frac{{73.32}}{{100.0}}$ |
| 4 | 31 | $\frac{{83.16}}{{76.1}}$ | $\frac{{11.57}}{{10.6}}$ | $\frac{{94.73}}{{86.7}}$ | $\frac{{10.65}}{{9.7}}$ | $\frac{{0.79}}{{0.7}}$ | $\frac{{11.44}}{{10.4}}$ | $\frac{{3.01}}{{2.8}}$ | $\frac{{0.06}}{{0.1}}$ | $\frac{{14.51}}{{13.3}}$ | $\frac{{106.17}}{{97.1}}$ | $\frac{{109.24}}{{100.0}}$ |
| 5 | 34 | $\frac{{83.39}}{{78.4}}$ | $\frac{{11.55}}{{10.9}}$ | $\frac{{94.94}}{{89.3}}$ | $\frac{{8.82}}{{8.3}}$ | $\frac{{0.50}}{{0.5}}$ | $\frac{{9.32}}{{8.8}}$ | $\frac{{1.96}}{{1.8}}$ | $\frac{{0.12}}{{0.1}}$ | $\frac{{11.4}}{{10.7}}$ | $\frac{{104.26}}{{98.1}}$ | $\frac{{106.34}}{{100.0}}$ |
| 6 | 41 | $\frac{{103.81}}{{73.4}}$ | $\frac{{16.72}}{{11.8}}$ | $\frac{{120.53}}{{85.2}}$ | $\frac{{16.37}}{{11.5}}$ | $\frac{{0.78}}{{0.6}}$ | $\frac{{17.15}}{{12.1}}$ | $\frac{{3.64}}{{2.6}}$ | $\frac{{0.14}}{{0.1}}$ | $\frac{{20.93}}{{14.8}}$ | $\frac{{137.68}}{{97.3}}$ | $\frac{{141.46}}{{100.0}}$ |
Сравнительный анализ показал, что масса стволов, скелета крон и древесных фракций ольхи в гидрофильных серольшаниках практически не отличается от массы этих фракций в неморально-кисличных сероольшаниках близкого возраста, где в возрастном интервале от 10 до 40 лет масса стволов увеличивается в среднем примерно с 40 до 125, скелета крон − с 3.5–4.0 до 15.5, древесных фракций − с 43.5–44.0 до 140.5 т га–1 (Гульбе, 2012, 2017) (рис. 2). Масса листьев в гидрофильных сероольшаниках несколько больше, чем в неморально-кисличных, где она в эти годы также мало зависит от возраста и составляет в среднем 2.2 т га–1. Тем не менее, различия в массе листьев не сказываются на надземной фитомассе древостоев, которая, как и в гидрофильных сероольшаниках, возрастает с 45–49 до 143 т га–1.
Рис. 2.
Возрастная динамика фитомассы древостоев неморально-кисличных (1) и гидрофильных (2) сероольшаников: а – стволов (Phstw), б – скелета крон (Phsk), в – листьев (Phl), г – общей надземной (Phnads).
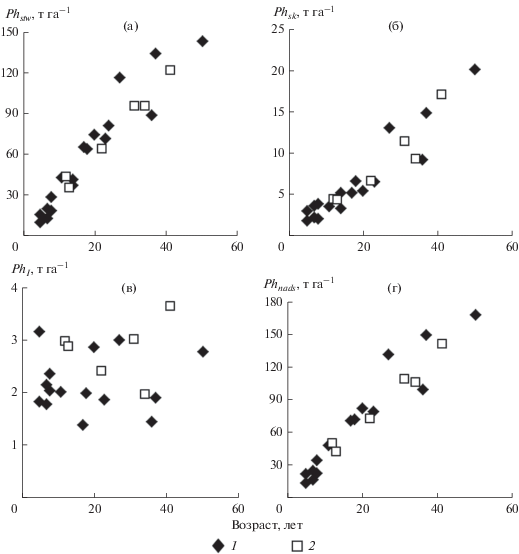
В гидрофильных сероольшаниках в результате аккумуляции органического вещества преимущественно в стволах деревьев эта фракция абсолютно преобладает в фитомассе надземной части древостоев, составляя в большинстве случаев 85–89%; лишь в 13-летнем древостое она несколько меньше (83%) (табл. 2). Доля скелета крон в надземной фитомассе также относительно постоянна (8–12%), хотя некоторое увеличение представительства скелета крон в фитомассе надземной части 41-летнего древостоя не исключает того, что в дальнейшем эта тенденция сохранится. Доля древесных фракций в 12-летнем древостое составляет 94%, в древостоях 22 лет и старше она несколько выше – 96–98%. Доля массы листьев в надземной фитомассе с возрастом, напротив, снижается с 6–7% в 12- и 13-летнем древостоях до 2–3% в более старых сообществах. Вклад генеративных органов в фитомассу незначителен (0.6% и менее).
Распределение надземной фитомассы по фракциям в древостоях гидрофильных сероольшаников близко к ее распределению в неморально-кисличных сероольшаниках, где в возрастном интервале от 10 до 40 лет стволы составляют в среднем 89% (в т. ч. доля древесины около 79%, коры – 10%), скелет крон – 9% и в целом древесные фракций – 98% (Гульбе, 1988) (рис. 2).
В рассматриваемом возрастном интервале годичная продукция надземной части древостоев гидрофильных сероольшаников варьирует в пределах 6.7–9.8 т га–1 год–1, в т. ч. годичная продукция стволов − 3.2–4.5 т га–1 год–1, скелета крон – 1.4–2.1 т га–1 год–1, листьев – 2.0–3.6 т га–1 год–1, и мало зависит от возраста (табл. 3). Уменьшение надземной годичной продукции 34-летнего древостоя до 6.7 т га–1 год–1, а также слагающих ее годичной продукции стволов и листьев до 3.2 т га–1 год–1 и 2.0 т га–1 год–1, соответственно, связано, видимо, с особенностями погодных условий в год наблюдений. Типичными для гидрофильных сероольшаников можно считать следующие значения годичной продукции фракций: стволы – 4 т га–1 год–1, скелет крон – 1.7, листья – 2.8, надземная часть в целом – 8.5 т га–1 год–1. Как следствие в рассматриваемый период относительно стабильно и распределение годичной продукции надземной части древостоев по фракциям. Доля стволов в годичной продукции надземной части древостоя составляет 47.0%, скелета крон – 19.7, листьев – 33.3%.
Таблица 3.
Годичная продукция надземной части древостоев гидрофильных сероольшаников
| № № | Возраст, лет |
Годичная продукция | Всего | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| стволов | крон | древесных фракций |
||||||||||
| древесины | коры | итого | ветвей | побегов текущего года |
скелета крон |
листьев | генера- тивных органов |
итого | ||||
| 1 | 12 | $\frac{{3.59}}{{39.7}}$ | $\frac{{0.75}}{{8.3}}$ | $\frac{{4.34}}{{48.0}}$ | $\frac{{1.08}}{{11.9}}$ | $\frac{{0.65}}{{7.2}}$ | $\frac{{1.73}}{{19.1}}$ | $\frac{{2.97}}{{32.9}}$ | $\frac{0}{0}$ | $\frac{{4.70}}{{52.0}}$ | $\frac{{6.07}}{{67.1}}$ | $\frac{{9.04}}{{100.0}}$ |
| 2 | 13 | $\frac{{2.78}}{{34.9}}$ | $\frac{{0.74}}{{9.3}}$ | $\frac{{3.52}}{{44.2}}$ | $\frac{{0.9}}{{11.3}}$ | $\frac{{0.66}}{{8.3}}$ | $\frac{{1.56}}{{19.6}}$ | $\frac{{2.88}}{{36.2}}$ | $\frac{0}{0}$ | $\frac{{4.44}}{{55.8}}$ | $\frac{{5.08}}{{63.8}}$ | $\frac{{7.96}}{{100.0}}$ |
| 3 | 22 | $\frac{{3.49}}{{42.5}}$ | $\frac{{0.50}}{{6.1}}$ | $\frac{{3.99}}{{48.6}}$ | $\frac{{0.87}}{{10.6}}$ | $\frac{{0.49}}{{6.0}}$ | $\frac{{1.36}}{{16.6}}$ | $\frac{{2.41}}{{29.3}}$ | $\frac{{0.45}}{{5.5}}$ | $\frac{{4.22}}{{51.4}}$ | $\frac{{5.35}}{{65.2}}$ | $\frac{{8.21}}{{100.0}}$ |
| 4 | 31 | $\frac{{3.90}}{{40.5}}$ | $\frac{{0.57}}{{5.9}}$ | $\frac{{4.47}}{{46.4}}$ | $\frac{{1.3}}{{13.5}}$ | $\frac{{0.79}}{{8.20}}$ | $\frac{{2.09}}{{21.7}}$ | $\frac{{3.01}}{{31.3}}$ | $\frac{{0.06}}{{0.6}}$ | $\frac{{5.16}}{{53.6}}$ | $\frac{{6.56}}{{68.1}}$ | $\frac{{9.63}}{{100.0}}$ |
| 5 | 34 | $\frac{{2.79}}{{41.9}}$ | $\frac{{0.42}}{{6.3}}$ | $\frac{{3.21}}{{48.2}}$ | $\frac{{0.87}}{{13.1}}$ | $\frac{{0.50}}{{7.5}}$ | $\frac{{1.37}}{{20.6}}$ | $\frac{{1.96}}{{29.4}}$ | $\frac{{0.12}}{{1.8}}$ | $\frac{{3.45}}{{51.8}}$ | $\frac{{4.58}}{{68.8}}$ | $\frac{{6.66}}{{100.0}}$ |
| 6 | 41 | $\frac{{3.54}}{{36.2}}$ | $\frac{{0.58}}{{5.9}}$ | $\frac{{4.12}}{{42.1}}$ | $\frac{{1.1}}{{11.2}}$ | $\frac{{0.78}}{{8.0}}$ | $\frac{{1.88}}{{19.2}}$ | $\frac{{3.64}}{{37.3}}$ | $\frac{{0.14}}{{1.4}}$ | $\frac{{5.66}}{{57.9}}$ | $\frac{{6.00}}{{61.3}}$ | $\frac{{9.78}}{{100.0}}$ |
Из известных определений другими авторами биологической продуктивности древостоев ольхи серой на территории Скандинавии и Восточной Европы в сходных с нашими почвенно-климатических условиях для сравнения могут быть использованы данные о фитомассе и продукции 14- и 40-летнего приручейных сероольшаников в Эстонии (Lõhmus et al., 1996, цит. по: Aosaar et al., 2012). Сероольшаник 40 лет характеризуется близкими значениями надземной фитомассы древостоя (131.8 т га–1), массы стволов (104.7 т га–1) и годичной продукции стволов (4.2 т га–1 год–1) с 41-летним гидрофильным серольшаником, а сероольшаник 14 лет существенно превосходит близкие по возрасту гидрофильные сероольшаники (93.1 т га–1; 81.7 т га–1 и 8.3 т га–1 год–1, соответственно). Можно предположить, что в данном случае учет только орографических факторов при типологической классификации фитоценозов недостаточен, и в одну группу “приручейные” попали сообщества с различной интенсивностью обменных процессов, а по величине фитомассы и годичной продукции молодой приручейный сероольшаник должен быть отнесен к более продуктивному типу леса.
В ходе роста и развития гидрофильных сероольшаников в возрастном интервале с 12 лет до 41 г. при четко выраженной возрастной динамике большинства дендрометрических показателей и фитомассы древостоев их годичная продукция и ее фракционная структура остаются практически неизменными. Аналогичная закономерность ранее была выявлена при изучении роста и развития древостоев березы, осины и ольхи серой, произрастающих в условиях неморально-кисличной группы типов леса. Период в развитии древостоев, когда значения годичной продукции (P) после достижения максимальных значений выходят на плато и сохраняют этот уровень в течение относительно продолжительного времени с некоторыми колебаниями конкретных значений P в сравнительно небольшом интервале, получил название инвариантного (Рождественский, Уткин, 1986; Рождественский и др., 1988; Уткин и др., 1988). Наблюдаемые колебания годичной продукции связаны, прежде всего, с погодными условиями отдельных лет, вызывающими изменения в структуре и функционировании фитоценозов.
Из-за возможных различий в продолжительности онтоценогенеза, сроках и временных рамках кульминации годичной продукции сравнительную оценку продукционного потенциала древостоев различных пород, произрастающих в различных экологических условиях, следует проводить в период кульминации годичной продукции. При ограниченном количестве детальной информации о биологической продуктивности лесных фитоценозов (вследствие большой трудоемкости ее получения) при решении подобных задач инвариантный период, когда годичная продукция максимальна или близка к ней, может быть использован в качестве базового. Такой подход ранее был реализован при сравнительном анализе продукционного процесса в древостоях разных пород: березняках, осинниках и сероольшаниках, − произрастающих в сходных условиях неморально-кисличной группы типов леса (Рождественский и др., 1988; Уткин и др., 1988).
Наличие инвариантного периода в развитии гидрофильных серооольшаников позволяет сравнить продукционную способность древостоев одной породы, а именно ольхи серой, произрастающих в разных экологических условиях, для решения вопроса о равноценности последних в отношении этой породы. В этот период годичная продукция надземной части древостоев гидрофильных сероольшаников в большинстве случаев несколько ниже величины этого показателя в неморально-кисличных сероольшаниках (10–12 т га–1 год–1) (рис. 3). Снижение уровня надземной годичной продукции происходит за счет уменьшения в гидрофильных сероольшаниках годичной продукции стволов по сравнению с неморально-кисличными, где она составляет 4.5–7.5 т га–1 год–1. Годичная продукция скелета крон древостоев в обоих типах сероольшаников довольно близка, за исключением нескольких наиболее продуктивных древостоев, где неморально-кисличные древостои имеют преимущество. Масса листьев в гидрофильных сероольшаниках зачастую соответствует максимальным значениям этого показателя в неморально-кисличных сероольшаниках, где она варьирует в пределах 1.7−3.4 т га–1.
Рис. 3.
Возрастная динамика годичной продукции древостоев неморально-кисличных (1) и гидрофильных (2) сероольшаников: а – стволов (Pstw), б – скелета крон (Psk), в – общей надземной (Pnads) и г – коэффициента продуктивности ассимиляционного аппарата (Pnads$Ph_{l}^{{ - 1}}$).
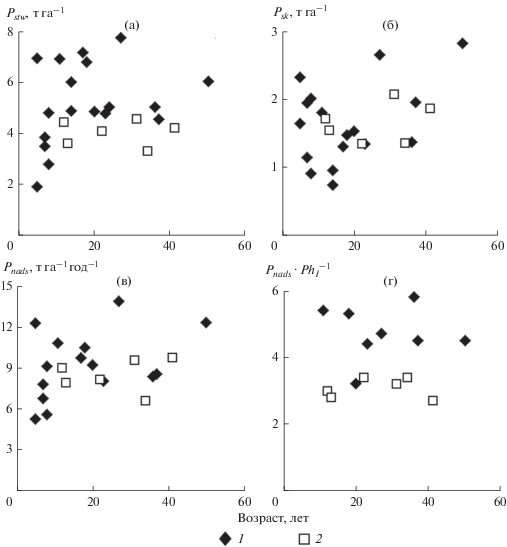
В гидрофильных сероольшаниках, как и в неморально-кисличных, понятие продукционной инвариантности распространяется и на фракционную структуру годичной продукции надземной части древостоя, выраженную в относительных единицах, но в данном случае представительство отдельных фракций в годичной продукции иное. Если в неморально-кисличных сероольшаниках стволы, скелет крон и листья составляли примерно 55, 19 и 26% от годичной продукции древостоя соответственно (Гульбе, 1988), в годичной продукции гидрофильных сероольшаников примерно на 8% меньше доля стволов и почти на эту же величину (7%) больше доля листьев. Доля скелета крон остается примерно на том же уровне.
Таким образом, при уменьшении величины надземной годичной продукции древостоев с увеличением увлажнения почвы происходит сокращение в ее составе массы и представительства фракции стволов, в то время как годичная продукция и доли скелета крон остаются на том же уровне, а годичная продукция и доля листьев даже несколько возрастают. Динамически равновесное состояние между потреблением древостоем питательных веществ и наличием доступных ресурсов среды на более низком уровне в условиях гидрофильной группы типов достигается перераспределением годичной продукции по фракциям. Аналогичная картина наблюдается в сосняках, произрастающих в разных условиях увлажнения, где “при меньшей величине годичной продукции деревьев на болоте она направляется преимущественно на формирование ассимиляционного аппарата и регенерацию ежегодного повышенного отпада корней, а не депонируется в стволовой древесине, как это имеет место в высокобонитетных суходольных древостоях” (Вомперский, 1991). Получает подтверждение мнение, что ствол является вместилищем “избытка” ассимилятов и при ухудшении условий для роста древесных растений и сокращении объема продуцированного вещества происходит уменьшение доли пластических веществ, идущих на формирование стволов (Waring, 1980). Я.К. Палуметс (1988) пришел к такому же выводу на основании анализа распределения фитомассы по фракциям в древостоях ели, где доля стволов, как правило, уменьшалась по мере ухудшения условий произрастания.
При использовании характеристик фитомассы и ее структуры в качестве критерия степени реализации продукционного потенциала ольхи серой в разных условиях произрастания гидрофильная и неморально-кисличная группы типов леса являются для нее равноценными. Если же рассматривать параметры и структуру не аккумулированного органического вещества (надземная фитомасса древостоя и ее структура), а синтезированного в течение вегетационного периода (надземная годичная продукции и ее структура), то обнаруживается преимущество неморально-кисличных сероольшаников, в которых величина надземной годичной продукции и годичной продукции стволов существенно выше. Неоднозначность полученных результатов при использовании этих критериев при оценке степени реализации продукционного потенциала древесной породы в различных условиях произрастания говорит о необходимости анализа как интегральной характеристики биологической продуктивности (фитомасса), так и ее текущего показателя в течение вегетационного периода (годичная продукция). Можно предположить, что для оценки реализации продукционного потенциала ольхи в различных условиях произрастания текущие показатели продукционного процесса характеризуются большей чувствительностью по сравнению с накопленными.
Близкие значения массы стволов и надземной фитомассы древостоев в сравниваемых типах сероольшаников при существенных различиях в годичной продукции этих фракций позволяют сделать предположение о меньшей интенсивности процессов отпада в гидрофильных сероольшаниках, для подтверждения которого необходимы дальнейшие исследования.
Коэффициент продуктивности (или эффективности работы) ассимиляционного аппарата (Уткин, 1975), равный отношению годичной продукции надземной части древостоя к массе листьев, в гидрофильных сероольшаниках в инвариантный период относительно постоянен и составляет в среднем 3.08 ± 0.12 т т–1 (σ = 0.30; n = 6). В неморально-кисличных сероольшаниках в интервале от 10 до 37 лет его величина также не зависит от возраста и равняется 4.77 ± 0.32 т т–1 (σ = = 0.86; n = 7), что на 1.69 т т–1 больше. Статистическая обработка данных показала, что гидрофильные и неморально-кисличные сероольшаники достоверно (доверительный уровень 95%) различаются между собой по коэффициенту продуктивности ассимилирующих органов. Полученные результаты свидетельствуют о перспективности использования коэффициента продуктивности ассимилирующих органов для оценки влияния условий произрастания на продуктивность различных древесных пород и необходимости дальнейших исследований в этом направлении.
Продукционный потенциал древесной породы и степень его реализации помимо величины и структуры годичной продукции в инвариантный период характеризуются его продолжительностью, возрастом древостоя в момент начала и окончания. По имеющимся данным временные параметры инвариантного периода в рассматриваемых типах сероольшаников довольно близки. Однако в полной мере сопоставить продукционный потенциал древостоев ольхи и степень его реализации в различных экологических условиях с этих позиций на данном этапе не представляется возможным, так как не установлены точный возраст древостоя в момент начала и окончания инвариантного периода и его продолжительность. Это связано, прежде всего, с отсутствием данных о биологической продуктивности гидрофильных сероольшаников на начальных стадиях их роста и развития, а также в возрасте старше 41 г. Кроме того, временные рамки инвариантного периода могут быть определены с известной долей условности, так как его начало и окончание могут варьировать вследствие индивидуальных особенностей роста и развития древостоя.
Заключение. Гидрофильные (таволговые) сероольшаники по запасу древесины и надземной фитомассе древостоев сопоставимы с неморально-кисличными сероольшаниками и относятся к высокопроизводительным лесным экосистемам. В ходе их роста и развития, как и в развитии насаждений мягколиственных пород неморально-кисличной группы типов леса, выявлен инвариантный период, в течение которого продукционная инвариантность проявляется, по крайней мере, в двух формах: по величине годичной продукции и по ее фракционному составу.
Наличие инвариантного периода в развитии гидрофильных серооольшаников расширяет понятие продукционной инвариантности и позволяет рассмотреть в этом аспекте древостои ольхи серой, занимающие различные местообитания, для решения вопроса об их продукционной равноценности в отношении данной породы. Установлено, что в указанный период древостои гидрофильных сероольшаников, почвы которых характеризуются повышенным увлажнением, уступают неморально-кисличным по величине годичной продукции надземной части древостоев и стволов, характеризуются меньшей величиной коэффициента продуктивности ассимилирующих органов по сравнению с неморально-кисличными. Таким образом, продукционный потенциал ольхи серой в условиях гидрофильной группы типов леса реализуется в меньшей степени. Полученные результаты могут быть использованы при решении целого ряда задач теоретического и прикладного характера: оценка сырьевого значения и экологической роли древостоев, бонитировка местообитаний, оценка продукционного разнообразия лесных биогеоценозов.
Список литературы
Борисова М.А., Богачев В.В., Маракаев О.А. Лесные формации западного флористического района Ярославской области // Известия Самарского НЦ РАН. 2012. Т. 14. № 1(4). С. 974–977.
Вомперский С.Э. Лес и болото: особенности круговорота веществ и проявления биосферной роли // Лесоведение. 1991. № 6. С. 54–64.
Гульбе Я.И. Фракционная структура фитомассы и годичной продукции древостоев и деревьев серой ольхи // Анализ продукционной структуры древостоев. М.: Наука, 1988. С. 85–103.
Гульбе Я.И. Динамика биологической продуктивности южнотаежных древостоев ольхи серой (на примере Ярославской области): Дис. … канд. биол. наук (03.00.16). М., 2012. 167 с.
Гульбе Я.И. Динамика биологической продуктивности южнотаежных древостоев ольхи серой // Комплексные стационарные исследования в лесах южной тайги (памяти М.В. Рубцова). М.: Товарищество научных изданий КМК, 2017. С. 99–124.
Ермолова Л.С., Гульбе Т.А., Гульбе Я.И. Фитоценотическая структура гидрофильных сероольшаников в южной тайге // Современная ботаника в России. Труды XIII съезда Русского ботанического общества и конференции “Научные основы охраны и рационального использования растительного покрова Волжского бассейна” (Тольятти 16–22 сентября 2013). Т. 2: Систематика и география сосудистых растений. Сравнительная флористика. Геоботаника. Тольятти: Кассандра, 2013. С. 211–212.
Лесной план Ярославской области 2011 г. // Сайт Департамента лесного хозяйства Ярославской области. URL: http://www.yarregion.ru/depts/dlh/docsDocuments/ Л_%20п_Яр_обл_22_03_2011.rar (дата обращения: 10.12.2015)
Осипов В.В., Соколов А.А., Шахова О.В. Территориальное размещение типов леса в связи с рельефом и гидрографической сетью // Лесоводственные исследования в подзоне южной тайги. М.: Наука, 1977. С. 86–92.
Палуметс Я.К. Распределение фракций фитомассы ели европейской в зависимости от возраста и климатических факторов // Лесоведение. 1988. № 2. С. 34–40.
Рождественский С.Г., Уткин А.И. О продукционной инвариантности растительного покрова // Вертикально-фракционное распределение фитомассы в лесах. М.: Наука, 1986. С. 207–215.
Рождественский С.Г., Уткин А.И., Гульбе Я.И., Каплина Н.Ф. Древостои мелколиственных пород как инвариантные продукционные системы // Анализ продукционной структуры древостоев. М.: Наука, 1988. С. 214–224.
Усольцев В.А. Фитомасса и первичная продукция лесов Евразии. Екатеринбург: УрО РАН, 2010. 570 с.
Уткин А.И. О показателях лесных биогеоценозов // Бюл. МОИП. Отд. биол. 1975. Т. 80. № 2. С. 95–107.
Уткин А.И., Гульбе Я.И., Ермолова Л.С. Первичная продуктивность сероольшаников в Ярославской области // Лесоведение. 1980. № 3. С. 69–80.
Уткин А.И., Гульбе Я.И., Ермолова Л.С. Сероольшаники Большесельского района Ярославской области и их первичная биологическая продуктивность // Биологическая продуктивность лесов Поволжья. М.: Наука, 1982. С. 110–142.
Уткин А.И., Рождественский С.Г., Гульбе Я.И., Каплина Н.Ф. Продукционная инвариантность древостоев // Лесоведение. 1988. № 2. С. 12–23.
Юркевич И.Д., Гельтман В.С., Парфенов В.И. Сероольховые леса и их хозяйственное использование. Минск: Изд-во Академии наук БССР, 1963. 142 с.
Ярмишко В.Т. Конкурентные взаимовлияния ели и лиственных пород в зоне корневых систем // Лесное хоз-во, лесная, деревообрабатывающая и целлюлозно-бумажная промышленность. 1976. Вып. 4. С. 51–53.
Aosaar J., Vaik M., Uri V. Biomass production potential of grey alder (Alnus incana (L.) Moench.) in Scandinavia and Eastern Europe: A review // Biomass & Bioenergy. 2012. V. 45. P. 11–26.
Lõhmus K., Mander Ü., Tullus H., Keedus K. Productivity, buffering capacity and resources of grey alder forests in Estonia // Short Rotation Willow Coppice for Renewable Energy and Improved Environment. Uppsala: Swedish University of Agricultural Sciences, 1996. P. 95–105.
Mander Ü., Kuusemets V., Ivask M. Nutrient dynamics of riparian ecotones: a case study from the Porijõgi River catchment, Estonia // Landscape & Urban Planning. 1995. V. 31. P. 333–348.
Mander Ü., Kuusemets V., Lõhmus K., Mauring T. Efficiency and dimensioning of riparian buffer zones in agricultural catchments // Ecological Engineering. 1997. V. 8. P. 299–324.
Waring R.H. Site, leaf area, and phytomass production of trees // In: Benecke, U. and Davis, M.R. (eds). Mountain Environments and Subalpine Tree Growth. New Zealand Forest Service. Technical Paper 70. Wellington, New Zealand, 1980. P. 125–135.
Дополнительные материалы отсутствуют.