

Лесоведение, 2020, № 6, стр. 537-547
Структурно–функциональные изменения хвои сосны в условиях аэротехногенного загрязнения
В. В. Тужилкина a, *, С. Н. Плюснина a
a Институт биологии Коми научного центра Уральского отделения РАН
167982 Сыктывкар, ул. Коммунистическая, 28, ГСП-2, Россия
* E-mail: tuzhilkina@ib.komisc.ru
Поступила в редакцию 26.10.2018
После доработки 15.11.2019
Принята к публикации 06.06.2020
Аннотация
Исследовано влияние аэротехногенных выбросов целлюлозно-бумажного производства АО “Монди Сыктывкарский ЛПК” на состояние хвои сосны обыкновенной (Pinus sylvestris) в сосняках лишайниковых. Проведен сравнительный анализ структурно-физиологических показателей фотосинтетического аппарата сосны в фоновом районе и на загрязненной территории в зависимости от техногенной нагрузки в 1995 и 2015 гг. Показано, что сильные воздействия промышленных выбросов приводят к уменьшению накопления крахмала в хлоропластах и подавлению синтеза хлорофиллов и каротиноидов в хвое сосны. При ослаблении техногенной нагрузки загрязнение воздуха сернистыми и азотистыми соединениями не оказывает отрицательного влияния на ультраструктурную организацию клеток мезофилла, пигментный комплекс и фотосинтетическую способность хвои.
Известно, что аэротехногенное загрязнение окружающей среды является значимым фактором, влияющим на функционирование лесных экосистем на территории Европы. Целлюлозно-бумажное производство – одна из наиболее загрязняющих природную среду отраслей промышленности. Предприятия этого профиля при производительности 2000 т целлюлозы в сутки ежегодно выбрасывают в атмосферу около 40000 т SO2 (Сергейчик, 1984). В состав атмосферных выбросов комбинатов целлюлозно-бумажного производства входят различные серо- и азотсодержащие соединения, в т.ч. оксиды серы, метилмеркаптан, сероводород, диметилсульфид, оксиды азота и др. (Личутина и др., 2011).
Изучение влияния техногенного загрязнения на хвойные фитоценозы было проведено как за рубежом (Soikkeli, 1981; Huttunen, Laine, 1983), так и во многих промышленных регионах России: в Прибайкалье (Рунова, 1999), Архангельской области (Тарханов и др., 2004), Карелии (Дьяконов, 1996) и Республике Коми (Тужилкина и др., 1998; Торлопова, Робакидзе, 2003).
Мониторинговые исследования, охватывающие значительный временной промежуток деятельности промышленных предприятий, позволяют проследить процессы адаптации живых компонентов экосистемы к действию поллютантов. Связанное со снижением выбросов уменьшение токсической нагрузки на сообщества предоставляет возможность изучать процессы их естественного восстановления (Воробейчик и др., 2014), которые до настоящего времени недостаточно изучены. В отечественной литературе в основном представлены результаты мониторинговых исследований в зоне действия металлургических предприятий (Ярмишко и др., 2011, 2017; Сухарева, Лукина, 2014; Михайлова, 2017).
На севере России вокруг одного из крупнейших в Европе целлюлозно-бумажного предприятия АО “Монди Сыктывкарский лесопромышленный комплекс” (СЛПК) мониторинг лесов непрерывно проводится с 1994 г. (Торлопова, Робакидзе, 2003; Робакидзе, Торлопова, 2018; Тужилкина, Плюснина, 2014). СЛПК, наряду с Архангельским целлюлозно-бумажным комбинатом и Институтом экологических проблем Севера УрО РАН, одним из первых в Российской Федерации начал проводить анализ природоохранной деятельности по системе технологического нормирования. Это требовало перехода производства на передовые технологии, гарантирующие минимальное воздействие на экосистемы (Личутина, 2011). За период наблюдений объем газообразных выбросов сократился в 2–3 раза.
Сохранение жизнеспособности органов и растения в целом обеспечивается многообразными адаптационными механизмами, существующими на всех уровнях организации биологических систем. Независимо от того, какую устойчивость к повреждающему действию газообразных веществ проявляет растение – регенерационную, популяционную, габитуальную, физиолого-биохимическую – в конечном итоге о ней судят по состоянию основного ассимиляционного органа – листа (Кулагин, 1985; Гетко, 1989). Поэтому при проведении биомониторинга особое внимание следует уделять ассимиляционному аппарату древесных растений (Барахтенова, 1987).
Установлено, что повреждение в первую очередь проявляется на физиолого-биохимическом уровне, затем распространяется на ультраструктурный и клеточный, и лишь после этого развиваются видимые признаки повреждения – хлорозы и некрозы тканей листа, опадение листьев, торможение роста (Мальхотра, Хан, 1988). В связи с этим необходимы исследования структуры и функции ассимиляционных органов, наиболее чувствительных к экологическим изменениям среды (Кулагин, 1985; Гетко, 1989; Kukkola et al., 1997). Хвойные растения вследствие многолетнего существования фотосинтезирующих органов более подвержены негативному воздействию воздушных поллютантов, чем лиственные породы. При этом сосна лидирует в ряду хвойных пород по чувствительности к загрязнению (Антипов, 1979) и значительно повреждается в лишайниковых типах леса (Торлопова, Робакидзе, 2003).
Цель данной работы – сравнительная оценка структурно-функциональной организации хвои сосны в условиях аэротехногенного загрязнения выбросами целлюлозно-бумажного производства в 1995 и 2015 г.
ОБЪЕКТЫ И МЕТОДИКА
Исследования проводили в 1995 и 2015 г. в сосновых фитоценозах лишайникового типа в подзоне средней тайги на пробных площадях, заложенных для проведения локального мониторинга состояния лесов в зоне аэротехногенного воздействия СЛПК. Предприятие производит сульфатную целлюлозу и беленую химико-термомеханическую массу с полной переработкой на бумагу и картон (Личутина, 2011). В 90-ые годы прошлого столетия основными выбросами предприятия являлись оксиды азота (2300–3100 т · год–1) и углерода (17520–23150 т · год–1), сернистый ангидрид (379.0–765.6 т · год–1), неорганическая пыль (1700–2400 т · год–1). Примеси в виде хлора и двуокиси хлора составляли в сумме 3.7 т · год–1. В этот период СЛПК выбрасывал в атмосферу 24 983–30 708 т · год–1 пылевых и газообразных веществ (Государственный …, 1997). За последние годы отмечена тенденция снижения фоновых концентраций сернистого ангидрида и сероводорода на территории г. Сыктывкар в связи с модернизацией очистных сооружений на СЛПК. В 2006 г. на предприятии внедрена система бесхлорной отбелки целлюлозы. В настоящее время суммарное количество выбросов колеблется от 10 до 17 тыс. т · год–1, что в 2–3 раза ниже, чем в 1994–1996 гг. (Государственный …, 2016).
Экспериментальные участки располагались на расстоянии 7.3 и 11 км от СЛПК по вектору господствующих ветров летом (в северном и восточном направлениях) в лишайниковых типах леса. Фоновый участок находился в 56 км к северу от производства на Ляльском лесоэкологическом стационаре Института биологии Коми НЦ УрО РАН (табл. 1).
Таблица 1.
Характеристика древостоев экспериментальных и фонового участков
| Расстояние от СЛПК, км |
Состав древостоя |
Возраст, лет |
Средний диаметр, см | Средняя высота, м | Бонитет |
|---|---|---|---|---|---|
| 7.3 | 10С | 107–127 | 20 | 17 | IV |
| 11.0 | 10С | 77–127 | 20 | 19 | IV |
| 56.0 | 10С | 70–125 | 30 | 18 | IV |
Для анатомо-физиологических исследований отбирали образцы сформированной однолетней хвои с 5–10 80–90-летних деревьев сосны с опытных и фонового участков. Побеги срезали с середины кроны с южной стороны дерева. Сбор образцов проводили в июле. Анатомическую структуру изучали на временных препаратах поперечных срезов из средней трети хвои под микроскопом Axiovert 200 M (Carl Zeiss, Германия).
Электронно-микроскопические исследования. Фрагменты из центральной части хвои фиксировали в течение 4.5 ч в 2.5%-ом глутаральдегиде, приготовленном на фосфатном буфере с рН 7.4. Постфиксацию проводили в 1%-ом водном растворе осмиевой кислоты в течение 4 ч. После дегидратации в сериях растворов этилового спирта и ацетона образцы заключали в смолу Эпон-812 (FLUKA). Ультратонкие срезы были получены на ультрамикротоме PowerTome PC (Boeckeler Instruments, США). Наблюдения и съемку микропрепаратов проводили на электронном микроскопе Tesla BS 500 (Tesla, Чехословакия). Количественные характеристики клеток мезофилла и клеточных органелл получали по методике (Скупченко, 1990) в 20–60-кратной повторности.
Физиологические исследования. Для количественного и качественного анализа пигментов образцы хвои отбирали в пяти-шести биологических и четырех-пятикратной аналитических повторностях. Содержание пластидных пигментов определяли на спектрофотометре UV-1800 (Shimadzu, Япония) в ацетоновой вытяжке (Шлык, 1971). Фотосинтетическую способность оценивали по поглощению СО2, измеренному инфракрасным газоанализатором Li COR-6400 (Li COR, США) при насыщающей освещенности, температуре 20°С и влажности 70%. Темновое дыхание измеряли по выделению СО2 инфракрасным газоанализатором при 20°С, влажности 70%. Измерения фотосинтетической активности и дыхательной способности хвои проводили в контролируемой камере 6400-22L (Li COR, США).
Статистический анализ экспериментальных данных. Данные обрабатывали методами вариационной статистики с использованием Microsoft Office Excel 2007 и Statistica. Значимость отличий исследуемых показателей от контроля проверяли с помощью критерия Стъюдента.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Морфология и анатомия хвои. Форма поперечного сечения хвои сосны близка к полукругу или полуовалу. Его площадь зависит от размеров и числа гистологических элементов. Значительный объем (55–65%) занимает основная фотосинтезирующая ткань – складчатый мезофилл, клетки которого располагаются двумя-тремя рядами вокруг центрального цилиндра (рис. 1). Среди них формируются 8–12 смоляных ходов. Эндодерма, разграничивающая фотосинтезирующую и проводящие ткани, хорошо выражена. Парциальный объем центрального цилиндра, включающего паренхиму, в т.ч. трансфузионную, межпучковую склеренхиму и два проводящих пучка, составляет до 30%.
Рис. 1.
Поперечный срез хвои сосны обыкновенной. 1 – эпидерма, 2 – гиподерма, 3 – мезофилл, 4 – смоляной ход, 5 – эндодерма, 6 – проводящий пучок.
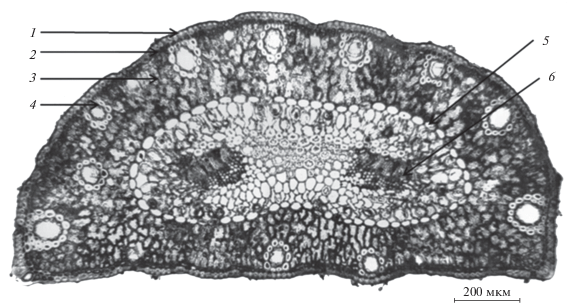
В сосняках лишайниковых в условиях многолетнего аэротехногенного пресса показаны изменения морфологических параметров хвои сосны. В период, когда объем выбросов СЛПК составлял около 30 тыс. т в год, длина однолетней хвои на загрязненной территории не отличалась от контроля, а площадь ее поперечного сечения была на 23% меньше, чем на фоновом участке (Ладанова, Плюснина, 1998; Тужилкина и др., 1998). После значительного сокращения объемов выбросов хвоя в зоне действия СЛПК была в 1.3–1.5 раза короче, а площадь ее поперечного сечения на 40% меньше, чем на фоновом участке (табл. 2). На протяжении всего периода наблюдений радиальные диаметры клеток тканей (эпидерма, гиподерма, мезофилл) достоверно не отличались на участках с разной степенью загрязнения. В ранних работах (Ладанова, Плюснина, 1998; Тужилкина и др., 1998) также было показано отсутствие значительных отклонений от контроля в парциальных объемах тканей на поперечном срезе однолетней хвои в сосняках лишайниковых около СЛПК. Изучение образцов 2015 г. показало, что изменение площади поперечного сечения происходит за счет уменьшения числа структурных элементов в листовых органах. Так, например, в хвое с контрольного участка в этот период между гиподермой и эндодермой чаще формировалось три ряда клеток мезофилла, а на загрязненных участках – два. Согласно полученным данным в хвое на загрязненной территории также уменьшалось число и диаметр смоляных ходов по сравнению с контролем (табл. 2), что ранее было отмечено также для ели, произрастающей вблизи лесопромышленного комплекса (Тужилкина, Плюснина, 2014). Эфирные масла, которые являются основными компонентами смолы и синтезируются в эпителиальных (терпеноидогенных) клетках смоляных ходов хвойных растений, выполняют защитные функции: они токсичны для большинства травоядных и насекомых-вредителей (Хелдт, 2011) и обладают, в т.ч. антифунгальной активностью (Cavaleiro et al., 2006). Сокращение смолоносной зоны в хвое потенциально может снижать ее сохранность и устойчивость к патогенным микроорганизмам.
Таблица 2.
Морфологическая характеристика хвои сосны на разном расстоянии от источника загрязнения (2015 г.)
| Показатель | Расстояние от источника эмиссии, км | ||
|---|---|---|---|
| 7.3 | 11 | 56 | |
| Длина хвои, мм | 35.1 ± 1.5*** | 39.4 ± 3.0*** | 53.2 ± 1.6 |
| Площадь сечения хвои, мм2 | 0.73 ± 0.09** | 0.71 ± 0.03*** | 1.17 ± 0.11 |
| Число смоляных ходов | 8.8 ± 0.6 | 7.6 ± 1.0* | 10.0 ± 1.2 |
| Диаметр смоляных ходов, мкм | 106.6 ± 7.5* | 106.0 ± 4.7** | 129.3 ± 9.5 |
| Парциальный объем смоляных ходов в хвое, % | 10.7 | 9.5 | 11.1 |
Известно, что морфологические показатели листовых органов зависят от многих природных факторов (Николаевский, 1979). В естественных условиях микрофиллия является характерной реакцией растений на ухудшение условий окружающей среды. Чаще всего это происходит при уменьшении доступной влаги и/или тепла (Горышина, 1989). В условиях разного режима аэротехногенного загрязнения на Кольском полуострове было показано отсутствие корреляции длины, ширины и толщины хвои сосны с уровнем загрязнения местообитания (Ярмишко, Лянгузова, 2013). Таким образом, уменьшение размерных параметров листа как в естественных, так и в антропогенных условиях часто является ответной реакцией растения на изменение экологических факторов и не всегда служит свидетельством его угнетения. Если при аэротехногенном загрязнении степень повреждения хвои (хлорозы, некрозы), ее сохранность близки к контрольным показателям, это может быть следствием оптимизации структуры фотосинтетического органа в данных конкретных условиях. В работе (Робакидзе и др., 2017) показано, что в сосняках лишайниковых в зоне действия СЛПК после снижения объемов выбросов наблюдается увеличение доли деревьев без потерь хвои, сокращение доли сосен с высокой степенью пожелтения кроны и процента поврежденных деревьев в древостоях в целом. Там же отмечено, что накопление серы в хвое и других элементов, содержащихся в выбросах, не отличается от контроля, а запас стволовой древесины древостоев увеличивается.
Изучение ультраструктурных и биохимических показателей позволяет более точно определить состояние ассимиляционного аппарата в условиях аэротехногенного загрязнения, когда видимые повреждения отсутствуют.
Ультраструктура клеток мезофилла. В период с высокими объемами выбросов, включающих хлорсодержащие компоненты, в зоне загрязнения клетки мезофилла характеризовались повышенной осмиофильностью, а хлоропласты – низким содержанием крахмала и высокой электронной плотностью стромы, что не позволяло детально рассмотреть тилакоидную систему (Тужилкина и др., 1998). В настоящее время в клетках мезофилла на загрязненных участках выявляются схожая с контролем осмиофильность и электронная плотность структур, мембраны органелл хорошо определяются (рис. 2). Показано снижение в 1.4 раза числа гран на срезе хлоропласта (табл. 3) по сравнению с фоновой территорией. Их недостаток компенсируется повышением до 1.5 раз числа пластид на срезе клетки. В итоге на загрязненной территории суммарное число фотосинтетических мембран на срезе клетки мезофилла было несколько выше, чем на фоновой. Содержание крахмала в хлоропластах сильно варьировало, особенно в контроле, где его было значительно меньше, чем на загрязненных участках. Крахмал – одно из запасных веществ растений, образуется в хлоропластах при полимеризации глюкозы (Хелдт, 2011). Запасные углеводы откладываются также в клеточной стенке. Показано, что ее толщина в мезофилле хвои на загрязненных участках выше, чем в контроле (табл. 3). Другие запасающие глобулы – крупные белковые и небольшие липидные – также присутствуют в цитоплазме клеток мезофилла. В клетках мезофилла установлено снижение числа митохондрий в условиях загрязнения.
Рис. 2.
Фрагменты клеток мезофилла (тангентальный срез) хвои сосны на расстоянии 56 км (а) и 7.3 км (б) от источника эмиссии в 2015 г. 1 – вакуоль, 2 – клеточная оболочка, 3 – крахмал, 4 – межклетник, 5 – митохондрия, 6 – хлоропласт.
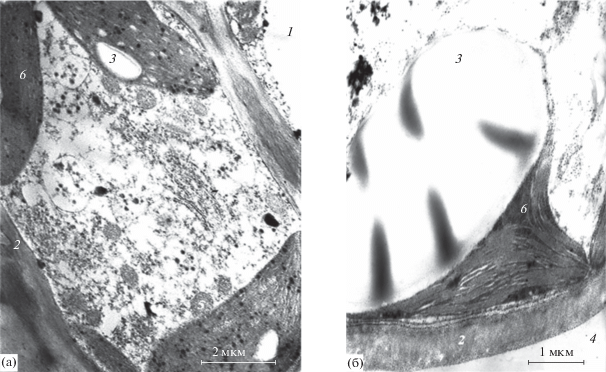
Таблица 3.
Характеристика клеток мезофилла однолетней хвои сосны на разном расстоянии от источника загрязнения (2015 г.)
| Параметр | Расстояние от источника эмиссии, км | ||
|---|---|---|---|
| 7.3 | 11 | 56 | |
| Число митохондрий на срез клетки | 17.8 ± 3.6*** | 24.9 ± 7.3** | 33.5 ± 8.7 |
| Площадь среза митохондрии, мкм2 | 0.34 ± 0.09 | 0.38 ± 0.07 | 0.35 ± 0.09 |
| Число хлоропластов на срез клетки | 14.2 ± 2.1*** | 15.6 ± 3.7*** | 10.7 ± 2.5 |
| Площадь среза хлоропласта без крахмала, мкм2 | 5.0 ± 1.2 | 7.4 ± 1.7 | 7.4 ± 1.6 |
| Число гран на срез хлоропласта | 33.3 ± 9.5*** | 34.1 ± 9.7*** | 47.7 ± 9.8 |
| Число тилакоидов в гране | 6.5 ± 1.8 | 5.3 ± 2.0 | 5.4 ± 1.1 |
| Доля хлоропластов с крахмалом, % | 90.9 ± 9.0*** | 81.0 ± 12.1*** | 58.7 ± 13.2 |
| Парциальный объем крахмального зерна в хлоропласте, % | 62.0 ± 8.2*** | 62.4 ± 19.4*** | 27.7 ± 15.1 |
| Толщина клеточной оболочки, мкм | 0.62 ± 0.05* | 0.70 ± 0.08*** | 0.54 ± 0.14 |
Пигментный комплекс. Известно, что одним из биохимических показателей реакции растений на изменение факторов внешней среды является содержание хлорофиллов и каротиноидов – главных фоторецепторов ассимилирующей клетки. Нами раннее (Тужилкина и др., 1998; Тужилкина, 2009) было установлено влияние загрязнения среды промышленными выбросами СЛПК на пигментную систему сосны и ели. Реакция пигментного комплекса хвои на загрязнение воздуха неодинакова и зависит от техногенной нагрузки и условий местопроизрастания. В 1995 г. в зоне значительного влияния, с максимальной техногенной нагрузкой в 20–100 раз превышающей фоновую, пигментный аппарат сосны проявлял повышенную чувствительность к токсикантам, выражающуюся в снижении синтеза фотосинтетических пигментов (рис. 3а). Следует отметить, что хвоя опытных деревьев в сосняке лишайниковом отличалась от контрольных не только по суммарному количеству зеленых и желтых пигментов, изменялось и соотношение зеленых пигментов. Так, изменения в пигментном комплексе под влиянием загрязнения происходили главным образом за счет снижения содержания хлорофилла b. Концентрация хлорофилла а в хвое экспериментальных деревьев уменьшилась на 25.7%, а хлорофилла b – на 34.7% по сравнению с контролем.
Рис. 3.
Содержание пигментов в однолетней хвое сосны в лишайниковом типе леса в 1995 г. (а) и в 2015 г. (б). 1 – фоновый район; 2 – загрязненный (7.3 км от СЛПК). Различия с контролем статистически значимы при: *p < 0.05; **p < 0.01.
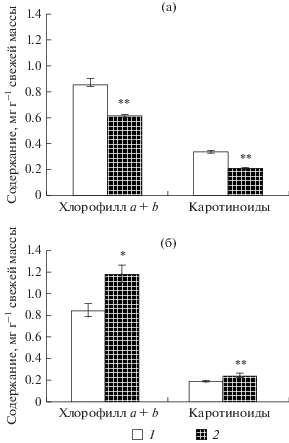
Исследования пластидного аппарата сосны, проведенные в 2015 г., показали, что в пигментном комплексе хвои произошли изменения по сравнению с 1995 г. При снижении техногенной нагрузки в хлоропластах сосны произошло усиление процессов образования пигментов в хвое. Концентрация хлорофиллов и каротиноидов повысилась в 1.2–1.3 раза по сравнению с контролем (рис. 3б). Согласно (Гетко, 1989) увеличение содержания хлорофиллов в условиях длительного техногенного пресса может быть связано с накоплением продуктов окисления углеводов – органических кислот цикла Кребса, продуктов гидролиза белков, необходимых для синтеза пигментов. При этом распределение хлорофилла по фотосинтетическим пулам у деревьев в зоне загрязнения, как показали наши исследования, достоверно не отличалось от контрольных. С приближением к источнику эмиссий отмечается низкая фотосинтетическая активность единицы хлорофилла (табл. 4), что, по-видимому, связано со снижением скорости обновления хлорофилла и числом молодых молекул (Гапоненко, 1971; Šestak, 1977). Это, в свою очередь, может оказать влияние на фотосинтетические процессы у деревьев и их устойчивость.
Таблица 4.
Физиолого-биохимические показатели однолетней хвои сосны на загрязненной и фоновой территории, июль 2015 г.
| Показатель | Расстояние от источника эмиссии, км | ||
|---|---|---|---|
| 7.3 | 11 | 56 | |
| Хлорофилл а, мг · г–1 сухой массы | 1.83 ± 0.1** | 1.57 ± 0.04* | 1.36 ± 0.07 |
| Хлорофилл b, мг · г–1 сухой массы | 0.65 ± 0.03* | 0.57 ± 0.04 | 0.49 ± 0.06 |
| Хлорофилл а: Хлорофилл b | 2.8 ± 0.2 | 2.8 ± 0.2 | 2.8 ± 0.2 |
| Доля хлорофилла в светособирающем комплексе, % | 58 | 59 | 58 |
| Каротиноиды, мг · г–1 сухой массы | 0.50 ± 0.05* | 0.46 ± 0.03 | 0.41 ± 0.02 |
| Отношение хлорофилл/каротиноиды | 5.0 ± 0.2 | 4.7 ± 0.1 | 4.5 ± 0.1 |
| Фотосинтез, мкмоль СО2 · м–2 · с–1 | 2.02 ± 0.30** | 3.12 ± 0.16 | 3.76 ± 0.30 |
| Темновое дыхание, мкмоль СО2 · м–2 · с–1 | 0.55 ± 0.05* | 0.42 ± 0.04 | 0.39 ± 0.04 |
| Ассимиляционное число, мг СО2 · мг хл.–1 · ч–1 | 1.63 | 2.74 | 3.76 |
Углекислотный газообмен. Техногенное загрязнение атмосферы и почвы оказывает существенное воздействие на процессы фотосинтеза и дыхания растений. Характер и степень влияния зависят от концентрации поллютанта (Oleksyn, 1984; Кайбияйнен и др., 1998). Измерения СО2-газообмена хвои сосны показали, что интенсивность фотосинтеза хвои деревьев в сосняках лишайниковых, произрастающих на расстоянии 7.3 км от источника выбросов, почти в 2 раза ниже по сравнению со скоростью поглощения углекислоты хвоей из фонового района (p < 0.05). Ингибирование процесса фотосинтеза хвои у сосны в условиях техногенного загрязнения атмосферы ранее было отмечено другими исследователями (Кайбияйнен и др., 1995; Тарханов, Бирюков, 2014). Торможение фотосинтеза сопровождалось повышением скорости темнового дыхания. Скорость выделения углекислоты на загрязненной территории (7.3 км от СЛПК) была в 1.4 раза выше по сравнению с интенсивностью дыхания хвои из контрольного района. Стимуляция процесса дыхания у ели в условиях промышленной зоны была отмечена нами ранее (Тужилкина, Плюснина, 2014). В промышленной зоне увеличение дыхания, как правило, наблюдается при низких концентрациях токсиканта, а при больших его дозах скорость дыхания падает (Кайбияйнен и др., 1998).
С ослаблением техногенной нагрузки (11 км от СЛПК) прослеживается тенденция снижения скорости фотосинтеза. Однако эти различия недостоверны. Дыхательная способность хвои не отличается от фоновой, что свидетельствует о сохранении стабильного уровня выделения СО2 сосны в условиях промышленного загрязнения.
ЗАКЛЮЧЕНИЕ
Мониторинг ассимиляционного аппарата сосны в лишайниковых сообществах на территории Республики Коми позволил выявить неодназначную реакцию структуры и функции хвои, содержания хлорофиллов и каротиноидов на воздействие аэротехногенных выбросов АО “Монди Сыктывкарский СЛПК” за 2 срока наблюдений (1995 и 2015 г.). В 1995 г. промышленные выбросы целлюлозно-бумажного производства в зоне значительного действия техногенной нагрузки оказывали отрицательное влияние на структуру клеток мезофилла и пигментную систему фотосинтетического аппарата сосны. Поллютанты способствовали ослаблению процессов накопления хлорофилла в пластидах, вызывали определенные изменения в пигментном комплексе, которые происходили главным образом за счет снижения содержания хлорофилла b (Тужилкина и др., 1998). Накопление крахмала в хлоропластах ассимилирующей паренхимы сокращалось. После снижения объемов выбросов была отмечена оптимизация структуры ассимилирующих органов, что сопровождалось уменьшением размеров хвои (длины и площади сечения) за счет сокращения числа структурных компонентов (числа рядов клеток мезофилла и смоляных ходов на поперечном срезе) и изменением численности органелл (увеличение числа хлоропластов и уменьшение числа митохондрий) в ассимиляционной паренхиме. Снижение затрат на формирование структурных компонентов хвои приводило к избыточному, по сравнению с контролем, накоплению запасных веществ в клетках мезофилла. В 2015 г., помимо увеличения числа хлоропластов, показано усиление процессов образования крахмала и пигментов в них, что отражает компенсаторный механизм адаптации хвои сосны в условиях промышленного загрязнения. Наши данные частично подтверждают гипотезу авторов (Мальхотра, Хан, 1988; Кравкина, 2001) о проявлении на структурно-биохимическом уровне адаптивных механизмов фотосинтетического аппарата в клетках хвои сосны, произрастающей в зоне загрязнения СЛПК. Установлено, что хлоропласты и их мембраны особенно чувствительны к действию загрязняющих веществ (Jäger, Klein, 1980; Кравкина, 1991) и могут служить в качестве биомаркеров оценки загрязнения окружающей среды (Vodka, 2017). Одной из важных структурных характеристик, определяющих интенсивность фотосинтеза, является тилакоидная система. В нашем исследовании действие поллютантов на число тилакоидов в гране не проявилось, в отличие от числа гран на срез хлоропласта, которое уменьшалось. Выбросы СЛПК вблизи источника эмиссии способствовали снижению фотосинтетической способности и увеличению интенсивности дыхания хвои, что в свою очередь обеспечивает энергетические потребности адаптации ее к поллютантам. С ослаблением техногенной нагрузки пигментная система, фотосинтетическая и дыхательная способности хвои не претерпевают изменений.
Таким образом, с 1995 по 2015 г. в сосняке лишайниковом зоны воздействия СЛПК у сосны обыкновенной при ослаблении загрязнения среды сернистыми и азотистыми соединениями выявлены структурно-биохимические перестройки в клетках мезофилла, направленные на стабилизацию процессов фотосинтеза и дыхания и способствующие устойчивости хвои к условиям загрязнения в районе действия целлюлозно-бумажного производства.
Авторы благодарят ведущих инженеров отдела лесобиологических проблем Севера Института биологии Коми НЦ УрО РАН С.Н. Кузина, С.П. Швецова и А.И. Патова за помощь в работе с электронным микроскопом и газоанализатором.
Список литературы
Антипов В.Г. Устойчивость древесных растений к промышленным газам. Минск: Наука и техника, 1979. 215 с.
Барахтенова Л.А. Влияние сернистого газа на фотосинтетический метаболизм у растений // Изв. АН СССР. Сер. биол. 1987. Вып. 2. С. 45.
Воробейчик Е.Л., Трубина М.Р., Хантемирова Е.В., Бергман И.Е. Многолетняя динамика лесной растительности в период сокращения выбросов медеплавильного завода // Экология. 2014. № 6. С. 448–458.
Гапоненко В.И. Обновление хлорофилла в фотосинтезирующем аппарате как физиологический процесс // Проблемы биосинтеза хлорофилла. Минск: Наука и техника, 1971. С. 78–137.
Гетко Н.В. Растения в техногенной среде. Минск: Наука и техника, 1989. 208 с.
Горышина Т.К. Фотосинтетический аппарат растений и условия среды. Л.: Изд-во Ленинградского университета, 1989. 204 с.
Государственный доклад о состоянии окружающей природной среды Республики Коми в 1996 г. Сыктывкар, 1997. 148 с.
Государственный доклад “О состоянии окружающей среды Республики Коми в 2015 г.”. Сыктывкар, 2016. 173 с.
Дьяконов В.В. Состояние лесов охраняемых территорий Карелии в связи с атмосферным загрязнением // Проблемы антропогенной трансформации лесных биогеоценозов Карелии. Петрозаводск: КНЦ РАН, 1996. С. 6–17.
Кайбияйнен Л.К., Софронова Г.И., Болондинский В.К. Влияние токсичных поллютантов на дыхание хвои и побегов сосны обыкновенной // Экология. 1998. № 1. С. 23–27.
Кайбияйнен Л.К., Хари П., Софронова Г.И., Болондинский В.К. Влияние длительности воздействия токсичных поллютантов на состояние устьиц и фотосинтез хвои // Физиология растений. 1995. Т. 42. № 5. С. 751–757.
Кравкина И.М. Влияние атмосферных загрязнителей на структуру листа // Ботанический журн. 1991. Т. 76. № 1. С. 3–9.
Кравкина И.М. Реакция пластидома и хондриома хлоренхимы листа на длительное аэротехногенное загрязнение // Физиология растений. 2001. Т. 48. С. 276–281.
Кулагин Ю.З. Индустриальная дендроэкология и прогнозирование // М.: Наука, 1985. 117 с.
Ладанова Н.В., Плюснина С.Н. Анатомо-морфологические изменения разновозрастной хвои сосны обыкновенной в зоне действия Сыктывкарского лесопромышленного комплекса // Лесной журн. 1998. № 1. С. 7–11.
Личутина Т.Ф., Боголицын К.Г., Гусакова М.А. Экологическая оценка деятельности предприятий целлюлозно-бумажной промышленности. Перспективные направления утилизации отходов // Российский химический журн. 2011. Т. LV. № 1. С. 101–107.
Мальхотра С.С., Хан А.А. Биохимическое и физиологическое действие приоритетных загрязняющих веществ // Загрязнение воздуха и жизнь растений / Под ред. Трешоу М.Л. М.: Гидрометиздат, 1988. С. 144–189.
Михайлова И.Н. Начальные этапы восстановления сообществ эпифитных лишайников после снижения выбросов медеплавильного завода // Экология. 2017. № 4. С. 277–281.
Николаевский В.С. Биологические основы газоустойчивости растений. Новосибирск: Наука, 1979. 278 с.
Робакидзе Е.А., Торлопова Н.В. Мониторинг состояния ельников в условиях загрязнения целлюлозно-бумажного производства // Растительные ресурсы. 2018. Т. 54. № 1. С. 42–58.
Робакидзе Е.А., Торлопова Н.В., Бобкова К.С. Состояние хвойных фитоценозов защитной зоны г. Сыктывкара // Сохранение лесных экосистем: проблемы и пути их решения: Матер. Всерос. науч-практич. конф. Киров: Радуга-ПРЕСС, 2017. С. 179–181.
Рунова Е.М. Влияние техногенного загрязнения на состояние хвойных древостоев: Автореф. дисс. …д-ра. с.-х. наук. Красноярск, 1999. 42 с.
Сергейчик С.А. Древесные растения и оптимизация промышленной среды. Минск: Наука и техника, 1984. 168 с.
Скупченко В.Б. Морфометрия на экране электронного микроскопа // Ботанический журн. 1990. Т. 75. № 10. С. 1463–1467.
Сухарева Т.А., Лукина Н.В. Минеральный состав ассимилирующих органов хвойных деревьев после снижения уровня атмосферного загрязнения на Кольском полуострове // Экология. 2014. № 2. С. 97.
Тарханов С.Н., Бирюков С.Ю. Морфоструктура и изменчивость биохимических признаков популяции сосны (Pinus sylvestris L.) в стрессовых условиях устья Северной Двины // Сибирский экологический журн. 2014. Т. 21. № 2. С. 319–327.
Тарханов С.Н., Прожерина Н.А., Коновалов В.Н. Лесные экосистемы бассейна Северной Двины в условиях атмосферного загрязнения: диагностика состояния. Екатеринбург: УрО РАН, 2004. 333 с.
Торлопова Н.В., Робакидзе Е.А. Влияние поллютантов на хвойные фитоценозы (на примере Сыктывкарского лесопромышленного комплекса). Екатеринбург: УрО РАН, 2003. 147 с.
Тужилкина В.В. Реакция пигментной системы хвойных на длительное аэротехногенное загрязнение // Экология. 2009. № 4. С. 243–248.
Тужилкина В.В., Ладанова Н.В, Плюснина С.Н. Влияние техногенного загрязнения на фотосинтетический аппарат сосны // Экология. 1998. № 2. С. 89–93.
Тужилкина В.В., Плюснина С.Н. Комплексная оценка состояния хвои Picea obovata (Pinaceae) в условиях аэротехногенного загрязнения // Растительные ресурсы. 2014. Т. 50. Вып. 4. С. 579–587.
Хелдт Г.-В. Биохимия растений. М.: БИНОМ, 2011. 471 с.
Шлык А.А. Определение хлорофиллов и каротиноидов в экстрактах зеленых листьев // Биохимические методы в физиологии растений. М.: Наука, 1971. С. 154–170.
Ярмишко В.Т., Горшков В.В., Лянгузова И.В., Баккал И.Ю. Экологический мониторинг лесных экосистем Кольского полуострова в условиях аэротехногенного загрязнения // Региональная экология. 2011. № 1–2(31). С. 21–29.
Ярмишко В.Т., Лянгузова И.В. Многолетняя динамика параметров и состояния хвои Pinus sylvestris L. в условиях аэротехногенного загрязнения на европейском Севере // Известия Санкт-Петербургской лесотехнической академии. 2013. № 203. С. 30–46.
Ярмишко В.Т., Лянгузова И.В., Лянгузов А.Ю. Изменение годичного прироста стволов Pinus sylvestris (Pinaceae) при снижении аэротехногенного загрязнения // Растительные ресурсы. 2017. Т. 53. № 4. С. 527–542.
Cavaleiro C., Pinto E., Goncalves M.J., Salgueiro L. Antifungal activity of Juniperus essential oils against dermatophyte, Aspergillus and Candida strains // J. Applied Microbiology. 2006. V. 100. P. 1333–1338.
Huttunen S., Laine K. Effects of air borne pollutants on the surface wax structure of Pinus sylvestris needles // Annales Botanici Fennici. 1983. V. 20. № 1. P. 79–86.
Jäger H.-J., Klein H. Biochemical and physiological effects of SO2 on plants // Angewandte Botanik. 1980. 54. P. 337–348.
Kukkola E., Huttunen S., Bӓck J., Rautio P. Scots pine needle injuries at subarctic industrial sites // Trees. 1997. V. 11. P. 378–387.
Oleksyn J. Effect of SO2, HF and NO2 on net photosynthesis and dark respiration rates of Scots pine needles of various ages // Photosynthetica. 1984. V. 18. № 2. P. 259–262.
Soikkeli S. Comparison of cytological injuries in conifer needles from several polluted industrial environments in Finland // Annales Botanici Fennici. 1981. V. 18. № 1. P. 47–61.
Šestak Z. Photosynthetic characteristics during ontogenesis of leaves. 1. Chlorophylls // Photosynthetica. 1977. V. 11. № 4. P. 367–387.
Vodka M.V. Chloroplasts ultrastructural changes as biomarkers of acid rain and heavy metals pollution // Biotechnologia Acta. 2017. V. 10. № 3. P. 57–64.
Дополнительные материалы отсутствуют.