

Лесоведение, 2022, № 2, стр. 115-131
Мелколиственные леса центра Русской равнины: экология и перспективы возобновления коренных сообществ
Н. Г. Беляева a, *, О. В. Морозова a, Т. В. Черненькова a, К. А. Корзников b, Е. Г. Суслова c
a Институт географии РАН
119017 Старомонетный переулок, 29, Москва, Россия
b Ботанический сад-институт ДВО РАН
690024 ул. Маковского, 142, Владивосток, Россия
c Московский государственный университет им. М.В. Ломоносова
119991 Ленинские горы, 1, Москва, Россия
* E-mail: belyaeva@igras.ru
Поступила в редакцию 08.07.2021
После доработки 12.07.2021
Принята к публикации 01.12.2021
- EDN: FRVMJV
- DOI: 10.31857/S0024114822020036
Аннотация
Мелколиственные леса центра Русской равнины имеют широкое распространение, и их площадь продолжает увеличиваться в связи с зарастанием брошенных сельскохозяйственных земель. В Московской области они занимают почти половину всех лесов региона. Рассмотрены состав, экология и динамика лесов с доминированием березы (Betula pendula, B. pubescens), осины (Populus tremula), ольхи серой (Alnus incana) и ольхи черной (Alnus glutinosa) в центре Русской равнины на примере Московского региона. На основе эколого-фитоценотической классификации выделены 11 групп ассоциаций, из которых 8 относятся к производным и 3 – к условно-коренным типам сообществ. Установлено, что основным экологическим фактором, влияющим на различия состава лесов, является увлажнение почв. Такие факторы, как освещенность, кислотность и богатство почв азотом, дифференцируют сообщества на следующих уровнях в рамках двух кластеров групп различного увлажнения почв. Участие основных коренных лесообразующих видов деревьев меняется в зависимости от экологических условий местообитаний мелколиственных лесов. Ель европейская (Picea abies) наиболее активна в среднем диапазоне экологических условий, в крайних значениях экологических факторов ее участие уменьшается. Липа мелколистная (Tilia cordata) и дуб черешчатый (Quercus robur) чаще отмечены в условиях увеличения богатства почв и хорошего дренажа, а сосна (Pinus sylvestris) – только в условиях хорошего освещения, которые складываются в березовых лесах кустарничково-травяно-сфагновой группы. В подросте условно-коренных серо- и черноольховых лесов высоко участие ольхи серой и ольхи черной, в то время как в производных березовых и осиновых сообществах преобладает в основном ель. Вероятно, при отсутствии нарушений березняки широкотравные, осинники широкотравные и влажнотравно-широкотравные скорее остальных будут заменены условно-коренными хвойно-широколиственными сообществами, поскольку на фоне преобладания ели в подросте в данных группах хорошо возобновляется липа, а также отмечается примесь широколиственных пород в древостое.
Основными лесообразующими породами хвойно-широколиственной зоны Русской равнины, согласно взглядам С.Ф. Курнаева (1968), являются ель европейская (Picea abies)22 и липа мелколистная (Tilia cordata). Такие леса должны занимать господствующее положение на водораздельных пространствах, особенно на покровных суглинках, подстилаемых моренными отложениями. Согласно альтернативной точке зрения (Разумовский, 2011), коренным типом растительности этой полосы являются дубравы с дубом черешчатым (Quercus robur). Однако вследствие многовековой хозяйственной деятельности коренные леса на этой территории повсеместно были сведены, а на их месте в настоящее время существуют разного рода производные33 сообщества, существенная доля которых приходится на мелколиственные леса.
Хвойно-широколиственные леса Русской равнины начали массово сводиться под пашни еще в XI–XII вв., а к началу XVI в. “уровень распашки приблизился к максимально возможному”. Поскольку поддерживать плодородие на постоянных полях не получалось, трехпольное земледелие сочеталось с переложным, по крайней мере, до второй половины XIX в. (Абатуров, 2000; Милов, 2006). Это приводило к высокому участию березовых и осиновых лесов в лесном покрове хвойно-широколиственной зоны. Создание культур еловых и сосновых лесов в XX веке и изменение способов ведения сельского хозяйства несколько снизили участие мелколиственных сообществ, однако в последние десятилетия наблюдается активное зарастание заброшенных сельскохозяйственных угодий (Potapov et al., 2015), что приводит к увеличению их доли в лесном покрове центра Русской равнины. На территории Московской области уже к концу XVIII в. леса в основном были производными мелколиственными (Рахилин, 1997; Беляева, Попов, 2016). По официальным данным, в настоящее время мелколиственные леса произрастают на 40.6% ее лесопокрытой площади (Лесной план, 2018), среди них березовые составляют 77% (Лесной план, 2018).
К мелколиственным видам деревьев иногда относят только березу и осину (Алехин, 1951), другие авторы включают также ольху серую и ивы (Ниценко, 1972). Ивовые леса из ивы козьей (Salix caprea), а также пойменные из ивы белой (Salix alba) и ивы ломкой (S. fragilis), которые тянутся прерывистыми полосами вдоль рек, формируют небольшие массивы в регионе. Не все мелколиственные леса являются производными. Условно-коренные44 типы сообществ (которые иногда рассматриваются как длительнопроизводные) приурочены главным образом к переувлажненным местообитаниям (заболоченным различного типа, приручьевым и пойменным). Авторы ранее опубликованных работ сообщают о высоком типологическом разнообразии производных и условно-коренных мелколиственных лесов (Ниценко, 1972; Абатуров и др., 1982; Василевич, 1996; Василевич, Щукина, 2001). Для территории Европейской России А.А. Ниценко (1972) на основе экологических условий местообитаний выделено 13 групп ассоциаций только для березовых лесов, а ассоциаций всех мелколиственных лесов, выделенных на основе доминирования видов, насчитывается несколько десятков. Такое дробное деление ограничивает статистическую обработку и корректный анализ в рамках данной работы. Для Северо-Запада России на основе доминантно-флористического подхода описана 31 ассоциация формаций сероольховых, березовых и осиновых лесов (Василевич, 1996; Бибикова, 1998; Василевич, 1998). Причины высокого ценотического разнообразия связаны с различным сукцессионным состоянием сообществ и с широким экологическим ареалом основных лесообразователей, в первую очередь, березы пушистой и березы повислой (Atkinson, 1992; Ветчинникова, 2004). Осина является гораздо более требовательной к условиям произрастания – она не переносит кислых и плохо дренированных почв (Ниценко, 1972; Possen et al., 2011), плохо растет на сухих и небогатых почвах (Blumenthal, 1942), однако хорошо возобновляется как семенным, так и вегетативным способом и заселяет вырубки даже активнее, чем береза (Ниценко, 1972), при условии подходящего местообитания и наличия источника семян. При сочетании экологических условий (достаточно дренированные и богатые местообитания) лесообразующими породами являются и береза, и осина, а в травяно-кустарничковом ярусе принимают участие виды неморальной и нитрофильно-неморальной групп.
Мелколиственные леса играют важную роль в лесном покрове: улучшают качество почв (увеличивается содержание калия, кальция, магния), выполняют водорегулирующую и противоэрозионную функции, участвуют в стоке атмосферной углекислоты, формируют местообитания для многих видов живых организмов, поддерживают биоразнообразие, обладают высокой продуктивностью (Ниценко, 1972; Perala, Alm, 1990; Brandtberg et al., 2000; Organisation …, 2003; Ferm, 1993; Гульбе и др., 2016). Одна из важнейших функций мелколиственных лесов – создание условий для спонтанного (естественного) восстановления условно-коренных сообществ. Несмотря на широкое распространение мелколиственных лесов, систематизированному изучению их видового и типологического состава исследователи уделяли гораздо меньше внимания по сравнению с условно-коренными хвойными, хвойно-широколиственными и широколиственными лесами.
Цель исследования – выявление типологического разнообразия мелколиственных лесов и оценка перспектив возобновления коренных типов сообществ на основе анализа состава подроста и экологической приуроченности разных типов сообществ на примере территории Московского региона.
ОБЪЕКТЫ И МЕТОДИКА
Район исследований включает всю территорию Московской области, которая располагается в центральной части Русской равнины (35°10′–40°15′ в.д., 54°12′–56°55′ с.ш.) и относится в основном к зоне широколиственно-хвойных лесов, а на юге – широколиственных (Курнаев, 1973; Грибова и др., 1980). Перепад высот составляет 90–320 м, а средние уклоны – 2° (Chernenkova et al., 2020).
В данной работе к мелколиственным сообществам отнесены березовые, осиновые, сероольховые и черноольховые леса. Традиционно считается, что береза повислая и береза пушистая в центре Русской равнины разделяются по экологическим предпочтениям: береза пушистая может произрастать в переувлажненных местообитаниях, в то время как береза повислая предпочитает хороший дренаж (Ветчинникова, 2004). На практике оба вида берез по морфологическим признакам трудно различимы, поскольку обладают высокой полиморфностью, к тому же в природе широко распространены гибриды этих видов (Perala, Alm, 1990). Молекулярно-генетические исследования показали (Маслов и др., 2019), что оба вида берез могут встречаться в различных условиях увлажнения. В связи с этим при анализе сообществ с участием берез мы вслед за А.А. Ниценко (1972) и В.И. Василевичем (1996) не выделяем формации из березы пушистой и березы повислой, а рассматриваем их в рамках одной формации березовых лесов.
Работа основана на анализе полного видового состава сосудистых растений и мохообразных 435 геоботанических описаний, собранных в 1998–2019 гг. и выполненных по стандартной методике. Нами принято традиционное обозначение ярусной структуры сообществ: А – древесный ярус (иногда с двумя ярусами), В – деревья и кустарники высотой 1–10 м; С – травяно-кустарничковый ярус (в том числе деревья и кустарники высотой ниже 1 м); D – мохово-лишайниковый ярус. Классификация сообществ проведена с использованием эколого-фитоценотического подхода (Черненькова, Морозова, 2017). Выделены синтаксоны уровня групп ассоциаций на основе представленности эколого-морфологических групп видов подчиненных ярусов и доминирующему виду (видам) в древостое. По содержанию синтаксоны согласуются с типологическими категориями, используемыми в отечественной геоботанической школе (Рысин, Савельева, 2007; Огуреева и др., 2008). Названия синтаксонов продублированы латинским вариантом в соответствии с кодексом номенклатуры (Нешатаев, 2001). Для формальной проверки классификации применен линейный пошаговый дискриминантный анализ в программе IBM SPSS Statistics 22. В качестве переменных предикторов использованы значения покрытий видов древостоя и сумма покрытий видов, относящихся к определенной эколого-ценотической группе (ЭЦГ). Отнесение видов к ЭЦГ выполнено по модифицированной схеме В.Э. Смирнова с соавторами (2006) c учетом диагностических видов классов растительности в системе Браун-Бланке (Ермаков, 2012; Mucina et al., 1993).
Для выявления приуроченности типов сообществ к экологическим условиям местообитаний проведена NMDS ординация описаний в среде статистического программирования R (R Core Team, 2019). На основе экологических шкал Элленберга (Ellenberg et al., 1991) для каждого описания в программе Juice 7.0 (Tichý, 2002) рассчитаны показатели кислотности почв (R), увлажнения почв (M), богатства почв азотом (N) и освещенности (L) с учетом покрытия вида (weighted by species cover). В видовом списке мелколиственных лесов региона (по нашим описаниям) 84% видов присутствуют в списке Элленберга (Ellenberg et al., 1991).
Для отображения изолиний участия коренных видов деревьев в составе сообществ (ели, сосны, липы и дуба) на ординационном пространстве посчитана сумма значений проективных покрытий каждого вида во всех ярусах (A, B, C) для каждого описания. По экологическим переменным (шкалам Элленберга) построено дерево решений в программе R. Для построения дерева решений по участию коренных видов деревьев в производных мелколиственных лесах были случайным образом отобраны по 10 описаний и использованы значения покрытий сосны, ели и широколиственных видов отдельно по ярусам A, B и C. Отдельно по среднему проективному покрытию подроста в ярусе В построены накопительные гистограммы для всех 11 групп.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Мелколиственные леса отнесены к 11 группам ассоциаций, из которых большая часть (6 групп) принадлежит формации березовых лесов, 2 – осиновых, 2 – черноольховых, 1 – сероольховых лесов (табл. 1).
Таблица 1.
Группы ассоциаций мелколиственных лесов Московской области
| № группы | Формация и группа ассоциаций | Число описаний |
|---|---|---|
| Формация березовых лесов Betuleta pubescentis-pendulae | ||
| 1 | Березовая мелкотравно-широкотравная Betuleta pendulo-pubescentis parvi-nemoraloherbosa |
36 |
| 2 | Березовая широкотравная Betuleta pendulo-pubescentis nemoraloherbosa |
154 |
| 3 | Березовая влажнотравно-широкотравная Betuleta pendulo-pubescentis nitrophilo-nemoraloherbosa |
12 |
| 4 | Березовая травяно-болотная Betuleta pendulo-pubescentis uliginoherbosa |
18 |
| 5 | Березовая разнотравная Betuleta pendulo-pubescentis varioherbosa |
23 |
| 6 | Березовая кустарничково-травяно-сфагновая Betuleta pendulo-pubescentisfruticuloso-herboso-sphagnosa |
10 |
| Формация осиновых лесов Populeta tremulae | ||
| 7 | Осиновая широкотравная Populeta tremulae nemoraloherbosa |
85 |
| 8 | Осиновая влажнотравно-широкотравная Populeta tremulae nitrophilo-nemoraloherbosa |
15 |
| Формация сероольховых лесов Alneta incanae | ||
| 9 | Сероольховая влажнотравно-широкотравная Alneta incanae nitrophilo-nemoraloherbosa |
28 |
| Формация черноольховых лесов Alneta glutinosae | ||
| 10 | Черноольховая влажнотравно-широкотравная Alneta glutinosae nitrophilo-nemoraloherbosa |
23 |
| 11 | Черноольховая травяно-болотная Alneta glutinosae uliginoherbosa |
31 |
Согласно проведенному дискриминантному анализу, общее качество классификации составляет 79.5%. Предсказанная принадлежность анализируемых синтаксонов к определенным классам различна. С наилучшим качеством (100%) выделяются сероольховые влажнотравно-широкотравные леса (группа 9), что в значительной степени объясняется доминированием серой ольхи в древостое только в этой группе. Самое низкое качество классификации наблюдается для березовой мелкотравно-широкотравной группы (66.7%) (группа 1), причем почти 17% описаний, отнесенных по экспертной оценке к данной группе сообществ, анализ распознал как березовые широкотравные леса. Это связано со сложностью отнесения сообществ к мелкотравно-широкотравным или же к широкотравным сообществам внутри одной формации, поскольку наблюдаются естественные плавные переходы соотношения видов мелкотравья и широкотравья. Сообщества остальных групп ассоциаций по результатам анализа были отнесены к предсказанному классу с вероятностью 70% и более. Если же анализ распознавал сообщество ближе к другой группе, то в большинстве случаев относил его в группу той же формации или того же характера травяно-кустарничкового яруса.
Ниже дана общая характеристика сообществ выделенных групп ассоциаций.
Группа 1. Березовые мелкотравно-широкотравные леса. Проективное покрытие (представлены среднее и стандартное отклонение) яруса А – 59 ± 18.5, B – 34 ± 21.1, C – 55 ± 25.6, D – 17 ± 15.4. Первый ярус древостоя сформирован березой (Betula pendula, B. pubescens), иногда с примесью Picea abies и Populus tremula. Второй ярус, если он выражен, образует Picea abies. В ярусе B обычен подрост ели, иногда присутствует Corylus avellana. В травяном ярусе преобладает Oxalis acetosella, часты бореальные виды – Maianthemum bifolium, Pyrola rotundifolia, Rubus saxatilis. С меньшим проективном покрытием, но высокой встречаемостью, отмечены Ajuga reptans, папоротники (Athyrium filix-femina, Dryopteris carthusiana, D. filix-mas), Galeobdolon luteum, Lysimachia nummularia. В нескольких описаниях высокое покрытие отмечено для Aegopodium podagraria и хвощей (Equisetum pratense и E. sylvaticum). Ярус мхов образован Pleurozium schreberi, Hylocomium splendens, Rhytidiadelphus triquetrus, Cirriphyllum piliferum.
Группа 2. Березовые широкотравные леса. Проективное покрытие яруса A – 64.8 ± 17.4, B – 40.9 ± 21.6, C – 66.3 ± 17.9, D – 8.5 ± 9.2. Первый ярус древостоя сформирован Betula pendula и B. pubescens, как примесь встречаются Acer platanoides, Picea abies, Populus tremula, Quercus robur, Tilia cordata. P. abies также формирует второй древесный ярус, который развит не всегда. Ярус В очень хорошо выражен и состоит в основном из Corylus avellana и подроста P. abies. В травяном ярусе с относительно высоким проективным покрытием встречается группа неморальных видов: Aegopodium podagraria, Carex pilosa, Galeobdolon luteum, Pulmonaria obscura, иногда Mercurialis perennis. Часто встречаются, но не образуют высокого проективного покрытия виды: Ajuga reptans, Asarum europaeum, папоротники (Athyrium filix-femina, Dryopteris carthusiana, D. filix-mas), Ranunculus cassubicus, Stellaria holostea. Моховой ярус развит слабо, к наиболее часто встречающимся видам мхов относятся Atrichum undulatum, Cirriphyllum piliferum, Plagiomnium cuspidatum.
Группа 3. Березовые влажнотравно-широкотравные леса. Проективное покрытие яруса A – 57.5 ± 18.5, B – 33.8 ± 20.6, C – 71.7 ± 18.8, D – 9.3 ± 6.1. Лесной полог сформирован Betula pendula и B. pubescens с примесью Populus tremula. Во втором древесном ярусе, при его наличии, присутствуют Picea abies, Betula pendula и B. pubescens, редко – Alnus incana, Quercus robur, Salix caprea. Ярус В разреженный, его образуют Corylus avellana, Frangula alnus, Sorbus aucuparia, а также подрост Picea abies, Betula pendula и B. pubescens. Травяно-кустарничковый ярус состоит из двух подъярусов. Верхний подъярус образован Filipendula ulmaria, папоротниками (Athyrium filix-femina, Dryopteris filix-mas, D. carthusiana), иногда встречаются группировки Urtica dioica. Во втором подъярусе обычны Ajuga reptans, Deschampsia cespitosa, Geum rivale, Ranunculus repens, ряд бореальных видов – Equisetum sylvaticum, E. pratense, Luzula pilosa и Rubus saxatilis. В ярусе мхов наиболее часто встречаются Atrichum undulatum и Cirriphyllum piliferum.
Группа 4. Березовые травяно-болотные леса. Проективное покрытие яруса A – 63.6 ± 10.0, B – 28.6 ± 15.0, C – 82.5 ± 10.2, D – 32.2 ± 25.9. Древостой сформирован преимущественно Betula pubescens, иногда с примесью Populus tremula, Alnus glutinosa, изредка с участием Padus avium, Salix caprea и S. pentandra. Древостои, как правило, одноярусные, второй ярус, если он развит, образован Picea abies. В ярусе В чаще всего встречаются Frangula alnus и Salix cinerea. В травяном ярусе преобладают Filipendula ulmaria, Calamagrostis canescens, Phragmites australis, а также осоки (Carex acuta, C. appropinquata, C. elongata), образующие кочки, часто встречается Deschampsia cespitosa. Между кочками произрастают Carex vesicaria, Scirpus sylvaticus, Ranunculus repens. Иногда встречаются Comarum palustre, Caltha palustris, Thyselium palustre. В ярусе мхов наиболее обычны Climacium dendroides и Aulacomnium palustre.
Группа 5. Березовые разнотравные леса. Проективное покрытие яруса A – 57.8 ± 20.4, B – 18.7 ± ± 16.4, C – 71.9 ± 18.8, D – 9.0 ± 11.1. Древостои двухъярусные. Первый состоит из Betula pendula и B. pubescens с примесью Populus tremula и Salix caprea. Второй сложен деревьями Betula pendula, B. pubescens, Picea abies, Populus tremula, Salix caprea, редко с участием широколиственных пород. Ярус В развит слабо, но иногда велико покрытие подроста Picea abies, часто встречаются Frangula alnus, Sorbus aucuparia, Padus avium, Corylus avellana. В травяном ярусе по проективному покрытию преобладают виды лугово-опушечной группы: Fragaria vesca, Chamaenerion angustifolium, Veronica chamaedrys, Festuca rubra, Poa pratensis. Часто встречаются Angelica sylvestris, Deschampsia cespitosa, Lysimachia nummularia, Ajuga reptans, Convallaria majalis, Solidago virgaurea, Taraxacum officinale. Моховой покров почти не выражен, наиболее развиты куртинки Atrichum undulatum.
Группа 6. Березовые травяно-кустарничково-сфагновые леса. Проективное покрытие яруса A – 49.0 ± 20.8, B – 31.5 ± 21.9, C – 63.5 ± 16.0, D – 90.5 ± 1.6. Древостой одноярусный, сформирован Betula pubescens, в редких случаях с примесью Pinus sylvestris, Picea abies или Populus tremula. Ярус В развит слабо, его образуют Frangula alnus и подрост Picea abies, Pinus sylvestris, часто встречается Salix cinerea. В травяно-кустарничковом ярусе преобладают болотные кустарнички и травы: Chamaedaphne calyculata, Vaccinium uliginosum, Eriophorum vaginatum, Carex lasiocarpa. В ярусе мхов обычны Polytrichum commune, Aulacomnium palustre, Climacium dendroides, Sphagnum magellanicum и Sphagnum spp.
Группа 7. Осиновые широкотравные леса. Проективное покрытие яруса A – 62.5 ± 18.3, B – 42.8 ± 23.7, C – 64.4 ± 21.1, D – 8.8 ± 7.9. Первый древесный ярус состоит из Populus tremula с примесью Betula pendula и B. pubescens, зачастую также с участием Picea abies, Tilia cordata, Quercus robur. Второй ярус обычно развит, сложен преимущественно деревьями Picea abies, с участием широколиственных видов (в т.ч. Ulmus glabra). Ярус В хорошо развит, его образует Corylus avellana, часто отмечается подрост Picea abies, Quercus robur, Acer platanoides, Tilia cordata. Травяной ярус состоит преимущественно из неморальных трав и отличается полидиминантностью: Aegopodium podagraria, Asarum europaeum, Carex pilosa, Galeobdolon luteum, Galium odoratum, Mercurialis perennis, Pulmonaria obscura. Обычны Ajuga reptans, папоротники (Athyrium filix-femina, Dryopteris filix-mas, D. carthusiana), Paris quadrifolia, Ranunculus cassubicus, Stellaria holostea. В моховом покрове чаще других видов встречаются Atrichum undulatum, Cirriphyllum piliferum, Eurhynchium angustirete, Oxyrrhynchium hians, Plagiomnium cuspidatum, Sciuro-hypnum curtum.
Группа 8. Осиновые влажнотравно-широкотравные леса. Проективное покрытие яруса A – 61.7 ± 21.7, B – 44.3 ± 19.6, C – 76.3 ± 13.8, D – 18.1 ± 15.1. Первый ярус древостоя сформирован Populus tremula со значительной примесью Betula pendula и B. pubescens, изредка – Quercus robur, Picea abies, Alnus incana, A. glutinosa. Второй ярус развит и присутствует почти всегда. Образован, в основном, Picea abies, изредка Padus avium, Acer platanoides, Betula pendula, B. pubescens, Quercus robur, Tilia cordata. Ярус В хорошо развит, в основном образован Corylus avellana, Lonicera xylosteum, иногда встречаются Frangula alnus и Sorbus aucuparia. Обилен подрост Picea abies, подрост других древесных видов (Populus tremula, Quercus robur, Tilia cordata) встречается реже и с меньшим покрытием. В травяном ярусе преобладают Athyrium filix-femina, Crepis paludosa, Filipendula ulmaria, Geum rivale, Pulmonaria obscura. Часты Ajuga reptans, Asarum europaeum, Dryopteris carthusiana, Equisetum pratense, Lysimachia nummularia, L. vulgaris, Paris quadrifolia, Ranunculus cassubicus. В моховом ярусе обычны Atrichum undulatum, Brachythecium rutabulum, Cirriphyllum piliferum, Climacium dendroides, Plagiomnium cuspidatum, Rhizomnium punctatum, Sciuro-hypnum curtum.
Группа 9. Сероольховые влажнотравно-широкотравные леса. Проективное покрытие яруса A – 58.0 ± 14.0, B – 33.2 ± 16.4, C – 94.5 ± 8.5, D – 28.1 ± 19.2. Древостой одноярусный, состоит из Alnus incana с примесью Padus avium. В подросте яруса В представлена в основном ольха серая. Picea abies и широколиственные виды почти всегда отсутствуют. Среди кустарников в ярусе В изредка встречаются Corylus avellana, Lonicera xylosteum, Sambucus racemosa. Травяной ярус состоит из двух подъярусов. Верхний образован Urtica dioica, Campanula latifolia, Filipendula ulmaria, Rubus idaeus. Нижний – Aegopodium podagraria, Chrysosplenium alternifolium, Galeobdolon luteum, Geum rivale, Glechoma hederacea, Lamium maculatum, Mercurialis perennis, Myosoton aquaticum. Моховой ярус сформирован Plagiomnium undulatum, часто встречаются Brachythecium rutabulum, Oxyrrhynchium hians.
Группа 10. Черноольховые влажнотравно-широкотравные леса. Проективное покрытие яруса A – 70.2 ± 14.3, B – 20.7 ± 16.3, C – 84.7 ± 18.3, D – 8.3 ± 8.5. Древостой образован Alnus glutinosa, иногда с примесью Betula pubescens и Picea abies. Другие древесные виды встречаются единично. Второй подъярус, который отмечается редко, образован Alnus glutinosa, Picea abies, редко с примесью Padus avium, Acer platanoides. В ярусе В отмечаются Padus avium, Ribes nigrum, Sorbus aucuparia. В подросте яруса В – Picea abies, Alnus glutinosa. Для травяно-кустарничкового яруса характерна мозаичность, сформированная приствольными повышениями, мочажинами и кочками из Carex elongata. Доминируют Urtica dioica, Filipendula ulmaria, Impatiens noli-tangere. Часто встречаются Athyrium filix-femina, Crepis paludosa и неморальные травы (Milium effusum, Paris quadrifolia, Ranunculus cassubicus). В мочажинах встречаются Scirpus sylvaticus, редко Carex vesicaria и C. riparia. Среди наземных мхов преобладают Brachythecium rutabulum, Climacium dendroides, Plagiomnium cuspidatum.
Группа 11. Черноольховые травяно-болотные леса. Проективное покрытие яруса A – 76.8 ± 12.9, B – 30.3 ± 15.7, C – 92.7 ± 8.1, D – 23.3 ± 13.9. Древостой образован Alnus glutinosa, иногда с примесью Betula pendula и B. pubescens, редко c участием Picea abies, и широколиственных видов деревьев. Ярус В разреженный, в нем чаще всего встречается Padus avium, Ribes nigrum, Salix cinerea, а также Picea abies и Alnus glutinosa. В травяном ярусе выражено два подъяруса. В верхнем преобладают Urtica dioica и Filipendula ulmaria, иногда встречаются Phragmites australis и Calamagrostis canescens, кочкообразующие осоки Carex appropinquata и C. cespitosa. Мочажины между кочками могут быть обводнены, в них часто встречается Scirpus sylvaticus, иногда Carex vesicaria. В нижнем подъярусе наибольшие встречаемость и покрытие отмечены у Lysimachia nummularia и Ranunculus repens. Среди мхов преобладают Climacium dendroides, Aulacomnium palustre, Plagiomnium spp.
Производные и условно-коренные мелколиственные леса распространены в широком диапазоне экологических условий: от дренированных участков водоразделов до переувлажненных понижений (Ниценко, 1972). На графике дерева решений (рис. 1) видно, что основной фактор, дифференцирующий состав мелколиственных лесов (первый узел), – влажность почв. По этому фактору сообщества подразделяются на две большие группы. Более “сухие” объединяют березовые мелкотравно-широкотравные (группа 1), березовые широкотравные (группа 2), осиновые широкотравные (группа 7), березовые разнотравные (группа 5) и часть сероольховых влажнотравно-широкотравных (группа 9) лесов. Другая группа состоит из лесов со значительным увлажнением почв: березовые влажнотравно-широкотравные (группа 3), березовые травяно-болотные (группа 4), березовые травяно-кустарничково-сфагновые (группа 6), осиновые влажнотравно-широкотравные (группа 8), часть сероольховых влажнотравно-широкотравных (группа 9), черноольховые влажнотравно-широкотравные (группа 10) и черноольховые травяно-болотные (группа 11).
Рис. 1.
Дерево решений по экологическим переменным: M – увлажненность почв; L – освещенность; N – богатство почв азотом; R – кислотность почв. Расшифровка номеров групп приведена в таблице.
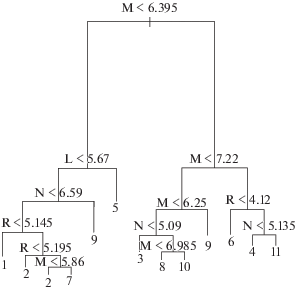
Если рассматривать левую часть графика дерева решений, то видно, что следующим после влажности узлом является разделение по условиям освещения, – при повышенных значениях освещенности выделяется березовая разнотравная группа (группа 5), в остальных группах более сухих лесов освещение меньше. Значимость данного фактора для березняков разнотравных подтверждается результатами ординации – на ординационной схеме они хорошо отделяются как по освещению, так и по пониженным значениям почвенного богатства и по повышенной кислотности почв (рис. 2). В местообитаниях с повышенным богатством почв (узел 3) встречается часть сероольховых лесов (группа 9). На четвертом узле идет разделение по реакции почв: более кислая реакция почв характерна для березовой мелкотравно-широкотравной группы (группа 1). На ординационной схеме (рис. 2) также видно, что для этих лесов характерно меньшее богатство почв, чем для широкотравной группы. Для березовой широкотравной и осиновой широкотравной групп характерна менее кислая реакция почв. При этом осиновые широкотравные леса (группа 7) встречаются в более увлажненных местообитаниях, чем березовые, хотя разделение происходит лишь по шестому узлу. Слабая дифференциация березовой и осиновой широкотравных групп хорошо видна также на ординационной схеме (рис. 2). Эти наиболее распространенные в регионе леса образуют плотное перекрывающееся скопление точек, находясь практически в одном экологическом пространстве. Для них характерны не переувлажненные достаточно богатые местообитания со средними значениями кислотности почв, а их подчиненные ярусы плохо освещены.
Рис. 2.
NMDS ординация сообществ. Расшифровка номеров групп дана в таблице, обозначение экологических факторов приведено на рис. 1.
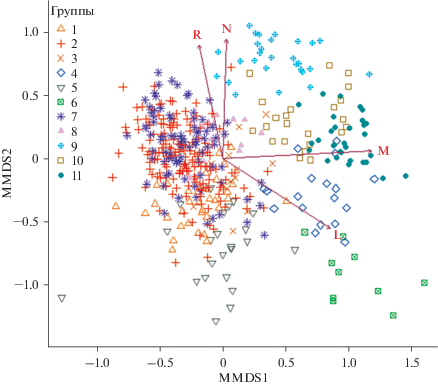
В правой части дерева в составе сообществ более влажных условий местообитаний второй узел также отвечает за влажность, таким образом, разделяя “влажные” (левая часть) и переувлажненные леса (правая часть) (рис. 1). Среди последних березовая кустарничково-травяно-сфагновая группа (группа 6) распространена в местообитаниях с более кислой реакцией почв (узел 3), также для нее характерна высокая освещенность подчиненных ярусов (рис. 2). Березовую и черноольховую травяно-болотные группы дифференцирует богатство почв (узел 4): чернольховая травяно-болотная (группа 11) встречается в чуть более богатых местообитаниях, чем березовая (группа 4). На схеме ординации (рис. 2) заметно также, что данные сообщества березовой формации встречаются в местообитаниях с более кислой реакцией почв.
Группу “влажных” лесов можно назвать влажнотравно-широкотравной, поскольку сюда входят 4 формации со сходным характером травяно-кустарничкового яруса (влажнотравно-широкотравные леса). Их первое разделение происходит по фактору богатства почв (узел 3), – при повышенном богатстве встречаются сероольховые леса (группа 9) (часть сообществ, не попавших в левый кластер более сухих лесов). Данные леса характеризуются максимальным богатством почв, что хорошо видно на ординации (рис. 2). Следующее разделение (узел 4) происходит также по богатству почв: в самых бедных местообитаниях отмечаются березовые влажнотравно-широкотравные леса. Осиновые (группа 8) и черноольховые (группа 10) влажнотравно-широкотравные леса дифференцирует влажность – в чуть более влажных местообитаниях отмечаются черноольховые сообщества.
В зависимости от конкретных экологических режимов местообитаний в различных типах сообществ участие основных коренных лесообразующих видов деревьев (ель, сосна, дуб, липа) варьирует в широких пределах. Оценка связи эдификаторных древесных видов с основными факторами среды позволяет оценить текущее их распределение и перспективы возобновления.
Наибольшее участие ели отмечается в оптимуме условий существования мелколиственных лесов со смещением в сторону чуть более кислой реакции почв. В наиболее экстремальных экологических условиях по режимам увлажнения и питания участие ели снижается (рис. 3). По нашим данным, участие сосны в большинстве групп не отмечается вовсе. Однако в условиях снижения конкуренции в неблагоприятных режимах питания и переувлажненности и лучшего освещения сосна начинает встречаться во всех ярусах. Это заметно на примере березовой кустарничково-травяно-сфагновой группы, где древостой сильно разрежен (рис. 3).
Рис. 3.
Распределение в экологическом пространстве мелколиственных лесов основных лесообразующих видов деревьев в составе всех ярусов, а – Picea abies; б – Pinus sylvestris; в – Tilia cordata и г – Quercus robur.
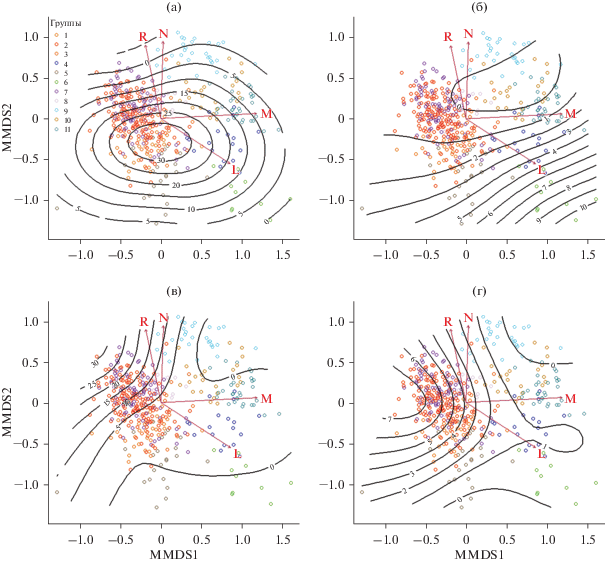
В лесном покрове региона отмечено большое число широколиственных видов деревьев (клен, ясень (Fraxinus excelsior), дуб, липа, вязы (Ulmus glabra и Ulmus laevis) и др.), однако наибольшее участие в сообществах у липы и дуба. Максимальная примесь липы отмечена в широкотравных группах (березовой и осиновой) (рис. 3), где почвенное богатство достаточно высоко и отсутствует переувлажнение, а высокое затенение не является лимитирующим фактором для ее возобновления. По нашим данным, участие дуба в мелколиственных сообществах ниже, чем липы. В целом наличие его примеси и возобновления увеличивается по мере уменьшения влажности, но меньше, в отличие от липы, зависит от богатства почв (рис. 3).
Различия в участии коренных лесообразующих видов деревьев (ели, сосны, липы и дуба) в разных ярусах в производных сообществах (березовых и осиновых) (группы 1–8) наглядно отслеживаются на графике дерева решений (рис. 4). На первом этапе происходит разделение сообществ по участию широколиственных видов деревьев в древостое. Правая часть дерева включает сообщества с более богатыми местообитаниями: широкотравные группы (осиновую и березовую), а также осиновую влажнотравно-широкотравную. Следующий узел в правой части дерева дифференцирует сообщества по участию широколиственных видов деревьев в составе яруса С. Их участие больше в широкотравных группах (2, 7). В древесном ярусе в группе березовых широкотравных лесов (2) участие широколиственных видов может быть как выше 4%, так и ниже, а в осиновой широкотравной оно всегда превышает 4% (узел 3). В группе осиновых широкотравных лесов (узел 4) еще и участие ели в ярусе В выше, чем в березовой. Для части сообществ осиновой широкотравной группы (7) характерно меньшее участие широколиственных пород в ярусе С, но при этом более высокое участие этих видов, а также ели в ярусе В. В осиновой влажнотравно-широкотравной группе (8) ниже участие как широколиственных видов, так и ели.
Рис. 4.
Дерево решений по проективному покрытию Picea abies, Pinus sylvestris и основных широколиственных видов (Tilia cordata, Quercus robur) в разных ярусах производных мелколиственных лесов: A – древостой; B – ярус В (крупный подрост); С – травяно-кустарничковый ярус (мелкий подрост). Расшифровка номеров групп приведена в таблице.
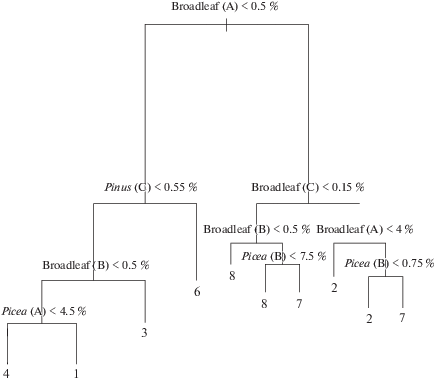
В левой части дерева в сообществах с меньшим участием широколиственных видов деревьев за следующий узел разделения сообществ отвечает доля сосны в ярусе С. Ее участие отмечено в березовой травяно-кустарничково-сфагновой группе (6) и однозначно ее отделяет от остальных. Следующий узел (узел 3) отделяет березовую влажнотравно-широкотравную группу по признаку более высокого участия широколиственных пород в ярусе В. При условии более низкого участия (узел 4) в березовой травяно-болотной группе (4) наблюдается более низкое участие ели в древостое, в то время как в березовой мелкотравно-широкотравной группе оно самое высокое из всех рассмотренных групп.
Таким образом, в березовой влажнотравно-широкотравной группе наблюдается наименьшее возобновление коренных пород, в то время как в осиновой широкотравной оно наибольшее. Березовая травяно-кустарничково-сфагновая группа является единственной, где довольно часто отмечается сосна. Березовые разнотравные леса (группа 5) не могут быть выделены в самостоятельную группу вследствие очень низкой выраженности подроста в целом и приблизительно равного участия видов в нем. Среди производных мелколиственных лесов осиновые леса (две группы) и березовые широкотравные сообщества отличаются более высоким участием широколиственных видов в древостое, в то время как березовая мелкотравно-широкотравная выделяется по высокому участию ели в древостое.
Состав и обилие подроста позволяют оценить перспективы развития лесов. Если рассматривать все группы лесов по участию в ярусе В основных ранне- и позднесукцессионных видов, то заметно, что практически во всех группах ель составляет большую часть подроста (рис. 5). Участие ели в подросте максимально в березовой мелкотравно-широкотравной группе (1) (15%), которая является самой бореальной по составу подчиненных ярусов; здесь редко встречаются другие виды деревьев (рис. 3, 4). В березовой широкотравно-влажнотравной группе (3) ель также составляет практически весь подрост, хотя он характеризуется меньшей величиной проективного покрытия (8%). Также очень высоко участие ели в подросте в березовой травяно-болотной группе (4) (14%). Ель составляет менее 1% только в сероольховой влажнотравно-широкотравной группе (9), где практически весь подрост представлен ольхой серой. Также достаточно низкое участие ели отмечается в березовой разнотравной группе (5), однако в данных сообществах общее проективные покрытие подроста в ярусе В всего около 5% в среднем – самое слабое возобновление деревьев среди всех мелколиственных лесов.
Рис. 5.
Среднее проективное покрытие основных древесных видов в ярусе B в группах ассоциаций мелколиственных лесов. Расшифровка номеров групп приведена в табл. 1.
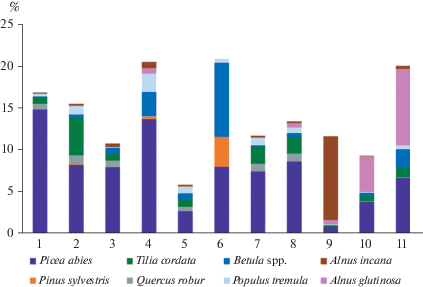
Из широколиственных пород в регионе основной является липа (Курнаев, 1968), больше всего ее в березовой (2) и осиновой широкотравной (7), а также в осиновой влажнотравно-широкотравной (8) группах (5%, 2% и 2% соответственно). В составе подроста этих трех групп все же преобладает ель, однако в березовой широкотравной группе соотношение ели и липы близко (8% и 5% соответственно). Дуб встречается практически во всех сообществах, однако его доля в подросте невелика и в среднем составляет лишь 1% и менее. Березовые кустарничково-травяно-сфагновые (группа 6) леса отличаются от остальных значительным участием в подросте сосны (4%) и березы (9%), хотя ель и здесь достаточно заметна (8%). В черноольховых группах (10, 11) в подросте в основном участвуют ель и черная ольха, причем в черноольховой травяно-болотной группе проективное покрытие этих видов выше (7% и 9% соответственно), чем во влажнотравно-широкотравной (по 4%). Береза, помимо кустарничково-травяно-сфагновых лесов, заметна в травяно-болотных лесах – березовых (группа 4) и черноольховых (группа 11), где ее проективные покрытие составляет чуть менее 3%.
ОБСУЖДЕНИЕ
Экологические факторы. В зависимости от географического охвата исследования и рассматриваемого набора сообществ на первый план выходят разные экологические факторы, в первую очередь влияющие на видовой состав лесов. Для относительно однородных по составу березовых бореальных лесов Норвегии (Bakkestuen et al., 2010) основным фактором, влияющим на дифференциацию состава сообществ, является изменение значений pH, а также концентрации в почве кальция, калия и серы. Для черноольховых лесов северо-запада России – увлажненность почв (Василевич, Щукина, 2001), а черноольховых лесов Словакии – помимо увлажненности еще и почвенное богатство (Slezák et al., 2011).
Основным экологическим фактором, влияющим на состав мелколиственных лесов центральной части Русской равнины, является влажность – наиболее контрастный из всех рассмотренных факторов. Перераспределение влаги является функцией рельефа. Мелколиственные леса могут произрастать в диапазоне от заболоченных местообитаний в понижениях и долинных элементах рельефа (березовые кустарничково-травяно-сфагновые леса, черноольховые травяно-болотные и др.) до наиболее сухих и дренированных - на моренных холмах и склонах. По увлажненности почв строго дифференцируются мелкотравно-широкотравные, широкотравные и разнотравные леса (как менее увлажненные) и влажнотравно-широкотравные, травяно-болотные и кустарничково-травяно-сфагновые сообщества (как более влажные). Единственной группой, в которой наблюдаются значительные различия по степени увлажнения, является сероольховая влажнотравно-широкотравная. Т.А. Работнов (1939) отмечает, что для овражно-долинных сероольховых лесов наблюдаются различия увлажнения и, соответственно, состава и доминантов травяного яруса в связи с положением сообщества в рельефе – близостью русла, выровненностью рельефа или склоновым положением. По мере увеличения увлажнения он выделяет леса с доминированием крапивы, крапивы и таволги и таволги. По нашим данным, также отмечается изменение участия более влаголюбивых видов в сообществах этой группы. Так, для части лесов, которые располагаются в более дренированных склоновых местообитаниях, характерен более неморальный состав подчиненных ярусов с доминированием Urtica dioica, Aegopodium podagraria, Galeobdolon luteum, Stellaria holostea. В местообитаниях с худшими условиями дренажа (например, выровненные поймы) существуют леса с доминированием Filipendula ulmaria, Urtica dioica, Geum rivale, значительным участием Chrysosplenium alternifolium, Lysimachia nummularia, Ranunculus repens, иногда Scirpus sylvaticus. Крапива является одним из доминантов в обоих вариантах сероольховых лесов. Участие более влаголюбивых видов увеличивается плавно, и четко разделить сообщества на две группы невозможно, к тому же большинство видов отмечается во всей группе сероольховых лесов, только с разным соотношением долевого участия.
Возобновление коренных видов деревьев и перспективы развития производных лесов. О перспективах развития производных сообществ можно судить по составу и состоянию подроста. Поселение тех или иных коренных пород деревьев под пологом леса при отсутствии нарушений зависит как от условий местообитаний, так и от возможности заноса семян (Korotkov et al., 2001). В результате при различном спектре видов в подросте прогнозируются и разные по длительности сукцессионные стадии, которые зависят от продолжительности жизни деревьев, выходящих в основной полог (Korotkov et al., 2001). По нашим данным, ель и липа наиболее активно участвуют в составе производных мелколиственных лесов, а участие дуба во всех группах примерно равное и небольшое (не более 1%). Липа чаще отмечается в широкотравных группах сообществ, где для нее благоприятны условия почвенного богатства и дренажа, а ее теневыносливость позволяет возобновляться при слабом освещении. Доля ели еще выше по сравнению с липой, наибольшее ее участие наблюдается в экологическом оптимуме мелколиственных лесов с небольшим смещением в сторону меньшего почвенного богатства и повышенной кислотности (березовые мелкотравно-широкотравные леса). Высокое участие ели может объясняться не только ее естественным возобновлением, но и созданием посадок. Из-за ненадлежащего ухода за культурами посадки еще на ранних этапах развития зарастают мелколиственными видами (особенно березой) и формируются производные мелколиственные леса с участием ели (Chernenkova et al., 2020). Так, треть лесных массивов березовой широкотравной группы в юго-западном Подмосковье, по нашим данным, составляют культуры ели и сосны, заросшие березой. Подрост дуба во всех производных сообществах (за исключением березовых кустарничково-травяно-сфагновых лесов) встречается примерно с одинаковым покрытием (около 1%). Таким образом, дуб, во-первых, менее “активен”, чем липа, в мелколиственных лесах (что может в некоторой степени подтверждать точку зрения С.Ф Курнаева о коренных липово-еловых лесах региона), а во-вторых, отсутствие варьирования значений его покрытий свидетельствует о меньшей требовательности к богатству почв по сравнению с липой (рис. 3). Клен – еще одна часто отмечаемая широколиственная порода, участие которой в сообществах в последние десятилетия увеличивается (Маслов, 2012). Однако клен не формирует древостои и встречается только в виде примеси. Вязы (вяз голый и вяз гладкий) участвуют в составе лесов довольно редко, хотя и формируют небольшие массивы в богатых и увлаженных местообитаниях.
Березовые (группа 2) и осиновые (группа 7) широкотравные, а также осиновые влажнотравно-широкотравные (группа 8) леса характеризуются максимальным возобновлением липы среди всех типов мелколиственных сообществ региона, они распространены в местообитаниях с высоким богатством почв и достаточным дренажем (за исключением осиновой влажнотравно-широкотравной группы). Также эти три группы лесов отличаются от остальных по более высокому участию широколиственных видов в древостое (рис. 5).
Сообщества березовой мелкотравно-широкотравной группы (1) отличаются значительным участием бореальных видов в подчиненных ярусах и абсолютным преобладанием ели в подросте. Для данных сообществ прогнозируется развитие в сторону бореальных и бореально-неморальных ельников.
Березовые влажнотравно-широкотравные (группа 3) и березовые травяно-болотные леса (группа 4) обычно встречаются небольшими массивами в понижениях, слабопроточных ложбинах разной степени переувлажненности. Здесь также в подросте преобладает ель. Широколиственные виды не отмечены в связи с высоким увлажнением местообитаний данных сообществ. Вероятно, коренными сообществами здесь являлись еловые леса с большим количеством влаголюбивых видов.
Березовые кустарничково-травяно-сфагновые леса (группа 6) характеризуются участием в подросте березы наравне с елью, а также только здесь постоянно отмечается сосна. Участие сосны связано с отсутствием конкуренции со стороны других видов деревьев из-за заболоченности местообитаний и хорошими условиями освещенности под разреженным пологом древостоя. Здесь ожидается смена на сосновые и еловые сфагновые и долгомошные леса, однако значительное участие березы в подросте может замедлять смену лесообразующих пород.
Березовые разнотравные леса (группа 5) с высоким покрытием видов лугово-опушечной группы являются наиболее нарушенными лесами, которые могут развиваться в мезофильных условиях при значительной рекреационной нагрузке, а в прошлом и хозяйственной деятельности (сенокосы, выпас) и обычно встречаются на периферии лесных массивов. В связи с этим, подрост деревьев здесь выражен слабо, и дальнейшее развитие этих сообществ зависит от условий местообитаний и заноса семян. Вероятно, сообщества этой группы дольше остальных будут восстанавливаться до условно-коренных лесов.
Таким образом, широкотравные (березовые и осиновые) и осиновые влажнотравно-широкотравные леса потенциально наиболее быстро по сравнению с другими группами производных мелколиственных лесов могут смениться условно-коренными липово-еловыми лесами. Березовые мелкотравно-широкотравные, влажнотравно-широкотравные и травяно-болотные леса скорее будут заменены еловыми лесами. Однако для прогнозирования смены производных лесов условно-коренными и ее скорости необходима также оценка жизненного состояния подроста. В случае, если оно не удовлетворительное, – смена состава верхнего полога может значительно замедлиться.
Сукцессионный статус групп сообществ мелколиственных лесов. Большинство групп сообществ березовых лесов (березовые мелкотравно-широкотравные, березовые широкотравные, березовые влажнотравно-широкотравные, березовые разнотравные), а также все осиновые (осиновые широкотравные, осиновые влажнотравно-широкотравные) являются однозначно производными, возникшими на месте хвойных, хвойно-широколиственных или широколиственных лесов. Происхождение части сообществ с участием березы не столь очевидно. С одной стороны, существует точка зрения (Ниценко, 1972; Абатуров и др., 1982), что все березовые леса являются производными, при этом березовые кустарничково-травяно-сфагновые возникли на месте долгомошных и сфагновых хвойных лесов (Ниценко, 1972; Абатуров и др., 1982), а также выгоревших переходных болот. С другой стороны, С.Ф. Курнаев (1968) отмечал коренной характер березняков из березы пушистой в замкнутых понижениях (травяно-болотные сообщества). По результатам наших исследований, в большинстве сообществ березовых лесов преобладает подрост не березы, а других видов (преимущественно ели), хотя на бедных почвах и при застойном увлажнении ель с возрастом может выпадать. Исключение составляет только березовая кустарничково-травяно-сфагновая группа, где в подросте наряду с елью и сосной достаточно много березы (проективное покрытие 9%), а также березовая травяно-болотная, в которой проективное покрытие подроста мелколиственных видов составляет 5%.
Черноольховые леса, распространенные по долинам водотоков и замкнутым понижениям (влажнотравно-широкотравные и травяно-болотные), мы рассматриваем как условно коренные. В данных лесах в подросте ель и ольха черная возобновляются примерно в равных долях, однако ель, по-видимому, не может успешно существовать в экологическом режиме черноольховых лесов, так как она редко выходит в первый ярус. Сероольховые леса в зависимости от местообитания могут быть как производными, так и коренными. Иногда они являются пионерными сообществами на месте антропогенных нарушений (Ликсакова, 2004). Однако гигрофитные неморальные леса (влажнотравно-широкотравные в рамках принятой классификации) считаются условно-коренными (Работнов, 1939; Коротков, Морозова, 1986; Ellenberg et al., 1992). В сероольховых влажнотравно-широкотравных лесах в подросте отмечается преобладание и высокое участие ольхи серой, другие виды деревьев отмечаются гораздо реже. Основываясь на этом, можно подтвердить условно-коренной характер данных сообществ.
ВЫВОДЫ
1. Мелколиственные леса центральной части Русской равнины разнообразны по типологическому составу. В рамках принятого эколого-фитоценотического подхода выделено 11 групп ассоциаций мелколиственных лесов, относящихся к 4 формациям.
2. Мелколиственные леса произрастают в широком диапазоне экологических условий хвойно-широколиственной зоны. При этом основным фактором, влияющим на варьирование видового состава мелколиственных сообществ, является увлажненность почв. По этому фактору четко разделяются, с одной стороны, более сухие мелкотравно-широкотравные, широкотравные и разнотравные леса, а с другой – влажнотравно-широкотравные, травяно-болотные и кустарничково-травяно-сфагновые сообщества.
3. Леса из ольхи серой и ольхи черной могут быть отнесены к условно-коренным. В сероольховых влажнотравно-широкотравных сообществах основной породой подроста является серая ольха, что подтверждает мнение некоторых исследователей о коренном статусе этих сообществ. В черноольховых лесах подрост черной ольхи и ели находится примерно в равных пропорциях, однако ель редко выходит в первый ярус.
4. Мелколиственные леса из березы и осины создают условия для восстановления условно-коренных сообществ. Ель является основной породой, которая возобновляется в мелколиственных лесах региона. Она отмечается во всех типах сообществ, причем максимально ее участие в мелкотравно-широкотравных березовых лесах. Отметим, что создание культур могло значительно увеличить участие ели в мелколиственных лесах.
5. В березняках травяно-кустарничково-сфагновых отмечается наибольшая освещенность под пологом древостоя, и наблюдается максимальное участие березы в ярусе B. По этой же причине, а также из-за отсутствия конкуренции сосна встречается здесь во всех ярусах.
6. Березовые и осиновые широкотравные, а также осиновые влажнотравно-широкотравные сообщества, вероятно, скорее остальных сменятся условно-коренными елово-липовыми лесами.
Благодарности. Для хранения и анализа материалов полевых геоботанических описаний использована база данных “FORDIV” (Св. о гос. регистрации № 2014620979). Авторы благодарят коллег, принявших участие в ее наполнении (М.В. Архипову, Н.Г. Кадетова, С.Ю. Попова, Е.В. Тихонову и многих других), а также выражают искреннюю признательность Природоохранному Фонду “Верховье” за многолетнее сотрудничество и предоставленные данные.
Список литературы
Абатуров А.В. Из истории лесов Подмосковья / Динамика хвойных лесов Подмосковья. М.: Наука, 2000. С. 22–32.
Абатуров Ю.Д., Зворыкина К.В., Ильюшенко А.Ф. Типы березовых лесов центральной части южной тайги. М.: Наука, 1982. 155 с.
Алехин В.В. Растительность СССР в основных зонах. М.: Советская наука, 1951. 483 с.
Беляева Н.Г., Попов С.Ю. Изменение лесистости бывшего Верейского уезда Московской области за последние 200 лет // Лесоведение. 2016. № 1. С. 44–54.
Бибикова Т.В. Классификация осиновых лесов северо-запада России // Ботанический журнал. 1998. Т. 83. № 3. С. 48–57.
Василевич В.И. Незаболоченные березовые леса северо-запада Европейской России // Ботанический журнал. 1996. Т. 81. № 11. С. 1–13.
Василевич В.И. Сероольшатники Европейской России // Ботанический журн. 1998. Т. 83. № 8. С. 28–42.
Василевич В.И., Щукина К.В. Черноольховые леса северо-запада Европейской России // Ботанический журнал. 2001. Т. 86. № 3. С. 15–26.
Ветчинникова Л.В. Береза: вопросы изменчивости (морфо-физиологические и биохимические аспекты). М.: Наука, 2004. 183 с.
Грибова С.А., Исаченко Т.И., Лавренко Е.М. Растительность Европейской части СССР. М.: Наука, 1980. 236 с.
Гульбе Я.И., Гульбе А.Я., Гульбе Т.А., Ермолова Л.С. Динамика биологической продуктивности молодняков березы на залежи // Актуальные проблемы лесного комплекса. 2016. № 46. С. 12–16. URL: https://cyberleninka.ru/article/n/dinamika-biologicheskoy-produktivnosti-molodnyakov-berezy-na-zalezhi
Емельянова Л.Г., Огуреева Г.Н. Биогеографическое картографирование. М.: Географический факультет МГУ, 2006. 132 с.
Ермаков Н.Б. Продромус высших единиц растительности России / Современное состояние основных концепций науки о растительности. Уфа: Гилем, 2012. С. 377–483.
Игнатов М.С, Игнатова Е.А. Флора мхов средней части Европейской России. M.: KMK. 2003. Т. 1–2. 960 с.
Коротков К.О., Морозова О.В. Класс Querco–Fagetea. Леса Валдайского лесничества // Классификация растительности СССР с использованием флористических критериев. М.: МГУ, 1986. С. 121–133.
Курнаев С.Ф. Лесорастительное районирование СССР. М.: Наука, 1973. 203 с.
Курнаев С.Ф. Основные типы леса средней части Русской равнины. М.: Наука, 1968. 356 с.
Лесной план Московской области. Федеральное агентство лесного хозяйства, Федеральное государственное бюджетное учреждение “Рослесинфорг”, Филиал ФГБУ “Рослесинфорг” “Мослеспроект”. М., 2018. Кн. 1. 83 с.
Ликсакова Н.С. Мелколиственные леса Чудовского района Новгородской области // Ботанический журн. 2004. Т. 89. № 8. С. 1319–1342.
Маслов А.А. Флуктуации и сукцессии в лесных сообществах на фоне изменения климата // Известия Самарского научного центра РАН. 2012. Т. 14. № 1–5. С. 1316–1319.
Маслов А.А., Баранов О.Ю., Сирин А.А. Идентификация видов берез в заболоченных лесах центра русской равнины по результатам молекулярно-генетического анализа // Лесоведение. 2019. № 3. С. 177–187.
Милов Л.В. Великорусский пахарь и особенности российского исторического процесса. М.: РОССПЭН, 1998. 568 с.
Нешатаев В.Ю. Проект Всероссийского кодекса фитоценологической номенклатуры // Растительность России. 2001. № 1. С. 62–70.
Ниценко А.А. Типология мелколиственных лесов Европейской части СССР. Л.: ЛГУ, 1972. 140 с.
Огуреева Г.Н., Булдакова Е.В., Архипова М.В. Региональный анализ биоразнообразия лесов Европейской части России / Биогеография в Московском университете. 60 лет кафедре биогеографии. М.: ГЕОС. 2008. С. 36–51.
Работнов Т.А. Типы сероольховых насаждений северо-западной части Московской области // Ботанический журн. 1939. № 1. С. 15–29.
Разумовский С.М. Труды по экологии и биогеографии (полное собрание сочинений). М.: КМК, 2011. 722 с.
Рахилин В.К. Леса Московского региона в XVIII веке / История изучения, использования и охраны природных ресурсов Москвы и Московского региона. М.: Янус-К. 1997. С. 120–124.
Рысин Л.П., Савельева Л.И. Кадастры типов леса и типов лесных биогеоценозов. М.: КМК. 2007. 144 с.
Смирнов В.Э., Ханина Л.Г., Бобровский М.В. Обоснование системы эколого-ценотических групп видов растений лесной зоны Европейской России на основе экологических шкал, геоботанических описаний и статистического анализа // Бюллетень МОИП. Серия Биологическая. 2006. Т. 111. № 2. С. 36–47.
Сукачев В.Н. Что такое фитоценоз? // Советская ботаника. 1934. № 5. С. 4–18.
Черепанов С.К. Сосудистые растения России и сопредельных государств. СПб.: Мир и семья, 1995. 992 с.
Черненькова Т.В., Морозова О.В. Классификация и картографирование ценотического разнообразия лесов // Лесоведение. 2017. № 4. С. 243–255.
Atkinson M.D. Betula pendula Roth (B. verrucosa Ehrh.) and B. pubescens Ehrh // The J. Ecology. 1992. V. 80. № 4. P. 837–870. https://doi.org/10.2307/2260870
Bakkestuen V., Aarrestad P.A., Stabbetorp O.E., Erikstad L., Eilertsen O. Vegetation composition, gradients and environment relationships of birch forest in six reference areas in Norway // Sommerfeltia. 2010. № 34. P. 1–223.
Blumenthal B. Studies into the occurrence and properties of aspen in Finland (Studied angående aspens förekomst och egenskaper i Finland) // Silva Fennica. 1942. № 56. P. 1–63.
Brandtberg P.-O., Lundkvist H., Bengtsson J. Changes in forest-floor chemistry caused by a birch admixture in Norway spruce stands // Forest Ecology and Management. 2000. V. 130. № 1–3. P. 253–264.
Chernenkova T.V., Kotlov I.P., Belyaeva N.G., Suslova E.G., Morozova O.V., Pesterova O.A., Arkhipova M.V. Role of Silviculture in the Formation of Norway Spruce Forests along the Southern Edge of Their Range in the Central Russian Plain // Forests. 2020. V. 11. № 7. 778 p.
Ellenberg H., Weber H.E., Düll R., Wirth V., Werner W., Paulissen D. Zeigerwerte von Pflanzen in Mitteleuropa // Scripta Geobotanica. 1991. № 18. P. 1–248.
Ellenberg H., Weber H.E., Dull R., Wirth V., Werner W., Paulssen D. Zeigerwerte von Pflanzen in Mitteleuropa. 2 Aufl. Göttingen: Verlag Erich Goltze GmbH & Co KG, 1992. 258 p.
Ferm A. Birch production and utilization for energy // Biomass and Bioenergy. 1993. V. 4. № 6. P. 391–404.
Korotkov V.N., Logofet D.O., Loreau M. Succession in mixed boreal forest of Russia: Markov models and non-Markov effects // Ecological Modelling. 2001. V. 142. № 1–2. P. 25–38. https://doi.org/10.1016/s0304-3800(01)00247-2
Mucina L., Grabherr G., Wallnöfer S., Geisselbercht L., Grass V., Gutermann W., Justin Ch., Wirth J. M. Die Pflanzengesellschaften Österrreichs. Teil III. Wälder und Gebüsche. Jena, 1993. 356 p.
Organisation for Economic Co-operation and Development (OECD). Consensus Document on the Biology of European White Birch (Betula pendula Roth). 2003. № 28. URL: http://www.olis.oecd.org/olis/2003doc.nsf/LinkTo/env-jm-mono(2003)12
Perala D.A., Alm A.A. Reproductive ecology of birch: a review // Forest Ecology and Management. 1990. V. 32. № 1. P. 1–38.
Possen B.J.H.M., Oksanen E., Rousi M., Ruhanen H., Ahonen V., Tervahauta A., Heinonen J., Heiskanen J., Kärenlampi S., Vapaavuori E. Adaptability of birch (Betula pendula Roth) and aspen (Populus tremula L.) genotypes to different soil moisture conditions // Forest Ecology and Management. 2011. V. 262. № 8. P. 1387–1399. https://doi.org/10.1016/j.foreco.2011.06.035
Potapov P.V., Turubanova S.A., Tyukavina A., Krylov A.M., McCarty J.L., Radeloff V.C., Hansen M.C. Eastern Europe’s forest cover dynamics from 1985 to 2012 quantified from the full Landsat archive // Remote Sensing of Environment. 2015. V. 159. P. 28–43.
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. 2019. URL: https://www.R-project.org/
Slezák M., Hrivnák R., Petrášová A. Syntaxonomy and ecology of black alder vegetation in the southern part of central Slovakia // Hacquetia. 2011. V. 10. № 2. P. 119–136.
Tichý L. JUICE, software for vegetation classification // J. Vegetation Science. 2002. V. 13. P. 451–453.
Дополнительные материалы отсутствуют.
Инструменты
Лесоведение