

Лесоведение, 2022, № 4, стр. 432-448
Продуктивность древесного яруса на верховых болотах в таежной зоне Западной Сибири
Н. Г. Коронатова a, *, Н. П. Косых a
a Институт почвоведения и агрохимии СО РАН
630090 Новосибирск, просп. Академика Лаврентьева, 8/2, Россия
* E-mail: koronatova@issa-siberia.ru
Поступила в редакцию 15.10.2019
После доработки 09.01.2020
Принята к публикации 06.04.2022
- EDN: ELOWBB
- DOI: 10.31857/S0024114822040052
Аннотация
Проведена количественная оценка фракций живой и мертвой органической массы древесного яруса верховых болот таежной зоны Западной Сибири, где доминирует болотная форма сосны обыкновенной Pinus sylvestris L. f. litwinowii с примесью сосны кедровой P. sibirica Du Tour. В результате дискриминантного анализа установлено значимое различие между модельными деревьями из южной, средней и северной тайги. Отдельно для каждой из таежных подзон получены полиномиальные уравнения регрессии, аппроксимирующие зависимости фракций биомассы от диаметра ствола. Густота древесного яруса в болотах линейно уменьшалась к северу от 20.3 ± 6.6 до 3.5 ± 0.9 тыс. дер. га–1, что сопровождалось снижением суммарной фитомассы от 1057 до 333 г м–2, в то время как максимальный запас сухостоя и сухих ветвей отмечен в средней тайге. Максимальная масса корней в толще 0-30 см была в средней тайге, а минимальная – в северной. Характерно увеличение запаса корней сосен с глубиной в южной и средней тайге и снижение – в северной. Показано изменение структуры фитомассы и соотношения фракций вдоль широтного градиента. Надземная первичная продукция древесного яруса в южной, средней и северной тайге оценена в 70.7, 27.3 и 4.7 г м–2 год–1 соответственно, с максимальным вкладом кедровой сосны в центре таежной зоны, не превышающим 5%. Вклад древесного яруса в общий запас фитомассы верховых болот уменьшался от 28% на юге территории до 9% на севере.
Торфяные болота широко распространены на территории Западной Сибири, где они занимают 26% площади (Романова, 1985), или 1000330 км2 (Yefremov, Yefremova, 2001), что составляет около 30% от всех торфяных болот умеренных широт (Moore, 2002). Поэтому болота данной территории играют существенную роль в формировании регионального, в том числе почвенного климата, регулировании речного стока, газовом обмене с атмосферой, а также вносят значимый вклад в глобальный круговорот углерода.
Преобладающим типом болот в Западной Сибири являются верховые, наиболее широко распространенные в таежной зоне, где занимают по разным оценкам от 216 000 км2 (Вомперский и др., 2005) до 336 000 км2 (Романова, 1985) и 416 000 км2 (Yefremov, Yefremova, 2001). При этом доля облесенных болот составляет в таежных подзонах от 50 до 58%, снижаясь до 39% в подтайге Западной Сибири (Вомперский и др., 2011).
Основной древесной породой верховых болот севера Европы и Сибири является сосна обыкновенная Pinus sylvestris L., что отличает евразийские облесенные болота бореальной зоны от североамериканских, где преобладают лиственница американская Larix laricina (Du Roi) и ель черная Picea mariana (Mill.) Britt., Sterns&Pegg (Vitt, 1994). Сосновые ценозы болот Северной Евразии приурочены к наиболее дренированным участкам (Вомперский, Глухов, 1982). Сосны на верховых болотах характеризуются низкорослостью в связи с высоким уровнем стояния болотных вод, бедностью олиготрофных торфяных почв (Пьявченко, 1967; Биогеоценологическое …, 1982; Глебов, 1988) и недостаточным развитием опорной функции корней в рыхлом сфагновом очесе (Панов, 2007), и при высоте менее 10 м не бонитируются (Глебов, 1988). Высокая вариабельность среды на болотах приводит к внутривидовому формообразованию у деревьев (Минаева, Сирин, 2011). В зависимости от линейных размеров и характера кроны выделяют несколько форм болотных сосен: P. sylvestris L. f. uliginosa Abol. высотой 10–12 м и кроной в верхней трети ствола; P. sylvestris L. f. litwinowii Sukacz. высотой 2–4 м и кроной в верхней части ствола; P. sylvestris L. f. willkommii Sukacz., имеющая вид деревца с обильным ветвлением от основания высотой 1–3 м; P. sylvestris L. f. pumila Abol., имеющая вид куста с обильным ветвлением от основания высотой до 1.5 м (Сукачев, 1973). Считается, что f. willkommii может быть как самостоятельной формой, так и переходить в f. pumila (Панов, 2007) либо быть подростом f. litwinowii (Лешок, Дыренков, 1988). Часто в качестве примеси к сосне обыкновенной на болотах встречается сосна сибирская кедровая Pinus sibirica Du Tour, которая в условиях олиготрофности и обводненности также приобретает специфические болотные формы (Храмов, Валуцкий, 1977; Велисевич, Чернова, 2014).
Согласно базе данных В.А. Усольцева (2007) о лесах Евразии, сведения о западносибирских болотных сосняках имеются главным образом для отдельных районов южнотаежной подзоны. Здесь параметры их фитомассы и продукции изучали Н.И. Пьявченко (1967), А.А. Храмов, В.И. Валуцкий (1977), Ф.З. Глебов (1988), С.П. Ефремов с соавт. (2005), Т.Т. Ефремова с соавт. (2008), Н.Г. Коронатова и Е.В. Миляева (2011), Е.А. Головацкая (2017). В более северных регионах данной территории такие исследования единичны (Махатков и др., 2007; Косых, Махатков, 2008).
Несмотря на значительную долю облесенных болот в Западной Сибири, при изучении круговорота углерода древесный ярус редко попадает в поле зрения исследователей по следующим причинам. Болотоведы-биогеоценологи сосредоточены на изучении основных продуцентов и эдификаторов – сфагновых мхов, а также травяно-кустарничкового яруса и используют методики, которые неприменимы для древесного яруса. Изучение запасов биомассы древесного яруса сфагновых болот требует модификации подходов, принятых в лесоведении, в связи с миниатюрностью болотных сосен. Хотя облесенные болота могут учитываться как леса (Торфяные болота …, 2001), низкопродуктивные сосновые ценозы верховых болот обычно не представляют интереса для исследователей. Малодоступность западносибирских болот также обусловливает их меньшую изученность по сравнению с аналогичными экосистемами Северной Европы.
Цель данной работы – получение количественных оценок фракций биомассы и первичной продукции древесного яруса на верховых болотах в таежной зоне Западной Сибири, а также выявление закономерностей их изменения по широтному градиенту.
ОБЪЕКТЫ И МЕТОДИКА
Ключевые участки располагались на верховых болотах разных таежных подзон Западно-Сибирской равнины (табл. 1). В пределах зоны распространения олиготрофных сфагновых болот деревья растут на повышенных, наиболее дренированных участках болотных микроландшафтов. К ним относятся гряды, которые входят в состав грядово-мочажинных и грядово-озерковых комплексов, а также сосново-кустарничково-сфагновые болота (рямы). В данной работе мы используем понятие “рям” как ландшафтную категорию для обозначения сосново-кустарничково-сфагновых гомогенных болот, которые встречаются на протяжении всей зоны олиготрофных сфагновых болот Западной Сибири (Храмов, Валуцкий, 1977; Боч, Мазинг, 1979; Лапшина, 2003; Филиппов, Лапшина, 2008; Терентьева и др., 2017). Основной породой, формирующей древесный ярус данных болот, является сосна обыкновенная Pinus sylvestris L. с преобладающей формой f. litwinowii, а на высоких рямах – f. uliginosa. Также в небольшом числе часто присутствует сосна сибирская (кедровая) P. sibirica. В северной тайге, в зоне распространения бугристых болот, некоторые исследователи отмечают наличие на мерзлотных буграх редкой угнетенной сосны обыкновенной (Кац, 1941), либо лиственницы и кедровой сосны (Боч, Мазинг, 1979), либо всех перечисленных древесных пород (Пьявченко, 1985). Нами, как и в других современных работах (Москаленко, 2009), зафиксировано присутствие молодых деревьев сосны обыкновенной и иногда сосны кедровой на плоско- и крупнобугристых болотах в районе Сибирских Увалов.
Таблица 1.
Характеристика ключевых участков
| Подзона | Название болота | Болотный микро-ландшафт или его элемент | Географические координаты | Состав древесного яруса* | Число учетных площа-док, шт. | Размер учетных площадок, м2 | Густота древесного яруса, тыс. дер. га–1** | Высота деревьев, м*** | Диаметр деревьев, мм*** | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| с.ш. | в.д. | сосны обыкновенной | сосны кедровой | сухостоя | среднее значение | разброс значений | среднее значение | разброс значений | ||||||
| Южная тайга | Верх-Тарка | Рям | 56°48′ | 78°42′ | 10С | 1 | 10 × 10 | 11.3 | 0 | 2.4 | 1.0 | 0.4–2.5 | 25 | 6–90 |
| Плотниково | Гряды | 56°48′ | 82°51′ | 10С, 10СедК |
5 | По размеру гряд | 6.7 37.0 44.6 |
0 0 3.1 |
0.9 3.0 3.1 |
н.д. 1.5 1.0 |
н.д. 0.6–2.7 0.2–3.1 |
н.д. 37 25 |
н.д. 13–60 3–80 |
|
| Рям | 56°51′ | 82°50′ | 10СедК | 2 | 10 × 10 | 11.1 11.0 |
0.1 0.2 |
1.2 0.8 |
1.4 1.3 |
0.6–2.0 0.5–2.7 |
31 26 |
10–50 5–85 |
||
| Средняя тайга | Юган | Рям | 59°45′ | 75°37′ | 10СедК, 9С1К |
3 | 10 × 10 | 24.0 15.6 4.9 |
0.5 1.9 0 |
1.1 1.4 1.0 |
1.9 2.1 1.7 |
0.7–5.1 1.0–4.3 0.8–3.1 |
45 49 49 |
6–200 13–160 20–100 |
| Гряды | 59°39′ | 75°33′ | 8С2К | 1 | 1.8 × 72 | 5.9 | 3.6 | 1.9 | 1.6 | 0.7–2.5 | 42 | 22–67 | ||
| Мухрино | Высокий рям | 60°53′ | 68°39′ | 10СедКедБ | 1 | 10 × 10 | 8.6 | 0.6 | 0.3 | 3.1 | 0.7–10.6 | 37 | 14–160 | |
| Рям | 60°53′ | 68°41′ | 10СедК | 1 | 10 × 10 | 27.7 | 2.0 | 1.5 | 1.1 | 0.3–6.0 | 17 | 5–85 | ||
| Гряды | 60°53′ | 68°40′ | 9С1К | 4 | 1.5 × 10 | 13.0 | 2.0 | 1.7 | 1.2 | 0.3–3.3 | 26 | 5–90 | ||
| Нижневартовск | Гряды | 60°54′ | 76°49′ | 9С1К | 1 | 5 × 10 | 29.2 | 3.2 | 1.2 | 0.9 | 0.5–2.5 | 16 | 9–56 | |
| Первое | Гряды | 60°58′ | 69°45′ | 9С1К, 10С |
2 | 5.5 × 10, 5.5 × 2.5 | 2.9 12.4 |
0.4 0 |
1.5 2.2 |
0.5 1.7 |
0.5–3.0 0.6–3.5 |
39 28 |
18–75 15–70 |
|
| Чистое | Рям | 61°03′ | 69°27′ | 4С6К | 1 | 10 × 10 | 1.9 | 2.4 | 0.4 | 1.8 | 0.5–5.2 | 33 | 11–64 | |
| Неизвестное озеро | Гряды | 61°20′ | 73°05′ | 10С | 1 | 10 × 10 | 7.5 | 0 | 0.3 | 1.9 | 0.7–4.0 | 46 | 15–90 | |
| Федоровское | Рям | 61°25′ | 73°19′ | 9С1К | 1 | 10 × 10 | 8.5 | 0.8 | 0.6 | 1.5 | 0.6–3.0 | 38 | 10–110 | |
| Северная тайга | Пограничное | Высокий рям | 63°00′ | 75°12′ | 10C | 1 | 10 × 10 | 4.2 | 0 | 0.1 | 2.7 | 0.5–7.0 | 70 | 10–290 |
| Рям | 62°58′ | 75°11′ | 10С | 1 | 10 × 10 | 4.2 | 0 | 0.3 | 2.4 | 0.4–5.3 | 54 | 5–120 | ||
| Тету-Мамонтотяй | Мерзлотный бугор | 63°13′ | 75°43′ | 10С | 1 | 10 × 20 | 0.5 | 0 | 0.05 | 1.0 | 0.5–1.8 | 30 | 8–45 | |
| Западно-Ноябрьское | Гряды | 63°20′ | 75°22′ | 7С3К | 7 | 1.5 × 10 | 2.5 | 1.7 | 0.5 | 1.7 | 0.4–3.7 | 37 | 8–80 | |
| Рям | 63°20′ | 75°21′ | 10СедК | 1 | 10 × 10 | 6.0 | 0.2 | 0.8 | 1.1 | 0.3–2.6 | 21 | 5–65 | ||
В пониженных, наиболее обводненных болотных микроландшафтах – топях, мочажинах, ерсеях, которые образуют комплексы как с грядами, так и с мерзлотными буграми, – деревья не встречаются.
Сосны на верховых болотах отличаются низкорослостью и повышенной густотой (см. табл. 1) по сравнению с сосняками на суходолах, что потребовало модификации общепринятой методики. Во-первых, размер учетной площади сократили до 100 м2, поскольку на такой площади насчитывалось в среднем более 200 деревьев в южной тайге, где проводили первые исследования. В болотах средней и северной тайги применяли такую же методику, отработанную ранее в более южных районах. Небольшие размеры и вытянутая форма гряд обусловили выделение удлиненных прямоугольников для учета либо определяли площадь всей гряды. На выделенных площадях провели сплошной перечет деревьев, подроста и сухостоя. Во-вторых, диаметр деревьев измеряли на уровне мохового покрова, поскольку измерение на высоте 1.3 м невозможно при средней высоте древесного яруса, составляющей полтора метра, а измерение на уровне шейки корня затруднено в связи с обрастанием мхами и погребением ствола в моховом очесе, так что шейка оказывается на глубине 30–40 см от поверхности болота. Точность измерения диаметра составила 1 мм в связи с малым средним диаметром древесного яруса (около 36 мм). Такие же подходы к оценке таксационных показателей древесного яруса болот можно найти в работах А.А. Храмова и В.И. Валуцкого (1977), Е.А. Романовой (1980), С.Э. Вомперского и др. (1982), Е.А. Головацкой (2017). Высоту деревьев измеряли с точностью до 1 см. Возраст деревьев установили по годичным кольцам срубленных модельных деревьев. Для расчета запасов фракций биомассы и первичной продукции на учетных площадках отбирали по 5–8 модельных сосен высотой от 30 см до 3 м и диаметром ствола 3–70 мм, включая ствол, погруженный в моховой очес, с комлем. Всего с 2006 по 2015 гг. отобрано 115 модельных деревьев болотной формы f. litwinowii: 39, 52 и 24 экземпляра в болотах южной, средней и северной тайги, соответственно (от 5 до 15 на каждой учетной площади). Полевые работы велись во второй половине августа или в сентябре, когда вегетация заканчивается.
Для учета корней отбирали почвенные монолиты вглубь от уровня поверхности болота, т.е. от поверхности сфагнового покрова. Такой подход принят при оценке продуктивности верховых болот (Титлянова, 2007; Kosykh et al., 2008; Головацкая, 2009) и связан с тем, что верхний живой слой сфагнового покрова, мощностью в единицы сантиметров, с глубиной обычно без явных морфологических признаков переходит в отмерший очес и далее в торф, образуя единый слой со сходными физическими свойствами плотности, пористости, водоудержания. Для сосудистых растений этот почвенно-моховой слой является субстратом, поверх которого они поселяются, формируют побеги с фотосинтезирующими органами, в то время как корни трав и кустарничков обнаруживаются уже в поверхностном живом слое мха. Поэтому части растений, расположенные выше поверхности болота (мохового покрова) принято относить к надземной фитомассе, а ниже – к подземной. Монолиты объемом 1 дм3 отбирали послойно до глубины 30 см. Всего отобрано 104 монолита, из них 75 – в средней тайге, 12 – в южной и 17 – в северной (кроме бугристого болота). Для изучения распределения корней сосны в торфяной почве в ряме болотного комплекса Мухрино между двумя соснами, растущими на небольших кочках на расстоянии 2.05 м и с выровненным моховым покровом между ними, отбирали монолиты через каждые 15–20 см с отступом от стволов деревьев в 10 см. Отбор проводили послойно до глубины 40 см, поскольку болотные воды стояли ниже 45 см. Высота одного дерева была 135 см, диаметр 28 мм, другого – 173 см и 40 мм соответственно.
В лаборатории модельные деревья разбирали на фракции хвои, ветвей и ствола выше и ниже мохового покрова, а также сухих побегов (сухие ветви и не опавшая сухая хвоя вместе). Разделения ствола на фракции коры и древесины не проводили. Хвою и ветви, в свою очередь, делили на приросты текущего года и предыдущих лет. Фракции высушивали и взвешивали. Возраст модельных деревьев варьировал от 10 до 60 лет; один экземпляр, отобранный в среднетаежном Сургутском полесье, насчитывал 90 лет, что, по нашим данным, близко к предельному времени жизни сосен на олиготрофных сфагновых болотах. С возрастом увеличивались диаметр и высота деревьев. В среднем сосны от 20 лет и старше обладали диаметром ≥10 мм и высотой над уровнем мохового покрова 40–60 см.
Фитомассу разных фракций аппроксимировали полиномиальными уравнениями второй степени в зависимости от диаметра ствола на уровне мохового покрова, что обеспечило получение наибольшего коэффициента детерминации r2 (табл. 2). Наиболее надежные результаты получены для фракций ствола выше и ниже мохового покрова. Высокие значения коэффициентов детерминации получены для деревьев, которые растут в южной и северной тайге. Средняя тайга включала в данном исследовании наиболее протяженную по широте территорию с большим набором ключевых участков, что явилось причиной более низких значений коэффициентов детерминации. Недостатком использования этого типа уравнений явилось то, что в ряде случаев регрессионная кривая уходила в отрицательную область при наименьших значениях диаметра ствола. Поэтому на начальном этапе мы аппроксимировали массу фракций степенными уравнениями для деревьев с диаметром ствола менее 25 мм и полиномиальными – для более крупных модельных деревьев, а затем суммировали результаты для всего древесного яруса на учетной площади. Оказалось, что разница между результатами, полученными обоими способами, составляет от 0.4 до 4.1% как в большую, так и в меньшую сторону, в среднем 1.7%. То есть использование полиномиальных регрессий допустимо для расчета фитомассы всего древесного яруса, но непригодно для группы деревьев с минимальными диаметрами ствола. В северной тайге построение моделей проводили отдельно для деревьев сфагновых олиготрофных и плоскобугристых болот, поскольку их морфометрические и весовые показатели существенно отличались между собой. Число модельных деревьев с мерзлотного плоскобугристого болота составило четыре.
Таблица 2.
Параметры уравнения зависимости фракций фитомассы модельных деревьев от диаметра ствола на уровне мохового покрова, полинома второй степени*
| Фракция фитомассы | Коэффициенты | |||
|---|---|---|---|---|
| A | B | C | детерминации r2 | |
| Южная тайга (N = 39) | ||||
| Хвоя текущего года | 0.0254 | 0.0917 | –0.0582 | 0.956 |
| Хвоя прошлого года | 0.0306 | 0.0585 | –0.2133 | 0.885 |
| Трехлетняя хвоя | 0.0250 | –0.0995 | 0.7414 | 0.855 |
| Хвоя 4-х и более лет | 0.0060 | 0.2774 | –2.1373 | 0.863 |
| Ветви текущего года | 0.0052 | 0.0393 | –0.2324 | 0.886 |
| Ветви прошлого года | 0.0276 | –0.7971 | 4.9316 | 0.839 |
| Трехлетние ветви | 0.0083 | –0.0275 | 0.1549 | 0.937 |
| Ветви 4-х и более лет | 0.0705 | 1.0062 | –8.3616 | 0.878 |
| Надземная часть ствола | 0.0913 | 3.7093 | –22.6590 | 0.986 |
| Часть ствола, погребенного мхом, и комель | 0.1249 | 2.9011 | –15.4780 | 0.937 |
| Сухие ветви | –0.0395 | 3.2241 | –17.3580 | 0.639 |
| Средняя тайга (N = 52) | ||||
| Хвоя текущего года | –0.0075 | 1.2853 | –7.7307 | 0.696 |
| Хвоя прошлого года | –0.0047 | 1.3557 | –8.0100 | 0.730 |
| Хвоя трехлетняя | 0.0011 | 1.1785 | –7.5940 | 0.628 |
| Хвоя четырехлетняя и старше | 0.0039 | 1.1118 | –8.3242 | 0.584 |
| Ветви текущего года | –0.0015 | 0.2781 | –1.6787 | 0.692 |
| Ветви прошлого года | –0.0021 | 0.3247 | –1.5865 | 0.682 |
| Ветви трехлетние | –0.0022 | 0.3643 | –1.7657 | 0.502 |
| Ветви четырехлетние и старше | –0.0137 | 6.6325 | –51.1470 | 0.674 |
| Надземная часть ствола | 0.1374 | 3.8854 | –28.7280 | 0.955 |
| Часть ствола, погребенного мхом, и комель | 0.0959 | 4.0881 | –24.7150 | 0.868 |
| Сухие ветви | 0.0019 | 2.8488 | –24.5910 | 0.623 |
| Северная тайга (N = 24) | ||||
| Хвоя текущего года | 0.0039 | 0.1605 | –0.4664 | 0.882 |
| Хвоя прошлого года | 0.0016 | 0.6196 | –2.4420 | 0.916 |
| Хвоя трехлетняя | –0.0080 | 0.8018 | –3.6567 | 0.854 |
| Хвоя четырехлетняя и старше | –0.0054 | 0.7179 | –1.2208 | 0.447 |
| Ветви текущего года | 0.0018 | 0.0110 | 0.0737 | 0.916 |
| Ветви прошлого года | 0.0015 | 0.0732 | –0.2170 | 0.938 |
| Ветви трехлетние | –0.00007 | 0.1205 | –0.4435 | 0.943 |
| Ветви четырехлетние и старше | –0.0105 | 1.7930 | –8.5726 | 0.836 |
| Надземная часть ствола | 0.6220 | –10.2690 | 42.7260 | 0.995 |
| Часть ствола, погребенного мхом, и комель | 0.1833 | 0.3142 | –2.6689 | 0.971 |
| Сухие ветви | –0.0086 | 1.4013 | –8.2968 | 0.871 |
| Все таежные подзоны (N = 21) | ||||
| Шишки | –0.0007 | 0.3987 | –4.8177 | 0.360 |
Генеративные органы (стробилы, шишки) встречались среди модельных деревьев возрастом 16 лет и старше, диаметром не менее 8 мм и высотой над моховым покровом не менее 45 см. Поскольку доля модельных деревьев с генеративными органами составила чуть более 18%, для оценки фитомассы шишек построили уравнение аппроксимации без разделения по подзонам. Коэффициент детерминации в этом случае составил 0.360, так что полученный результат можно расценивать как ориентировочный.
Модельные деревья сосны кедровой (9 экз.) отобраны на двух болотах: болоте Чистом и грядах Западно-Ноябрьского болота, а полученные регрессионные зависимости применены для деревьев по всей Западной Сибири. Несмотря на достаточно высокие значения коэффициентов детерминации (около 0.9 для части ствола выше и ниже мохового покрова, 0.8 – для многолетних ветвей и хвои, 0.6 – для ветвей и хвои текущего года), малая выборка заставляет рассматривать полученные результаты как предварительные, а коэффициенты регрессионных уравнений не приводятся.
Сухостой на болотах чаще всего представлял собой стволы без ветвей, так что его запас рассчитывали по уравнениям аппроксимации для фракции живых стволов выше и ниже мохового покрова. Сухостой и сухие ветви вместе называли мортмассой.
Первичная продукция (нетто-продукция) характеризует интенсивность образования биомассы за определенный период. Надземную первичную продукцию определяли как сумму запасов хвои и ветвей текущего года. Прирост стволовой древесины напрямую не определяли, а рассчитывали, исходя из соотношения годичного радиального прироста ствола к годичному приросту ветвей, равному 2 : 1 (Храмов, Валуцкий, 1977; Вомперский, Иванов, 1982). Прирост многолетних ветвей в толщину считали равным нулю. Оценка подземной продукции деревьев не проведена, поскольку корректное определение возможно на основе изучения динамики запасов корней в течение лета, что в этой работе не было предусмотрено.
Для характеристики почвенных физико-химических условий роста деревьев в болотах проводили замеры портативными приборами в близлежащих топях и мочажинах, которые являются приемниками болотных вод, стекающих с гряд, рямов, бугров. Кислотность болотных вод определяли портативным рН-метром ИТ-1101 (ООО “Измерительная техника”, Москва), а общее содержание растворимых солей – солемером HANNA Dist-1 (HANNA Instruments, Германия).
Полиномиальная регрессия, уравнения и степень детерминации зависимости массы фракций деревьев от диаметра ствола на уровне мохового покрова были получены в программе Microsoft Excel 7.0. Для подтверждения гипотезы, что сосны, которые растут на сфагновых олиготрофных болотах в разных таежных подзонах, различаются параметрами фитомассы, в программе Statistica 8 применили процедуру дискриминантного анализа, где группирующим фактором выступили таежные подзоны, а независимыми переменными – запасы фракций фитомассы (хвоя текущего года и многолетняя, ветви текущего года и многолетние, ствол выше мохового покрова и часть ствола, погребенного мхом). Предварительно данные по разным фракциям фитомассы модельных деревьев подвергли логарифмическому преобразованию. Такой же анализ, где группирующим фактором был тип болотной экосистемы (рям и гряда), не выявил достоверных различий. В анализ включили модельные деревья олиготрофных сфагновых болот южной, средней и северной тайги и исключили модели из мерзлотного плоскобугристого болота северной тайги. Различия считали достоверными при значении уровня значимости Р < 0.05.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Почвы ключевых сфагновых болот торфяные олиготрофные, бугристых болот – торфяные мерзлотные. Болотные воды в почвах ключевых участков стояли на глубине 20–50 см от поверхности мохового покрова. На сфагновых олиготрофных болотах в южной, средней и северной тайге средние значения рН были 3.6, 3.7 и 3.8, а содержание солей – 39, 34 и 26 мг л–1 соответственно. В топях плоскобугристого мерзлотного болота рН 3.7, содержание солей – 20 мг л–1. Кислая реакция и чрезвычайно низкая минерализация типичны для верховых болот.
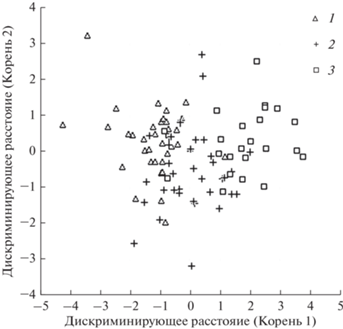
Дискриминантный анализ показал, что по запасам фракций фитомассы модельные деревья из южной и северной тайги достоверно различаются, а деревья из средней тайги занимают промежуточное положение, образуя в плоскости двух первых канонических дискриминантных функций совместные области как с северотаежными, так и с южнотаежными деревьями (рис. 1). Только первая дискриминантная функция статистически значимо (P < 0.00001) разделяла модельные деревья из разных подзон: собственное значение – 1.314104, коэффициент канонической корреляции – 0.75, критерий χ2 = 87.74226 при числе степеней свободы 12. При этом различия между средними значениями зависимой переменной дискриминантных функций во всех трех исследуемых группах статистически значимы (P < 0.0001). Наибольший вклад в дискриминацию трех исследуемых групп внесли фракции фитомассы хвои текущего года и прошлых лет (P < 0.0001), а также части ствола выше мохового покрова (P = 0.0024), в то время как фракции ветвей текущего года и части ствола, погребенного мхом, имели статистически значимое, но слабое влияние (P < 0.02), а фракции многолетних ветвей – несущественное (P > 0.05). Полученный результат послужил обоснованием расчета биомассы и продукции древесного яруса болот отдельно для каждой из таежных подзон.
Рис. 1.
Распределение модельных сосен в плоскости двух первых канонических дискриминантных функций: 1 – южная тайга; 2 – средняя тайга; 3 – северная тайга. Показатель лямбда Уилкса = 0.3833, f-критерий = 9.1256, P < 0.00001. Расстояния Махаланобиса между центроидами подзон следующие: южная тайга – средняя тайга 1.9006, средняя тайга – северная тайга 3.7255, южная тайга – северная тайга 8.8013.
Густота древесного ценоза составила 20.3 ± ± 6.6 тыс. дер. га–1 в южной тайге, 12.5 ± 2.6 – средней и 3.5 ± 0.9 – северной, линейно уменьшаясь с юга на север. Средняя высота на рямах и грядах (без учета высоких рямов) в южной тайге была 106 ± 3 см, средней – 103 ± 3 см, северной – 137 ± 11 см; средний диаметр у поверхности мохового покрова 25 ± 1, 20 ± 1 и 28 ± 2 мм соответственно. Известно, что древесному ярусу в болотах присуща абсолютная разновозрастность (Ефремов и др., 2005). Возраст сосен на ключевых участках во всех подзонах ограничивался обычно 80–100 годами, причем большая доля пришлась на деревья до 40 лет. Имеются сведения, что самые старые деревья на болотном массиве Мухрино достигают 250 лет и более при диаметре ствола около 18 см (Blanchet et al., 2016), что заставляет предположить их приуроченность к высоким рямам; на наших учетных площадках на этом же болоте таких старых и крупных сосен не встречалось.
Значительная доля сосны кедровой в составе древесного яруса отмечена только в двух местах – на Чистом болоте и грядах Западно-Ноябрьского болота, где средняя высота деревьев была 64 ± 16 и 26 ± 3 см, а средний диаметр – 17 ± 4 и 4 ± 1 мм соответственно. Возраст кедровой сосны на болотах за редким исключением не превышал 40 лет.
Суммарная усредненная надземная и подземная фитомасса древесного ценоза составила 1057, 613 и 333 г м–2 в южной, средней и северной тайге (табл. 3). Эти данные оказались ниже, чем по оценкам многих авторов. Так, фитомасса сосняка кустарничково-сфагнового болота в европейской части России была 39.4 т га–1 (Вомперский, Иванов, 1978), а такого же сосняка на юге Западной Сибири – 32.4 т га–1 (Пьявченко, 1967). Однако в обоих случаях речь шла о высоких рямах, что можно утверждать в первом случае в связи с приведенной оценкой высоты первого яруса деревьев (9 м), а во втором случае – по уточнению, что ценоз сложен не только f. litwinowii, но и f. uliginosa. Такие же высокие оценки (22.7–33.9 т га–1) есть в работе С.П. Ефремова с соавт. (2005): авторы уточняют, что данные приведены для низких рямов. Возможно, в их работе высокая оценка связана с иным методическим подходом, основанным на расчете запасов фракций фитомассы с помощью конверсионных коэффициентов в зависимости от объема стволовой древесины, либо обусловлена очень высокой густотой древесного яруса, которая, к сожалению, не указана в работе. С другой стороны, в работе Е.А. Головацкой (2017) средняя надземная фитомасса сосен на Бакчарском болоте, где мы тоже проводили исследование, близка к нашим оценкам. Усредненный по подзонам вклад кедровой сосны в общую фитомассу древесного ценоза составил в средней тайге 14%, а в остальных подзонах был незначителен.
Таблица 3.
Запасы фракций фитомассы древесного яруса, г м–2
| Фракция фитомассы | Южная тайга | Средняя тайга | Северная тайга |
|---|---|---|---|
| Сосна обыкновенная | |||
| Хвоя: | |||
| текущего года | 49.5 ± 13.1 | 14.1 ± 1.9 | 2.9 ± 0.2 |
| прошлого года | 56.9 ± 15.1 | 18.0 ± 2.3 | 3.6 ± 0.5 |
| трехлетняя | 41.7 ± 11.2 | 19.7 ± 2.5 | 1.9 ± 1.0 |
| четырехлетняя и старше | 18.8 ± 4.8 | 19.8 ± 2.6 | 3.4 ± 0.9 |
| Ветви: | |||
| текущего года | 10.6 ± 2.8 | 5.9 ± 0.8 | 0.9 ± 0.1 |
| прошлого года | 23.2 ± 6.9 | 3.8 ± 0.5 | 1.2 ± 0.1 |
| трехлетние | 14.0 ± 3.7 | 4.4 ± 0.6 | 0.8 ± 0.1 |
| четырехлетние и старше | 153.8 ± 40.0 | 82.1 ± 10.1 | 7.3 ± 1.6 |
| Ствол: | |||
| выше мохового покрова | 285.0 ± 72.9 | 185.1 ± 26.6 | 195.2 ± 47.4 |
| погребенный мхом + комель | 323.7 ± 83.8 | 156.1 ± 21.8 | 79.8 ± 12.4 |
| Генеративные органы | 6.3 ± 1.6 | 2.9 ± 0.9 | 1.1 ± 0.3 |
| Сухие ветви | 41.1 ± 9.2 | 39.3 ± 5.0 | 4.9 ± 1.2 |
| Сухостой: | |||
| выше мохового покрова | 29.6 ± 4.3 | 128.6 ± 50.0 | 49.0 ± 24.4 |
| ниже мохового покрова + комель | 38.9 ± 5.4 | 102.6 ± 36.8 | 19.0 ± 9.5 |
| Корни в слое 0–30 см | 73.6 ± 23.4 | 100.9 ± 11.1 | 34.6 ± 8.2 |
| Сосна кедровая | |||
| Хвоя | 0.4 ± 0.3 | 15.1 ± 9.0 | 0.4 ± 0.2 |
| Одревесневшие части: | |||
| надземные | 0.8 ± 0.7 | 41.1 ± 25.0 | 0.9 ± 0.5 |
| подземные + погребенные мхом | 0.4 ± 0.3 | 31.9 ± 18.9 | 0.9 ± 0.5 |
| Сухие ветви | 0.01 ± 0.01 | 0.7 ± 0.2 | 0.1 ± 0.1 |
Запасы отдельных фракций в южной, средней и северной тайге составили соответственно 167, 72 и 12 г м–2 для хвои; 202, 96 и 10 г м–2 – для ветвей; 587, 341 и 275 г м–2 – для ствола, включая часть, погребенную мхом. Запасы большинства фракций надземной и подземной фитомассы снижались от южной тайги к северной. Запасы хвои четырех лет и более, а также сухих ветвей, включая хвою, сходны в южной и средней тайге, а запас стволов выше мохового покрова сравним в средней и северной тайге.
В отличие от запаса фракций на единице площади, фитомасса фракций ствола в расчете на одно дерево увеличилась к северу, масса шишек была сходной в южной и северной тайге и уменьшилась в средней, а максимальная масса сухих ветвей пришлась на одно дерево в средней тайге (табл. 4). Масса хвои и веток на одном усредненном дереве также уменьшалась к северу, как и их площадные запасы.
Таблица 4.
Масса фракций фитомассы в расчете на одно дерево сосны обыкновенной
| Фракция фитомассы, г | Таежная подзона | ||
|---|---|---|---|
| южная | средняя | северная | |
| Хвоя | 82.2 | 57.3 | 33.5 |
| Ветви | 99.3 | 77.0 | 29.1 |
| Шишки | 3.1 | 2.3 | 3.1 |
| Ствол выше мохового покрова | 140.4 | 148.1 | 557.7 |
| Часть ствола, погребенного мхом | 159.4 | 124.9 | 227.9 |
| Корни | 36.3 | 80.7 | 98.9 |
| Сухие ветви | 20.2 | 31.5 | 13.9 |
| Сухостой | 33.7 | 185.0 | 194.3 |
Примечание. Масса фракций рассчитана как отношение фракций согласно табл. 3 к средней густоте деревьев в подзоне.
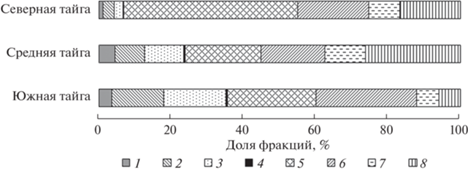
В структуре фитомассы характерно снижение с юга на север доли хвои, ветвей и шишек от общего запаса (рис. 2). Древесный ценоз в средней тайге отличался максимальным вкладом живых корней, сухих ветвей и сухостоя, а в северной – ствола выше мохового покрова. Полученные различия в структуре фитомассы соснового ценоза разных таежных подзон подтверждают важность ее учета при оценке продуктивности древесного яруса разных регионов (Усольцев и др., 2018).
Рис. 2.
Соотношение запасов фракций на единице площади в древесном ярусе болот таежных подзон: 1 – сухие ветви; 2 – хвоя; 3 – ветви; 4 – шишки; 5 – ствол выше мохового покрова; 6 – часть ствола, погребенная мхом, и комель; 7 – корни мелкие (до 2 мм в диаметре); 8 – сухостой.
Максимальный запас корней на единице площади наблюдался в средней тайге, а при пересчете на одно дерево приходилось в среднем 36, 81 и 99 г корней в толще 0–30 см в южной, средней и северной тайге соответственно. В этом же ряду отношение всей фитомассы к его корням составило 13, 5 и 9 в пересчете на одно усредненное дерево. Это в целом согласуется с данными об увеличении фитомассы сосущих корней к северу с понижением среднегодовой температуры, полученными для сосняков на минеральных почвах в Европе (Zadworny et al., 2016), что авторы объясняют адаптацией к снижению доступных ресурсов в более холодных почвах. Нарушение данной закономерности между средней и северной тайгой в нашей работе связано, вероятно, с разной обводненностью болот в этих подзонах.
В послойном вертикальном распределении корней сосен установлены следующие особенности. В северной тайге их запас снижался с увеличением глубины, составив 42% в толще 0–10 см, 37% – 10–20 см и 21% – 20–30 см. Снижение запаса корней с глубиной считается типичным для болотных сосняков (Згуровская, 1963). Однако в более южных районах нами получены данные, которые, напротив, свидетельствуют об увеличении запаса корней сосен с глубиной: в толще 0–10 см он составил 17 и 23%, 10–20 см – 39 и 32%, 20–30 см – 43 и 45% в южной и средней тайге соответственно. Такое распределение кажется труднообъяснимым для экосистем с близким стоянием болотных вод в почвенном профиле. Полагаем, что оно может быть связано с обогащением нижележащих горизонтов элементами питания в зоне периодического присутствия болотных вод, где деструкция торфа более интенсивна по сравнению с деструкцией вышележащего мохового очеса. Так, потери органического вещества при экспериментальной полевой деструкции в южной тайге за два года составили для мохового очеса из поверхностного слоя 26% (Вишнякова, Миронычева-Токарева, 2014), а для торфа, залегающего глубже, – 45% (Коронатова, 2010).
Более подробное изучение вертикального распределения фракций корней проведено на болоте Мухрино в средней тайге (рис. 3). Показано, что мелкие корни (диаметром менее 1 мм) были единственной фракцией на глубине 0–10 см и преобладали на глубине 20–40 см; фракция средних корней (1–3 мм) преобладала в толще 10–20 см и снижалась с глубиной; фракция крупных корней (более 3 мм) достигала максимума на глубине 30–40 см. Такое распределение отражало выполняемые функции и время жизни подземных органов: долгоживущие крупные проводящие корни находились на максимальной глубине, куда постепенно погружались с нарастанием вверх мохового покрова; отходящие от них средние корни нарастали вверх вслед за повышающейся поверхностью; короткоживущие, быстро обновляющиеся мелкие сосущие корни оказывались в обогащенных элементами питания местах – в поверхностном слое, куда поступает свежий опад, и ниже 20 см, где более интенсивны процессы деструкции органического вещества. Оказалось также, что в толще 30–40 см содержится 36% от общего запаса, т.е. в условиях достаточно низкого стояния болотных вод треть массы корней может быть сосредоточена ниже 30 см. Отрицательная геотропия корневых систем болотных сосен, превышение их радиуса над радиусом кроны характерны для изученных сосен и также установлены Л.Н. Згуровской (1963). Особенностью наших результатов является отсутствие снижения запаса корней с глубиной, что мы объясняем интенсивным линейным ростом сфагновых мхов на ключевых участках (Косых и др., 2017) и деструкционными процессам на нижней границе аэробной толщи торфа.
Рис. 3.
Распределение корней различного диаметра до глубины 40 см между двумя соснами, растущими на расстоянии 205 см друг от друга. Расстояние от первого дерева (см): I – 10–20; II – 38-48; III – 68–78; IV – 95–105; V – 128–138; VI – 158–168; VII – 185–195. Диаметр корней: 1 – <1 мм; 2 – 1–3 мм; 3 – >3 мм; 4 – все корни.
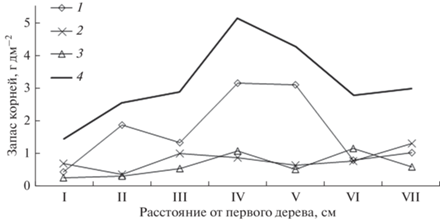
При изучении горизонтального распределения корней сосны в торфяной почве установлено, что максимальный запас сосредоточен на наибольшем удалении от стволов деревьев, причем он обеспечен в основном мелкими сосущими корнями (см. рис. 3). Запас средних и крупных корней постепенно возрастал по направлению к более высокому дереву.
Надземная первичная продукция древесного яруса в южной, средней и северной тайге составила 70.7 ± 17.7, 27.3 ± 3.9 и 4.7 ± 0.6 г м–2 год–1 соответственно, включая сосну кедровую, причем ее вклад в общую продукцию древесного яруса был незначителен на юге территории и увеличился до 5% в средней тайге и 2% – в северной.
Для оценки участия древесного ценоза болот в биотическом круговороте углерода важны соотношения компонентов, из которых складывается общий запас органического вещества растительности (табл. 5). В пределах зоны распространения сфагновых олиготрофных болот (таежная зона) с юга на север происходит увеличение доли ствола выше мохового покрова по сравнению с погребенным в мох, а также доли нефотосинтезирующей фитомассы по сравнению с фотосинтезирующей. Отношения хвои к корням и генеративным органам показывают, что к северу меньше пластических веществ направляется на постройку ассимиляционного аппарата и больше – на продукцию корней и органов размножения деревьев. Известно, что доля хвои сосен выше в олиготрофных условиях по сравнению с мезотрофными и евтрофными, поскольку в худших условиях больше органического вещества тратится на поддержание ассимиляционного аппарата (Вомперский, Иванов, 1978). В нашем случае в олиготрофных условиях доля хвои падает с увеличением суровости климата. Вероятно, в экстремальных условиях бóльшая доля ассимилятов направляется на развитие мелких сосущих корней (Zadworny et al., 2016) и органов размножения. Самая высокая доля мортмассы по сравнению с фитомассой была в средней тайге, самая низкая – в южной, что можно объяснить наиболее благоприятными условиями на юге территории и высокой обводненностью в ее средней части. Время оборота вещества в древесном ярусе, которое рассчитывается как отношение фитомассы к продукции (Титлянова, 1977), замедлилось с 15 лет в южной тайге до 72 лет в северной, что характеризует древесный ярус в круговороте как сток углерода, который депонируется на десятилетия.
Таблица 5.
Соотношение фракций фитомассы и время оборота вещества в зависимости от таежной подзоны
| Соотношение | Южная тайга | Средняя тайга | Северная тайга |
|---|---|---|---|
| Нефотосинтезирующая фитомасса : хвоя | 5.3 | 7.6 | 27.4 |
| Ствол над моховым покровом : часть ствола, погребенная мхом | 0.9 | 1.2 | 2.4 |
| Хвоя : корни | 2.3 | 0.7 | 0.3 |
| Хвоя : генеративные органы | 26.5 | 24.7 | 10.6 |
| Вся фитомасса : вся мортмасса | 9.6 | 2.3 | 4.3 |
| Время оборота вещества (фитомасса : продукция), год | 14.9 | 23.6 | 71.8 |
В отличие от остальных болот, мерзлотное болото Тету-Мамонтотяй лежит в зоне распространения плоскобугристых болот. Здесь сосны представлены молодняком возрастом около 20 лет, немногочисленная генерация включала 30–35-летние деревья. Древесный ярус обладал низкими запасами и незначительной годичной продукцией в силу своей разреженности: надземная фитомасса составила 33.9 г м–2, подземная – 6.6 г м–2, мортмасса – 0.4 г м–2, а надземная продукция – 2.4 г м–2 год–1. В то же время дендрометрические показатели деревьев этого болота оказались значительно выше, чем у деревьев олиготрофных сфагновых болот той же северотаежной подзоны. Так, при сравнении модельных деревьев возрастом 25 и 32 г. оказывается, что, в отличие от модельных деревьев средней и северной тайги, сосны плоскобугристых болот сравнимы или выше на 40 см, их диаметр больше на 4–7 мм, а часть ствола, погребенного мхом, в 2–2.5 раза короче. Запас фракций в расчете на одно усредненное дерево был выше, чем в южной тайге, и составил для хвои – 95 г, для ветвей – 121 г, в то время как запас сухих ветвей оказался самым низким – 9 г. На болоте Тету-Мамонтотяй отсутствовали деревья более старшего возраста, а также сухостой. Складывается впечатление, что сосны появились 2–3 десятилетия назад, возможно, в связи с глобальными климатическими изменениями. Известно, что на границах распространения леса чувствительность к климатическим изменениям максимальна и на севере проявляется в увеличении сомкнутости древостоев (Замолодчиков, Краев, 2016). Для юга Западной Сибири А.В. Глызин с соавт. (2005) отметили оптимум роста сосны в 1993 г. Возможно, в середине 1990-х гг. сложились благоприятные условия для роста сосен по всей Западной Сибири, что могло вызвать появление деревьев на данном болоте, однако это предположение требует подтверждений.
Исходя из опубликованных данных о площадях различных типов болот и доли облесенных болот на территории Западной Сибири (Вомперский и др., 2005, 2011), можно оценить запасы фитомассы древесного яруса в таежной зоне. При расчете на основе данных, опубликованных С.Э. Вомперским с соавт. (2011), мы принимали во внимание, что болотам с редкостойным древесным ярусом соответствуют низкие рямы и гряды с доминирующей формой f. litwinowii, сомкнутым – высокие рямы с f. uliginosa, для которых полученные результаты утраивали согласно имеющимся в литературе данным фитомассы низких и высоких рямов (Вомперский, Иванов, 1982). В таком случае общая фитомасса древесных ценозов на верховых болотах Западной Сибири в южной тайге равна 13.17 млн т, в средней – 124.47 млн т, в северной – 1.90 млн т., а суммарная – 139.54 млн т, что на порядок меньше, чем по оценкам С.П. Ефремова с соавт. (2005), и в 20 раз больше, чем по оценкам A.M. Peregon et al. (2008). Запасы сухостоя оцениваются в 1.17 млн т в южной, 46.95 млн т – в средней и 0.39 млн т – в северной тайге. Сопоставляя эти величины с данными о запасе сухостоя сосняков России (Замолодчиков и др., 2011), можно оценить вклад болот Западной Сибири в 4–9%.
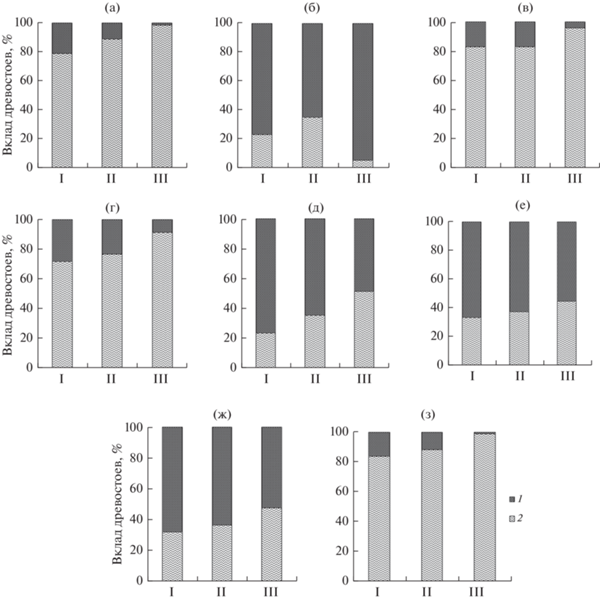
Рассмотрим вклад древесного яруса в общую фитомассу и продукцию болот, которые складываются участием деревьев, трав, кустарничков, лишайников и мохового покрова (рис. 4). Данные о продуктивности лишайниково-мохового и травяно-кустарничкового ярусов получены в тех же болотных массивах и опубликованы ранее (Косых, 2004; Косых, Махатков, 2008; Косых и др., 2003, 2008, 2017; Kosykh et al., 2008, 2017 и др.). Вклад хвои и подземной фитомассы деревьев был менее 20% и уменьшался к северу, в то время как участие деревьев в общем запасе нефотосинтезирующей надземной фитомассы, надземной и подземной мортмассы (без учета торфа и мохового очеса) оказалось значительным. Вклад деревьев в надземную первичную продукцию составил 16, 12 и 1% в южной, средней и северной тайге соответственно, что для южной тайги согласуется с опубликованной для Томской области оценкой в 18% (Пьявченко, 1967; Головацкая, 2017). Полученные результаты согласуются с выявленным ранее на автоморфных почвах республики Коми закономерным снижением продукции сосняков и увеличением вклада растений напочвенного покрова по мере продвижения в северном направлении (Осипов, Бобкова, 2019).
Рис. 4.
Вклад фракций фитомассы древесного яруса в общий запас растительных фракций в олиготрофных сфагновых болотах: (а) – хвоя; (б) – нефотосинтезирующая надземная фитомасса; (в) – подземная фитомасса; (г) – вся фитомасса; (д) – надземная мортмасса; (е) – подземная мортмасса; (ж) – вся мортмасса; (з) – надземная продукция. Запасы торфа и мохового очеса не включены. 1 – доля древесного яруса; 2 – суммарная доля мхов, кустарничков и трав. I – южная тайга; II – средняя тайга; III – северная тайга.
ЗАКЛЮЧЕНИЕ
В олиготрофных сфагновых болотах Западной Сибири биомасса и годичная продукция низкорослого древесного яруса, где преобладает P. sylvestris L. f. litwinowii, меняются вдоль широтного градиента. В связи с увеличением суровости климата с юга на север на единице площади уменьшались густота древесного яруса (от 20.3 ± 6.6 до 3.5 ± 0.9 тыс. дер. га–1), общая фитомасса (от 1057 до 333 г м–2) и запас большинства отдельных фракций. Запас фракций фитомассы в расчете на одно дерево также снижался с юга на север, кроме корней, в связи с более суровым климатом, и ствола, в связи с понижением уровня болотных вод в районе Сибирских Увалов. В средней тайге более высокая обводненность, чем в северной тайге, в сочетании с более суровым климатом, чем в южной тайге, послужили причиной максимальных запасов сухих ветвей и сухостоя (271 г м–2). Вклад кедровой сосны в фитомассу древесного яруса максимален в средней тайге, где достигает 14%.
В соотношении фракций древесного яруса олиготрофных сфагновых болот также прослеживаются закономерные изменения с юга на север: время оборота вещества в древесном ярусе замедлилось с 15 лет в южной тайге до 72 лет в северной, что сопровождалось увеличением доли нефотосинтезирующей фитомассы (ветвей, стебля, корней) и доли генеративных органов, которые приходятся на единицу фотосинтезирующей фитомассы. Самая высокая доля мортмассы по сравнению с фитомассой отмечена в средней тайге, самая низкая – в южной.
Установлены различия между древесным ярусом олиготрофных сфагновых и мерзлотных плоскобугристых болот: на мерзлотном болоте фитомасса и годичная продукция существенно выше в пересчете на одно дерево, но значительно снижаются в пересчете на единицу площади в связи с максимальной разреженностью деревьев.
Полученные оценки не претендуют на полноту, но позволяют сделать предварительные выводы об общих запасах биомассы низкорослого древесного яруса болот на территории Западной Сибири, которые составили в южной тайге 13.17 млн т, в средней – 124.47 млн т, в северной – 1.90 млн т, с вкладом сухостоя 4–9% от сухостоя сосняков на всей территории России. Вклад деревьев в общую фитомассу растительности олиготрофных болот достигает 20% в южной тайге и снижается до 9% в северной тайге, а вклад надземной первичной продукции снижается с 16 до 1% соответственно.
Список литературы
Биогеоценологическое изучение болотных лесов в связи с опытной гидромелиорацией / Под ред. Молчанова А.А. М.: Наука, 1982. 208 с.
Болота Западной Сибири, их строение и гидрологический режим / Под ред. Иванова К.Е., Новикова С.М. Л.: Гидрометеоиздат, 1976. 448 с.
Боч М.С., Мазинг В.В. Экосистемы болот СССР. Л.: Наука, 1979. 188 с.
Велисевич С.Н., Чернова Н.А. Морфогенез жизненных форм Pinus sibirica (Pinaceae) на олиготрофных болотах юга Западной Сибири // Ботанический журн. 2014. Т. 99. № 9. С. 988–1001.
Вишнякова Е.К., Миронычева-Токарева Н.П. Разложение сфагновых мхов в болотных экосистемах таежной и лесотундровой зон Западной Сибири // Торфяники Западной Сибири и цикл углерода: Матер. IV междунар. полевого симпозиума (г. Новосибирск, 4–17 августа 2014 г.). Томск: Изд-во Томского гос. университета, 2014. С. 160–162.
Вомперский С.Э., Глухов А.И. Геоморфологическое положение болотных лесов и безлесных болот различного генезиса // Биогеоценологическое изучение болотных лесов в связи с опытной гидромелиорацией. М.: Наука, 1982. С. 50–56.
Вомперский С.Э., Иванов А.И. Вертикально-фракционная структура и первичная продуктивность сосняков болотного ряда // Лесоведение. 1978. № 6. С. 13–23.
Вомперский С.Э., Иванов А.И. Первичная биологическая продуктивность болотных сосняков // Биогеоценологическое изучение болотных лесов в связи с опытной гидромелиорацией. М.: Наука, 1982. С. 94–132.
Вомперский С.Э., Лебков В.Ф., Иванов А.И. Таксационное строение болотных сосняков // Биогеоценологическое изучение болотных лесов в связи с опытной гидромелиорацией. М.: Наука, 1982. С. 57–94.
Вомперский С.Э., Сирин А.А., Сальников А.А., Цыганова О.П., Валяева Н.А. Оценка площади болотных и заболоченных лесов России // Лесоведение. 2011. № 5. С. 3–11.
Вомперский С.Э., Сирин А.А., Цыганова О.П., Валяева Н.А., Майков Д.А. Болота и заболоченные земли России: попытка анализа пространственного распределения и разнообразия // Известия РАН. Серия географическая. 2005. № 5. С. 39–50.
Глебов Ф.З. Взаимоотношения леса и болота в таежной зоне. Новосибирск: Наука, 1988. 184 с.
Глызин А.В., Размахнина Т.Б., Корсунов В.М. Дендрохронологические исследования в контактной зоне “лес – степь” как источник информации о ее динамике // Сибирский экологический журн. 2005. № 1. С. 79–83.
Головацкая Е.А. Биомасса и продукция древесного яруса сосново-кустарничково-сфагновых болот южной тайги Западной Сибири // Лесоведение. 2017. № 2. С. 102–110.
Головацкая Е.А. Биологическая продуктивность олиготрофных и эвтрофных болот южнотаежной подзоны Западной Сибири // Журн. Сибирского федерального университета. Серия: Биология. 2009. Т. 2. № 1. С. 38–53.
Ефремов С.П., Ефремова Т.Т., Блойтен В. Биологическая продуктивность и углеродный пул фитомассы лесных болот Западной Сибири // Сибирский экологический журн. 2005. № 1. С. 29–44.
Ефремова Т.Т., Аврова А.Ф., Ефремов С.П. Опыт построения бонитировочной шкалы местообитаний болотных сосняков южнотаежной подзоны Западной Сибири // Хвойные бореальной зоны. 2008. Т. 25. № 3–4. С. 269–276.
Замолодчиков Д.Г., Краев Г. Влияние изменений климата на леса России: зафиксированные воздействия и прогнозные оценки // Устойчивое лесопользование. 2016. № 4(48). С. 23–31.
Замолодчиков Д.Г., Зукерт Н.В., Честных О.В. Подходы к оценке углерода сухостоя в лесах России // Лесоведение. 2011. № 5. С. 61–71.
Згуровская Л.Н. Строение и рост корневых систем древесных растений на различных типах болот // Заболоченные леса и болота Сибири. М.: Изд-во АН СССР, 1963. С. 127–146.
Кац Н.Я. Болота и торфяники: Пособие для университетов. М.: Учпедгиз, 1941. 400 с.
Коронатова Н.Г. Исследование разложения торфа в болотах методом инкубации сухих и влажных образцов // Динамика окружающей среды и глобальное изменение климата. 2010. № 1. С. 65–71.
Коронатова Н.Г., Миляева Е.В. Продуктивность болотных сосняков южной тайги Западной Сибири // Интерэкспо Гео-Сибирь. Новосибирск: Сибирская гос. геодезическая академия, 2011. Т. 4. С. 259–262.
Косых Н.П. Болотные экосистемы таежной зоны Западной Сибири: фитомасса и продукция. Дис. … канд. биол. наук: 03.02.05. Томск, 2003. 149 с.
Косых Н.П., Махатков И.Д. Структура растительного вещества в лесо-болотных экосистемах средней тайги Западной Сибири // Вестник Томского гос. педагогического университета. 2008. Вып. 4(78). С. 77–80.
Косых Н.П., Миронычева–Токарева Н.П., Паршина Е.К. Биологическая продуктивность болот лесотундры Западной Сибири // Вестник Томского государственного педагогического университета. 2008. № 4(78). С. 53–57.
Косых Н.П., Миронычева-Токарева Н.П., Блейтен В. Продуктивность болот южной тайги Западной Сибири // Вестник Томского государственного университета. Приложение № 7, сер. Биологические науки (биология, почвоведение, лесоведение). 2003. С. 142–152.
Косых Н.П., Коронатова Н.Г., Лапшина Е.Д., Филиппова Н.В., Вишнякова Е.К., Степанова В.А. Линейный прирост и продукция сфагновых мхов в средней тайге Западной Сибири // Динамика окружающей среды и глобальное изменение климата. 2017. Т. 8. № 1(15). С. 3–13.
Лапшина Е.Д. Флора болот юго-востока Западной Сибири. Томск: Изд-во Томского ун-та, 2003. 296 с.
Лешок В.И., Дыренков С.А. Структура болотных сосняков Южной Карелии // Болотные экосистемы европейского Севера. Петрозаводск: Изд-во Карельского научного центра АН СССР, 1986. С. 78–93.
Махатков И.Д., Косых Н.П., Романцев С.А. Запасы фитомассы и годичная продукция верховых болот средней тайги // Торфяники Западной Сибири и цикл углерода: прошлое и настоящее: Матер. II междунар. полевого симпозиума (г. Ханты-Мансийск, 24 августа–2 сентября 2007 г.). Томск: Изд-во научно-технической литературы, 2007. С. 112–114.
Минаева Т.Ю., Сирин А.А. Биологическое разнообразие болот и изменение климата // Успехи современной биологии. 2011. Т. 131. № 4. С. 393–406.
Москаленко Н.Г. Изменение температуры пород и растительности под влиянием меняющегося климата и техногенеза в Надымском районе Западной Сибири // Криосфера Земли. 2009. Т. XIII. № 4. С. 18–23.
Осипов А.Ф., Бобкова К.С. Первичная продукция сосняков республики Коми // Лесоведение. 2019. № 5. С. 423–434. https://doi.org/10.1134/S0024114819050073
Панов В.В. Болотные формы Pinus sylvestris (Pinaceae) // Ботанический журн. 2007. Т. 92. № 5. С. 647–659.
Пьявченко Н.И. Лесное болотоведение (основные вопросы). М.: Изд-во АН СССР, 1963. 192 с.
Пьявченко Н.И. Биологическая продуктивность и круговорот веществ в болотных лесах Западной Сибири // Лесоведение. 1967. № 3. С. 32–43.
Пьявченко Н.И. Торфяные болота, их природное и хозяйственное значение. М.: Наука, 1985. 152 с.
Романова Е.А. Роль гидрологического режима в развитии древесной растительности на верховых болотах Нечерноземья // Значение болот в биосфере. М.: Наука, 1980. С. 147–152.
Романова Е.А. Растительность болот // Растительный покров Западно-Сибирской равнины. Новосибирск: Наука, 1985. С. 138–161.
Сукачев В.Н. О болотной сосне // Избранные труды. Л.: Наука, 1973. Т. 2. С. 13–24.
Терентьева И.Е., Сабреков А.Ф., Глаголев М.В., Лапшина Е.Д., Смоленцев Б.А., Максютов Ш.Ш. Новая карта болот южной тайги Западной Сибири для оценки эмиссии метана и диоксида углерода // Водные ресурсы. 2017. Т. 44. № 2. С. 209–220.
Титлянова А.А. Биологический круговорот углерода в травяных экосистемах. Новосибирск: Наука, 1977. 222 с.
Титлянова А.А. Чистая первичная продукция травяных и болотных экосистем // Сибирский экологический журн. 2007. Т. 14, № 5. С. 763–771.
Торфяные болота России: к анализу отраслевой информации / Под ред. А.А. Сирина, Т.Ю. Минаевой. М.: ГЕОС, 2001. 190 с.
Усольцев В.А. Биологическая продуктивность лесов Северной Евразии: методы, база данных и ее приложения. Екатеринбург: Изд-во УрО РАН, 2007. 636 с.
Усольцев В.А., Цепордей И.С., Шубаири С.О.Р., Дар Дж.А., Часовских В.П. Аддитивное аллометрические модели фитомассы деревьев и древостоев двухвойных сосен как основа региональных таксационных нормативов для Евразии // Эко-потенциал. 2018. № 1(21). С. 27–47.
Филиппов И.В., Лапшина Е.Д. Типы болотных микроландшафтов озерно-болотных систем Среднего Приобья // Динамика окружающей среды и глобальные изменения климата. Новосибирск, 2008. Т. 8. Вып. 1. С. 115–124.
Храмов А.А., Валуцкий В.И. Лесные и болотные фитоценозы восточного Васюганья (структура и биологическая продуктивность). Новосибирск: Наука, 1977. 222 с.
Blanchet G., Guillet S., Calliari B., Corona Ch., Edvardsson J., Stoffel M., Bragazza L. Impacts of regional climatic fluctuations on radial growth of Siberian and Scots pine at Mukhrino mire (central-western Siberia) // Science of the Total Environment. 2016. V. 574. P. 1209–1216. https://doi.org/10.1016/j.scitotenv.2016.06.225
Kosykh N.P., Koronatova N.G., Granath G. Effect of temperature and precipitation on linear increment of Sphagnum fuscum and S. magellanicum in Western Siberia // Russian J. Ecology. 2017. V. 48. № 3. P. 203–211. https://doi.org/10.1134/S1067413617030080
Kosykh N.P., Koronatova N.G., Naumova N.B., Titlyanova A.A. Above- and below-ground phytomass and net primary production in boreal mire ecosystems of Western Siberia // Wetlands Ecology and Management. 2008. V. 16. № 2. P. 139–153. https://doi.org/10.1007/s11273-007-9061-7
Moore P.D. The future of cool temperate bogs // Environmental Conservation. 2002. V. 29. P. 3–20. https://doi.org/10.1017/S0376892902000024
Peregon A.M., Maksyutov Sh.Sh., Kosykh N.P., Mironycheva–Tokareva N.P. Map-based inventory of wetland biomass and net primary production in western Siberia // J. Geophysical Research. 2008. V. 113. G01007. https://doi.org/10.1029/2007JG000441
Vitt D.H. An overview of factors that influence the development of Canadian peatlands // Memoirs of the Entomological Society of Canada. 1994. V. 169. P. 7–20.
Yefremov S.P., Yefremova T.T. Stocks and forms of deposited carbon and nitrogen in bog ecosystems of West Siberia // Proceedings of the International Field Symposium “West Siberian Peatlands and Carbon Cycle: Past and Present”. 2001. P. 148–151.
Zadworny M., McCormack M.L., Mucha J., Reich P.B., Oleksyn J. Scots pine fine roots adjust along a 2000-km latitudinal climatic gradient // New Phytologist. 2016. V. 212. № 2. P. 389–399. https://doi.org/10.1111/nph.14048
Дополнительные материалы отсутствуют.
Инструменты
Лесоведение