

Лесоведение, 2022, № 6, стр. 595-616
Oценка и приоритизация территорий для сохранения биоразнообразия на примере центра Европейской части России
Е. Н. Букварева a, *, А. А. Алейников b, О. А. Климанова c, Л. А. Титова c, Т. В. Свиридова d, А. В. Щербаков e
a Центр охраны дикой природы
117312 Москва, ул. Вавилова, д. 41, Россия
b Центр по проблемам экологии и продуктивности лесов РАН
117997 Москва, ул. Профсоюзная, д. 84/32, Россия
c Географический факультет МГУ им. М.В. Ломоносова
119991 Москва, ул. Ленинские горы д. 1, ГСП – 1, Россия
d Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский проспект, д. 33, Россия
e Биологический факультет МГУ им. М.В. Ломоносова
119991 Москва, ул. Ленинские горы д. 1, ГСП – 1, Россия
* E-mail: bukvareva@gmail.com
Поступила в редакцию 17.01.2022
После доработки 05.05.2022
Принята к публикации 07.06.2022
- EDN: VSQCFN
- DOI: 10.31857/S0024114822060031
Аннотация
Для адекватного учета и эффективного управления биоразнообразием и экосистемными услугами на обширной и чрезвычайно гетерогенной территории России необходим многоуровневый подход, интегрирующий задачи сохранения биоразнообразия на разных иерархических уровнях (разнообразие экосистем и видов), и на разных уровнях территориального управления. На примере Центрального федерального округа РФ рассмотрена предварительная методика приоритезация территорий для сохранения биоразнообразия на трех уровнях управления (федеральный округ – субъекты – муниципальные районы). Для приоритезации территорий использованы показатели редкости генерализованных типов экосистем в пределах этих территорий и значения территорий для сохранения “краснокнижных” видов животных и растений. Показано, что приоритетные для сохранения биоразнообразия генерализованные типы экосистем различаются как на разных территориальных уровнях, так и на разных территориях внутри одного уровня. Также возникает противоречие между управленческими задачами сохранения разнообразия видов, требующих обширных местообитаний, и сохранения редких экосистем, имеющих малую площадь. Эти противоречия могут быть решены на основе разработки природоохранных стратегий для разных уровней территориального управления.
Задача сохранения экосистем суши и биоразнообразия, а также интеграции их ценности в национальное и местное территориальное и экономическое планирование содержится в Целях устойчивого развития ООН (Mohieldin, Caballero, 2015). В России поставлена задача формирования экосистемного учета (United …, 2021) в рамках системы природно-экономического учета (ЭУ СПЭУ). Осознание ценности экосистем для качества жизни населения отражено в документах по реализации Государственной программы РФ “Комплексное развитие сельских территорий” (Постановление …, 2019), предполагающей проекты по улучшению качества природной среды в рамках деятельности органов местного самоуправления. Утверждение Стратегии социально-экономического развития Российской Федерации с низким уровнем выбросов парниковых газов до 2050 года (Распоряжение …, 2021) актуализирует работы по оценке качества и состояния экосистем лесов и представляемых ими экосистемных услуг. Формально задачи сохранения биоразнообразия реализуются в рамках национальных проектов РФ и учитываются на уровне субъектов Федерации и федеральных округов. В этой связи становится актуальной задача интеграции фундаментальных научных данных о зависимостях между биоразнообразием, экосистемным функционированием (ЭФ) и экосистемными услугами (ЭУ) в системы планирования территориального развития и сохранения биоразнообразия.
На обширной территории России адекватный учет экосистем, биоразнообразия и экосистемных услуг для управления ими может быть обеспечен только на основе многоуровневого подхода, учитывающего специфику регионов и разных уровней территориального управления (Экосистемные услуги …, 2020).
Цель статьи – проанализировать возможности и проблемы использования многоуровневого подхода (с учетом как иерархических уровней биоразнообразия, так и территориальных уровней управления) для приоритизации территорий и экосистем при принятии управленческих решений в области сохранения биоразнообразия. Первая часть статьи содержит краткий обзор современных представлений о связях между биоразнообразием и ЭФ/ЭУ на разных масштабах и в разных условиях. Затем представлена предварительная методика многоуровневой оценки важности экосистем и территорий для сохранения биоразнообразия, разработанная в рамках проекта TEEB-Russia, и проанализированы основные результаты ее апробации на примере Центрального федерального округа РФ. В заключительной части статьи рассмотрены главные проблемы и вопросы, которые требуют решения для дальнейшей разработки методики многоуровневой оценки территорий в целях сохранения биоразнообразия.
1. БИОРАЗНООБРАЗИЕ КАК ОСНОВА ЭКОСИСТЕМНЫХ ФУНКЦИЙ И ЭКОСИСТЕМНЫХ УСЛУГ
В настоящее время биоразнообразие признано одним из необходимых условий благополучия людей и достижения целей устойчивого развития, а в более узком научном смысле – ключевым фактором экосистемного функционирования, ослабление которого ведет к утрате жизненно важных экосистемных услуг (Cardinale et al., 2012; Tilman et al., 2014; IPBES, 2018; Van der Plas, 2019; Тебенькова и др., 2019; Лукина и др., 2020).
Для обеспечения ЭФ и ЭУ важны все иерархические уровни биоразнообразия - генетическое и фенотипическое разнообразие в популяциях и видах, разнообразие видов внутри сообществ и экосистем, разнообразие экосистем внутри ландшафтов и территорий различного размера (Букварева, Алещенко, 2013; Shin et al., 2019; Лукина и др., 2020; Arneth et al., 2020). Задача сохранения внутривидового генетического разнообразия важна, прежде всего, в отношении редких и исчезающих видов, а также эксплуатируемых промысловых объектов. Сохранение лесных генетических ресурсов рассматривается как основа обеспечения людей качественной лесной продукцией, а также эффективного выполнения лесами важнейших экосистемных услуг (Graudal et al., 2020). Однако далее в статье мы рассматриваем только разнообразие видов и типов экосистем.
Современное понимание взаимосвязей между биоразнообразием и ЭФ основано, прежде всего, на результатах сотен экспериментов, в которых измеряли показатели ЭФ (продуктивность, биомасса, запасы углерода, эффективность использования ресурсов и др.) в сообществах, искусственно составленных из разного числа видов. Наибольшее количество подобных экспериментов были проведены с травянистыми растениями на небольших участках или в контейнерах, размер которых не превышал нескольких квадратных метров, также использовали экспериментальные сообщества водных, наземных и почвенных беспозвоночных животных, водорослей, простейших и бактерий (Eisenhauer et al., 2019; Van der Plas, 2019). Относительно небольшая часть экспериментов была проведена с искусственными посадками деревьев на площадках в несколько десятков квадратных метров (Bruelheide et al., 2014; Verheyen et al., 2016). Эксперименты показали преобладание положительного влияния видового разнообразия на объем и устойчивость ЭФ. Однако практическое применение этих знаний в реальных условиях требует перехода к пространственно неоднородным территориям и масштабам, актуальным для управления в сфере охраны природы и природопользования (Cardinale et al., 2012; Brose, Hillebrand, 2016; Isbell et al., 2017).
Объединение классического для экологии тезиса о влиянии внешних условий на показатели видового разнообразия и ЭФ с современным пониманием того, как биоразнообразие влияет на ЭФ, формирует так называемую “новую парадигму” в области представлений о взаимодействии биоразнообразия и ЭФ (Loreau, 2010; Eisenhauer et al., 2019; Van der Plas, 2019). Специальные методы статистического анализа (в том числе – “structural equation modeling”, SEM) позволяют отделить воздействие внешних условий на биоразнообразие и ЭФ от воздействия биоразнообразия на ЭФ. Исследований экосистем в реальных условиях проведено значительно меньше, чем экспериментальных работ. Тем не менее показано, что в реальных условиях влияние биоразнообразия на ЭФ сопоставимо, а иногда превышает влияние абиотических факторов (Duffy et al., 2017; Van der Plas, 2019). Характер зависимостей между биоразнообразием и ЭФ (линейная позитивная или негативная, унимодальная, U-образная и т.п.), а также степень их статистической достоверности зависят от природных условий (климатические показатели, богатство почв, обеспеченность влагой и т.п.), степени антропогенной нарушенности (загрязнения, нарушенность местообитаний, интенсивность хозяйственного использования и др.), специфики биоценозов и популяций (межвидовые взаимодействия, трофическая структура популяций и т.п.). Тем не менее для травянистых, лесных, почвенных, пресноводных и морских сообществ выявлено преобладание положительных зависимостей между биоразнообразием и различными показателями ЭФ (Duffy et al., 2017; IPBES, 2018; Eisenhauer et al., 2019; Van der Plas, 2019).
Для лесных экосистем анализ более 700 тысяч площадок в 13 экорегионах по всему миру показал подавляющее преобладание положительных зависимостей между видовым богатством деревьев и продуктивностью экосистем (Liang et al., 2016). Однако, как показано для США (Watson et al., 2015), Китая (Baruffol et al., 2013; Chen et al., 2018) и европейских лесов (Vila et al., 2013), в разных типах леса и в разных условиях выявленные положительные зависимости различаются детальными характеристиками. В тропических лесах также обнаружены положительные зависимости (Cavanaugh et al., 2014; Poorter et al., 2015, 2017; Jucker et al., 2016b; Sullivan et al., 2017), хотя здесь частота их выявления не превышает частоту негативных зависимостей или же отсутствие любых зависимостей (Van der Plas, 2019). Ряд исследований, проведенных в Канаде (Paquette, Messier, 2011), Китае (Wu et al., 2015; Li et al., 2018; Liu et al., 2018), Европе (Jucker et al.,, 2016a; Ratcliffe et al., 2016), США (Potter, Woodall, 2014) и Японии (Mori, 2018a), показал, что положительное влияние разнообразия на ЭФ ослабевает и даже становится отрицательным при увеличении количества доступных для организмов ресурсов (в качестве показателей использовались климатические и биоклиматические индексы, а также средняя продуктивность лесов). Кроме того, было выявлено ключевое значение видового разнообразия для обеспечения мультифункциональности лесных экосистем (Schuldt et al., 2018). Зависимости между числом видов древесных растений и ЭФ могут различаться в лесных экосистемах разного сукцессионного статуса (Восточноевропейские …, 2004; Смирнова и др., 2006; Lasky et al., 2014; Cai et al., 2016; Schuldt et al., 2018).
Характер и выраженность зависимостей между биоразнообразием и ЭФ/ЭУ обусловлены масштабом, в котором проводятся исследования. При этом важны общая исследуемая площадь, размеры обследованных выделов лесов разного типа и размеры минимальных учетных площадок (Chisholm et al., 2013; McBride et al., 2014; Poorter et al., 2015; Barnes et al., 2016).
На уровне экосистем (сообществ) одного типа, существующих в относительно однородных природных условиях, проявляются те же причинно-следственные связи между разнообразием видов и ЭФ/ЭУ, что и в экспериментах. Преобладание положительных зависимостей в этом случае говорит о том, что утрата видов ведет к ослаблению ЭФ/ЭУ. Снижение числа видов в каждой отдельной экосистеме однозначно указывает на необходимость специальных мер по сохранению биоразнообразия для поддержания ЭФ/ЭУ в этом месте.
На ландшафтном уровне22 характер влияния биоразнообразия на ЭФ/ЭУ изменяется в зависимости от локальных условий и особенностей экосистем. Различные типы лесов, болот, лугов, составляющие “ландшафтную мозаику” природных экосистем, адаптированы к разным условиям среды и имеют разную степень нарушенности предшествующими воздействиями. Гипотеза оптимального биоразнообразия (Букварева, Алещенко, 2013) предполагает, что максимальная эффективность ЭФ достигается при оптимальных показателях разнообразия, снижаясь при любых отклонениях от оптимума. Ненарушенные природные сообщества, адаптированные к благоприятным и относительно стабильным локальным условиям среды, имеют более высокие оптимальные показатели видового разнообразия и ЭФ, а сообщества, адаптированные к скудным и нестабильным условиям – более низкие показатели того и другого. В подобных случаях более низкие показатели видового разнообразия (например, характерные для верховых болот) не говорят о меньшей ценности экосистем, так как именно такой уровень разнообразия обеспечивает их максимально эффективное и устойчивое функционирование в данных условиях. Однако при сравнении однотипных экосистем в пределах одного ландшафта пониженный уровень биоразнообразия в той или иной локальной экосистеме (например, на нарушенном осушенном верховом болоте или на участках леса, подверженных чрезмерному рекреационному воздействию) является опасным показателем ее деградации и вероятного снижения ЭФ/ЭУ. Такие нарушенные экосистемы менее ценны как поставщики ЭУ и нуждаются в восстановлении.
Кроме видового разнообразия, в каждой локальной экосистеме (α-разнообразие) важнейшим фактором эффективности и стабильности ЭФ/ЭУ на ландшафтном уровне является разнообразие экосистем и соответствующее ему видовое β-разнообразие, которое отражает смену видового состава в разных локальных экосистемах. Разные экосистемы производят разные ЭФ и ЭУ, обеспечивая мультифункциональность ландшафтов. Асинхронный ответ локальных экосистем на нарушающие воздействия и колебания условий среды обеспечивает устойчивость ландшафтного ЭФ в целом (Loreau et al., 2003; Olden, 2006). Положительное влияние разнообразия типов экосистем, сукцессионных стадий, конфигурации “ландшафтной мозаики” и β-разнообразия на мультифункциональность и устойчивость ЭФ показано для экспериментальных и реальных травянистых экосистем (Lamy et al., 2016; Grman et al., 2018; Hautier et al., 2018; Mori et al., 2018b) и на моделях лесных ландшафтов (Van der Plas et al., 2016).
На региональном уровне, который охватывает более обширные территории, причинно-следственные связи между биоразнообразием и ЭФ/ЭУ могут быть иными, чем на уровне одной экосистемы или ландшафта. На региональном масштабе возрастает вариабельность природных условий и степени антропогенных преобразований, может происходить смена биомов, радикально меняться структурно-функциональный тип экосистем. Иллюстрацией тому могут служить корреляции между индикаторами биоразнообразия и ЭФ/ЭУ, выявленные в пределах европейской территории России (далее – ЕТР) в рамках проекта TEEB-Russia 2 (Экосистемные услуги …, 2020). Например, на основе данных Атласа гнездящихся птиц Европейской России (Калякин, Волцит, 2020) на разных пространственных масштабах были выявлены разные зависимости между числом видов птиц и значениями некоторых ЭУ в квадратах 50 × 50 км. Еще одним примером может служить ЭУ обеспечения объема стока наземными экосистемами (далее – “экосистемный сток”). Показатель этой ЭУ вычислен как разность между наблюдаемым стоком и предполагаемым стоком c поверхности голого твердого грунта (Экосистемные услуги …, 2016, 2020). Для средних значений числа видов птиц и этой ЭУ в экорегионах выявлены негативная и унимодальная зависимости (рис. 1а). Для значений индикаторов в 50 км квадратах на ЕТР в целом проявлена негативная зависимость (рис. 1б). Однако внутри группы южных экорегионов (оранжевые точки на рис. 1б) зависимость положительна, а для группы северных, лесных и горных экорегионов (зеленые точки на рис. 1б) она негативна и сильнее выражена, чем для ЕТР в целом. Эти различия можно объяснить тем, что в группе северных, лесных и горных экорегионов показатели “экосистемного стока” и видового богатства изменяются на градиенте климатических условий противоположным образом: сток сокращается с севера на юг, а видовое богатство, напротив, возрастает. В группе южных экорегионов изменения этих индикаторов при движении с севера на юг однонаправленны – все показатели снижаются при движении от лесостепей к полупустыням. Для отдельных экорегионов эти зависимости могут быть разнонаправленными или же отсутствовать (рис. 1в).
Рис. 1.
Корреляции между числом видов птиц и величиной “экосистемного стока”: (а) зависимости между средними значениями индикаторов на ячейку карты в экорегионах; (б) зависимости между значениями в ячейках карты: серая линия – зависимость для всей выборки 50-км квадратов на ЕТР, зеленые точки и линия – значения и зависимость для 50-км квадратов группы северных, лесных и горных экорегионов; оранжевые точки и линия – значения и зависимость для 50-км квадратов группы южных экорегионов; (в) зависимости между значениями в ячейках карты в отдельных экорегионах. Средние значения индикаторов и зависимости для каждого из экорегионов показаны цветами, обозначенными в легенде.
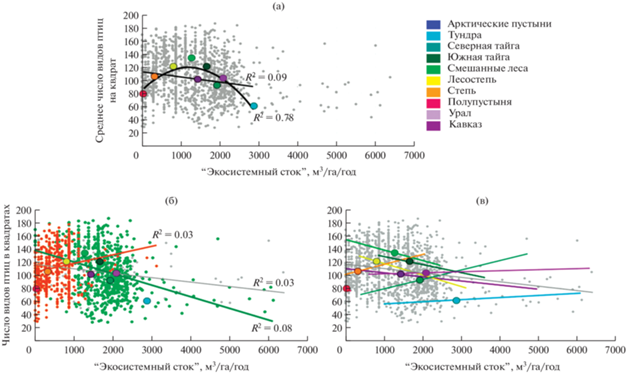
По результатам проекта TEEB-Russia 2 (Экосистемные услуги …, 2020) был сделан вывод о необходимости регионально-дифференцированных подходов к организации экосистемного учета в России, принимающего во внимание различия в зависимостях между биоразнообразием и ЭФ/ЭУ на разных пространственных масштабах.
Таким образом, для сохранения биоразнообразия как основы ЭФ/ЭУ необходима мультимасштабная экологическая политика (Isbell et al., 2017) на основе многомерного исследовательского подхода, который должен учитывать взаимные влияния биоразнообразия и ЭФ/ЭУ и их обоюдную зависимость от условий среды (Cardinale et al., 2009; Grace et al., 2016), а также разные иерархические уровни биоразнообразия (выше нами уже было рассмотрено разнообразие видов и экосистем).
2. ВОЗМОЖНЫЕ ПОДХОДЫ К МНОГОУРОВНЕВОЙ ОЦЕНКЕ И ПРИОРИТИЗАЦИИ ТЕРРИТОРИЙ ДЛЯ СОХРАНЕНИЯ БИОРАЗНООБРАЗИЯ НА ПРИМЕРЕ ЦЕНТРА ЕВРОПЕЙСКОЙ РОССИИ
В целях подготовки к началу формирования в России экосистемного учета в рамках проекта TEEB-Russia была разработана предварительная методика многоуровневой оценки важности33 (значимости) территорий и их приоритизации для сохранения экосистемного и видового разнообразия на разных уровнях территориального управления. Для учета природной зональности использованы лесные районы, поскольку их границы примерно совпадают с границами муниципальных образований. Учет на уровне лесных районов также целесообразен и для разработки мер по сохранению/восстановлению биоразнообразия при ведении лесохозяйственной деятельности, например, в рамках добровольной лесной сертификации.
Набор предлагаемых индексов включает внутри- и межуровневые индексы для оценки типов экосистем и территорий (табл. 1). Внутриуровневые индексы отражают важность (значимость) экосистем и территорий внутри соответствующего территориального уровня (ЦФО, лесного района, субъекта, муниципального района), а потому могут быть использованы для разработки мер по охране окружающей среды и социально-экономическому развитию этих территорий. Межуровневые индексы могут быть применены при необходимости учета задач сохранения биоразнообразия одновременно на нескольких уровнях.
Таблица 1.
Индексы для многоуровневой приоритизации типов экосистем и территорий в целях сохранения разнообразия экосистем. Названия индексов ценности типов экосистем выделены курсивом, индексов ценности территорий – полужирным шрифтом
| Индекс | Уровни решения задач | Формула для вычисления |
|---|---|---|
| Региональный ERi | Ценность каждого типа экосистем для региона | ERi = 100/SRi, где SRi – доля площади каждого типа экосистем в общей площади региона, %. |
| Лесорайонный EFi | Ценность каждого типа экосистем для лесного района | EFi = 100/SFi, где SFi – доля площади каждого типа экосистем в пределах лесного района, %. |
| Межуровневый индекс: лесорайонно-региональный EFRi |
Ценность каждого типа экосистем одновременно для двух уровней | Среднее значение между лесорайонным и региональным индексами для каждого типа экосистем |
| Суммарный индекс лесного района EFR | Важность лесного района для региона | Сумма индексов всех типов экосистем данного лесного района: ${{{\text{E}}}_{{{\text{FR}}}}} = \sum\limits_{i = 1}^n {{{E}_{{{\text{FR}}i}}}} $ |
| Субъектовый ESi | Ценность каждого типа экосистем для отдельного субъекта РФ | ESi = 100/SSi, где SSi – доля площади каждого типа экосистем в общей площади субъекта РФ, % |
| Межуровневые индексы: субъектно-региональный ESRi субъектно-лесорайонный ESFi субъектно-лесорайонно-региональный ESFRi |
Ценность каждого типа экосистем одновременно для двух или трех уровней | Вычисляются как средние значения между соответствующими индексами для каждого типа экосистем |
| Суммарные индексы субъекта РФ ESFR ESR |
Важность субъекта РФ для крупного региона | Cумма индексов ESFRi или ESRi для всех типов экосистем, представленных на территории субъекта: ${{{\text{E}}}_{{{\text{SR}}}}} = \sum\limits_{i = 1}^n {{{{\text{E}}}_{{{\text{SR}}i}}}} ;\,\,{{{\text{E}}}_{{{\text{SFR}}}}} = \sum\limits_{i = 1}^n {{{{\text{E}}}_{{{\text{SFR}}i}}}} $ |
| Муниципальный EMi | Ценность каждого типа экосистем для муниципалитета | EMi = 100/SMi где SMi – доля площади каждого типа экосистем в муниципалитете, %. |
| Межуровневые индексы: муниципально-субъектовый EMSi муниципально-лесорайонный EMFi муниципально-субъектно-лесорайонный EMSFi муниципально-субъектно-лесорайонно-региональный EMSFRi |
Ценность каждого типа экосистем одновременно для двух, трех или четырех уровней | Вычисляются как средние значения между соответствующими индексами для каждого типа экосистем |
| Суммарные индексы муниципального района EMS EMF EMSR EMSFR |
Важность муниципалитета для сохранения разнообразия экосистем с учетом разных уровней управления | Cумма индексов EMSi, EMFi, EMSFi или EMSFRi, в зависимости от выбранных уровней управления, для всех типов экосистем, представленных на территории муниципалитета: ${{{\text{E}}}_{{{\text{MS}}}}} = \sum\limits_{i = 1}^n {{{{\text{E}}}_{{{\text{MS}}i}}}} ;\,\,{{{\text{E}}}_{{{\text{MF}}}}} = \sum\limits_{i = 1}^n {{{{\text{E}}}_{{{\text{MF}}i}}}} ;{{E}_{{{\text{MSR}}}}}$ |
Апробация методики проводилась на основе использования общедоступных данных государственной статистики и открытых цифровых картографических материалов на примере Центрального федерального округа РФ (из-за специфики городской среды территория Москвы в ее прежних границах из анализа исключена). Границы лесных районов взяты с портала https://hcvf. ru/ru/maps/hcvf-russia. Подсчет площадей генерализованных типов экосистем произведен на основе специально созданной цифровой карты типов земельного (ландшафтного) покрова, которая интегрирует данные карты GLAD ARD Университета штата Мэриленд (Potapov et al., 2020), карты растительности ЦФО (Ершов и др., 2015) и карты сохранившихся участков степных экосистем по данным проекта “Совершенствование системы и механизмов управления ООПТ в степном биоме России” и портала “Сохранение степей России” (http://savesteppe.org/ru/steppe-project). На созданной карте были выделены пахотные угодья, застроенные территории, водные объекты, а также восемь типов наземных экосистем: темнохвойные, светлохвойные, лиственные, смешанные и заболоченные леса, а также болота, степные участки и участки с нестепной травянистой растительностью. К последнему типу были отнесены нераспаханные участки без древесной растительности, не относящиеся к степям. Очевидно, что выделенные типы не являются экосистемами в строгом научном смысле. Но более детальных цифровых карт экосистем для ЦФО в настоящее время в открытом доступе нет, поэтому на первом этапе апробации методики мы сочли возможным рассматривать их как генерализованные типы наземных экосистем.
Задачи сохранения видового разнообразия были решены на основе данных региональных Красных книг о числе “краснокнижных” видов птиц и млекопитающих в муниципальных районах. Использованы Красные книги областей ЦФО, изданные в следующие годы: Белгородская (2005), Брянская (2015), Владимирская (2010), Воронежская (2018), Ивановская (2017), Калужская (2017), Костромская (2009), Курская (2002), Липецкая (2014), Московская (2018), Орловская (2007), Рязанская (2011), Смоленская (1997), Тамбовская (2012), Тверская (2016), Тульская (2013) и Ярославская (2015) области. Кроме того, были использованы данные о числе и природоохранном статусе видов сосудистых растений в 8 областях (Владимирская, Воронежская, Московская, Тульская, Тамбовская, Ивановская, Рязанская, Липецкая), согласно данным проекта TEEB-Russia 2 (Экосистемные услуги …, 2020).
2.1. Оценка и приоритизация типов экосистем
Для оценки важности (значимости) различных типов экосистем для сохранения экосистемного разнообразия в пределах той или иной территории предлагается критерий редкости данного типа экосистем. Экосистемы, которые в пределах анализируемой территории имеют меньшую площадь, считаются редкими и имеющими приоритетное значение для сохранения экосистемного разнообразия. Такой критерий аналогичен подходу, используемому при сохранении видового разнообразия. В дальнейшем также могут быть разработаны и дополнительные показатели редкости экосистем, учитывающие их встречаемость и фрагментированность.
В пределах ЦФО округленные значения индексов экосистем изменяются от 2 (для наиболее распространенных типов, которые занимают 40–60% площади в пределах исследуемой территории) до 100 (для типов, которые занимают не более 1% площади). Оценка важности (значимости) разных типов экосистем существенно изменяется на разных пространственных масштабах (табл. 2). Так, для сохранения разнообразия экосистем в ЦФО наиболее важными оказываются степи. Но для сохранения разнообразия экосистем внутри лесных районов в пределах ЦФО (оценка по лесорайонному индексу) степи нигде не получили высший приоритет. Внутри всех районов, кроме южнотаежного, приоритетны темно- и светлохвойные леса. Относительно небольшие площади этих сообществ в пределах ЦФО объясняются длительной историей лесопользования (Евстигнеев, 2009; Браславская, 2020) и современными темпами рубок и пожаров, которые чаще всего приводят к смене хвойных лесов лиственными (Uvsh et al., 2020). В южнотаежном лесном районе приоритетным типом экосистем оказались болота, поскольку самые крупные болота в пределах ЦФО находятся в районе смешанных лесов. Стоит подчеркнуть, что лесорайонные коэффициенты, вычисленные в пределах ЦФО, не отражают полностью ситуации в лесных районах.
Таблица 2.
Индексы важности (значимости) генерализованных типов экосистем на разных территориальных уровнях. Индексы для лесных районов подсчитаны в пределах ЦФО. Места типа экосистем в рейтинге оценок внутри соответствующей территории выделены цветом
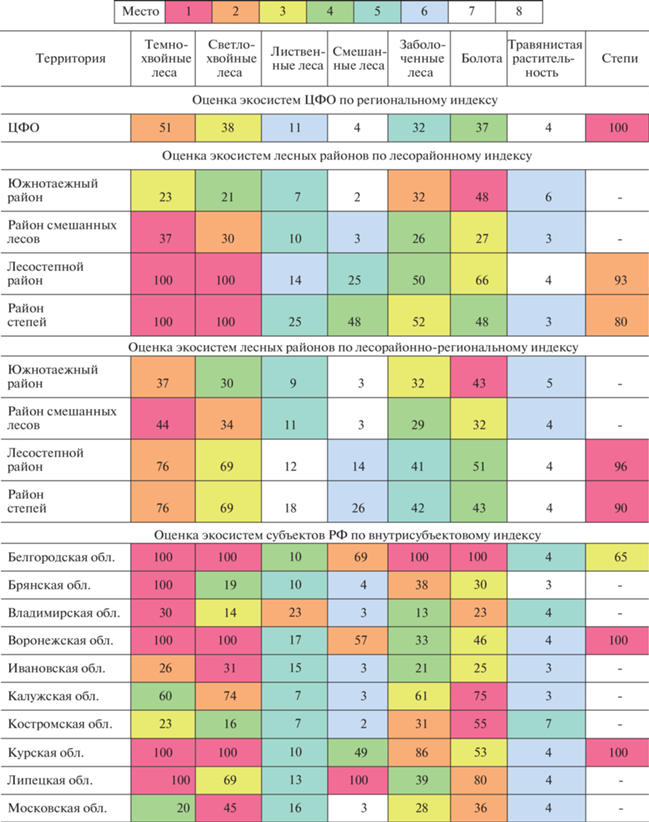

Внутри субъектов РФ результаты приоритизации типов экосистем также различны. В большинстве областей так же, как и в лесных районах, приоритетными оказались хвойные леса. Но в Костромской и Калужской областях высшую оценку получили болота, а в Тверской области – заболоченные леса. В Липецкой области, наряду с темнохвойными лесами, приоритетными оказались смешанные леса, которые ни в одной из других областей не получили высшей оценки. Внутри муниципальных районов приоритетные типы экосистем также различаются, причем нередко они не совпадают с таковыми в соответствующих субъектах Федерации (пример для муниципальных районов Ивановской области показан в табл. 2).
Очевидно, что результаты этой предварительной оценки во многом определяются точностью выделения экосистем на использованной нами цифровой карте земельного покрова, а генерализованные типы экосистем включают в себя как обычные экосистемы, так и редкие. Например, лиственные леса, широко представленные в большинстве регионов ЦФО, содержат как широко распространенные производные мелколиственные леса, так и редкие широколиственные. Для более точной оценки необходимы уточнения цифровых карт, в том числе с использованием лесотаксационных материалов.
Использование межуровневых индексов позволяет учесть задачи сохранения разнообразия экосистем одновременно на двух или более территориальных уровнях. Например, при использовании для оценки экосистем лесных районов лесорайонно-регионального индекса лучше учитывается приоритетность степей для ЦФО, и они становятся главными в южных лесных районах (табл. 1). В целом при использовании межрегиональных индексов различия между территориями одного уровня сглаживаются.
Использованные нами индексы вычисляются по площади разных типов экосистем (табл. 1). В дальнейшем в оценку необходимо включать и другие важные для сохранения биоразнообразия показатели состояния лесных экосистем – фрагментированность и возраст. Кроме того, необходимо сохранять не только типологическое, но и сукцессионное разнообразие экосистем. Поскольку периодические антропогенные нарушения, в основном пожары и рубки, возвращают экосистемы на более ранние сукцессионные стадии, то важными для сохранения следует считать сообщества на поздних стадиях сукцессии. Наилучшим образом разнообразие сукцессионных стадий, сформированное естественным путем, сохраняется в пределах крупных малонарушенных лесных территорий (МЛТ), где могут присутствовать не только ненарушенные экосистемы, но и старовозрастные леса, сформированные после нарушений разных типов (Алейников, 2021). Тем не менее для сохранения лесных экосистем приоритетными должны быть естественные разновозрастные леса, которые в освоенных человеком регионах можно считать специфическим типом редких экосистем, находящимся на грани исчезновения.
Одним из показателей значения экосистем для сохранения биоразнообразия является их принадлежность к категориям лесов высокой природоохранной ценности (в первую очередь, МЛТ) и водоохранных/нерестозащитных лесов, которые зачастую остаются не только последними рефугиумами регионального биоразнообразия, но и обеспечивают связность экосистем. Особой категорией являются уникальные для того или иного региона экосистемы, которые выделяются экспертно, как это, например, было сделано для северо-запада России (Сохранение …, 2011).
По критерию редкости экосистем самыми важными для сохранения биоразнообразия оказываются территории, наиболее сильно преобразованные человеком (что показано далее, например, на рис. 2а и 3б). Между индексами важности территорий муниципалитетов для сохранения разнообразия экосистем и долей площади в них природных экосистем выявлена отрицательная корреляция (коэффициент корреляции Пирсона для внутримуниципальных индексов равен –0.764**). Эта отрицательная корреляция отражает взаимосвязь двух аспектов антропогенной угрозы для природных экосистем: угрозы сокращения площади и угрозы утраты биоразнообразия. При использовании для оценки также критериев малой нарушенности и возраста экосистем наименее преобразованные обширные природные территории, бóльшая часть которых расположена в северной половине ЦФО, также получают высокую оценку.
2.2. Приоритизация территорий
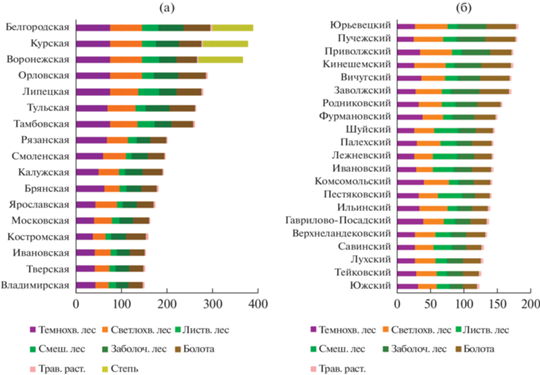
Приоритизация территорий для сохранения разнообразия экосистем осуществляется на основе коэффициентов их важности (значимости), которые являются суммой индексов важности экосистем внутри этих территорий (табл. 1). На рис. 2а показано ранжирование субъектов РФ по их важности для сохранения разнообразия экосистем в ЦФО, в лесных районах и в самих субъектах (по субъектно-лесорайонно-региональному индексу, см. табл. 1). На рис. 2б показан пример ранжирования муниципальных районов по их важности для сохранения разнообразия экосистем в пределах отдельного субъекта РФ (Ивановская обл.) с учетом задач сохранения разнообразия экосистем в лесном районе смешанных лесов, а также в самих муниципалитетах (по муниципально-лесорайонно-субъектовому индексу).
На рис. 3а показан пример пространственного распределения индексов важности экосистем для сохранения их разнообразия внутри отдельных лесных районов (лесорайонный индекс, см. табл. 1). Максимальную важность имеют редкие типы экосистем (с минимальной площадью), изолированные участки которых сосредоточены в южных областях ЦФО. В северной части ЦФО значительные территории заняты экосистемами с относительно большой площадью и невысокими индексами. На рис. 3б показана важность территории муниципалитетов для сохранения разнообразия экосистем в лесных районах и в самих муниципалитетах (муниципально-лесорайонный индекс, см. табл. 1). Оценка важности муниципалитетов возрастает с севера на юг, так как более южные районы, как правило, более сильно трансформированы человеком и угроза утраты в них разнообразия экосистем выше.

2.3. Использование показателей видового разнообразия для приоритизации территорий
В настоящее время в России отсутствует система мониторинга видового разнообразия для всей территории страны. Наилучшее покрытие территории (вся ЕТР) имеют данные о видовом богатстве птиц, собранные в рамках проекта “Атлас гнездящихся птиц Европейской России” (Калякин, Волцит, 2020). Для всей территории страны имеются сведения из региональных Красных книг о точках регистрации редких видов с указанием муниципальных районов (в некоторых региональных книгах муниципалитеты, где найдены виды, не указаны). Подходы составителей региональных Красных книг к выбору видов и степень изученности субъектов РФ существенно различаются. Поэтому сравнивать муниципалитеты в пределах крупного региона, включающего несколько субъектов РФ, можно не по числу отмеченных там “краснокнижных” видов, а по показателю доли от общего числа видов в региональной Красной книге. В дальнейших оценках может быть учтена также категория редкости видов.
При апробации методики мы использовали показатель доли числа “краснокнижных” видов, отмеченных в отдельных муниципалитетах, от общего числа видов, занесенных в региональную Красную книгу. Виды разных категорий редкости имели одинаковый “вес”. Число “краснокнижных” видов птиц и млекопитающих по муниципалитетам определено по данным региональных Красных книг, число сосудистых растений определено А.В. Щербаковым по литературным и гербарным данным в рамках проектов “Флора Окского бассейна” и “Флора Центрального Черноземья”.
Между индексами важности муниципалитетов для сохранения разнообразия экосистем и суммарным числом “краснокнижных” видов птиц и млекопитающих были выявлены либо слабые отрицательные корреляции, либо отсутствие зависимости. Для сосудистых растений ни в одной из восьми избранных областей ЦФО негативные корреляции не являются статистически значимыми, а для Тульской области проявляется тенденция к положительной корреляции (табл. 3). Это свидетельствует о том, что в проанализированных восьми областях ЦФО площади выделенных на карте участков редких экосистем недостаточны для обитания “краснокнижных” видов птиц и млекопитающих, но достаточны для сосудистых растений. Не исключено также, что более детальные обследования территорий могут изменить характер этих зависимостей, особенно в отношении небольших по размеру и малоподвижных видов.
Таблица 3.
Значения коэффициента корреляции Пирсона для зависимостей между долей числа видов, занесенных в региональные Красные книги, которые отмечены в муниципалитетах, и внутримуниципальным коэффициентом важности территории для сохранения разнообразия экосистем
| Регион (область) | Птицы и млекопитающие | Сосудистые растения |
|---|---|---|
| Владимирская (n = 16) | –0.504* | –0.400 |
| Воронежская (n = 33) | –0.510** | –0.77 |
| Московская (n = 39) | –0.399* | –0.059 |
| Тульская (n = 23) | 0.013 | 0.411 |
| Тамбовская (n = 23) | –0.728** | –0.394 |
| Ивановская (n = 21) | –0.174 | –0.219 |
| Рязанская (n = 25) | –0.550* | –0.077 |
| Липецкая (n = 18) | –0.622** | –0.402 |
| Все муниципалитеты 8 областей (n = 196) | –0.427** | –0.209** |
| Средние значения для 8 областей (n = 8) | –0.484 | –0.420 |
Задачи сохранения разнообразия видов и экосистем – два ключевых аспекта сохранения биоразнообразия, причем, не взаимозаменяющие, а взаимодополняющие друг друга. Из-за того, что необходимые площади местообитаний для сохранения разных видов различны, индексы относительно крупных и широко перемещающихся видов могут противоречить индексам, отражающим важность типов экосистем по критерию их редкости. Для более мелких по размеру и малоподвижных видов такое противоречие исчезает, причем именно отдельные, небольшие по площади участки редких типов экосистем оказываются критически необходимыми для сохранения “краснокнижных” видов растений и насекомых.
Дополнительным показателем для будущих оценок важности территорий может служить доля площади в их пределах ключевых орнитологических территорий (КОТР), которые выделяются по унифицированным качественным и количественным показателям видового разнообразия птиц (Свиридова и др., 2016) и представляют высокую ценность для сохранения разнообразия и качества экосистем (Экосистемные услуги …, 2020).
3. ЗАДАЧИ И ПРИОРИТЕТЫ СОХРАНЕНИЯ БИОРАЗНООБРАЗИЯ НА РАЗНЫХ ТЕРРИТОРИАЛЬНЫХ УРОВНЯХ: ПРОБЛЕМЫ И ВОПРОСЫ
Задачи и приоритеты охраны объектов биоразнообразия разных иерархических уровней на разных уровнях территориального управления должны различаться (табл. 4). В ряде случаев между приоритетами сохранения биоразнообразия на разных уровнях могут возникать кажущиеся противоречия, например, как показано выше – между приоритизацией типов экосистем по критерию их редкости и стремлением сохранить обширные малонарушенные массивы природных систем (см. раздел 2.1) или задачей сохранения видов, для выживания которых требуются обширные территории (см. раздел 2.3). Подобные противоречия между задачами сохранения биоразнообразия и поддержания/усиления экосистемных услуг (Sullivan et al., 2017) и задачами сохранения видового и экосистемного разнообразия (Bonn, Gaston, 2005) регулярно возникают в природоохранной практике. Однако они могут быть решены путем оптимизации природоохранных приоритетов и сценариев управления экосистемами (Socolar et al., 2015; Law et al., 2016), в том числе в области управления лесами (Trivino et al., 2016).
Таблица 4.
Задачи и приоритетные объекты для сохранения биоразнообразия наземных экосистем на разных территориальных уровнях. Курсивом выделены задачи и приоритетные объекты, касающиеся разнообразия экосистем
| Территориальные уровни организации биоразнообразия/ примеры объектов | Соответствие уровням государственного и ведомственного управления, а также местного самоуправления | Задачи сохранения разнообразия видов и экосистем | Приоритетные объекты сохранения биоразнообразия |
|---|---|---|---|
| Локальный/индивидуальная экосистема в однотипных локальных условиях | Выделы внутри муниципального образования/ лесничества | – Сохранение типичного для данного типа экосистем видового разнообразия – Сохранение популяций и местообитаний видов с минимальными требованиями к размеру местообитаний (мелкие и малоподвижные виды) – Сохранение участков (частей) местообитаний видов со средними и максимальными требованиями (крупные и мигрирующие виды) |
Редкие и исчезающие виды (занесенные в Красные книги РФ и субъектов РФ, а также локально исчезающие) |
| – Сохранение площади, предотвращение фрагментации данной экосистемы | – | ||
| Ландшафтный/совокупность индивидуальных экосистем | Муниципалитет Лесничество Субъект РФ |
– Сохранение разнообразия видов в пределах ландшафта или местности – совокупности
видов, характерных для сочетания индивидуальных экосистем – Сохранение популяций и местообитаний видов со средними требованиями к размеру местообитаний – Сохранение участков (частей) местообитаний видов с максимальными требованиями (крупные и мигрирующие виды) |
Редкие и исчезающие виды (КК РФ, региональные КК, а также локально редкие и исчезающие) |
| – Сохранение разнообразия экосистем (“ландшафтной мозаики”) – Сохранение связности (предотвращение фрагментации) природных экосистем |
– Редкие и исчезающие в данном ландшафте экосистемы – Наименее нарушенные экосистемы всех типов с типичным видовым разнообразием – Экосистемы на поздних стадиях сукцессии (старовозрастные леса) |
||
| Региональный/экорегион, биом, природная зона, бассейн крупной реки |
Группа субъектов РФ Федеральный округ Национальный уровень |
– Сохранение регионального и национального видового разнообразия – Сохранение популяций и ареалов видов с максимальными требованиями к размеру местообитаний |
Виды, занесенные в Красную книгу РФ |
| Сохранение разнообразия основных типов зональных и интразональных экосистем | – Редкие и исчезающие типы экосистем (например, европейские степи) – Уникальные экосистемы и природные комплексы – Малонарушенные природные территории (в т.ч. МЛТ) |
Для применения многоуровневого подхода на практике необходимо решить ряд вопросов:
– разработать методы пространственной интеграции характеристик природных систем и территориальных уровней управления, границы которых не совпадают;
– разработать подходы к интеграции задач сохранения разнообразия экосистем и разнообразия видов с учетом требований видов к размерам местообитаний и размерам сохранившихся редких типов экосистем;
– определить последовательность приоритизации объектов биоразнообразия на разных территориальных уровнях управления, которая позволили бы наиболее адекватно учитывать разный масштаб существующих типов природно-территориального деления, а также территориальные размеры объектов биоразнообразия;
– включить в оценку, кроме индексов редкости экосистем, определенных по их площади, ряд других показателей природоохранной ценности экосистем: возраст, степень нарушенности и фрагментированности, значение местообитаний редких, исчезающих и ключевых видов, важность отдельных экосистем для выполнения экосистемных услуг.
Важнейшей проблемой в России по-прежнему является недостаток требующихся для анализа исходных данных, а также затруднения при их получении и использовании, как объективные (разные единицы масштаба при сборе данных о различных группах организмов или ЭФ/ЭУ, не всегда поддающиеся унификации или интерполяции), так и субъективные (закрытость или сложность получения информации, в том числе высокая стоимость многих данных). В частности, для апробации рассмотренной методики даже для значительно генерализированных типов экосистем пришлось создавать собственную цифровую карту на основе интеграции данных из трех разных источников. Данные из федеральной Красной книги и из подавляющего большинства региональных Красных книг до сих пор не переведены в свободно доступную электронную форму (базу данных), поэтому для оценки важности территорий для сохранения “краснокнижных” видов необходима трудоемкая работа по поиску данных о регистрации видов в муниципалитетах в тексте Красных книг и занесению их в электронные таблицы.
ЗАКЛЮЧЕНИЕ
Анализ литературных источников и последних докладов международных проектов свидетельствует, что к сегодняшнему дню достигнут научный консенсус в понимании биоразнообразия как необходимой структурной основы выполнения экосистемных функций и услуг. Утрата биоразнообразия ослабляет и дестабилизирует ЭФ/ЭУ, что является угрозой для благополучия людей. Ключевая роль биоразнообразия в обеспечении устойчивого развития должна быть включена в принципы формирования экосистемного учета и использоваться для интерпретации его результатов при принятии решений в области природопользования и охраны природы. Для России, как для страны, имеющей крупнейшие в мире массивы природных экосистем, которые выполняют экосистемные услуги глобального значения, эта задача имеет первостепенную важность. Для выполнения ЭФ/ЭУ важны все иерархические уровни биоразнообразия – от внутрипопуляционного разнообразия до разнообразия экосистем. Также необходимо учитывать специфику “работы” биоразнообразия на разных пространственных масштабах.
Предварительная многоуровневая методика приоритизация территорий и экосистем для сохранения биоразнообразия на трех уровнях управления (федеральный округ – субъекты РФ – муниципальные районы), разработанная в рамках проекта TEEB-Russia, была апробирована на примере Центрального федерального округа РФ. Для учета природной зональности использовали лесные районы. Приоритизация генерализованных типов экосистем была основана на критерии редкости (показатель доли площади данного типа экосистем от общей площади территории). Более редкие типы экосистем считались приоритетными объектами охраны. Приоритизация территорий трех масштабных уровней (ЦФО, субъекты РФ и муниципалитеты) была основана на индексах ценности экосистем в пределах этих территорий и значении территорий для сохранения “краснокнижных” видов животных и растений.
Апробация методики показала, что приоритетные для сохранения биоразнообразия типы экосистем различаются как на разных территориальных уровнях (ЦФО, лесные районы, субъекты РФ, муниципалитеты), так и на разных территориях внутри одного уровня. Например, в большинстве областей ЦФО так же, как и в лесных районах, приоритетными оказались хвойные леса. Но в Костромской и Калужской областях высшую оценку получили болота, а в Тверской области – заболоченные леса. В Липецкой области, наряду с темнохвойными лесами, приоритетными оказались смешанные леса. В разных муниципальных районах приоритетные типы экосистем также различны, причем нередко они не совпадают с таковыми в соответствующих субъектах Федерации. Как было показано ранее результатами проекта TEEB-Russia, зависимости между биоразнообразием и ЭФ/ЭУ могут различаться на разных пространственных масштабах, что отражает специфику объектов биоразнообразия, которые имеют разные пространственные размеры, обитают и функционируют в разных природных и антропогенных условиях.
Выявленное противоречие между управленческими задачами сохранения редких экосистем и крупных ненарушенных природных массивов снимается путем разработки региональных природоохранных стратегий, учитывающих специфику сильно преобразованных человеком южных областей ЦФО и менее нарушенных северных областей, а также с помощью использования для приоритизации также критериев малой нарушенности и возраста экосистем.
Также выявлено противоречие между управленческими задачами сохранения разнообразия видов, требующих обширных местообитаний, и сохранения редких экосистем, имеющих малую площадь. Это противоречие проявилось в виде отрицательной корреляции между числом “краснокнижных” видов птиц и млекопитающих в муниципалитетах и индексами важности муниципалитетов для сохранения разнообразия экосистем. Однако для сосудистых растений статистически значимых негативных корреляций не выявлено. Это свидетельствуют о том, что площади отдельных участков редких экосистем недостаточны для обитания “краснокнижных” видов птиц и млекопитающих, но достаточны для сосудистых растений.
Таким образом, организация в России экосистемного учета в рамках системы природно-экономического учета (ЭУ СПЭУ) требует применения многоуровневого подхода, который должен учитывать задачи сохранения биоразнообразия на разных иерархических уровнях, прежде всего – разнообразия видов и разнообразия экосистем, а также специфику задач по сохранению биоразнообразия на разных уровнях территориального управления. Так, выявленные нами различия в приоритизации типов экосистем на разных территориальных уровнях и в пределах территорий одного уровня подчеркивают, что при принятии решений необходимо учитывать как межуровневые, так и внутриуровневые различия. Противоречия между управленческими задачами по сохранению разных объектов биоразнообразия (например, крупных и широко передвигающихся видов животных и редких типов экосистем) могут быть решены на основе разработки природоохранных стратегий для разных уровней территориального управления за счет выбора приоритетных объектов биоразнообразия с учетом требований видов к размерам местообитаний и размеров сохранившихся редких типов экосистем.
Список литературы
Алейников А.А. Историко-географические причины сохранности ненарушенных темнохвойных лесов Северного Урала // Лесоведение. 2021. № 6. С. 593–608.
Браславская Т.Ю. Леса и лесопользование на территории Звенигородской биостанции МГУ: XIX век // Russian Journal of Ecosystem Ecology. 2020. V. 5. № 2. P. 1–19.
Евстигнеев О.И. Неруссо-Деснянское полесье: история природопользования. Брянск: Государственный природный биосферный заповедник “Брянский лес”, 2009. 139 с.
Букварева Е.Н., Алещенко Г.М. Принцип оптимального разнообразия биосистем. М.: Товарищество научных изданий КМК, 2013. 522 с.
Восточноевропейские леса: история в голоцене и современность / Под ред. О.В. Смирновой. 2004. Кн. 1. М.: Наука, 479 с.
Ершов Д.В., Гаврилюк Е.А., Карпухина Д.А., Ковганко К.А. Новая карта растительности центральной части Европейской России по спутниковым данным высокой детальности // Доклады академии наук. 2015. Т. 464. № 5. С. 639–641.
Калякин М.В., Волцит О.В. Атлас гнездящихся птиц европейской части России. М.: Фитон XXI, 2020. 908 с.
Лукина Н.В., Гераськина А.П., Горнов А.В., Шевченко Н.Е., Куприн А.В., Чернов Т.И., Чумаченко С.И, Шанин В.Н., Кузнецова А.И., Тебенькова Д.Н., Горнова М.В. Биоразнообразие и климаторегулирующие функции лесов: актуальные вопросы и перспективы исследований // Вопросы лесной науки. Т. 3. № 4. 2020. С. 1–90.
Постановление Правительства Российской Федерации от 31.05.2019 г. №696 “Об утверждении государственной программы Российской Федерации “Комплексное развитие сельских территорий” и о внесении изменений в некоторые акты Правительства Российской Федерации” // Собрание законодательства Российской Федерации. 2019. № 23. Ст. 2953.
Распоряжение Правительства Российской Федерации от 29.10.2021 г. № 3052-р // Собрание законодательства Российской Федерации. 2021. № 45. Ст. 7556.
Свиридова Т.В., Зубакин В.А., Андреев А.В. Программа “Ключевые орнитологические территории России”: итоги 20 лет (1994–2014) // Инвентаризация, мониторинг и охрана ключевых орнитологических территорий России. М.: Союз охраны птиц России. 2016. № 7. С. 5–16.
Смирнова О.В., Бобровский М.В., Ханина Л.Г., Смирнов В.Э. Биоразнообразие и сукцессионный статус старовозрастных темнохвойных лесов Европейской России // Успехи современной биологии. 2006. Т. 126. № 1. С. 27–49.
Сохранение ценных природных территорий Северо-Запада России. Анализ репрезентативности сети ООПТ Архангельской, Вологодской, Ленинградской и Мурманской областей, Республики Карелии, Санкт-Петербурга / Под ред. Кобякова К.Н. СПб.: Кольский центр охраны дикой природы, 2011. С. 64–117.
Стратегия социально-экономического развития Российской Федерации с низким уровнем выбросов парниковых газов до 2050 года. Утверждена распоряжением Правительства Российской Федерации от 29 октября 2021 г. № 3052-р.
Тебенькова Д.Н., Лукина Н.В., Чумаченко С.И, Данилова М.А., Кузнецова А.И., Горнов А.В., Шевченко Н.Е., Катаев А.Д., Гагарин Ю.Н. Мультифункциональность и биоразнообразие лесных экосистем // Лесоведение. 2019. № 5. С. 341–356.
Экосистемные услуги России: Прототип национального доклада. Т. 1. Услуги наземных экосистем / Под ред. Е.Н. Букварёвой, Д.Г. Замолодчикова. М.: Изд-во Центра охраны дикой природы, 2016. 148 с.
Экосистемные услуги России: Прототип национального доклада. Т. 2. Биоразнообразие и экосистемные услуги: принципы учета в России / Под ред Е.Н. Букварёвой, Т.В. Свиридовой. М.: Изд-во Центра охраны дикой природы, 2020. 255 с.
Arneth A., Shin Y.-J., Leadley P., Rondinini C., Bukvareva E., Kolb M., Midgley G.F., Oberdorff T., Palomo I., Saito O. Post-2020 biodiversity targets need to embrace climate change // PNAS. 2020. V. 117. № 49. P. 30882–30891.
Barnes A.D., Weigelt P., Jochum M., Ott D., Hodapp D., Haneda N.F., Brose U. Species richness and biomass explain spatial turnover in ecosystem functioning across tropical and temperate ecosystems // Philosophical Transactions of the Royal Society B. 2016. V. 371: 20150279.
Barrufol M., Schmid B., Bruelheide H., Chi X., Hector A., Ma K., Michalski S., Tang Z., Niklaus P.A. Biodiversity Promotes Tree Growth during Succession in Subtropical Forest // PLoS ONE. 2013. V. 8. № 11: e81246.
Bonn A., Gaston K.J. Capturing biodiversity: selecting priority areas for conservation using different criteria. Biodiversity Conservation. 2005. V. 14. P. 1083–1100. https://doi.org/10.1007/s10531-004-8410-6
Brose U., Hillebrand H. Biodiversity and ecosystem functioning in dynamic landscapes // Philosophical Transactions of the Royal Society B. 2016. V. 371: 20150267.
Bruelheide H., Nadrowski K., Assmann T., Bauhus J., Both S., Buscot F., Chen X.-Y., Ding B., Durka W., Erfmeier A., Gutknecht J.L.M., Guo D., Guo L.-D., Härdtle W., He J.-S., Klein A.-M., Kühn P., Liang Y., Liu X., Michalski S., Niklaus P.A., Pei K., Scherer-Lorenzen M., Scholten T., Schuldt A., Seidler G., Trogisch S., von Oheimb G., Welk E., Wirth C., Wubet T., Yang X., Yu M., Zhang S., Zhou H., Fischer M., Ma K., Schmid B. Designing forest biodiversity experiments: general considerations illustrated by a new large experiment in subtropical China // Methods in Ecology and Evolution. 2014. V. 5. P. 74–89.
Cai H., Di X., Chang S.X., Jin G. Stand density and species richness affect carbon storage and net primary productivity in early and late successional temperate forests differently // Ecological Research. 2016. V. 31. № 4. P. 525–533.
Cardinale B.J., Duffy J.E., Gonzalez A., Hooper D.U., Perrings C., Venail P., Narwani A., Mace G.M., Tilman D., Wardle D.A., Kinzig A.P., Daily G.C., Loreau M., Grace J.B., Larigauderie A., Srivastava D.S., Naeem S. Biodiversity loss and its impact on humanity // Nature. 2012. V. 486. № 7401. P. 59–67.
Cardinale B.J., Bennett D.M. Nelson C.E., Gross K. Does productivity drive diversity or vice versa? A test of the multivariate productivity-diversity hypothesis in streams // Ecology. 2009. V. 90. № 5. P. 1227–1241.
Cavanaugh K.C., Gosnell J.S., Davis S.L., Ahumada J., Boundja P., Clark D.B., Mugerwa B., Jansen P.A., O’Brien T.G., Rovero F., Sheil D., Vasquez R., Andelman S. Carbon storage in tropical forests correlates with taxonomic diversity and functional dominance on a global scale // Global Ecology & Biogeography. 2014. V. 23. P. 563–573.
Chen S., Wang W., Xu W., Wang Y., Wan H., Chen D., Tang Z., Tang X., Zhou G., Xie Z., Zhou D., Shangguan Z., Huang J., He J.S., Wang Y., Sheng J., Tang L., Li X., Dong M., Wu Y., Wang Q., Wang Z., Wu J., Chapin F.S. III, Bai Y. Plant diversity enhances productivity and soil carbon storage // PNAS. 2018. V. 115. № 16. P. 4027–4032.
Chisholm R.A., Muller-Landau H.C., Rahman A.K., Bebber D.P., Bin Y., Bohlman S.A., Bourg N.A., Brinks J., Bunyavejchewin S., Butt N., Cao H., Cao M., Cárdenas D., Chang L.-W., Chiang J.-M., Chuyong G., Condit R., Dattaraja H.S., Davies S., Duque A., Fletcher C., Gunatilleke N., Gunatilleke S., Hao Z., Harrison R.D., Howe R., Hsieh C.-F., Hubbell S.P., Itoh A., Kenfack D., Kiratiprayoon S., Larson A.J., Lian J., Lin D., Liu H., Lutz J.A., Ma K., Malhi Y., McMahon S., McShea W., Meegaskumbura M., Razman M.S., Morecroft M.D., Nytch C.J., Oliveira A., Parker G.G., Pulla S., Punchi-Manage R., Romero-Saltos H., Sang W., Schurman J., Su S.-H., Sukumar R., Sun I.-F., Suresh H.S., Tan S., Thomas D., Thomas S., Thompson J., Valencia R., Wolf A., Yap S., Ye W., Yuan Z., Zimmerman J.K. Scale-dependent relationships between tree species richness and ecosystem function in forests // Journal of Ecology. 2013. V. 101. P. 1214–1224.
Duffy J.E., Godwin C.M., Cardinale B.J. Biodiversity effects in the wild are common and as strong as key drivers of productivity // Nature. 2017. V. 549. P. 261–264.
Eisenhauer N., Schielzeth H., Barnes A.D., Barry K., Bonn A., Brose U., Bruelheide H., Buchmann N., Buscot F., Ebeling A., Ferlian O., Freschet G.T., Giling D.P., Hättenschwiler S., Hillebrand H., Hines J., Isbell F., Koller-France E., König-Ries B., de Kroon H., Meyer S.T., Milcu A., Müller J., Nock C.A., Petermann J.S., Roscher C., Scherber C., Scherer-Lorenzen M., Schmid B., Schnitzer S.A., Schuldt A., Tscharntke T., Türke M., van Dam N.M., van der Plas F., Vogel A., Wagg C., Wardle D.A., Weigelt A., Weisser W.W., Wirth C., Jochum M. A multitrophic perspective on biodiversity-ecosystem functioning research // Advances in Ecological Research. 2019. V. 61. P. 1–54.
Graudal L., Loo J., Fady B., Vendramin G., Aravanopoulos F. A., Baldinelli G., Bennadji Z., Ramamonjisoa L., Changtragoon S., Kjær E. D. Indicators of the genetic diversity of trees – State, Pressure, benefit and response. State of the World’s Forest Genetic Resources – Thematic study. Rome, FAO, 2020. 92 p.
Grace J.B., Anderson T.M., Seabloom E.W., Borer E.T., Adler P.B., Harpole W.S., Hautier Y., Hillebrand H., Lind E.M., Pärtel M., Bakker J.D., Buckley Y.M., Crawley M.J., Damschen E.I., Davies K.F., Fay P.A., Firn J., Gruner D.S., Hector A., Knops J.M., MacDougall A.S., Melbourne B.A., Morgan J.W., Orrock J.L., Prober S.M., Smith M.D. Integrative modelling reveals mechanisms linking productivity and plant species richness // Nature. 2016. V. 529. P. 390–393.
Grman E., Zirbel C.R., Bassett T., Brudvig L.A. Ecosystem multifunctionality increases with beta diversity in restored prairies // Oecologia. 2018. V. 188. № 3. P. 837–848.
Hautier Y., Isbell F., Borer E.T., Seabloom E.W., Harpole W.S., Lind E.M., MacDougall A.S., Stevens C.J., Adler P.B., Alberti J., Bakker J.D., Brudvig L.A., Buckley Y.M., Cadotte M., Caldeira M.C., Chaneton E.J., Chu C., Daleo P., Dickman C.R., Dwyer J.M., Eskelinen A., Fay P.A., Firn J., Hagenah N., Hillebrand H, Iribarne O., Kirkman KP, Knops J.M.H., La Pierre K.J., McCulley R.L., Morgan J.W., Pärtel M., Pascual J., Price J.N., Prober S.M., Risch A.C., Sankaran M., Schuetz M., Standish R.J., Virtanen R., Wardle G.M., Yahdjian L., Hector A. Local loss and spatial homogenization of plant diversity reduce ecosystem multifunctionality // Nature Ecology and Evolution. 2018. V. 2. № 1. P. 50–56.
The IPBES regional assessment report on biodiversity and ecosystem services for Europe and Central Asia / Rounsevell M., Fischer M., Torre-Marin Rando A. and Mader A. (eds.). Secretariat of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. Germany, Bonn, 2018. 892 p.
Isbell F., Gonzalez A., Loreau M., Cowles J., Díaz S., Hector A., Mace G.M., Wardle D.A., O’Connor M.I., Duffy J.E., Turnbull L.A., Thompson P.L., Larigauderie A. Linking the influence and dependence of people on biodiversity across scales // Nature. 2017. V. 546. P. 65–72.
Jucker T., Avacaritei D., Barnoaiea I., Duduman G., Bouriaud O., Coomes D.A. Climate modulates the effects of tree diversity on forest productivity // Journal of Ecology. 2016a. V. 104. P. 388–398.
Jucker T., Sanchez A.C., Lindsell J.A., Allen H.D., Amable G.S. Coomes D.A. Drivers of aboveground wood production in a lowland tropical forest of West Africa: teasing apart the roles of tree density, tree diversity, soil phosphorous, and historical logging // Ecology and Evolution. 2016b. V. 6. P. 4004–4017.
Lamy T., Liss K.N., Gonzalez A., Bennett E.M. Landscape structure affects the provision of multiple ecosystem services // Environmental Research Letters. 2016. V. 11: 124 017.
Law E.A., Bryan B.A., Meijaard E., Mallawaarachchi T., Struebig M.J., Watts M.E., Wilson K.A. Mixed policies give more options in multifunctional tropical forest landscapes. Journal of Applied Ecology. 2017. V. 54. № 1. P. 51–60. https://doi.org/10.1111/1365-2664.12666
Lasky J.R., Uriarte M., Boukili V.K., Erickson D.L., Kress W.J., Chazdon R.L. The relationship between tree biodiversity and biomass dynamics changes with tropical forest succession // Ecology Letters. 2014. V. 17. P. 1158–1167.
Li S., Lang X., Liu W., Ou G., Xu H., Su J. The relationship between species richness
and aboveground biomass in a primary Pinus kesiya forest of Yunnan, southwestern China // PLoS ONE. 2018. Vol. 13. № 1: e0191140.
Liang J., Crowther T.W., Picard N., Wiser S., Zhou M., Alberti G., Schulze E.D., McGuire A.D., Bozzato F., Pretzsch H., de-Miguel S., Paquette A., Hérault B., Scherer-Lorenzen M., Barrett C.B., Glick H.B., Hengeveld G.M., Nabuurs G.J., Pfautsch S., Viana H., Vibrans A.C., Ammer C., Schall P., Verbyla D., Tchebakova N., Fischer M., Watson J.V., Chen H.Y., Lei X., Schelhaas M.J., Lu H., Gianelle D., Parfenova E.I., Salas C., Lee E., Lee B., Kim H.S., Bruelheide H., Coomes D.A., Piotto D., Sunderland T., Schmid B., Gourlet-Fleury S., Sonké B., Tavani R., Zhu J., Brandl S., Vayreda J., Kitahara F., Searle E.B., Neldner V.J., Ngugi M.R., Baraloto C., Frizzera L., Bałazy R., Oleksyn J., Zawiła-Niedźwiecki T., Bouriaud O., Bussotti F., Finér L, Jaroszewicz B., Jucker T., Valladares F., Jagodzinski A.M., Peri P.L., Gonmadje C., Marthy W., O’Brien T., Martin E.H., Marshall A.R., Rovero F., Bitariho R., Niklaus P.A., Alvarez-Loayza P., Chamuya N., Valencia R., Mortier F., Wortel V., Engone-Obiang N.L., Ferreira L.V., Odeke D.E., Vasquez R.M., Lewis S.L., Reich P.B. Positive biodiversity-productivity relationship predominant in global forests // Science. 2016. V. 354(6309): aaf8957.
Liu X., Trogisch S., He J.-S., Niklaus P.A., Bruelheide H., Tang Z., Erfmeier A., Scherer-Lorenzen M., Pietsch K.A., Yang B., Kühn P., Scholten T., Huang Y., Wang C., Staab M., Leppert K.N., Wirth C., Schmid B., Ma K. Tree species richness increases ecosystem carbon storage in subtropical forests // Proceedings of the Royal Society. 2018. V. 285: 2018124020181240.
Loreau M., Mouquet N, Gonzalez A. Biodiversity as spatial insurance in heterogeneous landscapes // PNAS. 2003. V. 100. № 22. P. 12765–12770.
Loreau M. Linking biodiversity and ecosystems: towards a unifying ecological theory // Philosophical Transactions of the Royal Society. 2010. V. 365. P. 49–60.
McBride P.D., Cusens J., Gillman L.N. Revisiting spatial scale in the productivity–species richness relationship: fundamental issues and global change implications // AoB Plants. 2014. V. 6: plu057.
Mohieldin M., Caballero P. Protect, restore and promote sustainable use of terrestrial ecosystems, sustainably manage forests, combat desertification, and halt and reverse land degradation and halt biodiversity loss // UN Chronicle. 2015. V. 51. № 4. P. 34–35.
Mori A.S. Environmental controls on the causes and functional consequences of tree species diversity. Journal of Ecology. 2018 a. V. 106. P. 113–125.
Mori A.S., Isbell F., Seidl R. β-diversity, community assembly, and ecosystem functioning. Trends in Ecology and Evolution. 2018 b. V. 33. P. 549–564.
Olden J.D. Biotic homogenization: a new research agenda for conservation biogeography // Journal of Biogeography. 2006. Vol. 33. P. 2027–2039.
Paquette A., Messier C. The effect of biodiversity on tree productivity: from temperate to boreal forests // Global Ecology and Biogeography. 2011. V. 20. P. 170–180.
Poorter L., van der Sande M.T., Thompson J., Arets E.J.M.M., Alarcón A., Álvarez-Sánchez J., Ascarrunz N., Balvanera P., Barajas-Guzmán G., Boit A., Bongers F., Carvalho F.A., Casanoves F., Cornejo-Tenorio G., Costa F.R.C., de Castilho C.V., Duivenvoorden J.F., Dutrieux L.P., Enquist B.J., Fernández-Méndez F., Finegan B., Gormley L.H.L., Healey J.R., Hoosbeek M.R., Ibarra-Manríquez G., Junqueira A.B., Levis C., Licona J.C., Lisboa L.S., Magnusson W.E., Martínez-Ramos M., Martínez-Yrizar A., Martorano L.G., Maskell L.C., Mazzei L., Meave J.A., Mora F., Muñoz R., Nytch C., Pansonato M.P., Parr T.W., Paz H., Pérez-García E.A., Rentería L.Y., Rodríguez-Velazquez J., Rozendaal D.M.A., Ruschel A.R., Sakschewski B., Salgado-Negret B., Schietti J., Simões M., Sinclair F.L., Souza P.F., Souza F.C., Stropp J., ter Steege H., Swenson N.G., Thonicke K., Toledo M., Uriarte M., van der Hout P., Walker P., Zamora N., Peña-Claros M. Carbon storage in tropical forests // Global Ecology and Biogeography. 2015. V. 24. P. 1314–1328.
Poorter L., van der Sande M.T., Arets E.J.M.M., Ascarrunz N., Enquist B.J., Finegan B., Licona J.C., Martínez-Ramos M., Mazzei L., Meave J.A., Muñoz R., Nytch C.J., de Oliveira A.A., Pérez-García E.A., Prado-Junior J., Rodríguez-Velázques J., Ruschel A.R., Salgado-Negret B., Schiavini I., Swenson N.G., Tenorio E.A., Thompson J., Toledo M., Uriarte M., van der Hout P., Zimmerman J.K., Peña-Claros M. Biodiversity and climate determine the functioning of Neotropical forests // Global Ecology and Biogeography. 2017. V. 26. P. 1423–1434.
Potapov P., Hansen M.C., Kommareddy I., Kommareddy A., Turubanova S., Pickens A., Adusei B., Tyukavina A., Ying Q. Landsat Analysis Ready Data for Global Land Cover and Land Cover Change Mapping // Remote Sensing. 2020. V. 12. № 3:426.
Potter K.M., Woodall C.W. Does biodiversity make a difference? Relationships between species richness, evolutionary diversity, and aboveground live tree biomass across U.S. forests // Forest Ecology and Management. 2014. V. 321. P. 117–129.
Ratcliffe S., Liebersgesell M., Ruiz-Benito P., Madrigal Gonzalez J., Munoz Costaneda J. M., Kandler G., Lehtonen A., Dahlgren J., Kattge J., Penuelas J., Zavala M. A., Wirth C. Modes of functional biodiversity control on tree productivity across the European continent // Global Ecology and Biogeography. 2016. V. 25. P. 251–262.
Schuldt A., Assmann T., Brezzi, M., Buscot F., Eichenberg D., Gutknecht J., Härdtle W., He J-S, Klein A-M, Kühn P., Liu X., Ma K., Niklaus P.A., Pietsch K.A., Purahong W., Scherer-Lorenzen M., Schmid B., Scholten T., Staab M., Tang Z., Trogisch S., von Oheimb G., Wirth C., Wubet T., Zhu C.-D., Bruelheide H. Biodiversity across trophic levels drives multifunctionality in highly diverse forests // Nature Communications. 2018. V. 9: 2989.
Shin Y.J., Arneth A., Chowdhury R., Midgley G.F., Leadley P., Agyeman Boafo Y., Basher Z., Bukvareva E., Heinimann A., Horcea-Milcu A.I., Kindlmann P., Kolb M., Krenova Z., Oberdorff T., Osano P., Palomo I., Pichs Madruga R., Pliscoff P., Rondinini C., Saito O., Sathyapalan J., Yue T. Chapter 4: Plausible futures of nature, its contributions to people and their good quality of life / Global assessment report of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. Brondízio E.S., Settele J., Díaz S., Ngo H.T. (eds). IPBES secretariat, Bonn, Germany. 2019. 168 p.
Socolar J.B., Gilroy J.J., Kunin W.E., Edwards D.P. How Should Beta-Diversity Inform Biodiversity Conservation? Trends in Ecology and Evolution. 2015. V. 31. № 1. P. 67–80. https://doi.org/10.1016/j.tree.2015.11.005
Sullivan M.J.P., Talbot J., Lewis S.L., Phillips O.L., Qie L., Begne S.K., Chave J., Cuni-Sanchez A., Hubau W., Lopez-Gonzalez G., Miles L., Monteagudo-Mendoza,A., Sonke B., Sunderland,T., ter Steege H., White L.J.T., Affum-Baffoe K. Diversity and carbon storage across the tropical forest biome // Scientific Reports. 2017. V. 7: 39102.
Tilman D., Isbell F., Cowles J.M. Biodiversity and ecosystem functioning // Annual Review of Ecology, Evolution, and Systematics. 2014. V. 45. P. 471–493.
Triviño M., Pohjanmies T., Mazziotta A., Juutinen A., Podkopaev D., Le Tortorec E., Mönkkönen M. Optimizing management to enhance multifunctionality in a boreal forest landscape. Journal of Applied Ecology. 2017. V. 54. № 1. P. 61–70. https://doi.org/10.1111/1365-2664.12790
United Nations. System of Environmental-Economic Accounting—Ecosystem Accounting (SEEA EA). White cover publication, pre-edited text subject to official editing. 2021. Available at: https://seea.un.org/ecosystem-accounting.
Uvsh D., Gehlbach S., Potapov P.V., Munteanu C., Bragina E.V., Radeloff V.C. Correlates of forest-cover change in European Russia, 1989–2012 // Land Use Policy. 2020. V. 96. P. 104 648–104 688.
Van der Plas F. Biodiversity and ecosystem functioning in naturally assembled communities // Biological Reviews. 2019. V. 94. P. 1220–1245.
Van der Plas F., Manning P., Soliveres S., Allan E., Scherer-Lorenzen M., Verheyen K., Wirth C., Zavala M.A. Ampoorter E., Baeten L., Barbaro L., Bauhus J., Benavides R., Benneter A., Bonal D., Bouriaud O., Bruelheide H., Bussotti F., Carnol M., Castagneyrol B., Charbonnier Y., Coomes D.A., Coppi A., Bastias C.C., Dawud S.M., De Wandeler H., Domisch T., Finér L., Gessler A., Granier A., Grossiord C. Guyot V., Hättenschwiler S., Jactel H., Jaroszewicz B., Joly F.X., Jucker T., Koricheva J., Milligan H., Müller S., Muys B. Nguyen D., Pollastrini M., Ratcliffe S., Raulund-Rasmussen K., Selvi F., Stenlid J., Valladares F., Vesterdal L., Zielínski D., Fischer M. Biotic homogenization can decrease landscape-scale forest multifunctionality // PNAS. 2016. Vol. 113. № 13. P. 3557–3562.
Verheyen K., Vanhellemont M., Auge H., Baeten L., Baraloto C., Barsoum N., Bilodeau-Gauthier S., Bruelheide H., Castagneyrol B., Godbold D., Haase J., Hector A., Jactel H., Koricheva J., Loreau M., Mereu S. Contributions of a global network of tree diversity experiments to sustainable forest plantations // Ambio. 2016. V. 45. № 1. P. 29–41.
Vila M., Carrillo-Gavilan A., Vayreda J., Bugmann H., Fridman J., Grodzki W., Haase J., Kunstler G., Schelhaas M., Trasobares A. Disentangling Biodiversity and Climatic Determinants of Wood Production // PLoS ONE. 2013. V. 8. № 2: e53530.
Watson J.V., Liang J., Tobin P.C., Lei X., Rentch J.S., Artis C.E. Large-scale forest inventories of the United States and China reveal positive effects of biodiversity on productivity // Forest Ecosystems. 2015. V. 2:22. P. 1-16.
Wu X., Wang X., Tang Z., Shen Z., Zheng C., Xia X., Fang J. The relationship between species richness and biomass changes from boreal to subtropical forests in China // Ecography. 2015. V. 38. P. 602–613.
Дополнительные материалы отсутствуют.
Инструменты
Лесоведение