

Микробиология, 2019, T. 88, № 6, стр. 673-684
Экология приустьевых акваторий малых рек южного Байкала по данным химических и микробиологических исследований в весенний период
Т. И. Земская a, *, А. С. Захаренко a, И. И. Русанов b, С. В. Букин a, Т. В. Погодаева a, О. Г. Нецветаева a, В. Г. Иванов a, Ю. Р. Штыкова a, Н. В. Пименов b
a Лимнологический институт СО РАН
664033 Иркутск, Россия
b Институт микробиологии им. С.Н. Виноградского, ФИЦ Биотехнологии РАН
119071 Москва, Россия
* E-mail: tzema@lin.irk.ru
Поступила в редакцию 17.05.2019
После доработки 10.06.2019
Принята к публикации 29.07.2019
Аннотация
В марте 2018 г. были проведены исследования воды, льда и снежного покрова в устьях и приустьевых акваториях рек юго-восточной и юго-западной частей озера Байкал, а также на двух фоновых станциях в пелагиали озера. Определены химический состав, общая численность микроорганизмов (ОЧМ), темновая ассимиляция углекислоты (ТАУ), количество органотрофных и санитарно-значимых бактерий в расплавах ледовых кернов и подледной воде устьев. Полученные результаты свидетельствуют о закислении льда в акваториях рек Переемная, Крестовка и Большая Черемшаная за счет поступления атмосферных осадков, обогащенных выбросами близрасположенных ТЭЦ, и влияния ложа рек. Показано, что отрицательного воздействия на ОЧМ, количество органотрофных бактерий и продукционные характеристики эти процессы не оказывают. По результатам измерения ТАУ ледовые сообщества микроорганизмов более активны в реках и приустьевых акваториях юго-восточной части озера (0.43 мкг C л−1 сут−1), в сравнении с таковыми юго-западной части Байкала (0.26 мкг C л−1 сут−1). Для водных образцов обнаружена сопряженность микробиологических показателей с содержанием биогенных элементов, суммой ионов и РОВ, также положительная корреляция отмечена между ОЧМ и рН (r = 0.66). Низкая активность микробных сообществ при высокой ОЧМ в устьях рек озера Байкал может быть следствием влияния антропогенного воздействия на продукционный потенциал микробиоценозов.
Байкал является самым глубоким и древним озером на Земле. Многочисленными исследованиями было показано стабильное состояние вод в пелагиали озера (Вотинцев, 1961; Falkner et al., 1991; Грачев, 2002), несмотря на наличие отдельных районов с высоким антропогенным влиянием (район Байкальского целлюлозно-бумажного комбината – БЦБК). Рассматривая проблему современного состояния Байкала, следует отметить, что эта экосистема весьма сильно изменялась в геологическом прошлом, и обладает достаточно мощной адаптивной системой, обеспечивающей приспособляемость к различным изменениям природной среды и климата, в том числе и катастрофическим. В озеро постоянно поступают отличающиеся по составу воды притоки подводных источников (Вотинцев и соавт., 1965; Сороковикова и соавт., 2015; Томберг и соавт., 2016), насыщенные метаном и иногда сероводородом (Намсараев и соавт., 1994; Гранина, 2008), а также нефтепродукты природного и антропогенного происхождения (Хлыстов и соавт., 2007). Поэтому острые проблемы локального загрязнения прибрежных участков акватории Байкала, его поверхностных вод существуют реально (Тимошкин и соавт., 2014; Ханаев и соавт., 2016; Timoshkin et al., 2016). Водные системы “река–приустьевая акватория” являются первым реагирующим, аккумулирующим и трансформирующим фильтром на пути к пелагиали озера Байкал. Наличие так называемых “барьерных зон”, защищающих пелагиаль озера Байкал от чуждых его экосистеме веществ, было показано в районах впадения крупных рек – Селенги и Верхней Ангары (Максименко и соавт., 2008, 2012). Приустьевые зоны небольших притоков, количество которых насчитывает более 330, рассматривались лишь эпизодически (Максимов, 1995). В последние годы именно в этих зонах наблюдается увеличение концентрации соединений фосфора и азота, а также массовое развитие неспецифичных для литорали озера Байкал водорослей (Тимошкин и соавт., 2014; Ханаев и соавт., 2016; Timoshkin et al., 2016). Вблизи поселков и портов летом регистрируются повышенные количества кишечной палочки – индикатора неочищенных фекальных стоков (Максимова, Максимов, 1989; Дрюккер и соавт., 1993; Штыкова и соавт., 2016). В прошлом столетии повышенная численность этих микроорганизмов отмечалась как вблизи поселков и портов, так и в районе промышленных сбросов Байкальского ЦБК, куда попадали бытовые стоки г. Байкальска (Максимова, Максимов, 1989; Дрюккер и соавт., 1993; Штыкова и соавт., 2016). Наблюдения за бактериями кишечной группы используются не только для оценки фекального загрязнения, но и как индикаторы органических веществ, поступающих в воду в результате антропогенного воздействия (Lemarchand et al., 2004; Stewart et al., 2007; Stoeckel, Harwood, 2007). Показано, что при высоких концентрациях питательных веществ продлевается период персистенции Escherichia coli в разных средах (Morris, Lewis, 1992; Wanjugi et al., 2016). На выживание бактерий в пресноводных экосистемах влияют и другие факторы, такие как седиментация, рН среды обитания, осмотические эффекты и концентрация органических и питательных веществ (Toothman et al., 2009; Surbeck et al., 2010; Schang et al., 2018).
Целью данной работы являлось исследование общей численности микроорганизмов (ОЧМ), численности органотрофных бактерий (ОБ), общего числа мезофильных аэробных и факультативно анаэробных микроорганизмов (общего микробного числа – ОМЧ) и темновой ассимиляции углекислоты (ТАУ) в зонах впадения в озеро Байкал небольших притоков, характеризующихся разной минерализацией, рН, Сорг, а также рекреационной нагрузкой.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
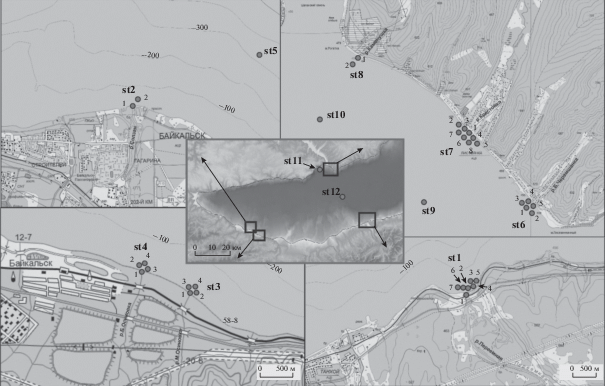
Районы исследования и отбор проб. Исследования воды, льда и снежного покрова проводили в марте 2018 г. в устьях и приустьевых акваториях рек юго-восточной (Переемная, Солзан, Большая и Малая Осиновки) и юго-западной (Каменушка, Крестовка, Большая Черемшаная) частей озера Байкал, а также на двух фоновых станциях в пелагиали озера (рис. 1). Вертикальное распределение микроорганизмов, использующих легкоминерализуемое органическое вещество, исследовали в районе впадения в озеро р. Каменушка, на расстоянии до 13 км от устья (st. 10–12). Воду отбирали с помощью батометра системы Нискина в устьях рек и в окружностях, на расстоянии 50–100 м от устья. Снег и лед отбирали в месте майны.
Рис. 1.
Карта-схема отбора проб в реках и приустьевых акваториях в Южном Байкале. Использованы карты Google.
Химический анализ проводили общепринятыми в гидрохимии пресных вод методами (Wetzel, Likens, 1991). Определение катионов (Na+, K+, Ca2+, Mg2+) выполняли атомно-абсорбционным и пламенно-эмиссионным методами (относительная погрешность 2–3%), анионов (Clˉ, ${\text{SO}}_{4}^{{2 - }},$ ${\text{HCO}}_{3}^{ - },$ ${\text{NO}}_{3}^{ - }$) – методом ВЭЖХ (относительная погрешность 5–10%; Барам и соавт., 1999). Определение биогенных элементов осуществляли колориметрическим методом: относительная погрешность метода для фосфат-ионов – ±1.5%, нитрит-ионов – ±3–5%. Определение аммонийного азота выполнено индофенольным методом, относительная погрешность до ±5%. Достоверность полученных результатов концентраций биогенных элементов подтверждали регулярным проведением контроля качества анализов в рамках международной программы EANET по тестированию стандартных образцов поверхностных вод. Измеренные концентрации главных ионов контролировали путем расчета ошибки ионного баланса (R1) и сравнением рассчитанной и измеренной удельной электропроводности (R2). Определение органических веществ проводили методами химического потребления кислорода (ХПК) оценивая бихроматную (БО) и перманганатную окисляемость (ПО). Ошибка использованных методов не превышает 10% (Wetzel, Likens, 1991). Содержание растворенного органического углерода (РОВ) определяли методом каталитического высокотемпературного окисления образцов при 850°С с последующим определением СО2 с помощью ИК детектора на высокотемпературном анализаторе углерода Vario TOC cube (“Elementar Analysensysteme GmbH”, Германия). В качестве конечного результата взяты средние значения трех измерений одного образца. Стандартное отклонение не превышает 0.01%.
Содержание метана определяли методом “Headspace” (Большаков, Егоров, 1987) на газовом хроматографе “Эхо_EW” с пламенно-ионизационным детектором (Новосибирск, Россия). Ошибка метода определения 7%. Измерение рН проводили в воде, размороженном снеге и расплавах ледовых кернов с помощью прибора pH 3310 (“WTW”, Германия).
Микробиологический анализ. В исследуемых пробах воды, льда и снега определяли ОЧМ, численность ОБ, общее число мезофильных аэробных и факультативно анаэробных микроорганизмов (ОМЧ), колиформных бактерий и спор сульфитредуцирующих клостридий. Для подсчета ОЧМ пробы воды и расплавов фиксировали 4% формалином, окрашивали флуорохромным красителем ДАФИ (4,6-диамидино-2-фенилиндол) (Porter, 1980) и просматривали под эпифлуоресцентным микроскопом AxioImager.M1 (“Carl Zeiss”, Германия). Для определения численности ОБ использовали среду с рыбо-пептонным агаром (РПА:10) (Горбенко, 1961). Чашки Петри с посевами инкубировали при 22°С в течение 7 сут, после чего учитывали количество выросших колоний. ОМЧ подсчитывали при росте микроорганизмов на мясо-пептонном агаре (МПА) при температуре 37°С в течение 24 ч и при температуре 22°С в течение 72 ч. Для подсчета колиформных бактерий и спор сульфитредуцирующих клостридий использовали метод мембранной фильтрации. Учет колиформных бактерий проводили на среде Эндо при температуре 37°С в течение 24 ч, подтверждая их наличие с помощью оксидазного теста (OXItest “Erba Lachema s. r. o.”, Чешская Республика) и инкубацией на лактозо-пептонной среде при температуре 37 и 44°С в течение 24 ч. Споры сульфитредуцирующих клостридий определяли на железосульфитном агаре при температуре 44°С в течение 16–18 ч (ISO 9308-1, 2000; Практикум по микробиологии, 2005).
Темновую ассимиляцию углекислоты микроорганизмами измеряли радиоуглеродным методом. Для этого воду из свежевскрытой майны отбирали в предварительно затемненные 100 мл флаконы и помещали в сумку-холодильник. От свежего керна откалывали нижнюю часть (10–15 см) льда, укладывали в темный пакет и транспортировали в стационарную лабораторию, где лед растапливали при комнатной температуре в темноте, пробу воды тщательно перемешивали и переливали в затемненные 100 мл флаконы.
Пробы воды и расплавов льда во флаконах перед инкубацией выдерживали несколько часов в холодильнике. Затем в каждый флакон вносили по 100 мкл раствора 14С-бикарбоната (10 мкКи на пробу). Затемненные флаконы закрепляли на горизонтальных крестовинах и помещали в подготовленную лунку для дальнейшей подледной инкубации в условиях “in situ”. В качестве контроля использовали склянки с пробами, фиксированными 0.5 мл 0.05 н HCl, до 14С-бикарбоната. Каждую пробу исследовали в двух повторностях с одним контролем. После суточной инкубации, склянки с пробами извлекали из лунки и немедленно фиксировали 0.05 н HCl. Для определения скорости включения 14С в состав биомассы микроорганизмов пробы фильтровали при 0.1 атм. через капроновые фильтры с диаметром пор 0.2 мкм. Фильтры заливали сцинтилляционной жидкостью и просчитывали на сцинтилляционном счетчике TRI-Carb TR (“Packard”, США). Для выявления включения 14С во фракцию растворенного органического вещества, фильтрат помещали в колбы установки для отгонки, продували воздухом в течение 40–45 мин и добавляли в колбу 10 г персульфата калия (K2S2O8). При постоянном кипячении, в течение 1.5 ч, отгоняли углекислоту, образующуюся в результате химического окисления органических соединений (мокрое сжигание), и улавливали в сцинтилляционный коктейль с органическим щелочным поглотителем. Расчет интенсивности ТАУ проводили по ранее описанной формуле (Pimenov, Bonch-Osmolovskaya, 2006).
Эффективность продукции бактериопланктона характеризуется величиной коэффициента Р/В, выражаемого отношением суточной продукции к величине биомассы бактериопланктона. Коэффициент Р/В, в ряде публикаций называемый µ = Р/В (сут–1), вычисляется при использовании величины суточной продукции бактериопланктона (Р), рассчитываемой по показателям темновой ассимиляции углекислоты, и величины микробной биомассы (В), получаемой прямыми микроскопическими методами на основании подсчета общей численности и измерения среднего объема клеток (Sorokin, 1990; Cooper et al., 2015).
Микробное окисление метана в пробах подледной воды определяли радиоуглеродным методом (14СН4). Воду наливали из батометра в 30 мл пенициллиновые флаконы с промывкой и изливом (без пузырька воздуха) и закрывали резиновой пробкой. После введения метки, пробы инкубировали 1 сут в подледной воде в условиях “in situ”. Для каждой пробы использовали 2 повторности и контроль (пробу, предварительно фиксированную 1 н КОН). При обработке фиксированных после инкубации проб в лабораторных условиях определяли переход углеродной метки из субстрата (меченый метан) в продукты микробного окисления: углекислоту, биомассу микроорганизмов (БМ) и внеклеточный растворенный органический углерод (РОУ). Обработку проб и расчеты интенсивности метанокисления проводили по ранее описанной методике (Русанов и соавт., 1998).
Для статистической обработки применяли метод главных компонент с нормализацией и центрированием данных, который был выполнен и визуализирован с использованием программного обеспечения BioVinCi версии 1.1.5 (“BioTuring Inc.”, США).
РЕЗУЛЬТАТЫ
Химический состав ледяных кернов характеризовался низкими концентрациями основных ионов, что, в первую очередь, обусловлено процессом обессоливания при образовании льда. В их составе преобладали ионы кальция и гидрокарбоната (рис. 2a), которые характерны для снегового покрова и ледяных кернов литорали озера (Нецветаева и соавт., 2013; Бондаренко и соавт., 2013). Значения рН в ледяных кернах из юго-западной части озера Байкал соответствовали слабокислым условиям (рН 6.23–6.62), содержание РОВ варьировало от 1.1 до 1.8 мг л–1, сумма ионов – от 3.19 до 5.93 мг л–1, при значении 2.2 мг л–1 на фоновой станции (табл. 1S). В ледяных кернах из рек Крестовка и Б. Черемшаная и их приустьевых акваторий отмечено повышенное содержание ионов хлора (0.96 мг л–1).
Рис. 2.
Диаграмма рассеяния точек в пространстве первых двух главных компонент, построенная на основе химического состава, микробиологических данных, концентраций биогенных элементов, рН, Σi, РОВ и ТАУ в ледовых (а, б) и водных (в, г) образцах. Стрелками отображены векторы, направленные по градиентам изменения исследуемых показателей в образцах.
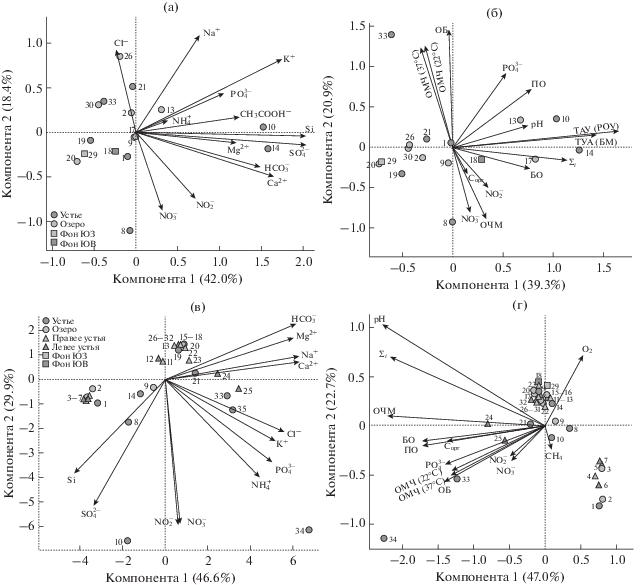
Таблица 1.
Пределы значений химических и биологических показателей в ледовой, речной и озерной водах Байкала
| Показатель | Значения в ледовой воде | Значения в устьях рек и приустьевой зоне | Значения в пелагиали Байкала, ссылка |
|---|---|---|---|
| ${\text{HCO}}_{3}^{ - },$ мг л–1 | 0–21.45 | 10.25–73.23 | 16.0–16.4 Folkner et al., 1991; Khodzher et al., 2018 |
| Cl–, мг л–1 | 0–0.96 | 0.10–4.34 | 0.4–0.5 Folkner et al., 1991; Khodzher et al., 2018 |
| ${\text{NO}}_{2}^{ - },$ мг л–1 | 0–0.039 | 0–0.096 | Не обнаружены |
| ${\text{NO}}_{3}^{ - },$ мг л–1 | 0–2.57 | 0.30–12.13 | 0.30–0.45 Sakirko et al., 2015 |
| ${\text{SO}}_{4}^{{ - 2}},$ мг л–1 | 0–2.73 | 5.30–21.63 | 5.3–5.5 Folkner et al., 1991; Grachev et al., 2004 |
| ${\text{NH}}_{4}^{ + },$ мг л–1 | 0.008–0.82 | 0.001–0.067 | Не более 0.05 устное сообщение, Сакирко |
| ${\text{PO}}_{4}^{{ - 3}},$ мг л–1 | 0–0.025 | 0.004–0.496 | 0.02–0.05 Sakirko et al., 2015 |
| Σi, мг л–1 | 2.2–33.84 | 33.43–125.88 | 94.3–96.3 Folkner et al., 1991; Grachev et al., 2004 |
| РОВ, мг л–1 | 1.1–3.3 | 0.85–4.85 | 1.05–1.3 Yoshioka et al., 2002 |
| О2, мг л–1 | – | 12.5–14.3 | 10.5–12.7 Khodzher et al., 2016 |
| рН | 5.76–7.42 | 6.7–8.16 | 7.6–8.0 Grachev et al., 2004 |
| ОЧМ, кл. мл–1 | 0.056 × 106–0.580 × 106 | 0.107 × 106–3.973 × 106 | 0.347–1.27 × 106 Максимова, Максимов, 1989 0.5–1.4 × 106 Straskrabova et al., 2005 |
| ОБ, КОЕ мл–1 | 0–546 | 1–6880 | До 200 Максимова, Максимов, 1989 |
| ОМЧ (22°С), число КОЕ в 1 мл | 0–401 | 0–6301 | Нет данных |
| ОМЧ (37°С), число КОЕ в 1 мл | 0–344 | 0–202 | 0–21 Дрюккер и соавт., 1993 |
| Темновая ассимиляция углекислоты (бактериальная продукция), мкг С л–1 сут–1 | В биомассу 0.01–0.37 В РОУ 0.02–0.47 |
В биомассу 0.07–0.69 В РОУ 0.19–1.29 |
0.032–1.43 Максимова, Максимов, 1989 1.11–1.63 Максимова, Максимов, 1989 |
В ледяных кернах из юго-восточной оконечности озера Байкал наблюдали смещение рН в сторону пониженных значений только в приустьевой акватории р. Переемная (рН 5.76–6.41) (рис. 2а, 2б). Значения рН в образцах из других рек соответствовали нейтральным или слабокислым условиям. Содержание РОВ в ледяных кернах этой акватории озера варьировало от 1.4 до 3.3 мг л–1, при концентрации 1.1 мг л–1 на фоновых станциях (табл. 1S). Сумма ионов изменялась в более широком диапазоне и составляла 2.9–33.84 при 13.01 мг л–1 на фоновой станции. В ледяных образцах р. Солзан отмечались повышенные концентрации нитрит- и нитрат ионов (0.039 и 2.57 мг л–1 соответственно), а в реках Малая и Большая Осиновки – ионов кремния (0.84 и 0.95 мг л–1) и сульфата (2.47 и 2.73 мг л–1 соответственно) (рис. 2а, 2б).
Микробиологические исследования. ОЧМ в ледяных кернах всех исследованных акваторий варьировала от 56 до 580 тыс. кл. мл–1, органотрофных бактерий – от 0 до 546 КОЕ мл–1, ОМЧ – от 0 до 401 КОЕ мл–1 (табл. 1 и 1S). Значения ОЧМ не превышали данных, полученных в 2007–2008 гг. при анализе разных слоев ледяных кернов в литорали и пелагиали в юго-западной части Байкала (Бондаренко и соавт., 2013). Колиформные бактерии встречались в единичных количествах, и лишь в речных образцах Большой Черемшаной количество ОБ (546 КОЕ мл–1) было сопоставимо с общим микробным числом мезофильных аэробных и факультативно анаэробных микроорганизмов (ОМЧ 401 и 345 КОЕ мл–1 соответственно), культивированных на среде с высоким содержанием органического субстрата. Следует отметить, что микроорганизмы ледяных сообществ в реках юго-восточной части озера более активны – средняя величина интенсивности суммарной ТАУ для ледяных образцов этой части озе-ра (0.43 мкг C л−1 сут−1) была в 7 раз выше, чем среднее для юго-западных станций (0.06 мкг C л−1 сут−1) (табл. 2). Тем не менее, величины интенсивности ТАУ в ледяных образцах были существенно (в 2–32 раза) ниже, в сравнении с подледной водой. В процессе ТАУ от 0.01 до 0.37 мкг C л−1 сут−1 переходило в биомассу бактериопланктона и от 0.02 до 0.47 мкг C л−1 сут−1 фиксировалось во внеклеточном РОУ (табл. 2). Анализ данных с помощью метода главных компонент не выявил в образцах достоверного влияния рН и РОВ на ОЧМ, ОБ и ОМЧ (рис. 2а, 2б). Положительная сопряженность отмечена лишь между рН и значениями ТАУ (r = 0.61 для БМ и 0.5 для РОУ).
Таблица 2.
Темновая ассимиляция углекислоты в ледовых кернах из рек и приустьевых акваторий Южного Байкала
| № станции | Темновая ассимиляция углекислоты | Р | В | Р/В | ||||
|---|---|---|---|---|---|---|---|---|
| БМ | РОУ | общая | БМ | РОУ | мкг С л–1 сут–1 | (по С) | ||
| мкг С л–1 сут–1 | % от общего | мкг С л–1 сут–1 | ||||||
| Ст. 1 | Река Переемная | |||||||
| Устье | 0.06 | 0.05 | 0.11 | 56 | 44 | 1.26 | 1.26 | 1.00 |
| Озеро | 0.09 | 0.06 | 0.15 | 59 | 41 | 1.74 | 1.47 | 1.18 |
| Ст. 2 | Река Солзан | |||||||
| Устье | 0.05 | 0.05 | 0.11 | 51 | 49 | 1.08 | 1.62 | 0.67 |
| Озеро | 0.06 | 0.04 | 0.10 | 56 | 44 | 1.13 | 2.46 | 0.46 |
| Ст. 3 | Река Малая Осиновка | |||||||
| Устье | 0.33 | 0.42 | 0.75 | 44 | 56 | 6.54 | 1.11 | 5.89 |
| Озеро | 0.30 | 0.33 | 0.62 | 48 | 52 | 5.93 | 1.68 | 3.53 |
| Ст. 4 | Река Большая Осиновка | |||||||
| Устье | 0.25 | 0.47 | 0.72 | 35 | 65 | 5.05 | 2.55 | 1.98 |
| Озеро | 0.27 | 0.46 | 0.73 | 37 | 63 | 5.42 | 3.81 | 1.42 |
| Ст. 5 | Фоновая станция напротив г. Байкальск | |||||||
| Ф | 0.37 | 0.19 | 0.56 | 66 | 34 | 7.48 | 2.48 | 3.02 |
| Ср. ю-в | Средние величины для юго-восточной части оз. Байкал | |||||||
| 0.20 | 0.23 | 0.43 | 46 | 54 | 3.96 | 2.05 | 1.93 | |
| Ст. 6 | Река Большая Черемшаная | |||||||
| Устье | 0.02 | 0.02 | 0.04 | 45 | 55 | 0.36 | 0.86 | 0.42 |
| Озеро | 0.02 | 0.03 | 0.05 | 45 | 55 | 0.44 | 1.04 | 0.43 |
| Ст. 7 | Река Крестовка | |||||||
| Устье | 0.03 | 0.05 | 0.08 | 40 | 60 | 0.64 | 0.89 | 0.72 |
| Озеро | 0.03 | 0.05 | 0.08 | 39 | 61 | 0.64 | 0.68 | 0.94 |
| Ст. 8 | Река Каменушка | |||||||
| Устье | 0.02 | 0.03 | 0.05 | 41 | 59 | 0.45 | 2.04 | 0.22 |
| Озеро | 0.02 | 0.03 | 0.05 | 44 | 56 | 0.46 | 0.88 | 0.52 |
| Ст. 9 | Фоновая станция напротив п. Листвянка | |||||||
| ЛФ | 0.01 | 0.03 | 0.04 | 25 | 75 | 0.22 | 0.85 | 0.26 |
| Ср. ю-з | Средние величины для юго-западной части оз. Байкал | |||||||
| 0.02 | 0.03 | 0.06 | 40 | 60 | 0.46 | 1.04 | 0.44 | |
| Среднее | Средние величины для всей исследованной акватории | |||||||
| 0.11 | 0.14 | 0.26 | 43 | 57 | 2.22 | 1.46 | 1.52 | |
Минерализация воды в исследуемых реках и приустьевых акваториях варьировала от 33 до 126 мг л–1) (табл. 1). Вода насыщена кислородом (12.7–14.3 O2 мг л–1), что обеспечивает интенсивность деструкционных процессов. Значения ПО, БО и РОВ сопряжены, коэффициенты корреляции имели высокую достоверность (табл. 4S ). Воды исследуемых притоков характеризуются как слабокислые, нейтральные или щелочные. Содержание РОВ в реках и приустьевых акваториях юго-восточной части озера Байкал варьировало от 1.03 до 2.74 мг л–1, юго-западной – от 0.85 до 4.85 мг л–1(табл. 2S).
В воде большинства исследованных рек ОЧМ, численность ОБ и ОМЧ соответствуют значениям, отмечаемым в пелагиали озера (Максимова, Максимов, 1979, табл. 1). По количеству микроорганизмов выделяются воды рек и приустьевых акваторий Большой Черемшаной и Крестовки, где регистрируются высокие значения ОЧМ (1396–3973 тыс. кл. мл–1), ОБ (до 6680 кл. мл–1) и ОМЧ (до 6301 КОЕ). На этих станциях отмечены значительные количества колиформных бактерий (1000 КОЕ в 100 мл) и спор сульфитредуцирующих бактерий.
Анализ распределения в толще вод ОБ и санитарно-значимых бактерий в акватории р. Каменушка показал высокие количества в поверхностных слоях приустьевой акватории и на разных глубинах водной толщи по направлению в открытый Байкал на расстоянии до 3 км от устья (рис. 1S). Аналогичные профили распределения с максимальными значениями в трофогенном слое характерны для мезофильных аэробных и факультативно анаэробных бактерий, культивированных при 22°С (до 120 КОЕ мл−1), тогда как при 37°С культивирования отмечались единичные колонии (не более 4 кл мл−1) по всему водному столбу. Анализ численности исследуемых показателей в речных водах и в приустьевых акваториях большинства исследованных рек, за исключением р. Крестовка, показал, что уже на расстоянии 50 м от устьев значения показателей соответствуют отмечаемым на фоновых станциях.
Темновая ассимиляция углекислоты (ТАУ) в водных образцах из юго-восточной части Байкала варьировала от 0.28 до 1.78 мкг С л−1 сут−1 (при среднем значении – 1 мкг С л−1 сут−1). Величины меньшие, чем средние, зафиксированы на станциях в акваториях рек Солзан и Переемная. Выше средних показатели интенсивности микробной ассимиляции углекислоты отмечены на фоновой станции в этой части озера, а максимальные значения обнаружены в районе рек Малая и Большая Осиновка (табл. 3). Следует также отметить, что в устьях рек этой оконечности Байкала активность ТАУ была ниже, чем в зоне смешения речной и озерной вод. Аналогичная закономерность отмечена ранее при исследовании зоны смешения реки Верхняя Ангара (Максименко и соавт., 2012). На прибрежных станциях юго-западной части озера данная закономерность справедлива только для интенсивности продукции биомассы бактериопланктона. При этом средняя величина общей ТАУ для станций юго-западной части (1 мкг С л−1 сут−1) полностью совпадала со средними значениями для юго-восточной и, соответственно, средней величиной для всех обследованных точек (табл. 3). В воде исследованных акваторий в процессе ТАУ от 18 до 46% углерода бикарбоната включалось в биомассу бактериопланктона и 54–82% ассимилировалось в РОУ. В среднем для водных проб исследованных районов только треть ассимилированного углерода бикарбоната была детектирована в биомассе, а две трети во внеклеточном растворенном органическом веществе. В пробах ледяных кернов это соотношение было более равномерным, при средних величинах 43% в БМ бактериопланктона и 57% в РОУ (табл. 2). А интервал величин включенного углерода бикарбоната составил 25–66% в биомассу и 34–75% в растворенное органическое вещество.
Таблица 3.
Темновая ассимиляция углекислоты в воде рек и приустьевых акваторий Южного Байкала
| № станции | Темновая ассимиляция углекислоты | Р | В | Р/В | ||||
|---|---|---|---|---|---|---|---|---|
| БМ | РОУ | общая | БМ | РОУ | мкг С л–1 сут–1 | (по С) | ||
| мкг С л–1 сут–1 | % от общего | мкг С л–1 сут–1 | ||||||
| Ст. 1 | Река Переемная | |||||||
| Устье | 0.09 | 0.19 | 0.28 | 32 | 68 | 1.77 | 1.23 | 1.44 |
| Озеро | 0.07 | 0.28 | 0.35 | 21 | 79 | 1.48 | 1.67 | 0.89 |
| Ст. 2 | Река Солзан | |||||||
| Устье | 0.20 | 0.40 | 0.60 | 33 | 67 | 3.94 | 2.07 | 1.91 |
| Озеро | 0.35 | 0.41 | 0.75 | 46 | 54 | 6.93 | 5.37 | 1.29 |
| Ст. 3 | Река Малая Осиновка | |||||||
| Устье | 0.28 | 0.84 | 1.13 | 25 | 75 | 5.69 | 2.09 | 2.72 |
| Озеро | 0.66 | 0.80 | 1.46 | 45 | 55 | 13.23 | 15.27 | 0.87 |
| Ст. 4 | Река Большая Осиновка | |||||||
| Устье | 0.50 | 0.91 | 1.40 | 35 | 65 | 9.92 | 2.72 | 3.65 |
| Озеро | 0.69 | 1.09 | 1.78 | 39 | 61 | 13.71 | 16.79 | 0.82 |
| Ст. 5 | Фоновая станция напротив г. Байкальск | |||||||
| Ф | 0.60 | 0.77 | 1.36 | 44 | 56 | 11.92 | 15.63 | 0.76 |
| Ср. ю-в | Средние величины для юго-восточной части оз. Байкал | |||||||
| 0.38 | 0.63 | 1.01 | 38 | 62 | 7.62 | 6.98 | 1.09 | |
| Ст. 6 | Река Большая Черемшаная | |||||||
| Устье | 0.22 | 0.35 | 0.57 | 39 | 61 | 4.43 | 10.60 | 0.42 |
| Озеро | 0.32 | 1.29 | 1.61 | 20 | 80 | 6.34 | 24.51 | 0.26 |
| Ст. 7 | Река Крестовка | |||||||
| Устье | 0.23 | 1.07 | 1.30 | 18 | 82 | 4.66 | 11.39 | 0.41 |
| Озеро | 0.24 | 0.77 | 1.01 | 23 | 77 | 4.76 | 12.25 | 0.39 |
| Ст. 8 | Река Каменушка | |||||||
| Устье | 0.19 | 0.48 | 0.66 | 28 | 72 | 3.71 | 10.94 | 0.34 |
| Озеро | 0.28 | 0.35 | 0.63 | 44 | 56 | 5.50 | 11.07 | 0.50 |
| Ст. 9 | Фоновая станция напротив п. Листвянка | |||||||
| ЛФ | 0.43 | 0.90 | 1.33 | 32 | 68 | 8.61 | 8.76 | 0.98 |
| Ср. Ю-з | Средние величины для юго-западной части оз. Байкал | |||||||
| 0.27 | 0.74 | 1.02 | 27 | 73 | 5.44 | 12.79 | 0.43 | |
| Среднее | Средние величины для всей исследованной акватории | |||||||
| 0.33 | 0.68 | 1.01 | 33 | 67 | 6.70 | 9.95 | 0.67 | |
Микробное окисление метана. Содержание метана в подледной воде на разных станциях варьировало от 8 до 993 нмоль л−1. Максимальные величины выявлены в устьях рек Переемная, Каменушка (614 и 993 нмоль л−1 соответственно) и приустьевой зоне реки Крестовка (1975 нмоль л−1). Следует отметить, что эти значения были выше отмечаемых в пелагиали Байкала даже в районах разгрузки газообразных и жидких углеводородов (Захаренко и соавт., 2015). Интенсивность микробного окисления метана была невысокой и полностью коррелировала с его концентрацией в пробах (табл. 3S). Разброс величин суммарной интенсивности метанокисления изменялся от 0.35 до 51.68 нл СН4 л−1 сут−1 , среднее значение – 6.85 нл СН4 л−1 сут−1 для всей исследованной акватории. Доля углерода метана (в процентах от суммарного МО), перешедшего в продукты окисления (в СО2, БМ и РОУ), не зависела ни от концентрации метана, ни от интенсивности его окисления. В среднем по акватории доли углерода метана, выявляемого после его микробного окисления в углекислоте, биомассе и растворенном органическом веществе, составили 40, 10 и 50% соответственно. При этом разброс величин доли углерода метана (в процентах от общего), обнаруживаемого внутри каждого из продуктов метанокисления, был незначителен: от 20 до 50% – в углекислоте, от 7 до 19% – в биомассе и от 40 до 60% – в растворенном органическом веществе.
ОБСУЖДЕНИЕ
Как показали наши исследования, реки Южного Байкала и их приустьевые акватории различаются по химическому составу, значениям рН, содержанию РОВ и микробиологическим показателям между собой, а также отличаются от отмечаемых значений в пелагиали озера (табл. 1, рис. 2). Известно, что в зонах смешения речных и озерных рек происходят как выпадение минеральной и биологической взвесей, так и активные процессы трансформации органических веществ, существенный вклад в которые вносят микробные сообщества (Гордеев, Лисицин, 1978; Мицкевич, Намсараев, 1994). В приустьевых акваториях рек нами зарегистрированы высокие количества бактерий, участвующих в деструкции легко минерализуемых органических веществ, и, на некоторых станциях, активные процессы ТАУ и МО. Как и отмечено ранее (Ходжер и соавт., 2002; Сороковикова и соавт., 2002; Оболкин и соавт., 2013; Томберг и соавт., 2016), одними из факторов, влияющих на гидрохимические показатели в реках Южного Байкала, являются поступление атмосферных осадков, обогащенных выбросами расположенных поблизости ТЭЦ, а также состав пород и почв, с которыми соприкасаются воды рек. В частности, пониженные значения рН и низкая минерализация вод в реке Переемная и ее акватории обусловлены протеканием вод по дренирующим кислым бескарбонатным породам. Но, несмотря на закисление снегового покрова и вод этой реки, отрицательного воздействия этого процесса на численность микроорганизмов и на продукционные характеристики не наблюдается. По данным измерений ТАУ и Р/В коэффициента, микробные сообщества в акватории этой реки находятся в активном состоянии.
Водные образцы реки Солзан и ее приустьевой акватории, а также образцы рек Большая и Малая Осиновки группируются в другой кластер. По значениям рН и сумме ионов вода в этих реках сопоставима с характерными для пелагиали озера значениями (табл. 1), лишь в устьях рек отмечаются повышенные концентрации некоторых ионов (сульфата, нитрата, нитрита). Известно (Государственный доклад…, 2017 г.), что после закрытия Байкальского ЦБК в этом районе регистрируется загрязнение подземных вод стоками из карт шлам-лигнина, оказывающих отрицательное воздействие на их качество. Отличительной особенностью этих акваторий являются высокие значения ТАУ в ледовых образцах и воде устьев рек в сравнении с приустьевой зоной. Причем высокие значения коэффициента Р/В (удельной эффективности бактериопланктона) при существенно меньшей ОЧМ свидетельствуют о высоком проценте метаболически активных клеток, обеспечивающих включение углерода не только в биомассу бактериопланктона, но и в другие органические соединения.
Еще один кластер включает воды фоновых и приустьевых акваторий некоторых (рис. 2в, 2г) рек, где компонентный состав вод сходен с составом воды в пелагиали озера (Грачев и соавт., 2004; Khodzher et al., 2016, 2018). В устьях рек этих акваторий интенсивность ТАУ и численность бактериопланктона ниже отмечаемой в зонах смешения речной и озерной вод. Ранее было показано, что одним из ключевых медиаторов темновой ассимиляции углекислоты бактериопланктоном может быть низкая доступность лабильного органического углерода (Alonso-Saez et al., 2010). Вполне вероятно, что для образцов этого кластера, с низкими значениями ХПК и небольшим вкладом органотрофных бактерий в ОЧМ, определяющим интенсивность продукционных процессов фактором является доступность субстрата.
Воды рек и приустьевых акваторий Большой Черемшаной и Крестовки, где также отмечено закисление снежного покрова за счет выбросов оксида азота (Obolkin et al., 2016) от крупных ТЭЦ Иркутска и Ангарска, не подвергаются ацидификации в такой степени, как воды реки Переемная. Воды этих рек и приустьевых акваторий характеризуются по данным ТАУ и Р/В низкой активностью микробных сообществ и высокими значениями санитарно-значимых бактерий, что свидетельствует об интенсивном поступлении в реки легко минерализуемого органического вещества, в том числе фекальных загрязнений. Это согласуется с данными о массовом развитии в реках Крестовка и Б. Черемшаная представителей нанопланктонных жгутиковых родов Cryptomonas и Chlamydomonas, являющихся видами-индикаторами загрязнения (Bondarenko et al., 2012). Наличие E. coli в приустьевых районах рек и в пелагиали отмечалось в летний период и в 80-е годы прошлого столетия, при более низкой рекреационной нагрузке (Максимова, Максимов, 1989; Максимов, 1995). Высокий коли-индекс отмечался в реках Большая Осиновка, Солзан, как входящих в акваторию, подверженную поступлению сточных вод от Байкальского ЦБК и хозяйственно-бытовых стоков г. Байкальска (Максимова, Максимов, 1989; Максимов, 1995). В настоящий период наиболее высокая рекреационная нагрузка наблюдается в западной части Южного Байкала, куда с водами рек поступают хозяйственно-бытовые сточные воды из поселков, не оснащенных централизованными очистными сооружениями. Ранее было экспериментально доказано выживание и развитие при низких температурах патогенных и условно-патогенных бактерий в байкальской воде (Панасюк и соавт., 2002). Согласно исследованиям Schang et al. (2018), E. coli может сохраняться до 24 сут в осадках прибрежной зоны эстуариев, взмучивание которых приводило к увеличению в водной толще количества E. coli в 20 раз. Эти данные объясняют обнаружение колиформных бактерий в пелагиали озера Байкал, куда они могут поступать из поверхностных осадков устьевых акваторий рек при штормовых ветрах. Вместе с тем, анализ данных измерений ТАУ свидетельствует, что с речными водами в литораль Байкала поступают микроорганизмы, не способные проявлять активность в изменившихся условиях. Для сравнения можно привести результаты измерения ТАУ подледных сообществ в Баренцовом море и меромиктических озерах Шира и Шунет (Саввичев и соавт., 2000, 2005), где при низкой ОЧМ и в отсутствие антропогенных воздействий отмечены более высокие значения этого показателя. Очевидно, что в приустьевых акваториях рек Южного Байкала с повышенной антропогенной нагрузкой происходит изменение продукционного потенциала микробиоценозов, и, таким образом, их роль как “биологического фильтра” на пути к пелагиали озера Байкал снижается.
Список литературы
Барам Г.И., Верещагин А.Л., Голобокова Л.П. Применение микроколоночной высокоэффективной жидкостной хроматографии с УФ-детектированием для определения анионов в объектах окружающей среды // Аналитическая химия. 1999. Т. 54. С. 962–965.
Baram G.I., Vereshchagin A.L., Golobokova L.P. Microcolumn high-performance liquid chromatography with UV detection for the determination of anions in environmental materials // J. Analyt. Chem. 1999. V. 54. P. 854–857.
Большаков А.М., Егоров А.В. Об использовании методики фазово-равновесной дегазации при газометрических исследованиях // Океанология. 1987. Т. 27. № 5. С. 861–862.
Bolshakov A.M., Egorov A.V. Using a phase-equilibrium degassing method for gasometric studies // Oceanology. 1987. V. 27. P. 861–862.
Бондаренко Н.А., Белых О.И., Голобокова Л.П., Артемьева О.В., Логачева Н.Ф., Тихонова И.В., Липко И.А., Косторнова Т.Я., Парфенова В.В., Ходжер Т.В. Химический состав, бактериальные и водорослевые сообщества льда озера Байкал // Гидробиол. журн. 2013. № 1. С. 14–28.
Вотинцев К.К. Гидрохимия озера Байкал. М.: Изд-во АН СССР, 1961. 310 с.
Вотинцев К.К., Глазунов И.В., Толмачева А.П. Гидрохимия рек бассейна озера Байкал. М.: Наука, 1965. 495 с.
Государственный доклад о состоянии и об охране окружающей среды Иркутской области в 2016 году / Министерство природных ресурсов и экологии Иркутской обл. Иркутск: ООО “Мегапринт”, 2017. 274 с.
Грачев М.А. О современном состоянии экологической системы озера Байкал / Под ред. проф. Эджингтона Д.Н. Новосибирск: Изд-во СО РАН, 2002. 156 с.
Грачев М.А., Домышева В.М., Ходжер Т.В., Коровякова Н.В., Голобокова Л.П., Погодаева Т.В., Верещагин А.Л., Гранин Н.Г., Гнатовский Р.Ю., Косторнова Т.Я. Глубинная вода озера Байкал – природный стандарт пресной воды // Химия в интересах устойчивого развития. 2004. № 12. С. 417–429.
Горбенко Ю.А. О наиболее благоприятном количестве сухого питательного агара в средах для культивирования морских микроорганизмов // Микробиология. 1961. Т. 30. № 1. С. 168–172.
Гордеев В.В., Лисицин А.П. Средний химический состав взвеси рек мира и питание океанов речным осадочным материалом // Докл. АН СССР. 1978. Т. 238. № 1. С. 255–258.
Гранина Л.З. Ранний диагенез донных осадков озера Байкал. Новосибирск: Акад. изд-во “ГЕО”, 2008. 159 с.
Дрюккер В.В., Косторнова Т.Я., Моложавая О.А., Афанасьев В.А. Оценка качества воды оз. Байкал по санитарно-бактериологическим показателям // География и природные ресурсы. 1993. № 1. С. 60–64.
Захаренко А.С., Пименов Н.В., Иванов В.Г., Земская Т.И. Окисление метана в водной толще районов газо- и нефтепроявлений Среднего и Южного Байкала // Микробиология. 2015. Т. 84. № 1. С. 98–106.
Zakharenko A.S., Pimenov N.V., Ivanov V.G., Zemskaya T.I. Detection of methane in the water column at gas and oil seep sites in central and southern Lake Baikal // Microbiology (Moscow). 2015. V. 84. P. 90–97.
Максименко С.Ю., Земская Т.И., Павлова О.Н., Иванов В.Г., Бурюхаев С.П. Микробное сообщество водной толщи на биогеохимическом барьере “река Селенга-озеро Байкал” // Микробиология. 2008. Т. 77. № 5. С. 660–667.
Maksimenko S.Y., Zemskaya T.I., Pavlova O.N., Ivanov V.G., Buryukhaev S.P. Microbial community of the water column of the Selenga River-Lake Baikal biogeochemical barrier // Microbiology (Moscow). 2008. V. 77. P. 587–594.
Максименко С.Ю., Иванов В.Г., Бурюхаев С.П., Шимараев М.Н., Земская Т.И. Структура микробных сообществ в зоне впадения р. Верхняя Ангара в оз. Байкал // Гидробиол. журн. 2012. Т. 48. № 2. С. 55–68.
Максимов В.В. Микробиология крупных рек Байкала и их приустьевых акваторий. Автореферат дис. … канд. биол. наук, Иркутск, 1995. 25 с.
Максимова Э.А., Максимов В.Н. Микробиология вод Байкала. Иркутск: Изд-во Иркут. ун-та, 1989. 168 с.
Мицкевич И.Н., Намсараев Б.Б. Численность и распределение бактериопланктона в Карском море в сентябре 1993 г. // Морская биология. 1994. Т. 34. № 5. С. 704–708.
Mitskevich I.N., Namsaraev B.B. The distribution of bacterioplankton in the Kara Sea in September 1993 // Oceanology. 1995. V. 34. P. 641–645.
Намсараев Б.Б., Дулов Л.Е., Дубинина Г.А., Земская Т.И., Гранина Л.З., Карабанов Е.Б. Участие бактерий в процессах синтеза и деструкции органического вещества в микробных матах озера Байкал // Микробиология. 1994. Т. 63. № 2. С. 345–351.
Namsaraev B.B., Dulov L.E., Dubinina G.A., Zemskaya T.I., Granina L.Z., Karabanov E.B. Bacterial synthesis and destruction of organic-matter in microbial mats of Lake Baikal // Microbiology (Moscow). 1994. V. 63. P. 193–197.
Нецветаева О.Г., Чипанина Е.В., Оболкин В.А., Зимник Е.А., Сезько Н.П., Лопатина И.Н., Ходжер Т.В. Особенности химии атмосферных осадков cтанций Листвянка (Иркутская область) и Приморская (Приморский край) в 2005–2011 гг. // Оптика атмосферы и океана. 2013. Т. 26. № 6. С. 466–471.
Панасюк Е.Ю., Дрюккер В.В., Парфенова В.В. Изучение влияния низкой температуры и популяции автохтонной микрофлоры на выживаемость патогенных и потенциально-патогенных бактерий в воде в условиях эксперимента // Медицинский научно-производственный журнал “Сибирь-Восток”. 2002. Т. 3 (51, март). С. 4–7.
Практикум по микробиологии: Учебное пособие для студентов высших учебных заведений / Под ред. Нетрусова А.И. М.: Издательский центр “Академия”, 2005. 608 с.
Русанов И.И., Саввичев А.С., Юсупов С.К., Пименов Н.В., Иванов М.В. Образование экзометаболитов в процессе микробного окислении метана в морских экосистемах // Микробиология. 1998. Т. 67. № 5. С. 710–717.
Rusanov I.I., Savvichev A.S., Yusupov S.K., Pimenov N.V., Ivanov M.V. Production of exometabolites in the microbial oxidation of methane in marine ecosystems // Microbiology (Moscow). 1998. V. 67. P. 590–596.
Саввичев А.С., Русанов И.И., Пименов Н.В., Мицкевич И.Н., Байрамов И.Т., Лейн А.Ю., Иванов М.В. Микробиологические исследования северной части Баренцева моря в начале зимнего сезона // Микробиология. 2000. Т. 69. № 6. С. 819–830.
Savvichev A.S., Rusanov I.I., Pimenov N.V., Mitskevich I.N., Bairamov I.T., Lein A.Y., Ivanov M.V. Microbiological explorations in the northern part of the Barents Sea in early winter // Microbiology (Moscow). 2000. V. 69. P. 698–708.
Саввичев А.С., Русанов И.И., Рогозин Д.Я., Захарова Е.Е., Лунина О.Н., Брянцева И.А., Юсупов С.К., Пименов Н.В., Дегерменджи А.Г., Иванов М.В. Микробиологические и изотопно-геохимические исследования меромиктических озер Хакасии в зимний сезон // Микробиология. 2005. Т. 74. № 4. С. 552–561.
Savvichev A.S., Rusanov I.I., Rogozin D.Y., Zakharova E.E., Lunina O.N., Bryantseva I.A., Yusupov S.K., Pimenov N.V., Degermendzhi A.G., Ivanov M.V. Microbiological and isotopic-geochemical investigations of meromictic lakes in Khakasia in winter // Microbiology (Moscow). 2005. V. 74. P. 477–485.
Сороковикова Л.М., Синюкович В.Н., Коровякова И.В., Голобокова Л.П., Погодаева Т.В., Нецветаева О.Г. Формирование химического состава воды притоков Южного Байкала в современных условиях // География и природные ресурсы. 2002. № 4. С. 52–57.
Сороковикова Л.М., Синюкович В.Н., Томберг И.В., Маринайте И.И., Ходжер Т.В. Оценка качества вод притоков озера Байкал по химическим показателям // География и природные ресурсы. 2015. Т. 36. № 1. С. 37–45.
Sorokovikova L.M., Sinyukovich V.N., Tomberg I.V., Marinaite I.I., Khodzher T.V. Assessing the water quality in the tributary streams of Lake Baikal from chemical parameters // Geography and Natural Resources. 2015. V. 36. P. 31–39.
Тимошкин О.А., Бондаренко Н.А., Волкова Е.А., Томберг И.В., Вишняков В.С., Мальник В.В. Массовое развитие зелeных нитчатых водорослей родов Spirogyra Link и Stigeoclonium Kutz. (Chlorophyta) в прибрежной зоне Южного Байкала // Гидробиол. журн. 2014. № 5. С. 15–26.
Томберг И.В., Сороковикова Л.М., Нецветаева О.Г., Сезько Н.П., Жученко Н.А. Химический состав и тенденция закисления снеговых вод и вод притоков Южного Байкала // Оптика атмосферы и океана. 2016. Т. 29. № 6. С. 516–520.
Ханаев И.В., Дзюба Е.В., Кравцова Л.С., Грачев М.А. Влияние массового развития зеленых нитчатых водорослей на воспроизводство желтокрылки Cottocomephorus grewingkii (Dybowski, 1874) (Cottidae) в условиях экологического кризиса озера Байкал // Докл. Академии наук (Общая биология). 2016. Т. 467. № 1. С. 119–121.
Khanaev I.V., Dzyuba E.V., Kravtsova L.S., Grachev M.A. The effect of bloom of filamentous green algae on the reproduction of yellowfin sculpin Cottocomephorus grewingkii (Dybowski, 1874) (Cottoidae) during ecological crisis in Lake Baikal // Dokl. Biol. Sci. 2016. V. 467. № 1. P. 63–64.
Хлыстов О.М., Горшков А.Г., Егоров А.В., Земская Т.И., Гранин Н.Г., Калмычков Г.В., Воробьева С.С., Павлова О.Н., Якуп М.А., Макаров М.М., Москвин В.И., Грачев М.А. Нефть в озере мирового наследия // ДАН. 2007. Т. 414. № 5. С. 656–659.
Khlystov O.M., Gorshkov A.G., Zemskaya T.I., Granin N.G., Vorob’eva S.S., Pavlova O.N., Yakup M.A., Makarov M.M., Grachev M.A., Egorov A.V., Kalmychkov G.V., Moskvin V.I. Oil in the lake of world heritage // Dokl. Earth Sci. 2007. V. 415. № 1. P. 682–685.
Ходжер Т.В., Семенов М.Ю., Оболкин В.А., Домышева В.М., Голобокова Л.П., Кобелева Н.А., Нецветаева О.Г., Потемкин В.Л., Сергеева М.В. Мониторинг кислотных выпадений в Байкальском регионе // Химия в интересах устойчивого развития. 2002. Т. 10. № 5. С. 699–705.
Штыкова Ю.Р., Суслова М.Ю., Косторнова Т.Я., Суханова Е.В., Пестунова О.С., Горшкова А.С., Галачьянц А.Д., Парфенова В.В. Мониторинг санитарно-микробиологического состояния пелагиали озера Байкал и устьев впадающих в него крупных рек с 2010 по 2015 г. // Изв. Иркутского гос. ун-та. Сер. “Биология. Экология”. 2016. Т. 17. С. 51–62.
Alonso-Saez L., Galand P.E., Casamayor E.O., Pedros-Alio C., Bertilsson S. High bicarbonate assimilation in the dark by Arctic bacteria // ISME J. 2010. V. 4. P. 1581–1590.
Bondarenko N.A., Belykh O.I., Golobokova L.P., Artemyeva O.V., Logacheva N.F., Tikhonova I.V., Lipko I.A., Kostornova T.Ya., Parfenova V.V., Khodzher T.V., Ahn T.-S., Zo Y.-G. Stratified distribution of nutrients and extremophile biota within freshwater ice covering the surface of Lake Baikal // J. Microbiol. 2012. V. 50. P. 8–16.
Cooper L.W., Savvichev A.S., Grebmeier J.M. Abundance and production rates of heterotrophic bacterioplankton in the context of sediment and water column processes in the Chukchi Sea // Oceanography. 2015. V. 28. № 3. P. 84–99.
Falkner K.K., Measures C.I., Herbelin S.E., Edmond J.M., Weiss R.F. The major and minor element geochemistry of Lake Baikal // Limnol. Oceanogr. 1991. V. 36. P. 413–423.
ISO 9308-1:2000. Drinking Water. Detection and Enumeration of Escherichia coli and Coliform Bacteria. Part 1. Membrane Filtration Method. Geneva: International Standards Organization, 2000.
Khodzher T.V., Domysheva V.M., Sorokovikova L.M., Golobokova L.P. Part II. Method and case studies for understanding and monitoring the landscapes of Siberia. Ch. 3. Methods for monitoring the chemical composition of Baikal water // Novel Methods for Monitoring and Managing Land and Water Resources in Siberia / Eds. Mueller L., Sheudshen A.K., Eulenstein F. New York: Springer, 2016. P. 113–132.
Khodzher T.V., Domysheva V.M., Sorokovikova L.M., Tomberg I.V., Sakirko M.V. Hydrochemical studies in Lake Baikal: history and nowadays // Limnol. Freshwater Biol. 2018. № 1. P. 2–9.
Lemarchand K., Masson L., Brousseau R. Molecular biology and DNA microarray technology for microbial quality monitoring of water // Crit. Rev. Microbiol. 2004. V. 30. P. 145–172.
Morris D.P., Lewis W.M. Nutrient limitation of bacterioplankton growth in Lake Dillon, Colorado // Limnol. Oceanogr. 1992. V. 37. P. 1179–1192.
Obolkin V., Khodzher T., Sorokovikova L., Tomberg I., Netsvetaeva O., Golobokova L. Effect of long-range transport of sulphur and nitrogen oxides from large coal power plants on acidification of river waters in the Baikal region, East Siberia // Int. J. Environ. Stud. 2016. V. 73. P. 452–461.
Pimenov N.V., Bonch-Osmolovskaya E.A. In situ activity studies in thermal environments // Methods in Microbiology / Eds. Rainey F.A., Oren A. Amsterdam: Elsevier Ltd., 2006. V. 14. P. 29–53.
Porter K.G. The use of DAPI for identifying and counting aquatic microflora // Limnol. Oceanogr. 1980. V. 25. P. 943–948.
Sakirko M.V., Domysheva V.M., Pestunov D.A., Netsvetaeva O.G., Panchenko M.V. Concentration of nutrients in the water of Southern Baikal in summer // Proc. SPIE. 2015. V. 9680. № 968045. P. 1–7.
Schang C., Lintern A., Cook P.L.M., Rooney G., Coleman R., Murphy H.M., Deletic A., McCarthy D. Escherichia coli survival and transfer in estuarine bed sediments // River Res. Applicat. 2018. V. 34. P. 606–614.
Sorokin Yu.I. Comparative evaluation of isotopic methods for estimation of microbial production in the sea // Archives de Hydrobiologie Beigh Ergebnis Limnologie. 1990. V. 34. P. 153–164.
Stewart J.R., Santo Domingo J.W., Wade T.J., Sadowsky M.J. Microbial Source Tracking // Fecal Pollution, Public Health, and Microbial Source Tracking. / Eds. Santo Domingo J. and Sadowsky M Washington: Amer. Soc. Microbiol, 2007. Ch. 1. P. 1‒32.
Stoeckel D.M., Harwood V.J. Performance, design, and analysis in microbial source tracking studies // Appl. Environ. Microbiol. 2007. V. 73. P. 2405–2415.
Straskrabova V., Izmest’yeva L.R., Maksimova E.A., Fietz S., Nedoma J., Boroveca J., Kobanova G.I., Shchetinina E.V. Pislegin primary production and microbial activity in the euphotic zone of Lake Baikal (Southern Basin) during late winter // Global and Planetary Change. 2005. V. 46. P. 57–73.
Surbeck C.Q., Jiang S.C., Grant S.B. Ecological control of fecal indicator bacteria in an urban stream // Environ. Sci. Technol. 2010. V. 44. P. 631–637.
Timoshkin O.A., Samsonov D.P., Yamamuro M., Moore M.V., Belykh O.I., Malnik V.V., Sakirko M.V., Shirokaya A.A., Bondarenko N.A., Domysheva V.M., Fedorova G.A., Kochetkov A.I., Kuzmin A.V., Lukhnev A.G., Medvezhonkova O.V., Nepokry-tykh A.V., Pasynkova E.M., Poberezhnaya A.E., Potaps-kaya N.V., Rozhkova N.A., Sheveleva N.G., Tikhonova I.V., Timoshkina E.M., Tomberg I.V., Volkova E.A., Zaitseva E.P., Zvereva Yu.M., Kupchinsky A.B., Bukshuk N.A. Rapid ecological change in the coastal zone of Lake Baikal (East Siberia): Is the site of the world’s greatest freshwater biodiversity in danger? // J. Great Lakes Res. 2016. V. 42. P. 487–497.
Toothman B.R., Cahoon L.B., Mallin M.A. Phosphorus and carbohydrate, limitation of fecal coliform and fecal enteroccoccus within tidal creek sediments // Hydrobiologia. 2009. V. 636. P. 401–412.
Yoshioka T., Ueda S., Khodzher T., Bashenkhaeva N., Tomberg I., Gorbunova L., Khanaev I., Zhdanov A. Dissolved organic matter in Lake Baikal and its watershed // Limnology. 2002. V. 3. P. 159–168.
Wanjugi P., Fox G.A., Harwood V.J. The interplay between predation, competition, and nutrient levels influences the survival of Escherichia coli in aquatic environments // Microb. Ecol. 2016. V. 72. P. 1–12.
Wetzel, R.G. and Likens, G.E., Limnological Analyses, New York: Springer, 1991.
Дополнительные материалы отсутствуют.