

Молекулярная биология, 2023, T. 57, № 1, стр. 3-9
Интерлейкин-11 при патологиях нервной системы
М. И. Айрапетов a, b, С. О. Ереско b, c, *, П. Д. Игнатова b, А. А. Лебедев b, Е. Р. Бычков b, П. Д. Шабанов b, d
a Санкт-Петербургский государственный педиатрический медицинский университет
194100 Санкт-Петербург, Россия
b Институт экспериментальной медицины,
197376 Санкт-Петербург, Россия
c Санкт-Петербургский государственный химико-фармацевтический университет
197101 Санкт-Петербург,, Россия
d Военно-медицинская академия им. С.М. Кирова Министерства обороны Российской Федерации
194044 Санкт-Петербург, Россия
* E-mail: erescko.sergei@yandex.ru
Поступила в редакцию 08.05.2022
После доработки 15.07.2022
Принята к публикации 15.07.2022
- EDN: AYDLGW
- DOI: 10.31857/S0026898423010020
Аннотация
Изучение роли цитокинов при различных патологических состояниях организма – одно из актуальных направлений современной биомедицины. В 1990 году в фиброцитоподобных стромальных клетках костного мозга обнаружили интерлейкин-11 (IL-11), повышенный интерес к которому наблюдается в последнее время. IL-11 локально экспрессируется нервными клетками и участвует в патогенетических механизмах формирования ряда заболеваний нервной системы. В представленном обзоре обобщены данные, указывающие на участие IL-11 в развитии патологий головного мозга. Вероятно, этот цитокин сможет найти применение в коррекции механизмов, связанных с формированием патологических состояний нервной системы.
ВВЕДЕНИЕ
Интерлейкин-11 (IL-11, он же AGIF) – белок, который относится к семейству IL-6-подобных цитокинов, впервые выделили в 1990 г. из фиброцитоподобных стромальных клеток костного мозга [1]. Первоначально показали, что IL-11 играет важную роль в процессах кроветворения, особенно в созревания мегакариоцитов [1]. В дальнейшем выявили участие этого цитокина и в других процессах.
IL-11 либо самостоятельно, либо вместе с другими цитокинами (IL-3, IL-4, IL-7, IL-12, IL-13, SCF, Flt-3 и GM-CSF) стимулирует гемопоэз мегакариоцитов, участвует в созревании В-клеток и активации Т-клеток, в пролиферации синовиальных фибробластов [1–3]. Кроме того, обнаружены негемопоэтические эффекты IL-11. Например, IL-11 обладает противовоспалительной активностью и оказывает защитное действие на эпителий слизистой оболочки желудочно-кишечного тракта и дыхательных путей [4]. Высокий уровень экспрессии IL-11 в дыхательных путях наблюдается при вирусиндуцированной бронхиальной астме, а сверхэкспрессия приводит к ремоделированию дыхательных путей у мышей, воспалению и к появлению симптомов, сходных с симптомами астмы [5]. Передача сигналов от IL-11 имеет решающее значение для Th2-опосредованного воспалительного ответа в легких, а ингибирование передачи сигналов IL-11 снижает активацию провоспалительных путей [6]. Таким образом, регуляцию путей передачи сигналов от IL-11 можно рассматривать как потенциальную терапевтическую мишень при заболеваниях легких. Другая широко известная функция IL-11 – способность ингибировать адипогенез (отсюда второе название этого цитокина – AGIF, adipogenesis inhibitory factor) [7].
Получены многочисленные данные о том, что IL-11 вовлечен в процессы нейрогенеза [8], нейровоспаления и нейродегенерации [3, 4, 9–12]. За последние годы показано, что IL-11 вовлечен в развитие патологических состояний нервной системы. Роль IL-11 при болезни Альцгеймера, аутоиммунном энцефаломиелите, рассеянном склерозе, при ишемическом повреждении головного мозга и других заболеваниях изучена на животных моделях. В нашем обзоре обобщены данные о вовлеченности IL-11 в молекулярные нейроиммунные механизмы при патологических состояниях нервной системы.
IL-11 И ВНУТРИКЛЕТОЧНЫЕ ПУТИ ПЕРЕДАЧИ СИГНАЛА
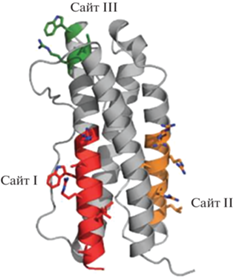
IL-11 – белок с молекулярной массой ~19 кДа, предшественник которого состоит из 199 аминокислотных остатков, включая лидерный пептид из 21 остатка. Ген IL-11, состоящий из пяти экзонов и четырех интронов, расположен на хромосоме 19 человека. IL-11 человека, крысы и мыши имеет высокую степень гомологии (более 80%), обогащен остатками пролина и не содержит остатков цистеина, что означает отсутствие дисульфидных связей во вторичной структуре. IL-11, как и другие члены семейства IL-6-подобных цитокинов, имеет форму пучка из четырех α-спиралей. Во взаимодействии с рецептором участвует его С-концевая область (рис. 1) [3, 13–16].
Рис. 1.
Структура IL-11. Отмечены сайты связывания цитокина с рецепторным комплексом. Рисунок построен по данным V. Barton и соавт. [17].
IL-11 продуцируется клетками многих тканей [18]. В 1993 г. IL-11 обнаружили в нейронах гиппокампа, где он стимулирует пролиферацию прогениторных клеток, и показали тем самым его участие в процессах нейрогенеза [8]. Стало очевидным, что в головном мозге экспрессируются также компоненты рецепторного комплекса IL-11 (IL-11Rα, gp130) [3, 4, 9–12, 19–21]. Экспрессия IL-11 выявлена в обонятельной луковице, амигдале, базальных ганглиях, таламусе, среднем мозге, мосте, продолговатом мозге, гиппокампе, коре головного мозга, мозжечке [18–21]. Иммуногистохимическим методом выявлена локализация ключевого компонента рецепторного комплекса – белка gp130 (он же IL6ST) – в головном мозге крысы [19]. При этом отмечена иммунореактивность gp130 как в глиальных, так и в нейрональных клетках. Окрашивание нейронов выявило две различные картины. На одной видно преимущественное окрашивание нейропилей в структурах конечного мозга, включая гиппокамп, кору головного мозга и хвостатое ядро. На другой окрашивание наблюдалась на цитоплазматической мембране тел нейронов, преимущественно в нижнем стволе мозга, в крупных нейронах ретикулярной формации, а также в спинномозговых и черепно-мозговых мотонейронах [19]. Электронно-микроскопический анализ показал, что оба типа иммунореактивности gp130 в основном связаны с цитоплазматической мембраной, но точно не локализуются в синаптических участках [19].
С помощью РНК-seq-анализа мРНК IL-11, IL-11Rα и gp130 выявили также в различных структурах головного мозга. При этом в незначительных количествах мРНК IL-11 представлена во всех типах клеток нервной системы; мРНК IL-11Rα и gp130 в наибольшем количестве обнаружена в астроцитах, микроглии и эндотелии [20, 21].
В настоящее время отсутствуют полученные с высоким разрешением структурные данные для сигнального комплекса IL-11, хотя имеются указания на сходство структур IL-11Rα и IL-6Rα. Методом иммунопреципитации показано, что сигнальные комплексы как IL-11, так и IL-6 имеют гексамерную структуру [17]. С помощью мутагенеза на IL-11 идентифицированы сайты I, II и III (рис. 1). Показано, что во взаимодействии IL-11 с рецепторным комплексом участвуют три сайта связывания (I, II и III) (рис. 1), расположенные аналогично сайтам I, II и III в IL-6. Сайт I позволяет IL-11 связываться с IL-11R, тогда как сайты II и III опосредуют связывание с gp130 [17]. Это стало еще одним фактом в пользу предположения о том, что активные сигнальные комплексы IL-6 и IL-11 используют в целом сходный механизм [17].
Взаимодействие IL-11 с рецепторным комплексом служит сигналом к активации JAK/SHP2-зависимого пути внутриклеточной передачи сигнала. Другой путь сигналинга начинается с фосфорилирования белков STAT1 и STAT3, что ведет к изменению скорости транскрипции генов Socs3 и Smad7. Синтезируемый белок Socs3 может, в свою очередь, ингибировать активность киназы JAK. Белок Smad7 регулирует экспрессию антиапоптотических, пролиферативных и проангиогенных генов [3, 13–16]. При этом стоит отметить, что IL-11 немного длиннее, чем IL-6. Это указывает на различия в способе связывания и геометрии сигнального комплекса [22, 23], а внеклеточный домен IL-11Rα имеет более гидрофобный характер, чем IL-6Rα, что предполагает существование различий в механизмах взаимодействия цитокинов IL-11 и IL-6 с рецепторным аппаратом [24].
Цитокины, в том числе IL-11, могут продуцироваться различными клетками, тогда как рецепторные субъединицы экспрессируются в более ограниченном наборе клеток, что определяет более специфичное влияние цитокинов на отдельные популяции клеток, которые могут активироваться непосредственно IL-11 [25]. Так, gp130 экспрессируется повсеместно, при этом IL-11Rα представлен на меньшем числе клеток, хотя этот рецептор более распространен, чем IL-6R. IL-11R выявлен на многих типах клеток, включая мегакариоциты, макрофаги, Т-клетки, остеокласты, гепатоциты, эпителиальные и эндотелиальные клетки [25]. По данным РНК-seq мРНК IL-11R экспрессируется в астроцитах, микроглии и эндотелии [20, 21].
Кроме того, часть рецепторного комплекса IL-11Rα может локализоваться не только на мембране клетки, но и существовать в растворимом виде (рис. 2) [25]. Металлопротеаза ADAM10, а также сериновые протеазы, нейтрофильная эластаза и протеиназа 3 могут отщеплять эктодомен от IL-11Rα, а образовавшийся в ходе отщепления эктодомен представляет собой растворимую форму рецептора – sIL-11R (soluble IL-11R). sIL-11R проявляет биологическую активность и связывает IL-11 в межклеточном пространстве, увеличивая число типов клеток, которые могут активироваться IL-11, но не экспрессируют собственный мембранный рецептор IL-11Rα. Такой путь передачи сигнала может реализовываться нейронами головного мозга, в которых на высоком уровне экспрессируется белок и мРНК gp130, но на крайне низком – мРНК IL-11Rα [20, 21].
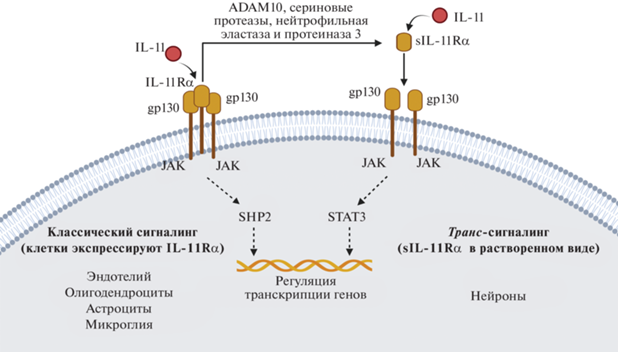
Участие IL-11 в нескольких путях внутриклеточной передачи сигнала делает его плейотропным цитокином, вызывающим порой противоположные эффекты, однако выбор сигнального пути, как это часто бывает, определяется множеством причин и в зависимости от того или иного состояния организма может усиливаться либо замедляться тот или иной внутриклеточный путь передачи сигнала. Несмотря на то, что IL-11 играет значимую роль в нервной ткани, внутриклеточные сигнальные пути этого цитокина все еще остаются неизученными и представляют интерес для дальнейших исследований.
ИНТЕРЛЕЙКИН-11 ПРИ ПАТОЛОГИЯХ НЕРВНОЙ СИСТЕМЫ
Показано, что IL-11 способен “корректировать” воспалительные пути в эпителиальных тканях дыхательной системы, где развиваются основные события при острой респираторной инфекции, вызываемой коронавирусом SARS-CoV-2. Рекомбинантный IL-11 используют в настоящее время для повышения числа тромбоцитов при миелодиспластическом синдроме [26]. Подтверждена эффективность и безопасность IL-11 при лимфосаркомах и солидных опухолях [27], а также при злокачественных опухолях женских половых органов [28]. Можно рассчитывать, что дальнейшее изучение IL-11 приведет к более широкому применению этого цитокина в клинической практике.
В этом разделе обобщены данные, которые показывают роль IL-11 в механизмах, связанных с развитием патологий головного мозга (табл. 1). В одном из первых исследований, проведенном на культуре клеток нейробластомы (модель болезни Альцгеймера), показано участие IL-11 в механизмах развития этой патологии [2]. При болезни Альцгеймера наблюдается отложение агрегатов β-амилоида в головном мозге, а повышенная выработка β-амилоидного пептида считается одним из ранних событий в патогенезе этого заболевания, приводящего к прогрессирующей нейродегенерации и деменции. Показано, что β-амилоид активирует L-фосфосеринфосфатазу в нейрональных клетках (клетки нейробластомы крысы B104), а IL-11 дозозависимым образом (5, 50, 100 нг/мл) ингибирует вызываемую при этом нейротоксичность. Предположили, что L-фосфосеринфосфатаза может участвовать в изменении клеточного метаболизма при болезни Альцгеймера путем усиления нейротоксичности, тогда как IL-11 может действовать через свою рецепторную систему как нейропротектор [2].
Обнаружено повышение содержания IL-11 в астроцитах и IL-11R на олигодендроцитах при рассеянном склерозе. В ходе исследований, проведенных на культурах астроцитов и олигодендроцитов, сделан вывод, что путь IL-11/IL-11R может участвовать в защите олигодендроцитов. Так, добавление рекомбинантного IL-11 (10 нг/мл) к культуре клеток спинного мозга плода человека привело к увеличению числа олигодендроцитов, повышенному ветвлению олигодендроглии, снижению апоптотической гибели клеток; кроме того, IL-11 потенцировал образование миелина [29]. В последующих исследованиях наблюдали развитие выраженного нейровоспаления в тканях спинного мозга у мышей с нокаутом гена IL-11Rα (IL-11Rα−/− и IL-11Rα+/−), которое характеризовалось инфильтрацией макрофагами с последующей демиелинизацией и снижением числа нервных клеток. Это данные свидетельствуют о значимости IL-11Rα в регуляции нейровоспаления [10].
Влияние экзогенного IL-11 оценивали на культуре прогениторных олигодендроцитов (OPC) крысы. Применение IL-11 повышало выживаемость клеток, уменьшая число клеток, гибнущих по пути апоптоза, и потенцируя процесс деления клеток. Кроме того, экзогенный IL-11 способен снижать активацию CD4 лимфоцитов, воздействуя на клетки CD11c++. IL-11 может регулировать продукцию эффекторных цитокинов, воздействуя на клетки CD11c++ [10]. Изучено также влияние рекомбинантного IL-11 при аутоиммунном энцефаломиелите, который моделировали у мышей. После появления признаков заболевания животным вводили рекомбинантный IL-11 в дозе 25 или 50 мкг/кг/сут. Оказалось, что IL-11 в дозе 50 мкг/кг/сут снижал тяжесть заболевания: наблюдалось снижение степени демиелинизации и потери олигодендроцитов, а также воспаления и числа CD3 лимфоцитов. Более низкие дозы IL-11 не оказывали значимого эффекта [10].
Повышение концентрации IL-11 выявлено в ликворе и сыворотке пациентов с рассеянным склерозом. Сообщается, что основным источником IL-11 в крови являются клетки CD4+, а содержание таких клеток (IL-11+CD4+) повышено у пациентов с рассеянным склерозом. Предполагается, что повышенный уровень IL-11 может индуцировать дифференцировку и пролиферацию клеток Th17, так как IL-11-индуцированная дифференцировка Th17 ингибировалась антителом к IL-11. При этом антитело к IL-6 не ингибировало дифференцировку клеток Th17, т.е. IL-11-индуцированная дифференцировка Th17 не зависит от IL-6. Участие клеток Th17 в развитии рассеянного склероза подтверждается результатами нескольких исследований. Клетки Th17 человека служат источником эффекторных цитокинов IL-17A, IL-17F, IL-21, IL-22 и TNF-α, которые опосредуют продукцию воспалительных цитокинов, хемокинов и металлопротеиназ, а также способствуют привлечению гранулоцитов и лимфоцитов к местам воспаления. Миграцию клеток в ЦНС обеспечивает рецептор хемокинов CCR6. Высокий уровень экспрессии CCR6 на IL-11+ CD4+ клетках может опосредовать их миграцию в ЦНС, что объясняет повышение уровня IL-11 в спинномозговой жидкости пациентов [30].
Появление очагов демиелинизации в белом веществе больных рассеянным склерозом считается результатом скоординированного аутоиммунного процесса, который включает активацию клеток микроглии в нервной ткани (местные резидентные макрофаги). На купризоновой модели рассеянного склероза у мышей, показано, что сверхэкспрессия IL-11, вызванная локальной доставкой в очаг повреждения гена IL-11 с помощью вирусного вектора, снижала степень демиелинизации с одновременным ускорением процесса ремиелинизации. Поскольку известно, что микроглия участвует в фагоцитозе миелина, полученные результаты могут указывать на способность IL-11 вносить коррективы в этот процесс. Однако механизм того, каким образом микроглия участвует в эффектах, вызванных IL-11, еще предстоит выяснить. В эксперименте in vitro на клеточной линии микроглии BV2 показано [11], что добавление рекомбинантного IL-11 (1–30 нг/мл) дозозависимо уменьшало степень фагоцитоза миелина. Кроме того, использование IL-11 привело к увеличению толщины миелинового слоя, что указывает на способность IL-11 усиливать процесс ремиелинизации, а не только замедлять демиелинизацию, однако механизм этого еще предстоит изучить [11].
Демиелинизация в купризоновой модели рассеянного склероза также может зависеть от секреции микроглией провоспалительных цитокинов, токсичных для олигодендроцитов и нервных клеток. Опубликованы данные о способности IL-11 ингибировать синтез TNF-α, IL-1β, IL-12, IL-6 и продукцию NO активированными макрофагами [31, 32], однако уровень провоспалительных медиаторов в этом эксперименте не указан.
Таблица 1.
Участие IL-11 в патологиях нервной системы
| Патологическое состояние | Объект | Функции IL-11 | Источник |
|---|---|---|---|
| Болезнь Альцгеймера | Культура нейробластомы крысы B104 | Выполняет нейропротекторную роль, ингибируя L-фосфосеринфосфатазу | [2] |
| Рассеянный склероз | Пациенты | Повышение IL-11 в астроцитах, IL-11R на олигодендроцитах, индуцирует дифференцировку и пролиферацию Th17, независимо от IL-6 | [11, 29, 30] |
| Первичная культура астроцитов и олигодендроцитов | Рекомбинантный IL-11 человека повышает число олигодендроцитов, снижает апоптоз, активирует ремиелинизацию | ||
| Мышь, культура клеток микроглии BV2 | Ограничивает пролиферацию и способность микроглии к фагоцитозу миелина, замедляет демиелинизацию, способствует ускорению процесса ремиелинизации | ||
| Аутоиммунный энцефаломиелит | Мышь, культура клеток OPCs | Замедляет нейродегенерацию, снижая потери олигодендроцитов и нейронов, снижает нейровоспаление и уровень цитотоксических лейкоцитов | [10] |
| Ишемия | Мышь | Снижает активацию нейроглии, провоспалительные цитокины (IL-1β, IL-6, TNFβ) и окислительный стресс в мозге, повышает уровень противовоспалительного цитокина TGF-β1 | [12] |
| Медуллобластома | Культура клеток медуллобластомы | Повышает химиорезистентность | [33] |
Изучено также участие IL-11 в ишемическом повреждении мозга, которое моделировали на мышах с помощью окклюзии средней мозговой артерии. Показано снижение экспрессии мРНК и белка IL-11 в первые 24 ч ишемии с дальнейшим постепенным его увеличением. Внутривенное введение рекомбинантного IL-11 в эксперименте выявило противовоспалительный эффект этого цитокина в дозах 20 и 40 мкг/кг, который приводил к снижению маркеров активации астроцитов и микроглии, а также уровня мРНК провоспалительных цитокинов (IL-1β, IL-6, TNF-β). При этом отмечено повышение уровня мРНК противовоспалительного цитокина (TGF-β1), уровня супероксид-дисмутазы и снижение уровня малонового диальдегида, что свидетельствует о снижении окислительного стресса в тканях мозга [12].
Стоит обратить внимание еще на один факт. В одном из недавних исследований показано, что цитокины семейства IL-6 могут опосредовать повышение химиорезистентности клеток медуллобластомы. Культуры клеток медуллобластомы, кондиционированные in vitro цитокинами IL-6, OSM, LIF или IL-11, проявляют повышенную активность JAK1/STAT3-сигнализации, при этом развивается резистентность к ряду препаратов. Применение ингибиторов gp130 или ингибиторов JAK-киназ позволило эффективно преодолеть резистентность экспрессирующих gp130 клеток медуллобластомы к винкристину [33]. Полученные данные могут указывать на ограниченную применимость рекомбинантного IL-11, однако необходимы дальнейшие исследования in vivo, а также изучение опосредованного влияния цитокинов семейства IL-6 на другие виды опухолевых клеточных линий.
ЗАКЛЮЧЕНИЕ
IL-11 впервые выделили из фиброцитоподобных стромальных клеток костного мозга еще в 1990 г. и тогда же его нашли в головном мозге, однако о функциях этого цитокина в нервной системе известно совсем немного. IL-11 обладает нейропротекторными и противовоспалительными свойствами, замедляет скорость развития нейровоспаления и апоптотической гибели нервных клеток, которая наблюдается при различных патологических состояниях нервной системы [34, 35].
IL-11 активирует процессы нейрогенеза (ускоряет скорость деления прогениторных нейронов и олигодендроцитов), стимулирует процесс ремиелинизации аксонов при рассеянном склерозе. Необходимы дополнительные исследования, которые расширят наши знания о роли этого цитокина при патологии центральной нервной системы.
Исследование выполнено за счет средств ИЭМ в рамках государственного задания по теме “Фармакологический анализ действия нейротропных средств, изучение внутриклеточных мишеней и создание систем направленной доставки”, шифр 0557-2019-0004.
Настоящая статья не содержит каких-либо исследований с участием людей или животных в качестве объектов исследования.
Авторы заявляют об отсутствии конфликта интересов.
Список литературы
Paul S.R., Bennett F., Calvetti J.A., Kelleher K., Wood C.R., O’Hara R.M., Jr., Leary A.C., Sibley B., Clark S.C., Williams D.A., Yang Y.C. (1990) Molecular cloning of a cDNA encoding interleukin 11, a stromal cell derived lymphopoietic and hematopoietic cytokine. Proc. Natl. Acad. Sci. USA. 87, 7512–7516.
Heese K., Nagai Y., Sawada T. (2000) Induction of rat L-phosphoserine phosphatase by amyloid-beta (1-42) is inhibited by interleukin-11. Neurosci. Lett. 288(1), 37–40.
Гук К.Д., Купраш Д.В. (2011) Интерлейкин-11, член семейства IL-6-подобных цитокинов. Молекуляр. биология. 45(1), 44–55.
Wilde M.I., Faulds D. (1998) Oprelvekin: a review of its pharmacology and therapeutic potential in chemotherapy-induced thrombocytopenia. BioDrugs. 10(2), 159–171.
Einarsson O., Geba G.P., Zhu Z., Landry M., Elias J.A. (1996) Interleukin-11: stimulation in vivo and in vitro by respiratory viruses and induction of airways hyperresponsiveness. J. Clin. Investig. 97(4), 915–924.
Tang W., Geba G.P., Zheng T., Ray P., Homer R.J., Kuhn C. (1996) Targeted expression of IL-11 in the murine airway causes lymphocytic inflammation, bronchial remodeling, and airways obstruction. J. Clin. Investig. 98(12), 2845–2853.
Keller D.C., Du X.X., Srour E.F., Hoffman R., Williams D.A. (1993) Interleukin-11 inhibits adipogenesis and stimulates myelopoiesis in human long-term marrow cultures. Blood. 82(5), 1428–1435.
Mehler M.F., Rozental R., Dougherty M., Spray D.C., Kessler J.A. (1993) Cytokine regulation of neuronal differentiation of hippocampal progenitor cells. Nature. 362(6415), 62–65.
Leng S.X., Elias J.A. (1997) Interleukin-11. Int. J. Biochem. Cell. Biol. 29, 1059–1062.
Gurfein B.T., Zhang Y., López C.B. (2009) IL-11 regulates autoimmune demyelination. J. Immunol. 183(7), 4229–4240.
Maheshwari A., Janssens K., Bogie J., Van Den Haute C., Struys T., Lambrichts I., Baekelandt V., Stinissen P., Hendriks J.J., Slaets H., Hellings N. (2013) Local overexpression of interleukin-11 in the central nervous system limits demyelination and enhances remyelination. Mediators Inflamm. 2013, 685317.
Zhang B., Zhang H.-X., Shi S.-T., Bai Y.-L., Zhe X., Zhang S.-J., Li Y.-J. (2019) Interleukin-11 treatment protected against cerebral ischemia/reperfusion injury. Biomed. Pharmacother. 115, 108816.
Горшкова Е.А., Недоспасов С.А., Шилов Е.С. (2016) Эволюционная пластичность цитокинов семейства IL-6. Молекуляр. биология. 50(6), 1039–1048.
McKinley D., Wu Q., Yang-Feng T., Yang Y.C. (1992) Genomic sequence and chromosomal location of human interleukin-11 gene (IL11). Genomics. 13(3), 814–819.
Yang Y.C., Yin T. (1992) Interleukin-11 and its receptor. Biofactors. 4(1), 15–21.
Stephen F.A., Thomas L.M., Alejandro A.S., Jinghui Z., Zheng Z., Webb M., David J.L. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs, Nucl. Acids Res. 25, 3389–3402.
Barton V., Hall M., Hudson K.R., Heath J.K. (2000) Interleukin-11 signals through the formation of a hexameric receptor complex. J. Biol. Chem. 275(46), 36197–36203.
Knut & Alice Wallenberg Foundation. IL-11. The Human Protein Atlas.
Watanabe D., Yoshimura R., Khalil M., Yoshida K., Kishimoto T., Taga T., Kiyama H. (1996) Characteristic localization of gp130 (the signal-transducing receptor component used in common for IL-6/IL-11/CNTF/LIF/OSM) in the rat brain. Eur. J. Neurosci. 8(8), 1630–1640.
Zhang Y., Chen K., Sloan S.A., Bennett M.L., Scholze A.R., O’Keeffe S., Phatnani H.P., Guarnieri P., Caneda C., Ruderisch N., Deng S., Liddelow S.A., Zhang C., Daneman R., Maniatis T., Barres B.A., Wu J.Q. (2014) An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J. Neurosci. 34(36), 11929–11947.
Zhang Y., Sloan S.A., Clarke L.E., Caneda C., Plaza C.A., Blumenthal P.D., Vogel H., Steinberg G.K., Edwards M.S., Li G., Duncan J.A., Cheshier S.H., Shuer L.M., Chang E.F., Grant G.A., Gephart M.G., Barres B.A. (2016) Purification and characterization of progenitor and mature human astrocytes reveals transcriptional and functional differences with mouse. Neuron. 89(1), 37–53.
Barton V., Hudson K.R., Heath J.K. (1999) Identification of three distinct receptor binding sites of murine interleukin-11. J. Biol. Chem. 274(9), 5755–5761.
Tacken I., Dahmen H., Boisteau O., Minvielle S., Jacques Y., Grötzinger J. (1999) Definition of receptor binding sites on human interleukin-11 by molecular modeling-guided mutagenesis. Eur. J. Biochem. 265(2), 645–655.
Metcalfe R.D., Aizel K., Zlatic C.O., Nguyen P.M., Morton C.J., Lio. (2020) The structure of the extracellular domains of human interleukin 11 α-receptor reveals mechanisms of cytokine engagement. J. Biol. Chem. 295(24), 8285–8301.
Lokau J., Agthe M., Flynn C.M., Garbers C. (2017) Proteolytic control of interleukin-11 and interleukin-6 biology. Biochim. Biophys. Acta Mol. Cell. Res. 1864(11 Pt B), 2105–2117.
Montero A.J., Estrov Z., Freireich E.J., Khouri I.F., Koller C.A., Kurzrock R. (2006) Phase II study of low dose interleukin 11 in patients with myelodysplastic syndrome. Leuk. Lymphoma. 47, 2049–2054.
Hatae M., Ariyoshi Y., Fukuoka M., Furuse K., Noda K., Hirabayashi K., Yakushiji M., Ogawa M., Takaku F. (2005) An early phase II clinical study of YM 294 (rhlL 11) in patients with solid tumors and malignant lymphoma. Gan To Kagaku Ryoho. 32, 489–496.
Hatae M., Noda K., Yamamoto K., Ozaki M., Hiraba yashi K., Nishida T., Ohashi Y., Ogawa M., Takaku F. (2005) A clinical study of YM 294 (rhlL 11) in patients with gynecologic cancer. Gan To Kagaku Ryoho. 32, 479–487.
Zhang Y., Taveggia C., Melendez-Vasquez C., Einheber S., Raine C.S., Salzer J.L., Brosnan C.F., John G.R. (2006) Interleukin-11 potentiates oligodendrocyte survival and maturation, and myelin formation. J. Neurosci. 26(47), 12174–12185.
Zhang X., Tao Y., Chopra M., Dujmovic-Basuroski I., Jin J., Tang Y., Drulovic J., Markovic-Plese S. (2015) IL-11 induces Th17 cell responses in patients with early relapsing-remitting multiple sclerosis. J. Immunol. 194(11), 5139–5149.
Trepicchio W.L., Wang L., Bozza M., Dorner A.J. (1997) IL-11 regulates macrophage effector function through the inhibition of nuclear factor-κB. J. Immunol. 159(11), 5661–5670.
Leng S.X., Elias J.A. (1997) Interleukin-11 inhibits macrophage interleukin-12 production. J. Immunol. 159(5), 2161–2168.
Sreenivasan L., Li L.V., Leclair P., Lim C.J. (2022) Targeting the gp130/STAT3 axis attenuates tumor microenvironment mediated chemoresistance in group 3 medulloblastoma cells. Cells. 11, 381.
Airapetov M., Eresko S., Lebedev A., Bychkov E., Shabanov P. (2021) The role of Toll-like receptors in neurobiology of alcoholism. BioSci. Trends. 15(2), 74–82.
Peterson P.K., Toborek M. (2014) Neuroinflammation Neurodegeneration. New York: Springer.
Дополнительные материалы отсутствуют.
Инструменты
Молекулярная биология