

Журнал общей биологии, 2019, T. 80, № 6, стр. 427-438
Проблемы применимости концепции модульной и структурно-функциональной организации цветковых растений к анализу структуры побегов у некоторых групп споровых растений
А. Г. Лапиров 1, *, Е. А. Беляков 1, 2
1 Институт биологии внутренних вод им. И.Д. Папанина РАН
152742 Некоузский р-н, Ярославская обл., пос. Борок, 109, Россия
2 Череповецкий государственный университет
162600 Череповец, просп. Луначарского, 5, Россия
* E-mail: lapir@ibiw.yaroslavl.ru
Поступила в редакцию 23.11.2018
После доработки 05.06.2019
Принята к публикации 20.08.2019
Аннотация
В работе рассмотрен вопрос применимости концепции модульной и структурно-функциональной организации к водным споровым растениям. В качестве последних исследованы Isoёtes lacustris L. и I. echinospora Durieu (Isoëtopsida: Isoëtaceae), Equisetum palustre L. и E. fluviatile L. (Equisetopsida: Equisetaceae), Salvinia natans (L.) All. (Polypodiopsida: Salviniaceae), Marsilea quadrifolia L. (Polypodiopsida: Marsiliaceae). Показано, что при изучении жизненных форм водных споровых растений использование трех категорий модулей (элементарного, универсального и основного), применяемых для цветковых растений, вызывает ряд вопросов. Основные из них связаны с рассмотрением строения элементарного модуля. Наибольшее внимание уделено таким его особенностям, как наличие (или отсутствие) спорообразующих элементов листа и пазушных либо внепазушных структур. Предлагаемая авторами трактовка этих позиций в характеристике элементарного модуля для водных споровых растений следующая: 1) лист с наличием или отсутствием спорообразующих элементов (для споровых растений, не обладающих аксиллярным ветвлением, за исключением хвощей); 2) внепазушные элементы (споровые растения, не обладающие аксиллярным ветвлением) – боковые побеги, формирующиеся из обособленных меристем или из подобных участков со скрытыми меристематическими свойствами, расположенные в определенной позиции по отношению к листьям, заканчивающиеся спорообразующими элементами (хвощи), либо без них (лептоспорангиатные папоротники, полушники). Проблем с выделением универсального и основного модуля у водных споровых растений практически нет. Выделение структурно-функциональных зон для водных споровых растений затруднено, поскольку их набор подробно разработан лишь для цветковых растений. Наибольшее сходство с последними в этом смысле отмечено для E. fluviatile и E. palustre. Однако и в этом случае необходимо расширить понятие “зона обогащения”, а вместо характерной для цветковых растений “зоны главного соцветия” ввести термин “зона главного (терминального) стробила”. Кроме того, для споровых (по аналогии с цветковыми) растений авторы предлагают использовать термины “полиспорогенный” и “моноспорогенный побег”.
Массовое изучение структурной организации растений в России и за рубежом началось в середине прошлого столетия (Савиных, 2008). При этом наибольшее внимание до недавнего времени уделялось цветковым растениям, в том числе различным представителям водных и прибрежно-водных растений (Лебедева, 2006; Лелекова, 2006; Петухова, 2008; Шабалкина, Савиных, 2011; Беляков, Лапиров, 2015; Беляков, 2016; Лапиров и др., 2017). Между тем водные споровые растения, входящие в состав ценотических комплексов водоемов и водотоков, с точки зрения структурно-функциональной организации (СФО) до сих пор исследованы слабо (Антонова, Лагунова, 1999; Шорина, 2001, 2007; Храпко, 2010; Державина и др., 2011).
Под водными споровыми растениями в данной работе понимаются представители класса полушниковые, или шильниковые (Isoëtopsida), порядка Isoëtales, рода полушник, или шильник, – Isoëtes; класса хвощовые (Equisetopsida), порядка Equisetales, рода хвощ – Equisetum (Филин, 1978а, б); а также класса полиподиопсиды (Polypodiopsida), порядков марсилеевые (Marsileales, рода марсилея – Marsilea) и сальвиниевые (Salviniales, рода сальвиния – Salvinia) (Махлин, Сурова, 1978а, б). Растения, входящие в состав этих родов, – все спороносные или бессемянные; исторически для них использовали собирательные термины “pteridophyta” (“птеридофиты”) и “ferns and fern allies” (папоротники и близкие к ним группы растений) – парафилетический комплекс растений (Smith et al., 2006).
Выбор именно этих групп растений для изучения СФО не случаен. Так, марсилеевые и сальвиниевые – два водных гетероспоровых семейства папоротников – давно вызывают пристальный интерес морфологов (Russow, 1872; Campbell, 1888; Sadebeck, 1902; Allsopp, 1963 – все цит. по: Schneider, Pryer, 2002). Они имеют очень много общих черт с водными покрытосеменными, среди которых упрощенные сосудистые системы, полиморфные вегетативные признаки и высокоспециализированные репродуктивные системы (Schneider, Pryer, 2002). Все сальвинии – растения, плавающие на поверхности стоячих и медленно текущих вод, а марсилеи – амфибийные растения, некоторые из которых большую часть жизни проводят в погруженном состоянии, другие – часть жизни проводят в воде, часть на суше, а третьи – преимущественно сухопутные растения (Махлин, Сурова, 1978а, б). Особую группу составляют современные хвощи – “…последние дошедшие до нас представители древнего и обособленного отдела членистых растений…, поэтому их изучение важно для познания биологии и эволюции высших растений в целом” (Скворцов, 2008, с. 3). Equisetum palustre и E. fluviatile – голарктические виды сезонного климата, с более слабыми проявлениями диморфизма побегов. Эти виды относятся к Palustre-типу, Palustre-подтипу и являются одними из немногих представителей влажных местообитаний (Скворцов, 2008).
Важное место в рассмотрении СФО принадлежит и р. Isoëtes L., представленному в Северной и Центральной Европе двумя погруженными видами – I. lacustris L. и менее многочисленным I. echinospora Durieu (Vöge, 2014). Это растения, относящиеся к древнему таксону Lycopodiophyta, известному с раннего девона, т.е. приблизительно 390 млн лет назад, сохранившие неизменными в течение длительного времени многие черты, позволяющие сравнивать общую организацию ископаемых и современных растений (Мейен, 1987; Yatsentyuk et al., 2001).
Вопрос, поставленный во главу угла данного исследования, предполагает анализ применимости концепции модульной и структурно-функциональной организации цветковых растений к структуре побегов у некоторых групп водных и околоводных споровых растений, изученных с этих позиций крайне слабо. В этом отношении (в отличие от остальных групп споровых растений) больше всего “повезло” наземным папоротникам, сопоставление модульной организации которых с модулями цветковых растений описано в ряде отечественных работ (Антонова, Лагунова, 1999; Гуреева, 2001; Шорина, 2001, 2007; Храпко, 2010; Державина и др., 2011).
МАТЕРИАЛЫ И МЕТОДЫ
В качестве объектов исследований выбраны виды, принадлежащие к отдельным таксономическим группам водных и околоводных споровых растений: Isoёtes lacustris L. и I. echinospora Durieu (р. Isoёtes L.), Equisetum palustre L. и E. fluviatile L. (р. Equisetum L.), а также ряд папоротников – Marsilea quadrifolia L. (р. Marsilea), Salvinia natans (L.) All. (р. Salvinia).
Морфологию полушников озерного и щетинистого (60 и 40 растений соответственно) исследовали на живом материале, собранном на озерах Тверской, Ярославской, Нижегородской и Московской областей в 2015–2018 гг. Для уточнения морфологических особенностей детально исследовали 130 гербарных листов (гербарий IBIW RAS). Морфологию побеговых систем хвощей болотного и приречного изучали на гербарном материале (просмотрено около 100 гербарных листов, IBIW) с территории Европейской России (в том числе и по сборам авторов с территорий Ярославской, Тверской, Нижегородской и других областей). Детально морфологию и ритм сезонного развития E. fluviatile удалось рассмотреть на основе гербарных сборов В.В. Богачёва (83 растения) за период 1968–1969 гг. При исследовании морфологии папоротников – марсилеи четырехлистной и сальвинии плавающей – использовали гербарные образцы (25 и 93 гербарных листа соответственно, главным образом с территории Нижней Волги), хранящиеся в IBIW. Кроме того, при написании работы авторами рассмотрен и проанализирован обширный материал, имеющийся как в отечественных, так и зарубежных научных работах, позволивший уточнить различные детали морфологии выбранных растений.
Структуру побеговых систем водных и околоводных споровых растений исследовали при помощи классического сравнительного морфологического метода (Серебряков, 1952). При характеристике модульной организации исследуемых растений применяли три категории модулей (Савиных, 2007, 2008): элементарный (ЭМ), универсальный (УМ) и основной (ОМ). ЭМ выделялись в зависимости от длины междоузлия, характерных особенностей листа и пазушных структур (если таковые имеются), наличия либо отсутствия придаточных корней. Структурно-функциональный анализ осуществляли с помощью комплекса различных подходов (Troll, 1964; Мусина, 1976; Серебрякова, Петухова, 1978; Серебрякова, Павлова, 1986; Борисова, Попова, 1990; Савиных, 2008; Савиных, Мальцева, 2008), на основании которых был рассмотрен спектр структурно-функциональных зон побегового тела растения.
РЕЗУЛЬТАТЫ
С учетом того, что “…анализ модулей в качестве конструктивных частей тела растений расширяет возможности сравнительной морфологии…” (Шорина, 2001, с. 129), ниже представлены структурные особенности всех типов модулей у изучаемых нами растений.
У представителей р. Isoёtes L. (I. lacustris и I. echinospora) элементарный модуль (ЭМ) представлен укороченным метамером (участок корневища), узлом с отходящим от него шиловидным листом и придаточными корнями (либо без них) (табл. 1). При этом в основании шиловидного листа может находиться либо мегаспорофилл, либо микроспорофилл. Кроме того, лист может быть стерилен, т.е. не нести спорофиллов. Забегая немного вперед, отметим, что наличие или отсутствие тех или иных вариантов ЭМ формирует разные типы универсального модуля.
Таблица 1.
Характеристика и структурные особенности ЭМ Isoёtes lacustris и I. echinospora
| Схематическое изображение ЭМ | Характеристика ЭМ | ||
|---|---|---|---|
 |
– укороченный метамер со стерильным листом; | ||
 |
– укороченный метамер с листом и мегаспорангием; | ||
 |
– укороченный метамер с листом и микроспорангием | ||
| Схематическое изображение элемента ЭМ | Описание схематического изображения элемента | Схематическое изображение элемента ЭМ | Описание схематического изображения элемента |
 |
– укороченный метамер; |  |
– мегаспорангий; |
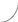 |
– шиловидный лист; | 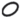 |
– микроспорангий |
У марсилеи четырехлистной (Marsilea quadrifolia) выделено восемь типов элементарных модулей (табл. 2). ЭМ представлен следующими элементами:
Таблица 2.
Характеристика и структурные особенности ЭМ Marsilea quadrifolia
| Схематическое изображение ЭМ | Характеристика ЭМ | Схематическое изображение ЭМ | Характеристика ЭМ |
|---|---|---|---|
 |
– удлиненный метамер с листом и придаточными корнями; | 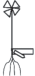 |
– удлиненный метамер с листом, боковым побегом и придаточными корнями; |
 |
– удлиненный метамер с листом, спорокарпиями и придаточными корнями; | 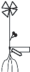 |
– удлиненный метамер с листом, спорокарпием, боковым побегом и придаточными корнями; |
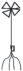 |
– укороченный метамер с листом и придаточными корнями; | 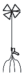 |
– укороченный метамер с листом, боковым побегом и придаточными корнями; |
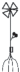 |
– укороченный метамер с листом, спорокарпиями и придаточными корнями; | 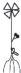 |
– укороченный метамер с листом, спорокарпием, боковым побегом и придаточными корнями |
| Схематическое изображение элемента ЭМ | Описание схематического изображения элемента | Схематическое изображение элемента ЭМ | Описание схематического изображения элемента |
 |
– удлиненный метамер; | 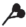 |
– спорокарпии; |
 |
– боковой побег; | ||
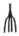 |
– придаточные корни; | 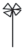 |
– развернутые листочки на черешке |
– узлом и нижележащим удлиненным или укороченным метамером корневища;
– отходящим от узла черешком (со спорокарпиями либо без них), оканчивающимся четырьмя листочками, два из которых (верхние) расположены супротивно друг относительно друга, а два других (нижние) – очередно (Махлин, Сурова, 1978а);
– наличием или отсутствием внепазушных побегов (пояснения см. ниже);
– ветвящимися придаточными корнями.
Заметим, что в разных условиях увлажнения спектр ЭМ, слагающих побеговую систему растения, может изменяться. Например, для растений M. quadrifolia, произрастающих в воде, характерно, в основном, наличие удлиненных метамеров (в составе ЭМ), а в обсыхающих местообитаниях – укороченных.
У сальвинии плавающей особенности строения ЭМ близки к таковым у марсилеи (табл. 3). Различия состоят лишь в отсутствии укороченных метамеров и корней, а также в отхождении от узла корневища трех листьев, два из которых плавающие на поверхности воды (простые и цельнокрайние, яйцевидно-эллиптические, на коротком черешке), третий погружен в воду и рассечен на многочисленные нитевидные сегменты (Махлин, Сурова, 1978б; Барабанщикова, 2012). Между тем “…наблюдения за формированием листьев на апикальной бластозоне побега с помощью сканирующего электронного микроскопа ясно показали, что предполагаемые три листа мутовки у сальвинии происходят из одной общей листовой бластозоны… и на самом деле представляют собой три листочка только одного листа – два противолежащих зеленых плавающих листочка и один погруженный в воду…” (Hagemann et al., 2008, p. 323). От отсутствия или наличия гроздьев из шаровидных сорусов у основания подводного листочка S. natans и спорокарпиев в нижней части черешка у M. quadrifolia, а также наличия или отсутствия бокового побега зависит численное и, соответственно, структурное разнообразие ЭМ. Отметим, что Н.С. Барабанщикова (2012) считает наиболее корректным относить побеги сальвинии к удлиненным, нежели к розеточным.
Таблица 3.
Характеристика и структурные особенности ЭМ Salvinia natans
| Схематическое изображение ЭМ | Характеристика ЭМ | Схематическое изображение ЭМ | Характеристика ЭМ |
|---|---|---|---|
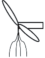 |
– удлиненный метамер с двумя цельными плавающими и одним погруженным нитевидным листочком; |  |
– удлиненный метамер с листочками и боковым побегом; |
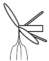 |
– удлиненный метамер с листочками и двумя боковыми побегами; | 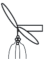 |
– удлиненный метамер с листочками и шаровидными сорусами; |
 |
– удлиненный метамер с листочками, шаровидными сорусами и боковым побегом; |  |
– удлиненный метамер с листочками, шаровидными сорусами и двумя боковыми побегами |
| Схематическое изображение элемента ЭМ | Описание схематического изображения элемента | Схематическое изображение элемента ЭМ | Описание схематического изображения элемента |
 |
– удлиненное междоузлие; |  |
– боковой побег; |
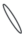 |
– плавающий на поверхности воды листочек; | 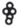 |
– шаровидные сорусы, прикрепленные к погруженному в воду листочку |
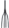 |
– погруженный в воду листочек; |
В отличие от папоротниковидных, у хвощей число и разнообразие ЭМ возрастает (так, у E. fluviatile число ЭМ достигает восьми; табл. 4). Это обусловлено наличием у этих растений геофильной (представленной разветвленным, достаточно долговечным корневищем) и надземной (представленной вегетативным либо спороносным ассимилирующим побегом) структур.
Таблица 4.
Характеристика и структурные особенности ЭМ Equisetum fluviatile
| Схематическое изображение ЭМ | Характеристика ЭМ | Схематическое изображение ЭМ | Характеристика ЭМ | ||
|---|---|---|---|---|---|
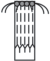 |
– удлиненный метамер с листьями, меристематическими зонами и корнями; |  |
– удлиненный метамер с листьями, меристематическими зонами, реализованным боковым побегом(ами) и корнями; | ||
 |
– удлиненный метамер с листьями, меристематическими зонами; |  |
– удлиненный метамер с листьями, меристематическими зонами и боковой(ыми) стерильной(ыми) веточкой(ами); | ||
 |
– удлиненный метамер с листьями и боковыми веточками; |  |
– удлиненный метамер с листьями, меристематическими зонами и боковой(ыми) веточкой(ами) со стробилом; | ||
 |
– удлиненный метамер с листьями и боковыми веточками со стробилами; |  |
– удлиненный метамер с листьями и главным (терминальным) стробилом | ||
| Схематическое изображение элемента ЭМ | Описание схематического изображения элемента | Схематическое изображение элемента ЭМ | Описание схематического изображения элемента | Схематическое изображение элемента ЭМ | Описание схематического изображения элемента |
 |
– удлиненный метамер; | 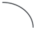 |
– боковой корень; | 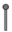 |
– боковая веточка со стробилом; |
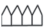 |
– листья; |  |
– боковой побег; |  |
– главный (терминальный) стробил |
 |
– меристематическая зона; |  |
– боковая стерильная веточка; | ||
Так, у E. fluviatile (также как и у E. palustre) ЭМ, как и в предыдущих случаях, представлен удлиненным элементарным метамером, узлом с мутовчато-расположенными листовыми влагалищами и почками (меристематическими зонами). Последние или не трогаются в рост, или развиваются в побеги ветвления следующего порядка или в стерильные либо спороносные веточки, на концах которых развивается ось стробила с мутовками спорангиофоров в виде шестиугольных щитков на ножках (Филин, 1978б).
Как указано выше, наличие или отсутствие тех или иных вариантов ЭМ способствует формированию разных типов универсальных модулей (УМ) – модулей следующего порядка, представляющих собой у цветковых растений (в том числе и у растений вод) новый структурный элемент – одноосный побег (Гатцук, 1974), формирующийся в результате деятельности одной меристемы. У наземных трав сезонного климата, по мнению Н.П. Савиных (2008), в качестве УМ выступает монокарпический побег (термин по: Серебряков, 1952). Аналогичная структурная единица характерна и для споровых растений вод. В зависимости от таксономической группы рассматриваемых нами споровых растений, можно выделить не более одного–двух типов УМ.
Например, для I. lacustris и I. echinospora УМ представлен полициклическим полиспорогенным (так как у побега наблюдается спороношение в течение длительного периода жизни) закрепляющимся в грунте моноподиально нарастающим укороченным (розеточным) побегом с листьями, несущими мега- и микроспорангии, либо без них (рис. 1).
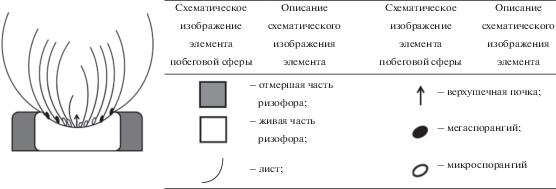
Для гидрофита M. quadrifolia УМ представлен полициклическим полиспорогенным закрепляющимся в грунте и моноподиально нарастающим удлиненным побегом со спорокарпиями в нижней части черешков листьев либо без них (рис. 2); у S. natans – моноциклическим моноспорогенным свободно плавающим моноподиально нарастающим удлиненным побегом с гроздьями из шаровидных сорусов, располагающимися при основании подводного листа, либо без них (рис. 3).
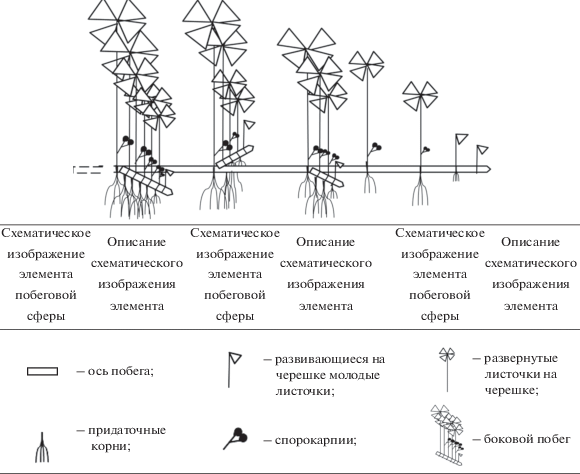
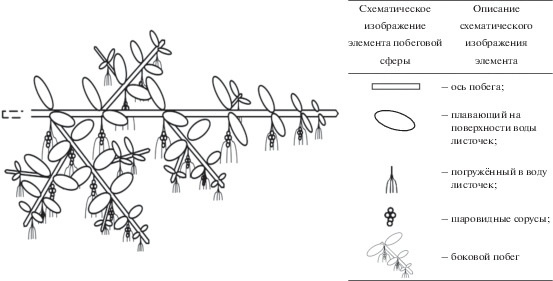
У E. fluviatile, как и у E. palustre, УМ представлен полициклическим полиспорогенным (характер нарастания побега по: Скворцов, 2008) анизотропно моноподиально нарастающим удлиненным побегом, укореняющимся в грунте, с терминальным стробилом либо без него (рис. 4).
Рис. 4.
Структурная организация побеговой сферы E. fluviatile с различными типами основных модулей (а–в). НЗТ – нижняя зона торможения, ЗВ – зона возобновления, СЗТ – средняя зона торможения, ЗО – зона обогащения, ВЗТ – верхняя зона торможения и ЗГС – зона главного (терминального) стробила.
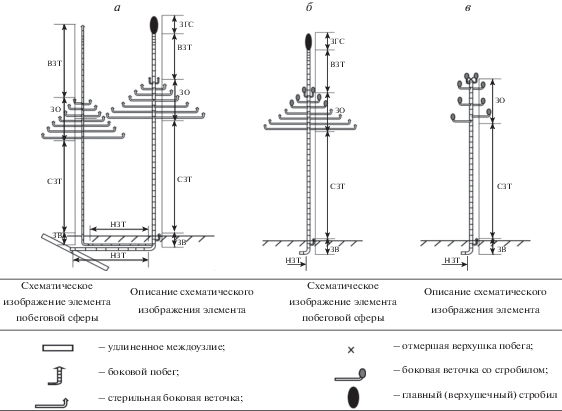
Выделение структурно-функциональных зон на основе УМ (одноосного побега) для водных споровых растений затруднено. Это связано с тем, что весь представленный в литературе набор структурно-функциональных зон (Борисова, Попова, 1990) подробно разработан лишь для цветковых растений. Наибольшее сходство с последними в этом смысле отмечено нами для E. fluviatile и E. palustre (рис. 4).
У этих растений мы выделяем:
– нижнюю зону торможения (НЗТ) – у рассматриваемых видов хвощей находится при основании побега и расположена в почве. Она имеет обособленные меристемы, которые впоследствии могут формировать побеги возобновления, но чаще далее они не развиваются;
– зону возобновления (ЗВ) – также располагается обычно в почве и, в структурном отношении, находится выше НЗТ. Здесь располагаются обособленные меристемы, которые формируют побеги возобновления следующих порядков ветвления;
– среднюю зону торможения (СЗТ) – частично может располагаться в почве, однако основная ее часть находится над уровнем грунта (у хвоща приречного – обычно в толще воды). СЗТ располагает обособленными меристемами, которые не реализуются ни в побеги возобновления, ни в побеги обогащения;
– зону обогащения (ЗО) – располагается выше СЗТ, где происходит развитие боковых ассимилирующих веточек либо со стробилами на терминальном конце, либо без них. Не случайно функции ЗО (фотосинтез и репродукция) у хвощей полностью сходны с таковыми цветковых растений. Отметим, что у отдельных видов, например у наземного вида – хвоща зимующего (Equisetum hyemale L.), ЗО у надземных ассимилирующих побегов как таковая отсутствует;
– верхнюю зону торможения (ВЗТ) – в рамках этой зоны имеющиеся обособленные меристемы также не трогаются в рост;
– зону главного (терминального) стробила (ЗГС). Данный термин введен нами потому, что по своей сути ЗГС полностью идентична зоне главного соцветия цветковых растений. Наличие или отсутствие ЗГС зависит от ритмологических особенностей надземного ассимилирующего побега. Ее отсутствие может быть связано как с физиологическими процессами, так и с механическим нарушением целостности апикальной меристемы побега.
Более сложно в структурно-функциональном аспекте побеговой системы дела обстоят у полушников и рассматриваемых в этой работе папоротников. Разбирая структурную организацию одноосных побегов I. lacustris и I. echinospora, следуя уже сложившейся системе, разработанной для цветковых растений, возникает вопрос о правомерности выделения здесь морфо-функциональных зон. Отсутствие меристематических зон, развивающихся в побеги возобновления на всем протяжении одноосного побега Isoëtes, позволяет нам возвести весь его побег в ранг зоны торможения по аналогии с цветковыми растениями. Наличие вблизи оснований листьев микро- и макроспорофиллов создает, однако, проблемную ситуацию в плане применения к этим растениям концепции структурно-функциональной организации. Аналогичное явление можно наблюдать и у водных папоротников – M. quadrifolia и S. natans. Здесь весь комплекс осевых структур является очень динамичным в своем развитии (за счет интенсивного ветвления) и часто в функциональном и ритмологическом плане может быть целиком представлен в качестве зоны возобновления. При этом наличие шаровидных сорусов (сальвиния) и спорокарпиев (марсилия) создает те же самые проблемы, как и в предыдущем случае.
В качестве высшего модуля, определяющего тип биоморфы растения, выступает основной модуль (ОМ), который формируется “…на основе или в течение жизни целого или части универсального модуля и во многом зависит от положения побегов в пространстве” (Савиных, 2008, c. 62). ОМ споровых и цветковых растений полностью аналогичны друг другу. У полушников ОМ соответствует вегетативно-неподвижному моноцентрическому ортотропно и моноподиально нарастающему короткокорневищному кистекорневому розеточному многолетнику. Жизненная форма S. natans представлена вегетативно-подвижным ацентрическим плавающим моноподиально длиннопобеговым однолетником с полной нормальной неспециализированной морфологической дезинтеграцией (Барабанщикова, 2012), в то время как M. quadrifolia представляет собой полицентрическое вегетативно-подвижное длиннопобеговое многолетнее растение. По жизненной форме E. fluviatile и E. palustre представляют собой вегетативно-подвижные полицентрические многолетние растения с гипогеогенными корневищами и надземными однолетними анизотропными удлиненными побегами. Кроме полушников, всем видам свойственна неспециализированная морфологическая дезинтеграция.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Как показано выше, проблем с выделением УМ и ОМ у водных споровых растений практически нет, а основные вопросы возникают при рассмотрении структуры элементарного модуля. Как известно, у сформировавшегося побега цветкового растения ЭМ представлен следующими структурами в разных сочетаниях (Савиных, 2008):
1) междоузлие (удлиненное или укороченное);
2) лист (низовой, срединной или верховой формации);
3) пазушные элементы – почки разных типов и их производные: соцветие, вегетативные ассимилирующие и вегетативно-генеративные побеги;
4) в ряде случаев придаточные корни.
И если позиции 1 и 4 этого перечня присутствуют у рассматриваемых водных споровых растений (кроме сальвинии плавающей, у которой корни отсутствуют) и практически мало отличаются от подобных структур у цветковых растений, то относительно 2-ой и 3-ей позиций между споровыми и цветковыми растениями существуют серьезные различия. Во-первых, существуют различные точки зрения на наличие (или отсутствие) пазушных элементов у водных споровых растений (Hagemann, 1989, 2008; Hébant-Mauri, 1984 – все цит. по: Vasco et al., 2013). Во-вторых, наличие спороносных органов на листьях требует иных подходов при трактовке листа как одного из элементов ЭМ. Для ответа на эти вопросы авторы данной статьи стремились обобщить свои представления и позиции ряда отечественных и зарубежных исследователей.
В одной из своих основополагающих работ “Геммаксиллярные растения и система соподчиненных единиц их побегового тела” Л.Е. Гатцук (1974) отмечала, что пазушные почки (gemmae axillares) – важнейший структурно-биологический признак, объединяющий жизненные формы покрытосеменных и хвойных. Именно поэтому она дала им название “геммаксиллярные растения” (с. 101). Аксиллярное (пазушное) ветвление Тролль (Troll, 1964) считал частным случаем бокового, а результат последнего – побеговые системы – называл холокладиями. Из ныне живущих организмов холокладиями, кроме геммаксиллярных растений, обладают лиственные мхи, хвощи, саговники, частично папоротники, и для всех них (кроме саговников) характерно экзогенное акропетальное развитие почек (Гатцук, 1974). У лептоспорангиевых же папоротников (к которым относятся изучаемые нами сальвиния и марсилия) боковые почки формируются из обособленных меристем или из подобных участков со скрытыми меристематическими свойствами, расположенных в определенной позиции по отношению к листьям (Wardlaw, 1943, 1945 – цит. по: Гатцук, 1974). Так, говоря о сальвинии плавающей, Н.С. Барабанщикова (2012, с. 155) отмечает, что “боковые побеги располагаются по одному в узле, попеременно то с правой, то с левой стороны от оси главного побега, отходя от стебля между одним из пары надводных листьев и подводным листом”. Об этом ранее писали М.Д. Махлин и Т.Д. Сурова (1978б), отмечая, что почки у сальвинии, из которых образуются латеральные корневища, формируются в узлах между листьями. Судя по рисунку из работы польских исследователей Галка и Змея (Gałka, Szmeja, 2013), боковые побеги S. natans в редких случаях могут располагаться и по два в узле (по одному с каждой стороны). По-видимому, именно за счет этих боковых побегов осуществляется один из типов вегетативного размножения и формирования рамет у сальвинии плавающей (Szmeja, Gałka, 2013). К сожалению, эти исследователи ничего не пишут о точном местоположении боковых побегов и не рассматривают особенности их формирования.
Другой позиции придерживаются немецкие и аргентинские исследователи (Hagemann et al., 2008), которые полагают, что ветвление побегов у сальвинии и марсилии происходит в пазушной позиции. Подобной точки зрения придерживаются и индийские ученые, отмечающие пазушное расположение почек на корневище сальвинии (Zutshi, Vass, 1971). В экспериментальных условиях румынские ученые показали (Brezeanu, Banciu, 2009), что только узловые части корневища Marsilea quadrifolia с предварительно сформированными пазушными меристемами способны к регенерации. Однако Хагеманн (Hagemann, 1989 – цит. по: Vasco et al., 2013) ранее отмечал, что почти у всех папоротников встречается внепазушное ветвление (пазушное наблюдается крайне редко). При этом почки, формирующие побеги, могут находиться в разных позициях относительно точки места прикрепления листа на стебле. По нашему мнению, данный вопрос требует дальнейшего подробного изучения.
У другого изучаемого нами гетероспорового папоротника – M. quadrifolia, длинное и тонкое корневище, ползущее по поверхности почвы подобно столону либо заглубленное в землю, дихотомически ветвится (Samiksha, 2018). Его боковые побеги формируются при основании черешка листа во внепазушной позиции, располагаясь в латеральном или наклонном положении.
Характер ветвления хвощей представляет собой одну из классических проблем морфологии (Stützel, Jaedicke, 2000). С одной стороны, предполагается, что их боковые побеги не формируются в пазухах листьев, а чередуются с редуцированными листьями (Серебряков, 1952; Филин, 1978б; Scagel et al., 1984 – цит. по: Husby, 2013; Stützel, Jaedicke, 2000); с другой стороны, с начала этого столетия ветвление считают экзогенным (Гатцук, 1974; Stützel, Jaedicke, 2000). Хотя тщательное исследование Хофмайстера (Hofmeister, 1851 – цит. по: Stützel, Jaedicke, 2000) показало эндогенный характер ветвления у хвощей, которое позднее было подтверждено и другими зарубежными работами (Barratt, 1920 – цит. по: Stützel, Jaedicke, 2000; Stützel, Jaedicke, 2000). Вместе с тем В.Р. Филин (1978б, с. 139) полагает, что “…ветви хвоща можно считать пазушными, если допустить, что в процессе эволюции большая часть листовой пластинки, в пазухе которой закладывается ветвь, приросла на протяжении целого междоузлия к стеблю”. Таким образом, до сих пор нет единства мнений в этом вопросе. По нашим наблюдениям, у E. fluviatile и E. palustre боковые побеги развиваются в основании междоузлий чуть ниже листьев, чередуясь с последними.
Таким образом, по точному выражению И.Г. Серебрякова (1952, с. 244) “…среди всех папоротникообразных с боковым ветвлением, природа как бы искала путей наилучших отношений между листьями и боковыми почками”. Причем у изучаемых нами споровых растений наблюдалось направление движения от полного отсутствия аксиллярных почек (полушники) через внепазушное положение почек (марсилия и сальвиния) к положению, когда почки находятся в непосредственной близости к листьям (хвощи).
С учетом всего вышеизложенного, рассматривая структуру ЭМ, позицию 3 в характеристике модуля предлагаем изложить следующим образом: внепазушные элементы (споровые растения, не обладающие аксиллярным ветвлением) – боковые побеги, формирующиеся из обособленных меристем или из подобных участков со скрытыми меристематическими свойствами, расположенные в определенной позиции по отношению к листьям, заканчивающиеся спорообразующими элементами (хвощи), либо без них (лептоспорангиатные папоротники, полушники).
Вместе с тем позиция 2 в характеристике модуля должна быть изложена в следующей редакции: лист с наличием или отсутствием спорообразующих элементов (для споровых растений, не обладающих аксиллярным ветвлением, за исключением хвощей).
Заметим, что почти у всех сохранившихся папоротников листья – доминирующие органы растения (Vasco et al., 2013). Подобное, по нашему мнению, характерно и для полушников. Несмотря на то, что листья хвощей уникальны по форме и функциям, доминирующим органом этих растений является стебель, осуществляющий большую часть фотосинтеза (Vasco et al., 2013).
Ранее мы отмечали, что разнообразие ЭМ приводит к формированию различных типов УМ. В пределах растения разнообразие УМ у цветковых растений нередко выше, чем у споровых. Так, например, у Sparganium emersum Rehm., S. natans L. и S. gramineum Georgi в течение одного вегетационного сезона формируется четыре УМ (Беляков, Лапиров, 2015). Значительное число последних демонстрирует высокие адаптационные способности цветковых растений при формировании побеговых систем. Низкий адаптационный потенциал короткокорневищных (розеточных) споровых, например полушников, обусловлен слабой способностью к вегетативному (чрезвычайно редко, по нашим наблюдениям) и генеративному размножению. Кроме того, специфические особенности местообитания (узкая экологическая амплитуда) делают этот вид еще более уязвимым. Однако это характерно не для всех водных споровых. Например, E. fluviatile, имеющий в составе своей побеговой системы два УМ, прекрасно приспособился к условиям местообитания в различных типах водоемов. По-видимому, это связано с наличием у него длинных и мощных корневищ, позволяющих этому виду не только завоевывать новые участки для жизни, но и удерживать ранее завоеванные площади. Последнее характерно для большинства длиннокорневищных цветковых растений.
ЗАКЛЮЧЕНИЕ
Хорошо известно, что в последнее время побег рассматривают как сложную структурно-функциональную (интегрированную и саморегулирующуюся) систему, обеспечивающую растению выполнение основных жизненно важных процессов ассимиляции и репродукции. Генетически запрограммированная морфология и архитектура побега, отличающаяся большой вариабельностью признаков и не меньшей пластичностью структурных элементов, определяет габитус растения и в большинстве случаев его жизненную форму (Берко, Козий, 1987). Оба этих тезиса применимы как для цветковых, так и для споровых растений. Между тем хорошо разработанная система структурно-функциональной зональности побегового тела цветковых растений в большинстве случаев слабо применима для отдельных видов споровых растений (например, полушников, сальвинии и марсилеи). Ключевое различие между споровыми и семенными растениями, касающееся их модульной организации, связано с полным отсутствием аксиллярных почек или расположением их во внепазушной позиции. Тем самым нарушается сама суть системы зонирования побеговой системы водных и околоводных споровых растений, что требует разработки иных подходов к этой проблеме либо совершенствования прежней системы. В последнем случае важным критерием, по нашему мнению, должно стать наличие или отсутствие макро- и микроспорангиев при основании листа. Между тем наиболее простым выходом из данной ситуации является полный отказ от системы зонирования побега некоторых споровых растений и рассмотрение их структуры только с точки зрения модульной организации.
Авторы признательны Н.М. Державиной и Н.П. Савиных за обсуждение работы и ценные замечания. Работа выполнена в рамках госбюджетной темы № АААА-А18-118012690099-2 “Растительный покров водоемов и водотоков России: структура и динамика” (руководитель канд. биол. наук, доцент А.Г. Лапиров) и при частичной финансовой поддержке гранта РФФИ № 18-34-00257 мол_а.
Список литературы
Антонова И.С., Лагунова Н.Г., 1999. О модульной организации некоторых групп высших растений // Журн. общ. биологии. Т. 60. № 1. С. 49–59.
Барабанщикова Н.С., 2012. Жизненная форма сальвинии плавающей (Salvinia natans (L.) All., Salviniaceae) // Актуальные проблемы современной биоморфологии. Киров: Радуга-ПРЕСС. С. 154–156.
Беляков Е.А., 2016. Биология некоторых представителей рода Sparganium L. (сем. Typhaceae). Автореф. дис. … канд. биол. наук. Сыктывкар: ИБ Коми НЦ УрО РАН. 21 с.
Беляков Е.А., Лапиров А.Г., 2015. Модульная и структурно-функциональная организация видов рода Sparganium L. в различных экологических условиях // Сиб. экол. журн. Т. 22. № 5. С. 785–799.
Берко И.Н., Козий Б.И., 1987. О методике моделирования морфологического строения монокарпического побега травянистых многолетников // V Всесоюз. шк. по теорет. морф. раст. Львов: Львовский гос. ун-т. С. 29–33.
Борисова И.В., Попова Г.А., 1990. Разнообразие функционально-зональной структуры побегов многолетних трав // Ботан. журн. Т. 75. № 10. С. 1420–1426.
Гатцук Л.Е., 1974. Геммаксиллярные растения и система соподчиненных единиц их побегового тела // Бюл. МОИП. Отд. биол. Т. 79. № 1. С. 100–113.
Гуреева И.И., 2001. Равноспоровые папоротники Южной Сибири. Систематика, происхождение, биоморфология, популяционная биология. Томск: Томский гос. ун-т. 158 с.
Державина Н.М., Волобуев С.В., Силаева Ж.Г., 2011. Модульная организация спорофита Polypodium vulgare L. // Ученые записки Орловского гос. ун-та. Сер. естественные, технические и медицинские науки. № 5. С. 207–212.
Лапиров А.Г., Беляков Е.А., Лебедева О.А., 2017. Биоморфология и ритм сезонного развития реликтового вида Lobelia dortmanna в олиготрофных озерах Тверской области // Regul. Mech. Biosyst. V. 8. № 3. P. 349–355.
Лебедева О.А., 2006. Биология шелковника волосистого (Batrachium trichophyllum (Chaix) Bosch.). Автореф. дис. … канд. биол. наук. Сыктывкар: ИБ Коми НЦ УрО РАН. 18 с.
Лелекова Е.В., 2006. Биоморфология водных и прибрежно-водных семенных растений северо-востока Европейской России. Автореф. дис. … канд. биол. наук. Пермь: Пермский гос. ун-т. 19 с.
Махлин М.Д., Сурова Т.Д., 1978а. Семейство Марсилеевые (Marsileaceae) // Жизнь растений. Т. 4. М.: Просвещение. С. 242–247.
Махлин М.Д., Сурова Т.Д., 1978б. Семейство Сальвиниевые (Salviniaceae) // Жизнь растений. Т. 4. М.: Просвещение. С. 248–254.
Мейен С.В., 1987. Основы палеоботаники. М.: Недра. 405 с.
Мусина Л.С., 1976. Побегообразование и становление жизненных форм некоторых розеткообразующих трав // Бюл. МОИП. Отд. биол. Т. 81. № 6. С. 123–132.
Петухова Д.Ю., 2008. Биоморфология столонно-розеточных гидрофитов. Автореф. дис. … канд. биол. наук. Сыктывкар: ИБ Коми НЦ УрО РАН. 19 с.
Савиных Н.П., 2007. Модульная организация растений // Онтогенетический атлас растений. Т. 5. Йошкар-Ола: МарГУ. С. 15–34.
Савиных Н.П., 2008. Применение концепции модульной организации к описанию структуры растения // Современные подходы к описанию структуры растений. Киров: Лобань. С. 47–69.
Савиных Н.П., Мальцева Т.А., 2008. Модуль у растений как структура и категория // Вестн. ТвГУ. Сер. Биология и экология. № 9. С. 227–233.
Серебряков И.Г., 1952. Морфология вегетативных органов высших растений. М.: Сов. Наука. 390 с.
Серебрякова Т.И., Павлова Н.Р., 1986. Побегообразование, ритм развития и вегетативное размножение в секции Potentilla рода Potentilla (Rosaceae) // Ботан. журн. Т. 71. № 2. С. 154–167.
Серебрякова Т.И., Петухова Л.В., 1978. Архитектурная модель и жизненные формы некоторых травянистых розоцветных // Бюл. МОИП. Отд. биол. Т. 83. № 6. С. 51–65.
Скворцов В.Э., 2008. Род Equisetum L. в российской и мировой флоре: биоморфология, изменчивость, таксономия. Автореф. дис. … канд. биол. наук. М.: МГУ. 22 с.
Филин В.Р., 1978а. Род полушник, или шильник (Isoëtes) // Жизнь растений. Т. 4. М.: Просвещение. С. 118–121.
Филин В.Р., 1978б. Род хвощ (Equisetum) // Жизнь растений. Т. 4. М.: Просвещение. С. 134–146.
Храпко О.В., 2010. Структурные особенности папоротников // Бюл. БСИ ДВО РАН. № 5. С. 160–166.
Шабалкина С.В., Савиных Н.П., 2011. Строение побеговых систем некоторых видов рода Rorippa Scop. с позиции модульной организации // Науч. ведомости Белгородского гос. уни-та. Сер. Естественные науки. № 9. Вып. 15/1. С. 16–22.
Шорина Н.И., 2001. Гомологии в модульной организации спорофитов папоротниковидных и семенных растений // Гомологии в ботанике: опыт и рефлексия. Тр. IX шк. по теорет. морф. раст. “Типы сходства и принципы гомологизации в морфологии растений”. СПб.: СПбСУ. С. 129–135.
Шорина Н.И., 2007. Биоморфологический и морфогенетический анализ модульной организации спорофитов и форм ветвления короткокорневищных папоротников // Биоморфологические исследования в современной ботанике. Владивосток: БСИ ДВО РАН. С. 487–491.
Brezeanu A., Banciu C., 2009. Comparative studies regarding ultrastructure Marsilea quadrifolia L. (Pteridophyta) leaf mesophyll cells in vivo and in vitro culture // Rom. J. Biol. Plant Biol. V. 54. № 1. P. 13–24.
Gałka A., Szmeja J., 2013. Phenology of the aquatic fern Salvinia natans (L.) All. in the Vistula Delta in the context of climate warming // Limnologica. V. 43 № 2. P. 100–105.
Hagemann W., Sota E.R., de la, Greissl R.P., 2008. Marsileaceae and Salvinlaceae – A critical comparison // Perspectives in Pteridophytes / Eds Verma S.C., Khullar S.P., Cheema H.K. Dehradun, India: Rishen Singh Mihendra Pal Singh. P. 313–334.
Husby C., 2013. Biology and functional ecology of Equisetum with emphasis on the giant horsetails // Bot. Rev. V. 79. № 2. P. 147–177.
Samiksha S., 2018. Useful Notes on the Life History of Marsilea. www.yourarticlelibrary.com/biology/useful-notes-on-the-life-history-of-marsilea-4960-words-2/7081/
Schneider H., Pryer K.M., 2002. Structure and function of spores in the aquatic heterosporous fern family Marsileaceae // Int. J. Plant Sci. V. 163. № 4. P. 485–505.
Smith A.R., Pryer K.M., Schuettpelz E., Korall P., Schneider H., Wolf P.G., 2006. A classification for extant ferns // Taxon. V. 55. № 3. P. 705–731.
Stützel T., Jaedicke A., 2000. Verzweigung bei Schachtelhalmen // Feddes Repert. V. 111. № 1–2. P. 15–22.
Szmeja J., Gałka A., 2013. Survival and reproduction of the aquatic fern Salvinia natans (L.) All. during expansion in the Vistula Delta, south Baltic Sea coast // J. Freshw. Ecol. V. 28. № 1. P. 113–123.
Troll W., 1964. Die Infloreszenzen. B. 1. Jena: Fischer Verlag. 615 p.
Vasco A., Moran R.C., Ambrose B.A., 2013. The evolution, morphology, and development of fern leaves // Front. Plant Sci. V. 4. P. 1–16.
Vöge M., 2014. Monitoring the vitality of Isoëtes lacustris by using a non-destructive method // Limnol. Rev. V. 14. № 3. P. 153–158.
Yatsentyuk S.P., Valiejo-Roman K.M., Samigullin T.H., Wilkstrom N., Troitsky A.V., 2001. Evolution of Lycopodiaceae inferred from spacer sequencing of chloroplast rRNA genes // Russ. J. Genet. V. 37. № 9. P. 1068–1073.
Zutshi D.P., Vass K.K., 1971. Ecology and production of Salvinia natans Hoffim in Kashmir // Hydrobiologia. V. 38. № 2. P. 303–320.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии