

Журнал общей биологии, 2020, T. 81, № 6, стр. 458-468
“Изюминка” популяционной биологии малолетних растений или почему размеры растений также важны
М. В. Марков *
Московский педагогический государственный университет
119991 Москва, Малая Пироговская, 1, Россия
* E-mail: markovsmail@gmail.com
Поступила в редакцию 12.06.2020
После доработки 23.07.2020
Принята к публикации 24.08.2020
Аннотация
Малолетние (одно- и двулетние) растения по своим биоморфологическим свойствам представляют категорию жизненных форм, стоящую особняком от растений всех остальных жизненных форм. Поэтому классические подходы отечественной популяционной биологии растений, связанные с анализом спектров онтогенетических состояний, оказываются ограниченно применимыми или неприменимыми вообще, т.е. для них необходима разработка особых подходов. Чрезвычайно высокая пластичность малолетних растений, как и поливариантность онтогенеза, придающие их популяциям поразительную разнородность в аспекте ритмов и темпов развития, жизненного состояния и плодовитости особей, обусловливают обязательность применения длительных наблюдений на постоянных площадках с регистрацией числа и размеров особей по числу слагающих их тела фитомеров (модулей) в их динамике, фенологических фаз. Закономерно несинхронное прорастание семян в популяциях многих видов приводит к поликогортности – последовательному появлению в активной части популяции когорт проростков, каждая из которых развивается в сложившейся к тому моменту более напряженной эколого-ценотической ситуации (усилившейся конкуренции) и потому состоит из особей более мелких размеров. Анализ размерных спектров зачастую сильно положительно асимметричных распределений по классам общей биомассы, предусматривающий корректное применение статистических приемов, способен дать ценную информацию о популяциях малолетних растений, длительности их жизни и детальную характеристику их реальных конкретных жизненных форм. Вопреки опубликованному ранее мнению суммарная плодовитость особей низших размерных классов оказывается заметно выше плодовитости немногочисленных высоко плодовитых наиболее крупных особей. Самые мелкие экземпляры наиболее эффективны в плане репродукции, демонстрируя высокие значения репродуктивного усилия, при том что в популяциях малолетников часто имеет место изометрия.
В известной книге “Размеры животных: почему они так важны” К. Шмидт-Ниельсен (1987) по вполне понятным причинам обращается за примерами почти исключительно к миру животных. Лишь вскользь, анализируя содержание таблицы 1.1 , он упоминает без комментариев калифорнийские секвойи, намного (в 10–15 раз!) превышающие по размерам синего кита – рекордсмена из мира животных. Еще одно упоминание представителей мира растений фигурирует в книге на графической иллюстрации (с. 92), заимствованной из тоже весьма известной работы Геммингсена (Hemmingsen, 1934), где среди облаков точек для видов животных, на основе которых рассчитана приведенная здесь же линия регрессии, обозначены точки для деревьев (бука). Такая ситуация может породить иллюзию, что для растений размеры тела не так важны, если важны вообще, а процедура, обозначенная в оригинальном заглавии книги К. Шмидта-Ниельсена труднопереводимым (по признанию осуществлявшего перевод книги Н.В. Кокшайского) словом “scaling” (шкалирование, масштабирование), к растениям обычно не применяется, что, конечно, далеко не так. Да, как совершенно справедливо отмечает переводчик в предисловии, “сама конструкция тела животного, его важнейших систем и органов в немалой степени определяется размерами, претерпевая в ходе эволюции то сравнительно легкие и закономерные, но иногда очень резкие и как бы внезапные преобразования в зависимости от масштаба” (Шмидт-Ниельсен, 1987, с. 6). Но размеры растений, приобретаемые в процессе свойственного им модульного, потенциально неограниченного роста, тоже очень важны, и притом на обоих “полюсах” – полюсе максимально возможных для особей вида размеров и полюсе минимальных, но уже допускающих переход в репродуктивное состояние, размеров. Миниатюризация и грандизация – два процесса, взаимодействующие в ходе эволюции малолетних растений, поэтому при сохранении шансов у отдельных особей вырастать до присущих виду максимальных размеров большая часть популяции представлена мелкими особями, и все они, являясь дискретными генетами, выгодно отличаются в методическом плане от многолетних растений дискретностью, наличием четких границ между особями в составе популяции. Проблемы с выделением счетных единиц, которая остро ощущается при работе с многолетними растениями, зачастую обладающими способностью к вегетативному размножению и формированию рамет, здесь не возникает, и к тому же имеется возможность использовать весовые показатели, определять массу как надземных органов, так и корневой системы, скелетная часть которой может быть с необходимой полнотой извлечена и взвешена.
МЕСТО МАЛОЛЕТНИХ РАСТЕНИЙ В РАЗМЕРНЫХ СПЕКТРАХ ВСЕХ ЦВЕТКОВЫХ РАСТЕНИЙ
Наверное, не будет ошибкой сказать, что изучение пластичности особей в популяциях растений, и в первую очередь малолетних, у которых это свойство выражено в очень яркой форме, представляет некую “изюминку” популяционной биологии. При этом амплитуда изменчивости размеров у малолетних растений даже в границах одной популяции впечатляет: размеры самой крупной генеративной особи у некоторых видов могут превышать размеры самой мелкой в 50 тыс. раз (Harper, 1977).
Отсюда понятно, что проблему логично рассматривать и на межвидовом (как это делал К. Шмидт-Ниельсен), и на внутривидовом и популяционном уровнях. Именно по этой причине в настоящей статье предпринята попытка рассмотреть наиболее важные и интересные вопросы, связанные с размерами тела (включая в них и корневую систему) у малолетних цветковых растений в активном и покоящемся (в форме жизнеспособных семян) состоянии, их изменчивостью и зависимостью от размеров особей их репродуктивных характеристик. В число рассматриваемых вопросов, принимая во внимание сильную изменчивость общих размеров особей у растений, включены и аллометрические показатели.
Способность к потенциально неограниченному росту реализуется далеко не всегда и не всеми особями. При этом в ходе эволюции цветковых растений, если принимать за иллюстрацию ее главной линии редукционный ряд жизненных форм, она становится даже более лимитированной. Закономерностям изменчивости размеров тела живых существ на межвидовом уровне посвящена очень интересная, но незаслуженно забытая работа Л.Л. Численко (1981). Взяв размеры тела (у растений высоту или длину главного побега) как очень удобный, легко измеримый и вместе с тем биологически важный признак, тесно связанный со многими (морфологическими, физиологическими, экологическими) свойствами организмов, известный из справочной литературы для большинства видов животных и растений, Л.Л. Численко провел их сопоставление. Были сравнены и кривые распределения (вершины которых обозначают положение оптимумов) по классам размеров числа видов, и средневидовые характеристики размеров (у растений – максимальной высоты) в таксонах. Среди других в монографии представлено логарифмированное распределение значений максимальной длины главного побега (в мм) для 15394 видов цветковых растений флоры СССР разных жизненных форм, которое, как и для других обширных таксонов, имело вид симметричной логнормальной кривой. На рис. 1 приведена эта кривая вместе с кривыми распределения 314 видов малолетних растений (Маевский, 2014) по классам значений максимальной и минимальной высоты, фигурирующих в определителе.
Рис. 1.
Кривые распределения по классам максимальной высоты для малолетников (Маевский, 2014) – 1; всех цветковых флоры СССР (данные Л.Л. Численко (1981), взятые им из издания “Флора СССР”) – 2; и классам минимальной высоты малолетников (Маевский, 2014) – 3.

Наряду с логнормальностью приведенных кривых в соответствии с законом логнормального распределения Геммингсена (Hemmingsen, 1934) налицо удивительное сходство полигона, полученного Л.Л. Численко, с полигоном распределения видов малолетников по классам максимальной высоты. Даже принимая во внимание логарифмическую трансформацию данных, нельзя не отметить это сходство и близость средних величин (2.65–2.71). Для подобающей оценки значимости этих средних процитируем дословно Л.Л. Численко (1981, с. 191): “Средневидовой размер тела таксонов надвидового ранга не есть пустая абстракция. Вместе с тем он и не является персонифицированной характеристикой некоего “среднего вида”, не существующего в природе. Средняя есть абстрактная и однозначная характеристика совокупности, и имеет смысл постольку, поскольку имеет смысл реальное существование самой совокупности как целого”. Население определенного биотопа (или таксона, представляющего в данном биотопе определенный диапазон размеров) имеет характер распределения, весьма близкий к модели, предложенной на основании изучения размеров тела как меры индивидуального ареала (Hutchinson, McArthur, 1959). По высказанному в монографии мнению Л.Л. Численко, представление об определенных упорядоченных отношениях размеров различных таксонов должно быть учтено так же, как и конструктивно-морфологические ограничения, налагаемые на размеры организмов при их экстремальных значениях. Под упорядоченностью отношения размеров в данном случае понимается расположение кривых или средних на определенных расстояниях друг от друга на логарифмической шкале размеров. Л.Л. Численко показал, что расстояние между средними очень близко к константе 0.50 логарифмических единиц. Эта константа не связана обязательно и исключительно с размерами тела. По словам Л.Л. Численко, ее можно выявить и при анализе формы тела (мы добавим, жизненной формы), плодовитости и числа таксонов, т.е. ряд других свойств обнаружит при “биотаксологическом” анализе ту же константу. Л.Л. Численко считал общий анализ относящихся к этой проблеме данных делом будущего, но отметил: что касается числового значения константы, то не исключена возможность, что оно связано с числом π, поскольку величина 0.50 логарифмических единиц равна логарифму числа π с точностью до второго знака. Для нас же будет весьма интересным констатировать, что именно такое значение (0.50) разделяет средние значения максимальной (2.72) и минимальной (2.16) высот для малолетников (рис. 1). Выше названная упорядоченность размерных соотношений фигурирует здесь в явной форме, несмотря на вполне понятную приблизительность данных, особенно о минимальной высоте (длине главного побега). Интересно также отметить, что размерный спектр, вобравший в себя данные о множестве видов, имеет положительную асимметрию и потому логнормален, как и размерный спектр, построенный для множества особей в составе популяции (см. ниже).
СВОЕОБРАЗИЕ КАТЕГОРИИ ЖИЗНЕННЫХ ФОРМ “МАЛОЛЕТНИЕ РАСТЕНИЯ” КАК ОБЪЕКТА ПОПУЛЯЦИОННОЙ БИОЛОГИИ РАСТЕНИЙ В ЕЕ КЛАССИЧЕСКОЙ ОТЕЧЕСТВЕННОЙ ВЕРСИИ
Отечественная популяционная биология растений, зародившаяся в недрах фитоценологии, к сожалению, не может похвастаться обилием методов, подходов и приемов исследования. Задавший тон и ставший по признанию мирового научного сообщества приоритетным (Urbanska, 1992) классическим подход к анализу возрастного состава популяций Т.А. Работнова (1950а, б) предусматривает продолжительные наблюдения за растениями (маркированными или закартированными) на постоянных пробных площадках (Markov, 1985) с регистрацией пополнения и убыли. Введением понятия “возрастность” (биологический возраст) были в определенной степени преодолены объективные сложности с выявлением абсолютного (календарного) возраста у многолетних травянистых растений, и дан старт выделению индикаторных морфологических признаков онтогенетических состояний, используемых затем для построения и анализа возрастного спектра популяции или спектра онтогенетических состояний. Несмотря на безусловную ценность и содержательность получаемых классическим методом результатов, а также несомненный технический прогресс (например, появление цифровой фотографии), позволяющий существенно снизить трудоемкость продолжительных наблюдений на постоянных площадях, число работ этого плана до обидного мало. Волна интереса к углубленному изучению популяций растений, возникшая благодаря подхваченному А.А. Урановым и ботаниками его школы стимулу “работновского” задела, способствовала развитию теории, уточнению терминологии и проведению ряда изящных исследований по классической методике (Заугольнова, 1985; Vorontzova, Zaugolnova, 1985; Zhukova, Ermakova, 1985; Shorina, Smirnova, 1985). Однако позднее продолжительных наблюдений из-за их сложности стало проводиться все меньше, а исследователи стали уделять больше внимания биоморфологической характеристике онтогенетических (возрастных) состояний. Появились совершенно неоправданные с точки зрения русского языка обороты: “дискретное описание онтогенеза” (ведь не описание дискретно, а онтогенез, который мы как плавно текущий непрерывный мультфильм при его создании для удобства разбиваем на “кадры”, т.е. представляем дискретным), “онтогенетическая структура популяций” вместо длинноватого, но более корректного выражения “спектр онтогенетических состояний особей в популяции” и т.п.
В продолжение темы классической статьи А.А. Уранова “Жизненное состояние вида в растительном сообществе” (1960) появлялись разные версии метода учета жизненного состояния. В том числе было введено понятие виталитета, и была предложена методика выявления так называемой виталитетной структуры популяций (Злобин, 1989а, б). Эта методика, привлекшая к себе внимание новизной терминологии, фигурировала в монографии Ю.А. Злобина (2009), рецензию на которую опубликовали в “Журнале общей биологии” Б.М. Миркин и Л.Г. Наумова (2010). Большой вклад в развитие популяционной биологии растений внес А.А. Уранов, выступивший наряду с И.Г. Серебряковым и зоологами Ф.И. Правдиным и С.П. Наумовым в качестве организатора единственной в бывшем СССР научной лаборатории по изучению популяций и возглавлявший ботанический отдел в этой лаборатории. А.А. Уранову вместе с его учениками (О.В. Смирновой, Л.Б. Заугольновой, Л.А. Жуковой, Ю.А. Злобиным и др.) удалось существенно уточнить терминологию, продвинуться в оценках жизненного состояния особей в популяции, выявлении пространственной неоднородности популяции и асинхронности развития ее популяционных локусов. Были разработаны и представления о динамике возрастных спектров популяций и подвергнута дальнейшей разработке классификация типов популяций по характеру их возрастных спектров. Не случайно представители школы И.Г. Серебрякова и А.А. Уранова продолжили исследование популяций с привлечением в качестве объектов видов самых разных жизненных форм, но (по примеру Т.А. Работнова), как правило, многолетних растений. Такую инерцию в выборе объектов можно в какой-то степени оправдать – ведь анализ спектра возрастных (онтогенетических) состояний дает по-настоящему хорошие и полезные для фитоценологии результаты только при работе с популяциями многолетних растений. Да и выделение самих онтогенетических состояний, достаточно долго (не менее одного сезона вегетации) сохраняющих свои индикаторные признаки, целесообразно только для многолетних (моно- и поликарпических) растений. Быстрое же развитие однолетних растений, приводящее к одновременному присутствию всех индикаторных признаков, которые диагностируют скоротечные возрастные изменения у одной особи (например, зацветание при наличии еще живых семядолей; рис. 2), не дает возможности получать полезную фитоценологическую информацию. Попытку некоторых отечественных ботаников (Абрамова, Нурмиева, 2011; Галкина, 2013) применять традиционную схему исследования возрастных (= онтогенетических) состояний и их спектра к популяциям малолетних растений (и даже незимующих однолетников) приходится квалифицировать не иначе как методическую инерционность или инерционность мышления по причине слабой информативности и полезности получаемых данных.

ПОРАЗИТЕЛЬНАЯ ИЗМЕНЧИВОСТЬ ОСОБЕЙ В ПОПУЛЯЦИЯХ МАЛОЛЕТНИХ РАСТЕНИЙ И ВАРИАНТЫ ЕЕ ИНТЕРПРЕТАЦИИ В ЛИТЕРАТУРЕ
Материал, собранный автором настоящей статьи в популяциях малолетников центра Русской равнины, позволил оценить меж- и внутрипопуляционную изменчивость и биоморфологическое разнообразие произрастающих здесь малолетних растений. Нами был разработан алгоритм популяционно-ботанического анализа малолетних растений (архитектура–жизненная форма–эколого-ценотическая стратегия), а также составлен дихотомический ключ для определения жизненной формы малолетнего растения (Марков, 1989).
Самое первое из нескольких допущений, фигурирующих в теории эволюции путем естественного отбора Дарвина, состоит в том, что особи какого-либо вида, образующие популяцию, не тождественны, но отличаются друг от друга, пусть иногда незначительно, по размерам, по скорости роста и развития, по реакциям на температуру и действие прочих экологических факторов (Бигон и др., 1989). Таким образом, внутривидовая и внутрипопуляционная изменчивость особей по размерам, или размерная дифференциация особей, поставлена на первое место в обосновании эволюционной теории.
Говоря о размерах растений разных видов и принимая во внимание метамерный (модульный) характер их роста, подразумевают некий средний размер особи в популяциях. Приблизительность такой информации очевидна, хотя она и дает ориентир при определении по определительным ключам, в которых обыкновенно фигурируют интервалы высот. Нельзя не отметить, что в отношении разных жизненных форм данные о размерах особей будут в разной степени информативны и точны из-за различий в уровне пластичности, свойственной разным жизненным формам. Однолетники, например, настолько пластичны, что средние значения размеров заведомо будут в высшей степени условными – слишком тесная зависимость от условий произрастания и способность переходить к репродукции при минимальных размерах и минимальном числе сформированных фитомеров в момент зацветания (Марков, 2010, 2016).
В настоящее время наметились две тенденции в изучении изменчивости как неотъемлемого свойства организмов и популяций. С одной стороны, появились интересные работы, где анализируются и уточняются новые представления о так называемой модификационной (или ненаследственной) изменчивости в связи с развитием созвучных эпигенетических представлений (Тиходеев, 2013). Факт поразительной внутривидовой и внутрипопуляционной изменчивости принято рассматривать как пример модификационной изменчивости (Васильченко, 1970). С другой стороны, в литературе стали применяться термины, которые не только не способствовали углублению понимания природы изменчивости, но оказались даже вредны, а их авторы, идя на поводу у искушения внести в науку что-то новое, подменяли уже устоявшиеся и зарекомендовавшие себя понятия и термины другими, как будто они сами по себе что-то объясняли. Таким термином, например, стал термин “поливариантность”, который был первоначально предложен Д.А. Сабининым как многовариантность онтогенеза и появился в публикации 1963 г. В этой своей первоначальной и приоритетной форме, предусматривающей протекание онтогенеза особи по одному из нескольких потенциально возможных вариантов (удачные иллюстрации у Д.А. Сабинина представлены в книге), понятие оказалось явно полезным. Данный термин полезен, в частности, для уяснения того, что у любого вида (в любой популяции) растения с его генофондом (генотипом, стратегией) всегда имеется некий набор разных тактических вариантов, путей онтогенеза, из которых особью реализуется в конкретной ситуации какой-то один. Однако в дальнейшем стали делать неоправданно сильный акцент на этом явлении, распространив на все виды внутрипопуляционной разнородности и, подменяя приоритетный классический термин “изменчивость”, создавать иллюзию чего-то нового, теоретически значимого. От того, что мы начнем именовать изменчивость морфологических признаков, включая размеры особей популяции, морфологической поливариантностью и смаковать этот термин, ничего нового и важного не родится. Закономерности возникновения изменчивости размеров под влиянием среды обитания и влияния одних особей на другие нам понятнее не станут. Поэтому, когда им попытались заменять все виды изменчивости и стали говорить о морфологической поливариантности (а не изменчивости), ничего нового, кроме путаницы, не появилось (Жукова, Глотов, 2001). Еще хуже было тогда, когда поливариантностью (Поливариантность развития…, 2006), объявив ее механизмом адаптации растений, пытались объяснять какие-то факты и явления, хотя поливариантность и механизмы ее возникновения сами нуждались в объяснении.
Точно так же, несмотря на имевшийся и употреблявшийся в нашей отечественной и зарубежной литературе термин “пластичность” онтогенеза (pasticity – Harper, 1961; Plastizität – Urbanska, 1992), подразумевавший способность особей, в первую очередь малолетних растений, переходить к репродукции при самых различных размерах (отличающихся порой в 50 тыс. раз), включая ничтожно малые, его стали использовать в несколько ином контексте и притом в дополнение к изменчивости (Злобин, 2009), как будто пластичность не есть один из очень важных вариантов изменчивости. Приоритетное понимание пластичности как реакции на загущение популяции или на уменьшение приходящихся на долю каждой особи ресурсов и отечественное понятие об аккомодации роста (Завадский, 1957) были без каких-либо обоснований и пояснений заменены индексом пластичности – показателем отношения амплитуды изменчивости признака к так называемому коэффициенту свободного развития к наибольшему из размеров, встреченному в различных вариантах природных условий (Злобин, 1989б).
Впечатляющая пластичность в приоритетном значении этого термина зиждется на метамерности (модульности) – формировании повторяющихся однотипных структурных элементов в теле растения.
Как в животном, так и в растительном мире проблема частичной компенсации опережающего увеличения объема при росте и увеличении размеров тела по сравнению с поверхностью решается, в принципе, сходным путем – благодаря усложнению формы органов и тела в целом за счет расчленения на мелкие взаимосвязанные друг с другом субъединицы. Более или менее однотипные и в той или иной степени автономные (особенно у растений) субъединицы, которые, дифференцируясь, накапливаются в ходе роста, и из которых складывается тело, называют метамерами (фитомерами) и/или модулями. Общая генетически закрепленная программа роста носит название “архитектурная модель”. Благодаря метамерности, многолетнее или однолетнее тело растения имеет ритмическое строение и, по существу, представляет пластический слепок онтогенеза особи. Структурная ритмичность – отражение временной ритмичности. Наиболее полным этот слепок оказывается в случае однолетних трав (Юрцев, 1976).
Не случайно Д.А. Сабинин (1963) определял рост растений как процесс новообразования элементов структуры, хотя он и работал в основном с показателем, определяемым весовым методом и уравнением RGR = (ln Wη – ln W0)/(tη – t0), где RGR – относительная скорость роста, W0 – начальная биомасса в момент времени t0, Wη – конечная биомасса в момент времени tη. С учетом метамерности строения растений и ритмичного формирования элементарных метамеров (фитомеров) апексом побега в ходе роста правомерно использовать для оценки интенсивности ростовых процессов особи темпы изменения числа ее метамеров (фитомеров и модулей). Это будет своеобразным “взвешиванием” растений в единицах числа элементарных или иных метамеров и сочетанием структурного и количественного подходов к изучению роста. При работе, например, с редкими охраняемыми растениями такой “неповреждающий” (индеструктивный) способ определения темпов роста обладает рядом серьезных преимуществ перед традиционным весовым методом. Действительно, прежде чем рассматривать накопление собственно биомассы, можно, отталкиваясь от относительной однотипности (и равновеликости) метамеров, слагающих тело растения, регистрировать увеличение в ходе роста числа метамеров (η) как своеобразных единиц биомассы и тогда работать с формулой: RGR' = = (lnη – lnη0)/(t – t0), где RGR' – относительная скорость роста, определяемая без уничтожения растений; η – число приобретенных за время роста метамеров, а η0 – начальное число метамеров. Речь идет, конечно, об элементарных метамерах или фитомерах, размеры которых (и они потому даже бывают указаны в определительных ключах) варьируют слабо по сравнению с варьированием их числа в составе особи растения.
Наряду с глубокими по содержанию публикациями по теме модульности (Гатцук, 1974, 2008), появлялись и такие, где опять-таки были зачем-то применены новые термины для обозначения разных модулей, не способствующие раскрытию сути и значимости явления (Савиных, 2002). Обиднее всего, что эти “новшества” были включены в новое издание классического учебника ботаники Т.И. Серебряковой (Васильев и др., 1979), негативно повлияв на его методическое качество и восприятие студентами (личное сообщение).
Особи в популяции любого вида растений различаются прежде всего по биомассе, которая у малолетних растений выступает как интегрированный показатель размеров особи и ее жизненного состояния. Общая биомасса (или общий вес) складывается из многих других параметров, в той или иной степени коррелятивно связанных с ней, но при характеристике роста и оценке жизненного состояния малолетних растений главенствующее место правомерно оставить именно за биомассой. В этой связи, на наш взгляд, нет противоречия между тем, что Ю.А. Злобин (1989а) призывает использовать весь комплекс морфометрических параметров, не замыкаясь на биомассе, и тем, что зарубежные исследователи, изучающие внутрипопуляционную дифференциацию как таковую, берут за основу именно гетерогенность особей популяции по биомассе. Именуют ее при этом иерархией использования ресурсов (White, Harper, 1970), иерархией доминантности или просто размерной иерархией (Ross, Harper, 1972; Weiner, Solbrig, 1984). Ю.А. Злобин полагает, что биомасса как показатель перегружена неживыми инертными структурами и в большей мере отражает прошлую историю особи, чем ее современный статус, но при этом оговаривается, что в первую очередь это относится к многолетним растениям. Здесь уместно напомнить высказывание Б.А. Юрцева (1976, с. 48) о том, что у однолетних растений побеговое тело представляет собой “наиболее полный пластический слепок онтогенеза”. Биомасса этого тела наилучшим образом информирует нас о жизненном состоянии и на разных промежуточных этапах роста, и в конце как итог онтогенеза.
Поэтому, не отвергая применимость и желательность анализа биомассы при работе с популяциями многолетних растений, рассмотрим примеры возникновения и развития размерной дифференциации в популяциях ряда малолетних травянистых растений. Корректный выбор количественных признаков, отражающих жизненное состояние особей популяции и дающих адекватное представление о структуре популяции, ее жизненном состоянии, масштабах репродукции, очень важен особенно при работе с редкими охраняемыми видами.
Представляется очень интересным и важным вопрос, каков минимальный пороговый размер, минимальная масса особи у того или иного вида, чтобы стала возможной репродукция. Пластичность допускает даже переход на уровень одного цветка и плода, и тогда число плодиков (орешков) в составе многоорешка или число семян пластически “подгоняется” к величине накопленной особью общей биомассы (рис. 2) (Марков, 1992).
Ю.А. Злобин (2009), разработавший методику виталитетного анализа популяций, в своей монографии находит нужным привести дословную цитату, из текста которой становится ясно, как оценивает виталитетный анализ Б.М. Миркин: “виталитетный подход… реализуется проще и часто дает не меньше информации, чем онтогенетический подход. При этом он более прагматичен, так как исследователь избавляется от необходимости субъективных домыслов о возрасте растений по их внешнему облику” (Миркин, Наумова, 1998, с. 137). Обычно для сравнения по размерам используют генеративные особи, поскольку методикой предписано брать в анализ особи одного возрастного состояния, но, как справедливо отмечает сам Ю.А. Злобин, при обследовании популяций редких видов генеративные особи часто могут вообще отсутствовать. В этом случае для установления виталитетного состояния популяций, как это было показано на примере Lilium martagon, можно использовать вегетирующие особи, но “строго одного онтогенетического состояния” (Злобин и др., 2013, с. 255). Отсюда понятно, что без обоснованного объективным анализом (а не субъективными домыслами!) выделения морфологически различимых возрастных (онтогенетических) состояний все равно никак не обойтись, а возрастной спектр, давно доказавший свою значимость в популяционной биологии многолетних растений, нельзя подменить виталитетным.
К тому же точности исследованию популяций малолетних растений никак не добавит методика анализа виталитета с разноской особей всего по трем классам (a, b, c) и рассмотрением урезанного таким образом размерного спектра (Злобин, 1980, 1989б). Статистический прием – построение гистограммы распределения – не случайно предполагает использование определенного числа классов и даже формулы для его вычисления. Главной в анализе внутрипопуляционной дифференциации по размерам (Марков, 2012) была и остается корректно построенная кривая (или гистограмма) изменчивости биомассы. Она является, по сути, детальным спектром жизненных состояний. По ней мы судим об амплитуде вариабельности массы и характере распределения, который необходимо учитывать при любой статистической обработке, и по ней подчас мы даже можем судить о динамических событиях, происходящих в популяциях. Чрезвычайно высокая пластичность малолетних растений, как и поливариантность онтогенеза, придающие их популяциям поразительную разнородность в аспекте ритмов и темпов развития, жизненного состояния и плодовитости особей, обусловливают обязательность применения длительных наблюдений на постоянных площадках с регистрацией числа и размеров особей по числу слагающих их тела фитомеров (модулей) в их динамике, фенологических фаз.
Закономерно несинхронное прорастание семян в популяциях многих видов малолетних растений приводит к поликогортности – последовательному появлению в активной части популяции когорт проростков, каждая из которых развивается в сложившейся к тому моменту более напряженной эколого-ценотической ситуации (усилившейся конкуренции) и потому состоит из особей более мелких размеров, пополняющих первые (низшие) классы, что и отражается на форме кривой распределения (= размерного спектра для логарифмически трансформированных данных о биомассе), приобретающей не только возрастающую положительную асимметрию, но и многовершинность (рис. 3). Таким образом, анализ размерных спектров зачастую сильно положительно асимметричных распределений по классам общей биомассы, предусматривающий корректное применение статистических приемов, способен дать ценную информацию об изменчивости размеров особей в популяциях малолетних растений, длительности их жизни и детальную характеристику их реальных конкретных жизненных форм. При этом корректность применения шкалы для оценки изменчивости по величине коэффициента вариации (CV) вызывает очень большие сомнения. Например, у признаков, подчиняющихся положительно асимметричному распределению, а таких признаков в популяциях растений достаточно много, величина CV может зашкаливать и превышать 100% (Богослов и др., 2019). В литературе неоднократно появлялись нарекания на адекватность коэффициента вариации как мерила изменчивости (Сукачев, 1953; Филипченко, 1978; Bendel et al., 1989).
Рис. 3.
Кривые распределения по классам логарифмов общей биомассы для двух выборок, взятых из агропопуляции Capsella bursa-pastoris в посеве озимой ржи 14 и 30 мая 1975 г. Пунктиром намечены границы когорт особей в составе популяции (I – осенняя когорта и II – весенняя когорта). W – общая биомасса, f – частоты в %.
УЧАСТИЕ ОСОБЕЙ РАЗНЫХ РАЗМЕРОВ В РЕПРОДУКЦИИ И ВОЗОБНОВЛЕНИИ ПОПУЛЯЦИЙ МАЛОЛЕТНИХ РАСТЕНИЙ
Размерный спектр популяций малолетних растений не был бы столь важным и информативным в их популяционной биологии, если бы не был связан напрямую с отражением особенностей репродукции, что вытекает из существования высокой положительной корреляции между массой особи и ее плодовитостью (числом формируемых семян). Здесь мы еще раз можем убедиться в чрезвычайной важности пластичности размеров особей малолетних растений и обосновать интерес к “изюминке” их популяционной биологии. Характер распределения по классам размеров (резко положительно асимметричный размерный спектр популяции) был основанием для очень важного вывода об участии в репродукции наиболее крупных (и потому наиболее плодовитых), но немногочисленных особей популяции. Долгое время считалось, что именно эти особи превалируют в формировании нового урожая семян популяции (Koyama, Kira, 1956; Obeid et al., 1967; Ford, 1975; Naylor, 1976; Gottlieb, 1977; Solbrig, 1981; Dolan, 1984; Mithen et al., 1984; Weiner, 1985; Bonan, 1988; Bernston, Weiner, 1991). Правда, еще в публикации 1990 г. автор настоящей статьи (Марков, 1990) усомнился в правильности этого заключения, проанализировав участие особей из разных размерных классов в воспроизводстве популяций (рис. 4). Публикация была в виде скромной монографии на русском языке и потому осталась незамеченной для наших зарубежных коллег. Но относительно недавно видные зарубежные исследователи (Chambers, Aarssen, 2009) обратили внимание на это интереснейшее явление – преимущественную роль мелких (и мельчайших) особей в формировании семенной продукции популяции, и первоначальное воззрение на генетический эффект в популяции пришлось корректировать.
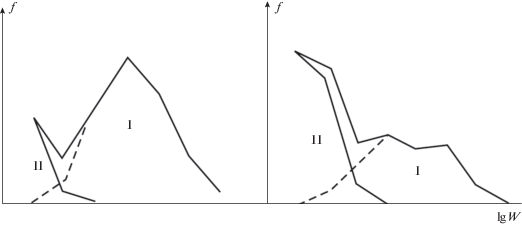
Рис. 4.
Участие особей разного размера в репродукции популяции клоповника густоцветкового (вверху) и рогоглавника (внизу). Кривые распределения по классам общей биомассы (W) на фоне гистограмм распределения суммируемого по тем же классам показателя репродукции Nr: числа односемянных плодиков (Ceratocephala) или числа двусемянных плодов (Lepidium) у особей разных размерных классов.

Выявленная теперь в популяциях многих видов (Capsella bursa-pastoris, Lepidium densiflorum, Polygonum scabrum и др.; Марков, 2012; Moles, 2018) эта закономерность заставляет взглянуть по-иному на выделенный по методике анализа виталитета (Злобин, 1984) “низший” класс “а”, из-за повышенной частоты которого популяцию записывают в разряд регрессивных.
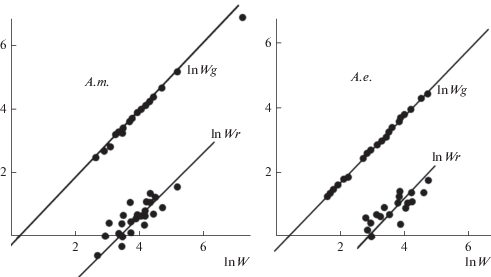
Как раз на примере малолетних растений с характерной для них пластичностью – широчайшим размахом варьирования общих размеров растений в генеративной фазе развития – удобно изучать аллометрию (рис. 5). На рис. 5 можно видеть иллюстрацию аллометрических связей, указывающих на изометрию (близость аллометрического коэффициента к 1.0) биомассы скелетных корней (Wr) и генеративных структур (Wg) в популяции двулетника проломника северного Androsace septentrionalis (lnWg = 1.01lnW – 0.32; lnWr = = 1.0 ln W – 3.2) и зимующего однолетника проломника удлиненного A. elongata (ln Wg = = 1.04 ln W – 0.41; ln Wr = 0.79 ln W – 2.08). Результаты показали также, что в популяциях ряда однолетников (Polygonum scabrum, Ceratocephala falcata) самые мелкие экземпляры наиболее эффективны в плане репродукции, демонстрируя самые высокие значения репродуктивного усилия (Марков, Плещинская, 1987; Марков, 2001).
Список литературы
Абрамова Л.М., Нурмиева С.В., 2011. Онтогенез циклахены дурнишниколистной (Cyclachaena xanthiifolia (Nutt.) Fresen) // Онтогенетический атлас растений. Т. 6. Йошкар-Ола: Марийский гос. ун-т. С. 66–70.
Бигон М., Харпер Дж., Таунсенд К., 1989. Экология: особи, популяции и сообщества. М.: Мир. Т. 1. 667 с.
Богослов А.В., Кашин А.С., Шилова И.В., Крицкая Т.А., Пархоменко А.С., Гребенюк Л.В., 2019. Пластичность и особенности общей и согласованной изменчивости признаков в популяциях Delphinium litvinowii Sambuk // Проблемы ботаники южной Сибири и Монголии. № 18. Барнаул: Алтайский гос. ун-т. С. 86–91.
Васильев А.Е., Воронин Н.С., Еленевский А.Г., Серебрякова Т.И., 1979. Ботаника. Анатомия и морфология. М.: Просвещение. 480 с.
Васильченко И.Т., 1970. О генетической и таксономической значимости модификаций у растений // Бот. журн. Т. 55. № 3. С. 357–363.
Галкина М.А., 2013. Онтогенез череды облиственной (Bidens frondosa) // Онтогенетический атлас растений. Т. 7. Йошкар-Ола: Марийский гос. ун-т. С. 147–150.
Гатцук Л.Е., 1974. Геммаксиллярные растения и система соподчиненных единиц их побегового тела // Бюлл. МОИП. Отд. биол. Т. 79. № 1. С. 100–113.
Гатцук Л.Е., 2008. Унитарные и модульные живые существа: к истории развития концепции // Вестн. Твер. гос. ун-та. Сер. Биология и экология. № 9. С. 29–41.
Жукова Л.А., Глотов Н.В., 2001 Морфологическая поливариантность онтогенеза в природных популяциях растений // Онтогенез. Т. 32. № 6. С. 455–461.
Завадский К.М., 1957. Перенаселение и его роль в эволюции // Бот. журн. Т. 42. С. 426–449.
Заугольнова Л.Б., 1985. Оценка степени динамичности ценопопуляций в пределах одного фитоценоза // Динамика ценопопуляций растений. М: Наука. С. 45–62.
Злобин Ю.А., 1980. О неравноценности особей в ценопопуляциях растений // Бот. журн. Т. 65. № 3. С. 311–322.
Злобин Ю.А., 1984. Ценопопуляционный анализ в фитоценологии. Владивосток: Изд-во АН СССР. 60 с.
Злобин Ю.А., 1989а. Принципы и методы изучения ценотических популяций. Казань: Изд-во Казанского гос. ун-та. 146 с.
Злобин Ю.А., 1989б. Теория и практика оценки виталитетного состава ценопопуляций растений // Бот. журн. Т. 74. № 6. С. 769–781.
Злобин Ю.А., 2009. Популяционная экология растений; современное состояние, точки роста. Сумы: Университетская книга. 263 с.
Злобин Ю.А., Скляр В.Г., Клименко А.А., 2013. Популяции редких видов растений: теоретические основы и методика изучения. Сумы: Университетская книга. 439 с.
Маевский П.Ф., 2014. Флора средней полосы европейской части России. 11-е изд. М.: Т-во науч. изд. КМК. 635 с.
Марков М.В., 1989. Алгоритм популяционно-ботанического анализа малолетних растений: архитектурная модель–жизненная форма–эколого-ценотическая стратегия // Биол. науки. № 11. С. 90–104.
Марков М.В., 1990. Популяционная биология розеточных и полурозеточных малолетних растений. Казань: Изд-во Казанского ун-та. 178 с.
Марков М.В., 1992. Роль многоорешков в обеспечении пластичности репродуктивной сферы у Ceratocephala falсata (L.) Pers и Myosurus minimus L. // Бюлл. МОИП. Отд. биол. Т. 97. № 4. С. 51–67.
Марков М.В., 2001. Особенности метамерного строения малолетних растений и аллометрический анализ репродукции в их популяциях // Бюлл. МОИП. Отд. биол. Т. 106. № 5. С. 83–90.
Марков М.В., 2010 Особенности онтогенеза лужницы Limosella aquatica L.: зацветающие проростки // Мат-лы I (VII) Междунар. конф. по водным макрофитам “Гидроботаника 2010”. Ярославль: “Принт Хаус”. С. 215–217.
Марков М.В., 2012. Популяционная биология растений. М.: Изд-во КМК. 387 с.
Марков М.В., 2016. К биоморфологии, анатомии и популяционной биологии двух представителей клады Ranunculinae // Современные концепции экологии биосистем и их роль в решении проблем сохранения природы и природопользования: Всерос. (с междунар. участием) науч. школа-конф., посвященная 115-летию со дня рождения А.А. Уранова. Пенза. С. 129–132.
Марков М.В., Плещинская Е.Н., 1987. Репродуктивное усилие у растений // Журн. общ. биологии. Т. 48. № 1. С. 77–83.
Миркин Б.М., Наумова Л.Г., 1998. Наука о растительности. Уфа: Гилем. 413 с.
Миркин Б.М., Наумова Л.Г., 2010. Злобин Ю.А. Популяционная экология растений: современное состояние, точки роста. Монография. Сумы: Университетская книга // Журн. общ. биологии. Т. 71. № 3. С. 268–271.
Поливариантность развития организмов, популяций и сообществ, 2006. Йошкар-Ола: Науч. Издание. Марийский гос. ун-т. 326 с.
Работнов Т.А., 1950а. Вопросы изучения состава популяций для целей фитоценологии // Проблемы ботаники. № 1. С. 465–483.
Работнов Т.А., 1950б. Жизненный цикл многолетних травянистых растений в луговых ценозах // Тр. Бот. ин-та АН СССР. Сер. 3. Геоботаника. № 6. С. 7–204.
Сабинин Д.А., 1963. Физиология развития растений. М.: Наука. 196 с.
Савиных Н.П., 2002. Модули у растений // Мат-лы международ. конф. по анатомии и морфологии растений (Санкт-Петербург, 14–18 октября 2002 г.). С. 95–96.
Сукачев В.Н., 1953. О внутривидовых и межвидовых взаимоотношениях среди растений // Бот. журн. Т. 38. № 1. С. 57–96.
Тиходеев О.Н., 2013. Механизмы модификационной изменчивости // Экол. генетика. Т. 11. № 3. С. 81–91.
Уранов А.А., 1960. Жизненное состояние вида в растительном сообществе // Бюлл. МОИП. Отд. биол. Т. 65. № 3. С. 77–92.
Филипченко Ю.А., 1978. Изменчивость и методы ее изучения М.: Наука. 240 с.
Численко Л.Л., 1981. Структура фауны и флоры в связи с размерами организмов. М.: Изд-во МГУ. 208 с.
Шмидт-Ниельсен К., 1987. Размеры животных: почему они так важны? М.: Мир. 259 с.
Юрцев Б.А., 1976. Жизненные формы: один из узловых объектов ботаники // Проблемы экологической морфологии. М.: Наука. С. 9–54.
Bendel R.B., Higgins S.S., Teberg J.E., Pyke D.A., 1989. Comparison of skewness coefficient, coefficient of variation, and Gini coefficient as inequality measures within populations // Oecologia. V. 78. P. 394–400.
Bernston G.M., Weiner J., 1991. Size structure of populations within populations: Leaf number and size in crowded and uncrowded Impatiens pallida individuals // Oecologia. V. 85. P. 327–331.
Bonan G.B., 1988. The size structure of theoretical plant populations: Spatial patterns and neigh borhood effects // Ecology. V. 69. P. 1721–1730.
Chambers J., Aarssen L.W., 2009. Offspring for the next generation: Most are produced by small plants within herbaceous populations // Evol. Ecol. V. 23. P. 737–751.
Dolan R.W., 1984. The effect of seed size and maternal source on individual size in a population of Ludwigia leptocarpa (Onagraceae) // Am. J. Bot. V. 71. P. 1302–1307.
Ford E.D., 1975. Competition and stand structure in some even-aged plant monocultures // J. Ecol. V. 63. P. 311–333.
Gottlieb L.D., 1977. Genotypic similarity of large and small individuals in a natural population of the annual plant Stephanomeria Exigua sp. Coronaria (Compositae) // J. Ecol. V. 65. P. 127–134.
Harper J.L., 1961. Approaches to the study of plant competition // Symp. Soc. Exp. Biol. V. 15. P. 1–39.
Harper J.L., 1977. Population biology of plants. L.: Academic Press. 892 p.
Hemmingsen A.M., 1934. A statistical analysis of the differences in body size in related species // Vidensk. Medd. Naturf. Foren. V. 98. P. 125–160.
Hutchinson G.E., McArthur G.E., 1959. A theoretical ecological model of size distributions among species of animals // Am. Nat. V. 43. № 869. P. 117–125.
Koyama H., Kira T., 1956. Interspecific competition among higher plants. VIII. Frequency distributions of individual plant weight as affected by the interaction between plants // J. Inst. Polytech. Osaka City Univ. Ser. D. V. 7. P. 73–94.
Markov M.V., 1985. Research on permanent quadrats in the USSR // The Population Structure of Vegetation / Handbook of Vegetation Science. Dordrecht: Junk Publishers. P. 111–119.
Mithen R., Harper J.L., Weiner J., 1984. Growth and mortality of individual plants as a function of “available area” // Oecologia. V. 62. P. 57–60.
Moles A.T., 2018. Being John Harper: Using evolutionary ideas to improve understanding of global patterns in plant traits // J. Ecol. V. 106. P. 1–18.
Naylor R.L., 1976. Changes in the structure of plant populations // J. Appl. Ecol. V. 13. P. 513–521.
Obeid M., Machin D., Harper J.L., 1967. Influence of density on plant to plant variation in fiber flax, Linum usitatissimum // Crop. Sci. V. 7. P. 471–473.
Ross M.A., Harper J.L., 1972. Occupation of biological space during Oseedling establishment // J. Ecol. V. 60. № 1. P. 77–88.
Shorina N.I., Smirnova O.V., 1985. The population biology of ephemeroids // The Population Structure of Vegetation / Handbook of Vegetation Science. Dordrecht: Junk Publishers. P. 225–240.
Solbrig O.T., 1981. Studies on the population biology of the genus Viola. 11. The effect of plant size on fitness in Viola sororia // Evolution. V. 35. P. 1080–1094.
Urbanska K.M., 1992. Populationsbiologie der Pflanzen. Stuttgart; Jena: G. Fischer. 373 s.
Vorontzova L.I., Zaugolnova L.B., 1985. Population biology of steppe plants // The Population Structure of Vegetation / Handbook of Vegetation Science. Dordrecht: Junk Publishers. P. 143–178.
Weiner J., 1985. Size hierarchies in experimental populations of annual plants // Ecology. V. 66. P. 743–752.
Weiner J., Solbrig O.T., 1984. The meaning and measurement of size hierarchies in plant populations // Oecologia. V. 61. № 3. P. 334–336.
White J., Harper J.L., 1970. Correlated changes in plant size and number in plant populations // J. Ecol. V. 58. № 2. P. 467–485.
Zhukova L.A., Ermakova I.M., 1985. Structure and dynamics of coenopopuations of some temperate grasses // The Population Structure of Vegetation / Handbook of Vegetation Science. Dordrecht: Junk Publishers. P. 179–205.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии