

Журнал общей биологии, 2020, T. 81, № 6, стр. 444-457
Являются ли сухопутные перемещения амфибийных рыб латеральными миграциями?
Д. Д. Зворыкин *
Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский просп., 33, Россия
* E-mail: d.zworykin@gmail.com
Поступила в редакцию 05.06.2020
После доработки 21.07.2020
Принята к публикации 20.08.2020
Аннотация
Первое сообщение о сухопутных перемещениях рыб-ползунов было опубликовано в научной литературе в конце XVIII в., однако до сих пор про это поведение известно крайне мало. Было высказано несколько гипотез о том, зачем амфибийные рыбы покидают воду, но ни одна из них на сегодняшний день не подтверждена. Короткие передвижения ползунов по суше между водоемами происходят в сезон дождей, когда погодные условия делают такое поведение возможным. На тот же период приходится пик нереста этих рыб. В последние десятилетия некоторые авторы стали априорно связывать между собой два этих явления, однако роль сухопутных перемещений в качестве нерестовых миграций не доказана. Кроме того, передвижения ползунов по суше стали называть латеральными миграциями, что не соответствует определению данного понятия. Очевидно, что сухопутные перемещения обеспечивают высокую пластичность жизненных стратегий ползунов, населяющих среду с изменчивой фрагментарностью. В то же время с большой вероятностью данные передвижения не являются миграциями и не всегда связаны с нерестом. Представляется перспективным применить к изучению жизненных циклов ползунов метапопуляционный подход, который позволил бы прояснить как структурную организацию популяций этих рыб, так и роль различных типов перемещений, обеспечивающих связь и обмен между субпопуляциями.
Лучеперые рыбы (Actinopterygii) представляют собой не только наиболее многочисленный, но и наиболее разнообразный класс современных позвоночных животных (Helfman et al., 2009). Одной из самых примечательных является экологическая группа рыб, часть жизненного цикла которых может проходить за пределами водной среды (Sayer, Davenport, 1991; Graham, 1997). Основную часть группы составляют различные пресноводные тропические виды, многих из которых иногда условно называют ползунами. Это, например, анабас (Anabas testudineus), рисовые угри (Monopterus albus), змееголовы (Channa argus), клариевые сомы (Clarias batrachus) и многие другие. В то же время к ней же относят эвригалинных илистых прыгунов (Periophthalmus variabilis), карпозубообразных (Kryptolebias marmoratus), диадромных речных угрей (Anguilla anguilla), некоторых морских собачек (Alticus arnoldorum).
В последние годы интерес к этим животным стремительно вырос, и они стали объектом целого ряда важных исследований, в том числе связанных с развитием представлений об эволюционном выходе позвоночных на сушу (Bayley et al., 2019; Mai et al., 2019; Du, Standen, 2020). К сожалению, в русскоязычной литературе большинство этих рыб упоминают лишь вскользь и никогда не рассматривают как целостный биологический феномен. В результате для них не существует даже общепринятого русского названия. Данная группа настолько разнообразна по формам своих взаимодействий с воздушной средой, что, как представляется, только само это взаимодействие как таковое и может использоваться для наименования всей группы. В данной работе я буду пользоваться переводом принятого английского термина “amphibious fish” – амфибийные рыбы. Все остальные варианты представляются менее удачными, а прилагательное “амфибийный” зафиксировано в словарях современного русского языка (Тихонов, 2002; Гришина, Ляшевская, 2020).
Характерной особенностью большинства амфибийных рыб является их способность не только к нахождению (иногда довольно продолжительному) на суше, но и к перемещению по поверхности земли, осуществляемому различными способами. Помимо вопросов, связанных с анатомическими и физиологическими адаптациями к такому образу жизни (Liem, 1987; Bressman et al., 2019; Wicaksono et al., 2020), принципиально важным является понимание экологической значимости такого поведения в терминах концепции жизненных стратегий животных. Показательным в этом отношении является название одного из первых обзоров, посвященных данному явлению: “Амфибийные рыбы: зачем они покидают воду?” (Sayer, Davenport, 1991). Иначе говоря, такой необычный для водных животных тип активности ставит перед исследователями целый ряд вопросов, связанных с адаптивной значимостью данного феномена, его связью с питанием, размножением, конкурентными взаимоотношениями, прессом хищников, паразитов и др.
Корректная интерпретация сухопутных передвижений рыб невозможна без понимания того, каким образом это поведение соотносится с организацией их жизненного цикла. В частности, коль скоро для многих рыб характерны миграции различного типа, важно понимать, являются ли сухопутные перемещения амфибийных рыб, подчас имеющие массовый характер, одной из форм миграционной активности. Если говорить о пресноводных тропических рыбах, составляющих, как уже было сказано, основу группы, речь следует вести, прежде всего, о так называемых латеральных миграциях, осуществляемых рыбами между руслом реки и пойменными водоемами. На первый взгляд, предположение о связи сухопутных передвижений рыб с латеральными миграциями выглядит правдоподобным. В публикациях последних лет эта гипотеза даже стала фигурировать в качестве априорно принимаемого факта, однако, как представляется, оснований для этого на сегодняшний день не существует.
Рассмотрение сухопутных перемещений амфибийных рыб в качестве элемента их жизненной стратегии требует также принятие во внимание их связи с циклическими процессами в окружающей среде, так как именно влияние среды во многом регулирует жизненный цикл организма (Wootton, Smith, 2015). В пресноводных биотопах наиболее важным в этом аспекте является годовой климатический цикл, прежде всего потому, что с чередованием периодов дождей и засухи связаны динамические изменения в водоемах тропического региона и зависящие от них колебания репродуктивной активности рыб. Наконец, еще одна важная проблема связана с влиянием на жизненный цикл таких рыб антропогенных факторов. В частности, большие площади пойменных земель многих стран Южной и Юго-Восточной Азии заняты рисовыми полями, обрабатываемыми пестицидами. Если сухопутные перемещения населяющих данный регион амфибийных рыб действительно являются миграциями, хозяйственная деятельность человека может катастрофически влиять на их жизненный цикл, поскольку миграции по определению являются его облигатным элементом (Никольский, 1963; Binder et al., 2011).
Данный обзор представляет собой попытку анализа основных литературных и собственных данных, связанных с сухопутными перемещениями амфибийных рыб в контексте современных представлений о миграционной активности и жизненных стратегиях рыб, обитающих во фрагментированной среде. Основное внимание при этом уделено рыбе-ползуну, или анабасу (Anabas testudineus), как одному из наиболее типичных, известных и широко распространенных представителей данной экологической группы.
ПЕРВЫЕ СООБЩЕНИЯ О ПОЛЗУНАХ
Анабас – не только одна из наиболее широко встречающихся, но и одна из самых знаменитых пресноводных рыб Юго-Восточной и Южной Азии. Начало ее известности было положено более 200 лет назад, в 1797 г., когда в Трудах Линнеевского общества вышла заметка лейтенанта Дальдорфа с сообщением о новом виде рыб, названном им Perca scandens (Daldorff, 1797). Позже оказалось, что приоритет в описании вида принадлежит Блоху, в 1792 г. назвавшему эту рыбу Anthias testudineus (Bloch, 1792). Однако о необычном поведении амфибийных рыб первым рассказал именно Дальдорф, сообщив, что с изумлением застал анабаса за весьма странным занятием – рыба не только ползала по земле, но и карабкалась вверх по трещине ствола пальмы, растущей неподалеку от пруда.
Справедливости ради заметим, что о ползающих по суше рыбах сообщали еще античные авторы, в частности Аристотель и Теофраст (Sharples, 1992). Однако сами они таких рыб не видели и по их рассказам затруднительно понять, о каких именно видах идет речь. В то же время благодаря античным писателям, с чьими трудами все ученые Нового времени были знакомы, сообщение Дальдорфа о ползающей рыбе не вызвало недоверия у научной общественности. А вот карабканье на дерево не давало покоя биологам вплоть до конца XX в. (Hamilton, 1822; Das, 1927; Smith, 1945; Graham, 1997). Во-первых, это был вопрос репутации солидного научного журнала и лично публикатора этого сообщения, знаменитого английского натуралиста, президента Королевского Общества, сэра Джозефа Бэнкса. Во-вторых, и рассказы аборигенов, и местные названия этой рыбы на разных языках свидетельствовали именно о такой особенности анабаса. К настоящему времени обсуждение этого вопроса затихло само собой, однако история изучения анабаса и амфибийных рыб в целом была бы неполной без публикации Дальдорфа.
Как ни удивительно, но и спустя два с лишним века сведения о биологии ползунов и, в частности, об их наземных перемещениях остаются разрозненными, противоречивыми и наполненными большим количеством сомнительной информации и спорных трактовок. Если ученых XVIII–XIX вв. занимало, действительно ли анабас забирается на пальму, чтобы напиться сладкого пальмового сока, то их коллег XX в. больше всего интересовала его анатомия и функциональная морфология. Они исследовали, как устроены лабиринтовый (супрабронхиальный) орган, благодаря которому рыба-ползун может дышать воздухом, жаберные крышки и грудные плавники, помогающие ей передвигаться по суше; они изучали, как она это делает, но очевидный, казалось бы, вопрос о том, зачем ползуны вылезают на сушу, стал обсуждаться лишь сравнительно недавно. Появившиеся в XXI в. многочисленные публикации, посвященные анабасу как перспективному объекту аквакультуры, не только не прояснили ситуацию, но во многих аспектах еще больше запутали ее (Зворыкин, 2020).
КУДА И ЗАЧЕМ ПОЛЗУТ ПОЛЗУНЫ?
Любые сообщения о таком необычном для рыб способе передвижения, а тем более любые его обсуждения должны были сопровождаться если не сведениями, то хотя бы какими-то предположениями, касающимися его назначения. За редким исключением, на протяжении всего XX в. размножение практически никем не рассматривалось, даже гипотетически, в качестве причины, побуждающей амфибийных рыб покидать воду. Дас (Das, 1927), например, писал, что эти рыбы выходят на сушу в поисках земляных червей. Смит (Smith, 1945) считал, что возможны три причины, по которым анабас может покидать воду, при этом ни одна из них, по его мнению, не связана с размножением: (1) поиск лучших условий жизни, (2) поиск корма, (3) бегство от врагов. Некоторые авторы писали про бегство ползунов из пересыхающих водоемов и поиск новых пригодных для жизни биотопов (Sao-Leang, Dom-Saveun, 1955; Graham, 1997; Poulsen, Valbo-Jørgensen, 2000).
По мнению Лиема (Liem, 1987), причиной сухопутных перемещений анабаса может быть повышение плотности популяции и возрастание внутривидовой конкуренции. Примечательно, что тот же автор допускает, что рисовый угорь может перемещаться по суше в поисках подходящего места для размножения, однако на другие виды рыб (в том числе на анабаса) это допущение автор не распространяет. Одной из первых попыток критического осмысления всего накопленного по данной теме материала стала упомянутая выше обзорная статья Сейера и Давенпорта (Sayer, Davenport, 1991). Однако ничего нового про нерест ползунов в этом обзоре не обнаруживается. Выход на сушу в качестве возможной формы нерестовой миграции упоминается только вскользь и только применительно к речным угрям и рисовому угрю.
Что касается версий, связанных с бегством от пересыхания, следует не забывать о том, что до сих пор нет ни достаточно убедительных подтверждений, ни опровержений повторяющимся уже несколько десятилетий сообщениям о том, что анабас и ряд других амфибийных рыб при пересыхании водоема могут зарываться в ил и пережидать там весь период засухи (Inger, Kong, 1962; Thakur, Das, 1986; Rahman, 1989; Graham, 1997; Tay et al., 2006). В наше время сообщения об этом в научной литературе также дополняются многочисленными видеороликами, демонстрирующими, как жители различных стран региона буквально выкапывают анабасов, клариевых сомов, змееголовов и некоторых других рыб живыми со дна пересохших водоемов. Однако хорошо видно, что под сухим слоем поверхности дна остаются влажные области, где, собственно, и обнаруживаются эти рыбы, способные дышать воздухом. Неизвестно, как долго эти слои сохраняют степень влажности, достаточную для выживания рыб, которые, в отличие от, например, протоптеров, не формируют кокон. Нет также подтверждений тому, что анабасы и большинство других ползунов впадают в состояние эстивации, достаточно хорошо изученное у тех же протоптеров (Fishman et al., 1986; Land, Bernier, 1995). Неудивительно, что многие исследователи полагают, что способность большинства рыб, даже способных дышать воздухом, пережидать таким образом засуху сильно преувеличена (Bruton, 1979).
Среди первых авторов, предположивших, что сухопутные перемещения рыбы-ползуна связаны с размножением, были индийцы Мукерджи и Мазумдар (Mookerjee, Mazumdar, 1946). Их три причины выхода анабаса из воды сформулированы так: (1) поиск безопасного места для нереста, (2) перегрев воды в жаркое время года и (3) высыхание водоема основного обитания. Позже другие индийские исследователи заметили, что пойманные на суше анабасы готовы к размножению и могут отнереститься в аквариуме без гормональной стимуляции, к которой обычно приходится прибегать для их размножения (Banerji, Prasad, 1974). Наконец, Давенпорт и Абдул Матин (Davenport, Abdul Matin, 1990) упомянули, что рыбы-ползуны чаще всего покидают воду в сезон дождей, совпадающий с сезоном их размножения. Таким образом, версия о связи сухопутных передвижений анабаса и экологически сходных с ним ползунов с нерестом получала новые и новые подкрепления, но в явном виде сформулирована и обоснована не была. Основная причина этого, по всей видимости, сводится к тому, что никто так и не осуществил никаких целенаправленных исследований данного феномена.
МИГРАЦИИ РЫБ: ПРОБЛЕМЫ ОПРЕДЕЛЕНИЯ
Этот раздел следует начать с цитат из четырех монографий, посвященных миграциям животных, в том порядке, в котором эти книги выходили в свет, от начала XX до начала XXI века. Они достаточно наглядно иллюстрируют изменение настроения исследователей, занимающихся данным феноменом:
“В последние два десятилетия проблемы миграций исследовались со все возрастающей энергией, результатом чего стало накопление важных знаний, которых, как я надеюсь, уже достаточно для выявления и определения общих принципов” (Meek, 1916, p. 7).
“Затруднительно дать общепринятое определение миграции, поскольку разные люди, особенно исследователи разных групп животных, используют этот термин по-разному” (Aidley, 1981, p. 1).
“Пожалуй, никакой другой тип поведения не “пострадал” так сильно из-за неразберихи с определениями, как миграции” (Dingle, 1996, p. 20).
“Миграции, так же как искусство, – это что-то такое, о чем все знают, про суть чего все имеют твердое мнение, но что никто не может четко определить” (Milner-Gulland et al., 2011, p. 1).
Действительно, в случае с миграциями мы, к сожалению, имеем дело с ситуацией, когда накопление информации не только не приводит к формированию целостной концепции и общепринятой терминологии, но, скорее, напротив, еще больше размывает смысл понятий, кажущихся интуитивно самоочевидными. Существенная часть путаницы и противоречий возникла из-за того, что теоретические представления о данном типе активности независимо друг от друга формировали специалисты по разным группам животных, недостаточно хорошо знакомые с материалом, касающимся других организмов. В результате представления о миграциях планктонных ракообразных, насекомых, рыб, птиц и копытных до сих пор плохо соотносятся друг с другом (Aidley, 1981; Rankin, 1985; Dingle, 1996, 2014).
Еще одним важным источником взаимных недопониманий стала разница в подходе, наблюдающаяся даже в работах, посвященных одним и тем же животным. Миграции исследовали и анализировали как один из типов поведения, как элемент жизненного цикла, жизненной стратегии, как форму адаптации, как демографический процесс, как реакцию на изменение климата и т.п. В частности, многие теоретики экологии, математики, строящие экологические модели, генетики, занимающиеся экологическими и популяционными процессами, стали называть миграциями самые разные, обычно масштабные перемещения, безотносительно к особенностям поведения и образа жизни конкретных видов животных. Подчас подразумевались даже не перемещения как таковые, а довольно абстрактные (в масштабе жизни отдельных особей) демографические процессы, связанные с такими явлениями, как изменение границ ареала, поток генов и пр. (Levins, 1969; Brower, Malcolm, 1991).
Из терминологической размытости проистекают не только взаимные недопонимания между исследователями, но и серьезные ошибки выводов и интерпретаций, связанные, в частности, с неверной трактовкой работ других авторов (Olson et al., 2019). Несомненно, любая концепция, равно как и обслуживающая ее терминология, может и должна изменяться, но эти изменения, очевидно, должны приводить к развитию, а не к обессмысливанию. Еще Н.В. Тимофеев-Ресовский с соавторами (1973, с. 77) отмечали, что в биологии многие понятия и термины “употребляются без достаточно строгой и однозначной формулировки”.
Широко распространенная сейчас точка зрения на научную терминологию сводится к тому, что автор публикации должен в явном виде пояснять в своей работе те термины, которые имеют неоднозначную и спорную трактовку. Это несомненно верно, но, как представляется, часто недостаточно. Понятия, кажущиеся очевидными автору, могут быть восприняты совершенно иначе специалистами из других областей биологии, что мы и наблюдаем в случае с амфибийными рыбами. В целом ряде публикаций, посвященных, казалось бы, конкретным вопросам организации жизненных стратегий и поведения конкретных видов ползунов, встречается использование слова “миграции” для определения чуть ли не любых перемещений (Das, 1927). Кроме того, отказ от анализа проблемы и поиска конвенциональных решений не приводит к исчезновению самой проблемы, в то время как в научной норме “любая теория так или иначе требует точных понятий, как бы она это не скрывала” (т.е. как бы ученые ни пытались убедить всех и самих себя, что это не является нужным и существенным) (Уайтхед, 1990, с. 328).
Фактически, доминирующая на сегодняшний день точка зрения на миграции животных сводится к тому, что точного определения у данного феномена, как у целостного явления, нет, как нет у него и четких границ, отделяющих миграции от всех остальных форм двигательной активности животных. Означает ли это, что сам термин “миграции” утратил всякое конкретное значение и может использоваться как угодно и в какой угодно области? На мой взгляд, это не так. Многие авторы отмечают, что связанная с миграциями рыб терминология – это проблема не формальная и не семантическая, а намного более серьезная именно в биологическом аспекте (Dingle, 1996; Dingle, Drake, 2007; Morais, Daverat, 2016).
Специальный анализ чрезвычайно запутанной ситуации, связанной с термином “миграции”, не является предметом данного обзора. Текущее положение вещей достаточно подробно разобрано с разных позиций в нескольких современных монографиях и обзорах (Milner-Gulland et al., 2011; Dingle, 2014). Тем не менее важно отметить, что, на мой взгляд, в ситуации, когда единая система представлений о важном биологическом феномене отсутствует, при анализе поведения и жизненных стратегий конкретных видов животных и их групп правильно опираться на концепции, сформировавшиеся и развивающиеся именно в этой области, равно как и на терминологию, используемую в данной концепции. В частности, в рамках темы данного обзора нас прежде всего интересует система взглядов и понятий, существующая на сегодняшний день в ихтиологии и связанная с образом жизни, поведением, жизненными циклами и стратегиями рыб.
МИГРАЦИИ ИЛИ ПРОСТО ПЕРЕМЕЩЕНИЯ?
Представления о передвижениях рыб в пространстве в целом и, прежде всего, об их миграциях активно развиваются на протяжении уже более века (Meek, 1916; Шмидт, 1947; Myers, 1949; Павлов, 1979; Jungwirth et al., 1998; Lucas et al., 2001; Brönmark et al., 2014; Morais, Daverat, 2016). Несмотря на некоторые различия в понимании миграционной активности рыб, можно выделить ряд признаков, по которым любые миграции выделяются большинством специалистов из всего многообразия перемещений в пространстве. Прежде всего, это их облигатный характер, т.е. понимание миграционных циклов как неотъемлемого элемента жизненной стратегии (Никольский, 1963; Павлов, Скоробогатов, 2014).
Не менее важно, что миграции рыб являются либо элементом онтогенеза (как, например, покатные миграции икры и молоди), либо циклическим процессом, который происходит с регулярной периодичностью в течение жизни особи и приводит к чередованию двух или более типов биотопов. В миграционное движение вовлекается большая часть популяции, однонаправленно перемещающаяся по максимально короткому маршруту (Northcote, 1984; Lucas et al., 2001; Binder et al., 2011; Wootton, Smith, 2015). На сегодняшний день нет никаких причин считать выход ползунов на сушу облигатным элементом их жизненной стратегии, а значит, нет и достаточно веских причин называть такие перемещения миграциями. То же касается и большинства других критериев истинных миграций рыб.
Результаты исследований последних лет, выполненных с использованием недоступных ранее методик, указывают на то, что пластичность жизненных стратегий рыб существенно выше, чем было принято считать, в том числе и в том, что касается их миграций. В частности, было показано, что диадромные (прежде всего амфидромные) миграции некоторых видов можно рассматривать в виде континуума стратегий (Augspurger et al., 2017). Так называемые частичные миграции, давно и хорошо известные у птиц (Lack, 1968), были не так давно обнаружены и у рыб (Chapman et al., 2012). Все это, очевидно, приведет к дальнейшему развитию и коррекции взглядов на феномен в целом, однако на сегодняшний день вся совокупность сравнительно скудных знаний о биологии ползунов вполне укладывается в традиционные представления.
Одним из основных типов миграций рыб являются потамодромные миграции, полностью происходящие в пресной воде. В середине ХХ в. Даге (Daget, 1959) ввел в научный обиход понятие латеральных миграций как одной из форм потамодромных нерестовых миграций, в ходе которых пресноводные рыбы, обитающие большую часть жизненного цикла в реке, в период размножения заселяют залитую половодьем пойму, а после нереста возвращаются обратно в русло. Широкое распространение этот термин получил, прежде всего, применительно к тропическим рыбам благодаря работам Уэлкомма (Welcomme, 1985). Такое понимание латеральных миграций мы находим в публикациях, посвященных рыбам Азии (Hoggarth et al., 1999; Chanh et al., 2000; Qin et al., 2019), Африки (Bénech, Peñáz, 1995; Waal, 1996) и Южной Америки (Fernandes, 1997; Castello, 2008; Nunes et al., 2019). В настоящее время данное понятие начинает все более широко использоваться и применительно к рыбам умеренных широт (Lucas et al., 2001; Górski et al., 2010).
Довольно быстро произошло увеличение семантического объема понятия, и термин “латеральные миграции” стал использоваться как sensu stricto, так и sensu lato. В последнем случае он стал (1) обозначать не только нерестовые, но и любые другие миграции рыб из русла реки в пойму и обратно (Winemiller, Jepsen, 1998; Welcomme et al., 2006; Valbo-Jorgensen et al., 2009), либо (2) включать перемещения рыб между руслом и не только поймой, но и притоками (Borcherding et al., 2002; Makrakis et al., 2012). На мой взгляд, второй вариант не очень удачен и является еще одним примером того, как неточное использование терминов вызывает путаницу и приводит к обессмысливанию связанных с ними теорий.
При разработке концепции латеральных миграций использовалась понятийная база, принятая в гидрологии и, в частности, в потамологии (Lowe-McConnell, 1987; Welcomme et al., 2006; Wootton, Smith, 2015). Понятие “основное русло” (main channel) используется в ней как eupotamon и включает в себя как русло главной реки, так и русла притоков (Amoros et al., 1987; Ward, Tockner, 2001; Hohensinner et al., 2018). В свою очередь, пойма представляет собой заливаемую в половодье водой часть речной долины (Важнов, 1976). Вне периода половодья пойма может быть как сухой, так и заболоченной, некоторые поймы высыхают полностью, в других остаются постоянные пойменные водоемы. Соответственно, степень фрагментированности пойменных биотопов, во-первых, может существенно различаться, а во-вторых, является величиной изменчивой.
Миграции рыб из основного русла в пойму связаны с их перемещениями между лотическими и лентическими условиями среды. Если же называть латеральными миграциями перемещение из реки в притоки (т.е. внутри eupotamon), то смысл самого понятия латеральных миграций исчезает. По тем же причинам не совсем правильно называть латеральными миграциями перемещения рыб в пределах поймы, т.е. без выхода из лентических условий, даже если при этом осуществляется переход из одного водоема в другой.
СТРУКТУРА И ДИНАМИКА В ПОПУЛЯЦИЯХ ПОЛЗУНОВ
Для типичной популяционной структуры ползунов характерен ряд особенностей, важнейшая из которых заключается в том, что большую часть года (в сухой сезон) популяция состоит из дискретных изолированных друг от друг субпопуляций (локальных популяций), населяющих главным образом постоянные пойменные водоемы и иногда русловое прибрежье. В сезон дождей суммарная площадь занимаемых популяцией биотопов существенно возрастает и появляется возможность для перемещения и перераспределения особей между субпопуляциями.
Такая популяционная структура типична для так называемых популяционных систем, описанных Ю.П. Алтуховым и Ю.Г. Рычковым (1970). Практически аналогичной, но более широко известной и более детально разработанной является концепция метапопуляций, которые понимаются как длительно существующие совокупности связанных локальных популяций. Обязательной характеристикой метапопуляции является ее пространственная структурированность и средовая фрагментированность, определяемые биотопической неоднородностью, а ее единство поддерживается обменом особями, периодически, но не постоянно перемещающимися между субпопуляциями (Levins, 1969; Nouhuys, 2016).
Примечательно, что метапопуляционный подход оказался весьма продуктивным в исследованиях популяций лягушек и некоторых других земноводных, которые, как и амфибийные рыбы, размножаются сезонно в пойменных, в том числе временных, водоемах (Smith, Green, 2005; Petranka, 2007). Метапопуляции описаны и у некоторых рыб, прежде всего лососевых и коралловых (Kritzer, Sale, 2006; Schtickzelle, Quinn, 2007), однако, насколько мне известно, к амфибийным рыбам, в частности ползунам, такой подход никто не применял.
Анализ популяционной организации ползунов через призму концепции метапопуляций весьма перспективен, поскольку данная концепция рассматривает популяционную динамику как систему нескольких процессов разного характера (в том числе стохастических) и разного уровня (Hanski, Ovaskainen, 2019). Степень фрагментированности среды в популяциях ползунов, как уже было отмечено выше, характеризуется высокой изменчивостью. Причем компонентами этой изменчивости являются как циклические, так и флуктуирующие процессы. В частности, сезонность вносит вклад в первую группу компонентов, а межгодовые различия – во вторую.
Устойчивое существование метапопуляций ползунов не может объясняться одной лишь структурой и динамикой локальных популяций, составляющих ее, а зависит также от взаимодействия между ними, т.е. от комплекса процессов разного масштаба: локальных и региональных. Именно в контекст процессов регионального масштаба, связывающих субпопуляции между собой, и интегрированы различные наземные и водные перемещения ползунов. Опираясь на такой подход, можно предположить, что перемещения амфибийных рыб не являются каким-то одним определенным типом миграций, а представляют собой поведение, позволяющее рыбам решать целый комплекс задач, связанных с разными типами ресурсов и разными факторами среды. При этом характер решаемых задач будет меняться вслед за изменениями в биотопах.
Очевидно, что для таких исследований придется преодолеть ряд методологических сложностей, в частности связанных с учетом численности как взрослых особей, так и кладок. Если в случае с лягушками это не вызывает больших проблем, то для изучения, например, анабаса будет представлять отдельную задачу. Как уже было сказано выше, нерест анабаса в природе до сих пор не описан. Эти рыбы размножаются преимущественно ночью в мутной воде, в заросших растениями биотопах. Нерест у них порционный, происходит в пелагиали, после чего мелкая икра свободно расплывается по поверхности воды (Зворыкин, 2012). При этом икра развивается быстро, а личинки поначалу также обладают положительной плавучестью (Дзержинский и др., 2019).
С другой стороны, у некоторых других ползунов, например змееголовов, можно наблюдать как брачные пары, так и кладки икры и родительское поведение (Roberts, 1989; Haniffa et al., 2003). Соответственно, с их учетом возникнет меньше проблем. Кроме того, исследователю жизненных стратегий и поведения придется выделить из всех демографических процессов, именуемых в метапопуляционных моделях миграциями, те типы передвижений, которые его, собственно, и интересуют.
Для многих рыб с высокой пластичностью жизненных стратегий характерны различия в выраженности двигательной и в широком смысле миграционной активности, зависящие от стабильности и неоднородности среды. Эти различия могут проявляться в следовании одной из альтернативных стратегий на протяжении всей жизни, в формировании как внутрипопуляционной, так и внутривидовой дифференциации. В то же время они могут приводить к ситуационно зависимой лабильности поведения и образа жизни, включая территориальность, оседлость, различные перемещения и миграции. Биотопически высокодифференцированные метапопуляции рыб, способных жить и перемещаться в двух средах, представляют ситуацию, в которой подобное разнообразие альтернативных стратегий может проявиться широко и многообразно. Причем высокая неоднородность среды часто связана с выраженностью миграционной активности обратной зависимостью, что мы и наблюдаем при сопоставлении популяций анабаса, населяющего пойму и русловое прибрежье, о чем еще будет сказано ниже (подробнее об экологической значимости неоднородности среды и о ее связи с миграциями см., например, Михеев, 2006; Blanquart, Gandon, 2014).
ЧЕРНЫЕ ИЛИ БЕЛЫЕ?
Начиная с рубежа XX–XXI вв. различные авторы начинают использовать термин “латеральные миграции” применительно к анабасу и другим ползунам, совершающим в период дождей сухопутные перемещения по пойме. При этом часто характер этих “миграций”, их назначение и отличие от разрозненных и разнонаправленных перемещений отдельных особей никак не рассматриваются (Jamsari et al., 2010; Sheenaja, Thomas, 2011; Termvidchakorn, Hortle, 2013). В то же время часто контекст использования явно или косвенно предполагает связь между этими передвижениями анабаса и его размножением (Chanh et al., 2000; Poulsen, Valbo-Jørgensen, 2000; Павлов, Зворыкин, 2014; Зворыкин, 2016).
В итоге в научной литературе спонтанно оформилась практика, в рамках которой такие выражения, как “сухопутные перемещения”, “латеральные миграции” и “нерестовые миграции”, стали использоваться применительно к ползунам как контекстуальные или даже семантические синонимы. Сухопутные перемещения рыб в период дождей стали с какого-то времени рассматриваться в качестве нерестовых миграций a priori, по совокупности косвенных данных. Для того чтобы понять, как в действительности соотносятся между собой данные понятия в случае ползунов, необходимо рассмотреть образ их жизни.
Известно широко используемое разделение рыб Юго-Восточной Азии (прежде всего бассейна Меконга) на “белую” и “черную” рыбу (Welcomme, 1985; Hoggarth et al., 1999; Chanh et al., 2000; Valbo-Jorgensen et al., 2009). Для “белой” реофильной рыбы, примером которой могут служить крупные карповые и сомы, характерны выраженные, в том числе латеральные, миграции. В то время как для “черной” рыбы типично круглогодичное пребывание преимущественно в пойме, где в наиболее засушливое время эти рыбы концентрируются в постоянных пойменных водоемах. Помимо мелких сомов, карповых и ряда других видов, к “черной” рыбе большинство авторов относят и всех ползунов (Sao-Leang, Dom-Saveun, 1955; Poulsen et al., 2002; Valbo-Jorgensen et al., 2009; Hortle, Bamrungrach, 2015; Sultana et al., 2020).
Для рыб других тропических регионов можно встретить и иные разделения на экологические группы, которых может быть, разумеется, и больше двух (Welcomme et al., 2006; Winemiller et al., 2008; Makrakis et al., 2012). Однако суть их та же – разделение рыб на в той или иной мере дискретные экологические группы видов, каждая из которых обладает неслучайным сочетанием черт жизненной стратегии, включая тип нереста, наличие и характер нерестовых миграций, предпочитаемые биотопы и др. Этот подход в значительной мере основан на классических представлениях об экологических гильдиях как о “группах видов, которые эксплуатируют один и тот же класс ресурсов окружающей среды сходным образом” (Root, 1967, p. 335). В частности, эта концепция использовалась при разработке близких к нашей теме представлений о репродуктивных гильдиях Балоном (Balon, 1975, 1990) и экологических гильдиях Уэлкоммом с соавт. (Welcomme et al., 2006). Несмотря на определенную условность таких схем и пластичность жизненных стратегий многих рыб, данный подход позволяет увидеть основные особенности этих стратегий в виде закономерной системы.
Существует целый ряд исследований (Welcomme, 1985; Halls et al., 1998; Chanh et al., 2000), в которых было показано, что анабас, как и другие ползуны, действительно круглогодично обитает в пойме, если там сохраняются непересыхающие водоемы. Эти рыбы могут не совершать никаких миграций, но при этом перемещаться на короткие расстояния как по воде, так и по суше из постоянных пойменных водоемов во временные и обратно. Известны и другие данные, согласно которым в некоторых местах анабас постоянно встречается не только в пойме, но и в основном русле реки (Halls et al., 1998; Poulsen, Valbo-Jørgensen, 2000). Однако и в таких случаях авторы, как правило, отмечают, что основным местом его обитания все равно остается пойма, в реке же он встречается реже и в меньших количествах (Smith, 1945; Chanh et al., 2000; Poulsen, Valbo-Jørgensen, 2000).
По моим полевым наблюдениям, в реке анабас встречается преимущественно в прибрежных заводях, заросших растительностью, слабопроточных и мелководных биотопах, во многом сходных с пойменными. Более того, недавно опубликованы данные (Sultana et al., 2020), согласно которым ползуны, населяющие водно-болотные угодья Чалан Бил в Бангладеш, перемещаются оттуда на нерест в реку, а в конце сезона дождей возвращаются обратно. Неизвестно, насколько такие перемещения соответствуют миграциям в ихтиологическом значении этого термина, но даже если они являются таковыми, их направление прямо противоположно латеральным миграциям.
ЛАТЕРАЛЬНЫЕ, НЕРЕСТОВЫЕ, СУХОПУТНЫЕ?
В целом приведенные выше данные согласуются с результатами наших собственных исследований, посвященных главным образом анабасу. За период с 2009 по 2014 год в разных районах Вьетнама в разное время года нами было собрано несколько десятков выборок рыб данного вида. Информация по некоторым из них представлена в табл. 1. Во всех случаях материал был собран в утренние часы, в мелководных участках, одним заходом, с использованием невода длиной 5 м и ячеей 7 мм. Учтены только половозрелые особи размером 48–142 мм. Эти данные не позволяют произвести полноценный статистический анализ, однако общие тенденции очевидны. Анабасы встречаются во всех исследованных биотопах, за исключением открытых участков рек, преобладая при этом в пойменных водоемах.
Таблица 1.
Уловы анабасов в различных биотопах, бассейнах и районах Вьетнама
| Тип биотопа | Сезон | Река (бассейн), провинция Вьетнама | Количество особей | Дата |
|---|---|---|---|---|
| Открытая прибрежная отмель реки | Начало сухого сезона | р. Зинь, Кханьхоа | 0 | 16.12.2009 |
| Заросший мелкий речной приток | Сухой сезон | р. Кинь, Бариа-Вунгтау | 26 | 20.12.2012 |
| Заросшая макрофитами прибрежная зона реки | Начало сухого сезона | р. Зинь, Кханьхоа | 16 | 25.12.2009 |
| Заросшая макрофитами прибрежная зона реки | Конец сухого сезона | р. Ба, Зялай | 60 | 12.02.2014 |
| Заросшая макрофитами прибрежная зона реки | Сезон дождей | р. Кай, Кханьхоа | 31 | 04.10.2010 |
| Пойма | Сухой сезон | р. Меконг, Тьензянг | 55 | 24.12.2012 |
| Пойма | Сезон дождей | р. Серепок, Даклак | 79 | 31.10.2010 |
| Пойма | Сезон дождей | р. Кон, Биньдинь | 83 | 23.11.2010 |
| Постоянный пойменный водоем | Начало сухого сезона | р. Зинь, Кханьхоа | 38 | 21.12.2009 |
| Пойма, каналы рисовых полей | Конец сезона дождей | р. Даранг, Фуйен | 72 | 03.01.2012 |
Следует обратить внимание на тот факт, что в разных районах Вьетнама ситуация разная. В бассейнах рек Меконг и Красная, где площади водно-болотных угодий существенны (Sterling et al., 2006; Зворыкин, Гусаков, 2014), амфибийные рыбы в большей степени приурочены к пойменным водоемам. В то время как в Центральном Вьетнаме пойменных земель меньше и существенная их часть занята рисовыми полями, обрабатываемыми пестицидами, в связи с чем все ползуны вынуждены заселять речные прибрежья и мелкие притоки.
Большая часть наблюдаемых сухопутных перемещений ползунов произошла в пойме, т.е. между пойменными водоемами, а не между руслом и поймой. По всей видимости, в соответствии с доминирующим образом жизни, ползуны действительно должны быть с некоторыми оговорками отнесены к “черной” рыбе, чаще всего не совершающей ни латеральных, ни каких-либо других протяженных миграций. Однако, благодаря пластичной жизненной стратегии, в некоторых районах они могут перемещаться не только между пойменными водоемами различного типа, но и между поймой и руслом, а также совершать короткие продольные перемещения в основном русле реки, включая ее притоки (Halls et al., 1998). Связь продольных перемещений с нерестом неизвестна, а осуществляться они могут, в отличие от сухопутных передвижений, большую часть года, включая сухой сезон.
Что касается сухопутных перемещений ползунов, то их в экологическом аспекте так до сих пор никто целенаправленно и не исследовал. Очевидно, что они действительно приурочены к сезону дождей. Осадки, влажность и другие погодные факторы являются критичными для такого рода поведения, поскольку высыхание кожных покровов может быть фатальным для рыб, продолжительное время находящихся на суше. В то же время по-прежнему достоверно неизвестно, что заставляет этих рыб покидать воду; как они выбирают направление своего движения; ориентируются ли они в пространстве, находясь на земле, и если да, то благодаря каким органам чувств; возвращаются ли они в биотопы, которые покинули; на какие расстояния перемещаются.
Примечательно также, что практически во всех наблюдениях за передвигающимися по суше ползунами сообщается о взрослых особях. Если нерест этих рыб действительно осуществляется во временных водоемах, куда они приползают в начале сезона дождей, скорость развития потомства должна быть достаточной для того, чтобы молодь была способна на сухопутное возвращение в основной водоем до того, как нерестовый биотоп высохнет. Однако никакие данные о таких массовых перемещениях молоди в конце влажного сезона мне не известны. При этом продолжительность сезона дождей во многих регионах Азии может составлять всего два–три месяца.
Отметим также, что ползуны чаще всего встречаются в тех же водоемах, где и многие другие виды “черной” рыбы. Очевидно, что способность к сухопутным передвижениям может быть полезна для перемещения из одного биотопа в другой, но не ясно, насколько часто реализуется эта способность, в каких ситуациях она играет какую-то действительно значимую роль. И уж совершенно точно ни анабасы, ни большинство других ползунов не относятся к экстремофилам, способным жить в недоступных другим видам биотопах (Riesch et al., 2015; Wang, Guo, 2019).
Для пресноводных тропических рыб пик нереста в сезон дождей является достаточно типичной и во многом объяснимой особенностью (Lowe-McConnell, 1987; Wootton, Smith, 2015). Соответственно, сухопутные перемещения ползунов и их нерест осуществляются в одно и то же время. Однако совпадение двух событий по времени вовсе не означает наличие связи между ними. Гипотеза о наличии такой связи в данном случае не изучена и не подтверждена. Более того, мне не известно вообще ни одной публикации, в которой был бы описан нерест анабаса в природе, а все выводы о нерестовом сезоне этих рыб построены на косвенных данных, основанных на гистологическом анализе гонад (Jacob, 2005; Bernal et al., 2015). Как расселяется молодь, тоже неизвестно, хотя и существуют отрывочные данные о скате икры и личинок по руслу р. Кай во Вьетнаме (Нездолий и др., 2014).
Различные известные типы перемещений ползунов схематично изображены на рис. 1. Данная схема составлена на основании трех источников данных. Первым послужили мои собственные наблюдения за анабасами, змееголовами и рисовыми угрями, проводившиеся в сезон дождей в поймах рек Зинь (11.2009), Серепок (10.2010), Кон (11.2010) и Даранг (03.2012). В качестве второго источника использовались опросы местных рыбаков различных районов Вьетнама. Данный метод в настоящее время признается важным инструментом экологического познания (Bao et al., 2001; Hind, 2015; Nunes et al., 2019). На сегодняшний день уже можно говорить о формировании целой методологической концепции, названной Local ecological knowledge (LEK) и основанной на данном принципе (Brook, McLachlan, 2008). Наконец, был использован ряд литературных источников, уже упомянутых в данном обзоре (Halls et al., 1998; Poulsen, Valbo-Jørgensen, 2000, и др.).
Рис. 1.
Направления основных перемещений и миграций ползунов: а, б – водные латеральные, в, г – сухопутные латеральные, д – водные продольные, е – водные и сухопутные по пойме (не латеральные и не продольные). 1 – основное русло, 2 – пойма, 3 – приток, 4 – временный пойменный водоем, 5 – постоянный пойменный водоем, 6 – временный водоток.
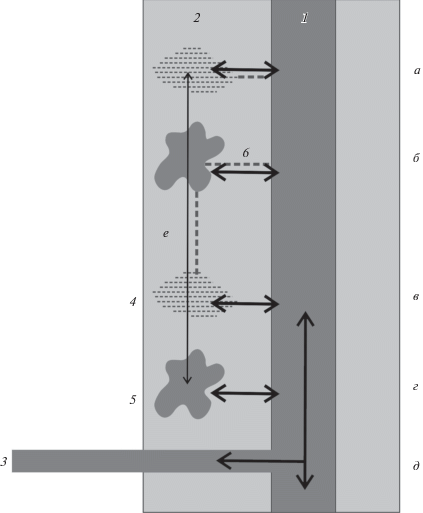
Видно, что перемещения ползунов могут быть как продольными, так и латеральными, и как водными, так и сухопутными в любых комбинациях. Очевидно также, что сами понятия “латеральный” и “сухопутный” применительно к перемещениям рыб относятся к разным понятийным категориям. Первое из них связано с направлением движения и является конкретным термином. Второе указывает на способ движения в рамках простой логической оппозиции “водный–сухопутный”. Третью категорию составляет назначение миграций или перемещений. В случае с ползунами мы пока вынуждены ограничиваться лишь предположениями о назначении и не можем называть какие бы то ни было из их перемещений ни нерестовыми, никакими иными, пока это не будет в явном виде доказано.
ЗАКЛЮЧЕНИЕ И ВЫВОДЫ
Таким образом, на сегодняшнем уровне знаний некорректно использовать понятия “сухопутные перемещения” и “латеральные миграции” ползунов в качестве синонимов и неверно априорно считать сухопутные перемещения нерестовыми миграциями. Большая часть этих перемещений, по всей видимости, вообще не является миграциями. Мы имеем дело с тремя разными феноменами, проявление и соотношение которых друг с другом у разных амфибийных рыб могут быть различными, а у большинства из них остаются неизученными.
Сухопутные перемещения рыб, несомненно, являются ярким феноменом, привлекающим внимание биологов и побуждающим к тому, чтобы атрибутировать его как не только уникальную, но и важную адаптацию. Склонность к адаптационизму, характерная для современной биологии (Wilkins, Godfrey-Smith, 2009; Koonin, 2016), располагает к этой в целом ошибочной тенденции в рассуждениях. В действительности достоверной информации, которая позволила бы адекватно оценить адаптивную значимость данного феномена, пока немного. Более того, совокупность имеющихся в распоряжении собственных и литературных данных, кратко изложенных в данном обзоре, позволяет предположить, что значимость сухопутных перемещений многих ползунов в их жизненном цикле, особенно как формы латеральных миграций, может быть существенно переоценена.
Именно то обстоятельство, что большая часть наблюдаемых сухопутных перемещений ползунов не имеет отношения к истинным миграциям в том смысле, который принят в современной ихтиологии, обеспечивает сравнительно высокую устойчивость этих рыб к антропогенным нагрузкам. Несмотря на существенное изменение среды обитания, связанное, прежде всего, с использованием пойменных земель в качестве сельскохозяйственных угодий, жизненный цикл ползунов фатально не нарушается, и они переходят к эксплуатации ресурсов других биотопов, главным образом руслового прибрежья. Такая пластичность стратегий была бы невозможна, если бы сухопутные перемещения являлись облигатным компонентом жизненного цикла, каковым по определению являются миграции, в том числе латеральные.
В случае с ползунами можно с той или иной степенью уверенности констатировать лишь небольшой набор подтвержденных фактов, касающихся их сухопутных передвижений. Эти рыбы предпочитают круглогодично оставаться в стоячих или слабопроточных водоемах поймы, но могут заселять и русловое прибрежье. Они ведут преимущественно оседлый образ жизни и перемещаются в основном на небольшие расстояния. Их сухопутные перемещения (назначение которых по-прежнему не вполне ясно) осуществляются главным образом в сезон дождей. Перемещения по суше не всегда являются миграциями и не обязательно связаны с нерестом. В свою очередь, размножение ползунов часто не связано ни с миграциями, ни с сухопутными перемещениями. Пик нереста ползунов приходится на сезон дождей, но отдельные порции икры могут выметываться и в сухое время года. Миграции ползунов если и осуществляются, то лишь в некоторых типах биотопов и только на короткие расстояния, при этом они могут быть как продольными, так и латеральными. Одним из перспективных подходов, который мог бы прояснить некоторые аспекты организации сухопутных и водных перемещений ползунов, является концепция метапопуляций.
Работа выполнена в рамках программы Эколан Э-3.2., раздел “Размножение, репродуктивное и пищевое поведение лабиринтовых рыб (Anabantoidei)” Российско-Вьетнамского Тропического научно-исследовательского и технологического центра. Автор благодарит Динь Тхи Хай Йен и Во Тхи Ха (Приморское отделение Тропического центра) за многолетнее сотрудничество в процессе исследований анабаса, а В.Н. Михеева и Д.С. Павлова (ИПЭЭ РАН) за внимательное прочтение рукописи, ценные замечания и рекомендации.
Список литературы
Алтухов Ю.П., Рычков Ю.Г., 1970. Популяционные системы и их структурные компоненты. Генетическая стабильность и изменчивость // Журн. общ. биологии. Т. 31. № 5. С. 507–525.
Важнов А.Н., 1976. Гидрология рек. М.: Изд-во МГУ. 339 с.
Гришина Е.А., Ляшевская О.Н., 2020. Грамматический словарь новых слов русского языка. http://dict.ruslang.ru/gram.php
Дзержинский К.Ф., Зворыкин Д.Д., Будаев С.В., 2019. Особенности раннего онтогенеза анабаса Anabas testudineus (Anabantoidei), связанные с динамикой плавучести // Вопр. ихтиологии. Т. 59. № 5. С. 579–589.
Зворыкин Д.Д., 2012. Размножение и репродуктивное поведение рыбы-ползуна Anabas testudineus (Perciformes, Anabantidae) в аквариуме // Вопр. ихтиологии. Т. 52. № 4. С. 469–479.
Зворыкин Д.Д., 2016. Филогенез репродуктивных стратегий у ползуновидных рыб (Anabantoidei) и в сестринских группах // Журн. общ. биологии. Т. 77. № 6. С. 464−481.
Зворыкин Д.Д., 2020. Некоторые тенденций пресноводной тропической аквакультуры и их влияние на исследования репродуктивной биологии анабаса Anabas testudineus (Anabantidae) // Рыбное хозяйство. № 4. С. 85–89.
Зворыкин Д.Д., Гусаков В.А., 2014. География, климат и водные ресурсы Вьетнама // Экология внутренних вод Вьетнама. М.: Т-во науч. изд. КМК. С. 9–14.
Михеев В.Н., 2006. Неоднородность среды и трофические отношения у рыб. М.: Наука. 191 с.
Нездолий В.К., Павлов Д.С., Нго Чи Тхьен, Нгуен Зуй Тоан, Нгуен Куок Кхань, 2014. Покатная миграция молоди рыб в реке Кай // Экология внутренних вод Вьетнама. М.: Т-во науч. изд. КМК. С. 300–319.
Никольский Г.В., 1963. Экология рыб. М.: Высш. шк. 368 с.
Павлов Д.С., 1979. Биологические основы управления поведением рыб в потоке воды. М.: Наука. 319 с.
Павлов Д.С., Зворыкин Д.Д., 2014. Миграции пресноводных рыб Вьетнама // Экология внутренних вод Вьетнама. М.: Т-во науч. изд. КМК. С. 279–285.
Павлов Д.С., Скоробогатов М.А., 2014. Миграции рыб в зарегулированных реках. М: Т-во науч. изд. KMK. 413 с.
Тимофеев-Ресовский Н.В., Яблоков А.В., Глотов Н.В., 1973. Очерк учения о популяции. М.: Наука. 278 с.
Тихонов А.Н., 2002. Морфемно-орфографический словарь русского языка. М.: АСТ, Астрель. 704 с.
Уайтхед А., 1990. Очерки науки и философии // Избранные работы по философии. М.: Прогресс. С. 304–336.
Шмидт П.Ю., 1947. Миграции рыб. 2-е изд. M.; Л.: Изд-во АН СССР. 362 с.
Aidley D.J., 1981. Questions about migration // Animal Migration. Cambridge, UK: Cambridge Univ. Press. P. 1–8.
Amoros C., Roux A.L., Reygrobellet J.L., Bravard J.P., Pautou G., 1987. A method for applied ecological studies of fluvial hydrosystems // Regul. River. V. 1. № 1. P. 17–36.
Augspurger J.M., Warburton M., Closs G.P., 2017. Life-history plasticity in amphidromous and catadromous fishes: A continuum of strategies // Rev. Fish Biol. Fisher. V. 27. № 1. P. 177–192.
Balon E.K., 1975. Reproductive guilds of fishes: A proposal and definition // J. Fish. Res. Board Can. V. 32. № 6. P. 821–864.
Balon E.K., 1990. Epigenesis of an epigeneticist: The development of some alternative concepts on the early ontogeny and evolution of fishes // Guelph Ichthyol. Rev. V. 1. P. 1–48.
Banerji S.R., Prasad D., 1974. Observation on reproduction and survival of Anabas testudineus (Bloch) in Bihar region (India) // J. Inland Fish. Soc. India. V. 6. P. 6–17.
Bao T.Q., Bouakhamvongsa K., Chan S., Chhuon K.C., Phommavong T. et al., 2001. Local Knowledge in the Study of River Fish Biology: Experiences from the Mekong // Mekong development series № 1. Phnom Penh: Mekong River Commission. 22 p.
Bayley M., Damsgaard C., Thomsen M., Malte H., Wang T., 2019. Learning to air-breathe: The first steps // Physiology. V. 34. № 1. P. 14–29.
Bénech V., Peñáz M., 1995. An outline on lateral fish migrations within the Central Delta of the Niger River, Mali // Hydrobiologia. V. 303. № 1–3. P. 149–157.
Bernal R.A.D., Aya F.A., Jesus-Ayson E.G.T., de, Garcia L.M.B., 2015. Seasonal gonad cycle of the climbing perch Anabas testudineus (Teleostei: Anabantidae) in a tropical wetland // Ichthyol. Res. V. 62. № 4. P. 389–395.
Binder T.R., Cooke S.J., Hinch S.G., 2011. The biology of fish migration // Encyclopedia of Fish Physiology: From Genome to Environment. San Diego: Academic Press. P. 1921–1927.
Blanquart F., Gandon S., 2014. On the evolution of migration in heterogeneous environments // Evolution. V. 68. № 6. P. 1617–1628.
Bloch M.E., 1792. Naturgeschichte der Ausländischen Fische. T. 6. Berlin: Auf Kosten des Verfassers. 126 s.
Borcherding J., Bauerfeld M., Hintzen D., Neumann D., 2002. Lateral migrations of fishes between floodplain lakes and their drainage channels at the Lower Rhine: Diel and seasonal aspects // J. Fish Biol. V. 61. № 5. P. 1154–1170.
Bressman N.R., Love J.W., King T., Horne C., Ashley-Ross M.A., 2019. Emersion and terrestrial locomotion of the northern snakehead (Channa argus) on multiple substrates // Integr. Org. Biol. V. 1. № 1. P. 1–20.
Brönmark C., Hulthén K., Nilsson P.A., Skov C., Hansson L.-A. et al., 2014. There and back again: Migration in freshwater fishes // Can. J. Zool. V. 92. № 6. P. 467–479.
Brook R.K., McLachlan S.M., 2008. Trends and prospects for local knowledge in ecological and conservation research and monitoring // Biodivers. Conserv. V. 17. № 14. P. 3501–3512.
Brower L.P., Malcolm S.B., 1991. Animal migrations: Endangered phenomena // Am. Zool. V. 31. № 1. P. 265–276.
Bruton M.N., 1979. The survival of habitat desiccation by air breathing clariid catfishes // Environ. Biol. Fish. V. 4. № 3. P. 273–280.
Castello L., 2008. Lateral migration of Arapaima gigas in floodplains of the Amazon // Ecol. Fresh. Fish. V. 17. № 1. P. 38–46.
Chanh S., Chhuon K.C., Valbo-Jørgensen J., 2000. Lateral fish migrations between Tonlé Sap River and its flood plain // 3th Tech. Symp. Mekong Fish. Phnom Penh, 8–9th December 2000. P. 102–114.
Chapman B.B., Skov C., Hulthén K., Brodersen J., Nilsson P.A. et al., 2012. Partial migration in fishes: Definitions, methodologies and taxonomic distribution // J. Fish Biol. V. 81. № 2. P. 479–499.
Daget J., 1959. Les migrations des poissons dans les eaux douces tropicales africaines // Notes et documents de la pêche et pisciculture. 5 DG. P. 1–5.
Daldorff, 1797. Natural history of Perca scandens // Trans. Linn. Soc. Lond. V. 3. № 1. P. 62–63.
Das B.K., 1927. The bionomics of certain air-breathing fishes of India, together with an account of the development of their air-breathing organs // Philos. Trans. R. Soc. Lond. B. Biol. Sci. V. 216. P. 183–219.
Davenport J., Abdul Matin A.K.M., 1990. Terrestrial locomotion in the climbing perch, Anabas testudineus (Bloch) (Anabantidea, Pisces) // J. Fish Biol. V. 37. № 1. P. 175–184.
Dingle H., 1996. Migration: The Biology of Life on the Move. 1st ed. N.Y.; Oxford: Oxford Univ. Press. 480 p.
Dingle H., 2014. Migration: The Biology of Life on the Move. 2nd ed. N.Y.; Oxford: Oxford Univ. Press. 326 p.
Dingle H., Drake V.A., 2007. What is migration? // BioScience. V. 57. № 2. P. 113–121.
Du T.Y., Standen E.M., 2020. Terrestrial acclimation and exercise lead to bone functional response in Polypterus senegalus pectoral fins // J. Exp. Biol. V. 223. № 11. P. jeb217554.
Fernandes C.C., 1997. Lateral migration of fishes in Amazon floodplains // Ecol. Freshw. Fish. V. 6. № 1. P. 36–44.
Fishman A.P., Pack A.I., Delaney R.G., Galante R.J., 1986. Estivation in Protopterus // J. Morphol. V. 190. № S1. P. 237–248.
Górski K., Winter H.V., Leeuw J.J., de, Minin A.E., Nagelkerke L.A.J., 2010. Fish spawning in a large temperate floodplain: The role of flooding and temperature // Freshwater Biol. V. 55. № 7. P. 1509–1519.
Graham J.B., 1997. Air-Breathing Fishes: Evolution, Diversity, and Adaptation. San Diego; L.: Academic Press. 299 p.
Halls A.S., Hoggarth D.D., Debnath K., 1998. Impact of flood control schemes on river fish migrations and species assemblages in Bangladesh // J. Fish Biol. V. 53. Suppl. A. P. 358–380.
Hamilton F., 1822. Account of the Fishes Found in the River Ganges and Its Branches. Edinburgh: George Ramsay & Co. 405 p.
Haniffa M.A., Nagarajan M., Marimuthu K., Raj A.J.A., 2003. Embryonic and larval development of spotted murrel, Channa punctatus (Bloch) // Indian J. Fish. V. 50. № 3. P. 355–362.
Hanski I., Ovaskainen O., 2019. Metapopulation models // Encyclopedia of Ecology. Elsevier. P. 136–144.
Helfman G.S., Collette B.B., Facey D.E., Bowen B.W., 2009. The Diversity of Fishes: Biology, Evolution, and Ecology. 2nd ed. Oxford: Wiley-Blackwell. 736 p.
Hind E.J., 2015. A review of the past, the present, and the future of fishers’ knowledge research: A challenge to established fisheries science // ICES J. Mar. Sci. V. 72. № 2. P. 341–358.
Hoggarth D.D., Cowan V.J., Halls A.S., Aeron-Thomas M., McGregor J.A. et al., 1999. Management Guidelines for Asian Floodplain River Fisheries. Pt. 2. Summary of DFID Research. Rome: FAO Fisheries Technical Paper № 384/2. 117 p.
Hohensinner S., Hauer C., Muhar S., 2018. River morphology, channelization, and habitat restoration // Riverine Ecosystem Management. Aquatic Ecology Series. V. 8. Cham: Springer. P. 41–65.
Hortle K.G., Bamrungrach P., 2015. Fisheries Habitat and Yield in the Lower Mekong Basin. Phnom Penh: MRC Technical Paper № 47. 80 p.
Inger R.F., Kong C.P., 1962. The Fresh-Water Fishes of North Borneo. Chicago: Chicago Natural History Museum. 268 p.
Jacob P.K., 2005. Studies on Some Aspects of Reproduction of Female Anabas testudineus (Bloch). Cochin, India: CUSAT. 245 p.
Jamsari A.F.J., Muchlisin Z.A., Musri M., Siti Azizah M.N., 2010. Remarkably low genetic variation but high population differentiation in the climbing perch, Anabas testudineus (Anabantidae), based on the mtDNA control region // Genet. Mol. Res. V. 9. № 3. P. 1836–1843.
Jungwirth M., Schmutz S., Weiss S., 1998. Fish Migration and Fish Bypasses. Oxford; Malden, MA: Blackwell Science. 438 p.
Koonin E.V., 2016. Splendor and misery of adaptation, or the importance of neutral null for understanding evolution // BMC Biol. V. 14. № 1. P. 114.
Kritzer J.P., Sale P.F., 2006. The metapopulation ecology of coral reef fishes // Marine Metapopulations. Amsterdam: Academic Press. P. 31–67.
Lack D., 1968. Bird migration and natural selection // Oikos. V. 19. № 1. P. 1–9.
Land S.C., Bernier N.J., 1995. Estivation: Mechanisms and control of metabolic suppression // Biochemistry and Molecular Biology of Fishes. V. 5. Oxford: Elsevier. P. 381–412.
Levins R., 1969. Some demographic and genetic consequences of environmental heterogeneity for biological control // Bull. Entomol. Soc. Amer. V. 15. № 3. P. 237–240.
Liem K.F., 1987. Functional design of the air ventilation apparatus and overland excursions by Teleosts // Fieldiana Zool. V. 37. P. 1–29.
Lowe-McConnell R.H., 1987. Ecological Studies in Tropical Fish Communities. Cambridge: Cambridge Univ. Press. 382 p.
Lucas M.C., Baras E., Thom T.J., Duncan A., Slavík O., 2001. Migration of Freshwater Fishes. Oxford: Blackwell Science Ltd. 420 p.
Mai H.V., Tran L.X., Dinh Q.M., Tran D.D., Murata M. et al., 2019. Land invasion by the mudskipper, Periophthalmodon septemradiatus, in fresh and saline waters of the Mekong River // Sci. Rep. V. 9. № 1. P. 14227.
Makrakis M.C., Miranda L.E., Makrakis S., Fontes Júnior H.M., Morlis W.G. et al., 2012. Diversity in migratory patterns among Neotropical fishes in a highly regulated river basin // J. Fish Biol. V. 81. № 2. P. 866–881.
Meek A., 1916. The Migrations of Fishes. L.: Edward Arnold. 427 p.
Milner-Gulland E.J., Fryxell J.M., Sinclair A.R.E. (eds.), 2011. Animal Migration: A Synthesis. Oxford: Oxford Univ. Press. 269 p.
Mookerjee H.K., Mazumdar S.R., 1946. On the life history, breeding and rearing of Anabas testudineus Bloch // J. Dep. Sci. Cal. Univ. V. 2. P. 101–140.
Morais P., Daverat F. (eds.), 2016. An Introduction to Fish Migration. Boca Raton: CRC Press. 315 p.
Myers G.S., 1949. Usage of anadromous, catadromous and allied terms for migratory fishes // Copeia. № 2. P. 89–97.
Northcote T.G., 1984. Mechanisms of fish migration in rivers // Mechanisms of Migration in Fishes. N.-Y.: Plenum. P. 317–355.
Nouhuys S., van, 2016. Metapopulation ecology // eLS. Chichester: John Wiley & Sons. P. 1–9.
Nunes M.U.S., Hallwass G., Silvano R.A.M., 2019. Fishers’ local ecological knowledge indicate migration patterns of tropical freshwater fish in an Amazonian river // Hydrobiologia. V. 833. № 1. P. 197–215.
Olson M.E., Arroyo-Santos A., Vergara-Silva F., 2019. A user’s guide to metaphors in ecology and evolution // Trends Ecol. Evol. V. 34. № 7. P. 605–615.
Petranka J.W., 2007. Evolution of complex life cycles of amphibians: Bridging the gap between metapopulation dynamics and life history evolution // Evol. Ecol. V. 21. № 6. P. 751–764.
Poulsen A.F., Valbo-Jørgensen J. (eds.), 2000. Fish Migrations and Spawning Habits in the Mekong Mainstream: A Survey Using Local Knowledge. Vientiane: Mekong River Commission. 150 p.
Poulsen A.F., Poeu O., Viravong S., Suntornratana U., Tung N.T., 2002. Fish Migrations of the Lower Mekong River Basin: Implications for Development, Planning and Environmental Management. Phnom Penh: MRC Technical Paper № 8. 62 p.
Qin X., Gong Z., Liu H., 2019. Lateral migration of fish between China’s second largest freshwater lake (Dongting Lake) and the mainstem of the Yangtze River // Environ. Biol. Fish. V. 102. № 4. P. 527–539.
Rahman A.K.A., 1989. Freshwater Fishes of Bangladesh. Dhaka: Dep. Zool., Univ. Dhaka. 364 p.
Rankin M.A. (ed.), 1985. Migration: Mechanisms and Adaptive Significance. Contrib. Marine Science. V. 27. Austin: Univ. Texas Press. 868 p.
Riesch R., Tobler M., Plath M. (eds.), 2015. Extremophile Fishes. Cham: Springer. 326 p.
Roberts T.R., 1989. The Freshwater Fishes of Western Borneo (Kalimantan Barat, Indonesia). San Francisco: California Academy of Sciences. 210 p.
Root R.B., 1967. The niche exploitation pattern of the blue-gray gnatcatcher // Ecol. Monogr. V. 37. P. 317–350.
Sao-Leang, Dom-Saveun, 1955. Aperçu général sur la migration et la reproduction des poissons d’eau douce du Cambodge // Proc. IPFC. V. 5. № 2–3. P. 138–142.
Sayer M.D.J., Davenport J., 1991. Amphibious fish: Why do they leave water? // Rev. Fish Biol. Fisher. V. 1. № 2. P. 159–181.
Schtickzelle N., Quinn T., 2007. A metapopulation perspective for salmon and other anadromous fish // Fish Fish. V. 8. P. 297–314.
Sharples R.W., 1992. “On fish” // Theophrastus. His Psychological, Doxographical and Scientific Writings. Rutgers University Studies in Classical Humanities. V. 5. New Brunswick, NJ: Transaction Publishers. P. 347–385.
Sheenaja K.K., Thomas K.J., 2011. Influence of habitat complexity on route learning among different populations of climbing perch (Anabas testudineus Bloch, 1792) // Mar. Freshw. Behav. Physiol. V. 44. № 6. P. 349–358.
Smith H.M., 1945. The Fresh-Water Fishes of Siam, or Thailand. Washington: Bull. U. S. Natl. Mus. V. 188. 447 p.
Smith M.A., Green D.M., 2005. Dispersal and the metapopulation paradigm in amphibian ecology and conservation: Are all amphibian populations metapopulations? // Ecography. V. 28. № 1. P. 110–128.
Sterling E.J., Hurley M.M., Minh L.D., 2006. Vietnam: A Natural History. New Haven; L.: Yale Univ. Press. 423 p.
Sultana N., Chowdhury M.A., Shahnawaj M., 2020. Seasonal movement of fish species of Rajshahi and Khulna division in Bangladesh // Int. J. Fish. Aquat. Stud. V. 8. № 3. P. 639–645.
Tay Y.L., Loong A.M., Hiong K.C., Lee S.J., Tng Y.Y.M. et al., 2006. Active ammonia transport and excretory nitrogen metabolism in the climbing perch, Anabas testudineus, during 4 days of emersion or 10 minutes of forced exercise on land // J. Exp. Biol. V. 209. № 22. P. 4475–4489.
Termvidchakorn A., Hortle K.G., 2013. A Guide to Larvae and Juveniles of Some Common Fish Species from the Mekong River Basin. MRC Technical Paper. Phnom Penh: Mekong River Commission. 248 p.
Thakur N.K., Das P., 1986. Synopsis of Biological Data on Koi, Anabas testudineus (Bloch, 1792). Barrackpore, India: Central Fisheries Research Institute. Bull. 40. 47 p.
Valbo-Jorgensen J., Coates D., Hortle K., 2009. Fish diversity in the Mekong River basin // The Mekong. Biophysical Environment of an International River Basin. N.Y.: Academic Press. P. 161–196.
Waal B.C.W., van der, 1996. Some observations on fish migrations in Caprivi, Namibia // South. Afr. J. Aquat. Sci. V. 22. № 1–2. P. 62–80.
Wang Y., Guo B., 2019. Adaption to extreme environments: A perspective from fish genomics // Rev. Fish Biol. Fisher. V. 29. № 4. P. 735–747.
Ward J.V., Tockner K., 2001. Biodiversity: Towards a unifying theme for river ecology // Freshwater Biol. V. 46. № 6. P. 807–819.
Welcomme R.L., 1985. River Fisheries. Rome: FAO Fisheries technical paper 262. 330 p.
Welcomme R.L., Winemiller K.O., Cowx I.G., 2006. Fish environmental guilds as a tool for assessment of ecological condition of rivers // River Res. Appl. V. 22. № 3. P. 377–396.
Wicaksono A., Hidayat S., Retnoaji B., Alam P., 2020. The water-hopping kinematics of the tree-climbing fish, Periophthalmus variabilis // Zoology. V. 139. P. 125750.
Wilkins J.F., Godfrey-Smith P., 2009. Adaptationism and the adaptive landscape // Biol. Philos. V. 24. № 2. P. 199–214.
Winemiller K.O., Jepsen D.B., 1998. Effects of seasonality and fish movement on tropical river food webs // J. Fish Biol. V. 53. Suppl. A. P. 267–296.
Winemiller K.O., Agostinho A.A., Caramaschi É.P., 2008. Fish ecology in tropical streams // Tropical Stream Ecology. L.: Academic Press. P. 107–146.
Wootton R.J., Smith C., 2015. Reproductive Biology of Teleost Fishes. Chichester, UK: John Wiley & Sons, Ltd. 496 p.
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии