

Журнал общей биологии, 2022, T. 83, № 1, стр. 71-80
Адаптивный потенциал амброзии полыннолистной (Ambrosia artemisiifolia L., Asteraceae) в связи с ее продвижением на север: опыт биоклиматического и эколого-географического анализа и моделирования распространения инвазивного вида
А. Н. Афонин 1, *, О. Г. Баранова 2, Ю. Ю. Кулакова 3, Ю. А. Федорова 4, Д. Р. Владимиров 5, А. В. Герус 6, Е. Ю. Герус 6, А. Я. Григорьевская 5, Т. Ю. Закота 6
1 Санкт-Петербургский государственный университет
199034 Санкт-Петербург, Университетская наб., 7/9, Россия
2 Ботанический институт им. В.Л. Комарова РАН
197376 Санкт-Петербург, ул. Проф. Попова, 2, Россия
3 Всероссийский центр карантина растений
140150 Раменское, р. п. Быково, ул. Пограничная, 32, Россия
4 Уфимский Институт Биологии УФИЦ РАН
450054 Уфа, просп. Октября, 69, Россия
5 Воронежский государственный университет
394068 Воронеж, ул. Хользунова, 40, Россия
6 Всероссийский научно-исследовательский институт защиты растений
196608 Санкт-Петербург, ш. Подбельского, 3, Россия
* E-mail: acer737@yandex.ru
Поступила в редакцию 13.10.2021
После доработки 03.11.2021
Принята к публикации 26.11.2021
- EDN: NNOFCH
- DOI: 10.31857/S0044459622010080
Аннотация
В основе оперативной генотипической адаптации растений к недостатку тепла лежит регуляция перераспределения ресурсов тепла между вегетативной и генеративной фазами развития. У короткодневных растений, цветущих при сокращающейся длине дня, перераспределение ресурсов тепла между фазами регулируется механизмом фотопериодической чувствительности. Растения, зацветающие при более длинном дне, переходят в генеративную фазу развития раньше и тем самым получают больше ресурсов тепла на созревание семян. При этом сокращается продолжительность и теплообеспеченность вегетативной фазы, в результате чего уменьшается вегетативный рост растений и потенциальная семенная продуктивность. Это приводит к снижению конкурентоспособности растений в ценозах. Оптимальный баланс между продолжительностью вегетативной и генеративной фаз в каждой зоне индивидуален и достигается в процессе естественного отбора генотипов на оптимальный для условий зоны порог фотопериодической чувствительности. В статье рассмотрены особенности порогов фотопериодической чувствительности растений амброзии полыннолистной Ambrosia artemisiifolia (Asteraceae) с южной и северной границ вторичного ареала в связи с особенностями динамики фотопериода и температурных условий их местообитаний. Дается прогноз возможности продвижения ареала к северу, которая связана как с потенциальной вариабельностью генотипов, так и с тенденциями потепления климата.
Условием выживания популяций однолетних видов растений является их способность стабильно из года в год формировать за период вегетации число семян, достаточное для перманентного возобновления популяции. Перманентность воспроизводства достигается сбалансированностью сезонной и многолетней динамики развития растений с локальной динамикой экологических факторов среды – абиотических и биотических. В условиях дефицита ресурсов тепла цикл онтогенеза растений должен укладываться в период вегетации, возможный на территории их распространения. Потребности растений натурализовавшейся популяции в тепле не могут выходить за пределы теплообеспеченности территории.
Жизненный цикл однолетних растений подразделяется на периоды вегетативного и репродуктивного развития. В вегетативный период происходит рост растений и формирование их габитуса. При этом решаются задачи конкуренции с другими растениями в ценозе и формируется потенциал для последующего продуцирования семян. В репродуктивный период, если рассматривать его от начала пыления и завязывания семян, прекращается вегетативный рост растений, и основные потоки ассимилятов перенаправляются на созревание семян. Оптимальный баланс в распределении ресурсов тепла между вегетативной и репродуктивной фазами может быть разным для разных климатических зон. Удлинение продолжительности и, соответственно, теплообеспеченности одной фазы снижает теплообеспеченность другой. Удлинение продолжительности репродуктивной фазы гарантирует большую вероятность вызревания семян даже в самые холодные годы, но при этом замедляется вегетативный рост растений, в связи с чем, в частности, снижается их конкурентоспособность.
Дата перехода от вегетативного к репродуктивному развитию определяется настройкой чувствительности растений к внешним сигнальным факторам. Для короткодневных видов сигнальным фактором является сокращение длины дня до определенного порогового значения. Зацветание растений при более длинном дне увеличивает период их репродуктивного развития, при более коротком – увеличивает период вегетативного развития.
В основе адаптивной стратегии инвазивных видов растений при продвижении их на север лежит прежде всего модификация систем чувствительности к пороговым значениям сигнальных факторов среды. Генетически обусловленные различия порога фотопериодической чувствительности у растений одной популяции и вариация этих порогов у разных популяций позволяют синхронизировать динамику развития растений с разнообразными локальными особенностями сезонной динамики экологических факторов среды в зоне инвазии. При оптимальном пороге фотопериодической чувствительности растения локальной популяции переходят от вегетативной к репродуктивной фазе развития в оптимальные даты. Оптимальной для перехода к генеративному развитию считается дата, которая обеспечивает наилучший баланс вегетативного и генеративного развития растений популяции, и в итоге как успешное противодействие биоценотическому прессу, так и стабильное и достаточное для перманентного существования популяции воспроизводство фертильных семян.
Амброзия полыннолистная (Ambrosia artemisiifolia L.) – вредоносный вид, засоряющий поля и вызывающий приступы астмы у людей, страдающих аллергией на его пыльцу. Амброзия была занесена в Евразию из Америки и начала свое распространение более столетия назад. В настоящее время северная граница ее массовой натурализации на европейской территории России проходит по Курской, Воронежской, Саратовской областям. Вторичный ареал амброзии до сих пор не стабилизировался. Поэтому перспективы расширения ареала вида вызывают большой интерес и тревогу. A. artemisiifolia – короткодневное растение, и ее цветение приходится на конец лета, когда длина дня уменьшается до порогового значения, которое является сигналом для начала цветения (Allard, 1945). Порог зацветания растений северных популяций амброзии приходится на более длинный день, чем южных, т.е. на более ранние даты (Dickerson, Sweet, 1971). Это увеличивает продолжительность репродуктивного периода и предоставляет дополнительные ресурсы тепла, необходимые завязавшимся семенам для созревания. Поэтому потенциал продвижения амброзии на север характеризуется прежде всего размахом вариации особей и популяций вида по значениям порога фотопериодической чувствительности.
В ранее проведенных географических опытах с популяциями амброзии полыннолистной разного происхождения были выявлены различия в сроках зацветания северных и южных популяций (Genton et al., 2005; Leiblein-Wild et al., 2014; Li et al., 2015; Scalone et al., 2016). Было установлено, что растения северных популяций в условиях более высоких широт переходят в генеративную фазу раньше, чем растения южных популяций. При этом связь потенциала распространения и натурализации растений, характеризующихся разными фотопериодическими порогами, с температурными условиями разных климатических зон количественно не оценивалась.
Цель нашего исследования – изучить вариацию порогов фотопериодической чувствительности растений популяций A. artemisiifolia разного происхождения, связав ее с региональной динамикой фотопериода и температурных условий периода созревания семян. На основе выявленных закономерностей определить экологический потенциал распространения A. artemisiifolia на север.
МАТЕРИАЛЫ И МЕТОДЫ
Наблюдения за динамикой развития растений популяций A. artemisiifolia проводили в 2019–2020 гг. в четырех регионах России: г. Славянск-на-Кубани (Краснодарский край), г. Воронеж, г. Москва и г. Уфа.
Растения местных популяций изучали в естественных условиях в Славянске-на-Кубани (45.23° с.ш., 38.16° в.д.), расположенном в зоне экологического оптимума амброзии, и в Воронеже (51.72° с.ш., 39.21° в.д.) – вблизи современной северной границы натурализации вида. Изучаемые растения отмечали бирками. В Краснодарском крае изучено 16 нумерованных растений, в Воронеже – 30. Фенологические наблюдения и биометрические измерения проводили на индивидуальных маркированных растениях по единой методике с интервалом в одну неделю. Наблюдения начинали проводить с начала июля – до перехода самых ранних растений в генеративную фазу. Измеряли высоту растений, динамику развития – число пар листьев на главном побеге, регистрировали даты перехода растений в генеративную фазу, начало и интенсивность пыления, дату появления первых зрелых плодов. С начала созревания плодов еженедельно оценивали динамику семенной продуктивности. Для этого проводили сбор созревших плодов со всего растения или, если растение крупное, с модельного среднего по семенной продуктивности побега. В последнем случае семенную продуктивность всего растения рассчитывали перемножением числа созревших плодов на модельном побеге на число побегов на растении.
Кроме наблюдений за местными популяциями, в 2020 г. по аналогичной методике в вегетационных опытах был изучен одинаковый географический набор, включающий растения южной и северных популяций. Изучение проводили на стационарах в Москве (55.64° с.ш., 38.10° в.д.) и Уфе (54.76° с.ш., 56.13° в.д.), расположенных несколько севернее границы натурализации амброзии. Для посева были использованы семена, собранные в 2019 г. Семена растений южной популяции были собраны в Славянске-на-Кубани на дамбе р. Протоки, координаты 45.25° с.ш., 38.16° в.д. Семена растений трех популяций с северной границы вторичного ареала (северные популяции) были собраны на пустыре в Воронеже (51.72° с.ш., 39.21° в.д.) и на обочинах автомобильных дорог вблизи с. Каплино (Белгородская обл.; 51.35° с.ш., 37.82° в.д.) и п. Горшечного (Курская обл.; 51.52° с.ш., 37.99° в.д.). Наблюдения проводили за 10 растениями популяций разного происхождения, выращиваемыми в естественных климатических условиях.
Перевод фенологических дат в градусо-дни (суммы активных температур выше 10°С) производили по среднесуточным температурным данным ближайших метеостанций. Корректность использования метеостанционных данных для конкретных условий стационаров поверялась данными, снятыми с логгеров, которые закладывали на стационарных площадках. За порог активной вегетации амброзии принимали температуру выше 10°С. При оценке связи динамики развития амброзии с температурным фактором рассчитывали кумулятивные показатели сумм активных температур с порогом выше 10°С (САТ10). По аналогичной методике проводили наблюдения и в географических опытах на стационарах.
Примененная при оценке экологического потенциала продвижения A. artemisiifolia на север методика эколого-географического анализа и моделирования описана в работе А.Н. Афонина и Ю.В. Соколовой (2018). Для проведения эколого-географического моделирования были составлены карты сумм температур за период созревания семян – от даты начала пыления (переход через фотопериодические пороги 14, 15 и 15.5 ч) до окончания активной вегетации: карты САТ10фп14, САТ10фп15, САТ10фп15.5.
ОПРЕДЕЛЕНИЕ СУММ ТЕМПЕРАТУР, НЕОБХОДИМЫХ ДЛЯ СОЗРЕВАНИЯ СЕМЯН A. ARTEMISIIFOLIA
Продолжительность созревания семян обусловлена температурным фактором. Считается, что длина дня не оказывает влияния на продолжительность созревания семян от момента их завязывания (Deen et al., 1998). Завязывание семян происходит при попадании пыльцы на рыльце пестика. В качестве маркера начала завязывания семян была принята фенологическая дата начала пыления. Суммы температур, необходимые для созревания первых семян маркированных растений, определяли как разницу между накопленными САТ10 на дату появления первых зрелых плодов на растении и САТ10 на дату начала пыления.
Средние значения САТ10 за период созревания семян отличались у исследованных южных и северных популяций в их естественных местообитаниях (табл. 1).
Таблица 1.
Суммы активных температур (САТ10), необходимые для появления первых зрелых семян A. artemisiifolia на растениях двух популяций разного происхождения в их естественных местообитаниях
| Параметры | Суммы активных температур, °С | |
|---|---|---|
| южная (славянская) | северная (воронежская) | |
| Среднее | 716 | 608 |
| Минимальное | 374 | 503 |
| Максимальное | 1009 | 794 |
| Стандартное отклонение (SD) | 167.6 | 68.1 |
| Количество растений | 16 | 30 |
Для успешной адаптации к условиям северной зоны начало пыления амброзии должно приходиться на дату, сумма температур от которой до конца вегетации гарантированно составляет не менее достаточных значений для вызревания минимального пула семян. Для северной воронежской популяции минимальное количество САТ10, достаточных для появления первых зрелых семян, составляет по нашим наблюдениям 608°С.
РОЛЬ ФОТОПЕРИОДИЧЕСКОЙ ЧУВСТВИТЕЛЬНОСТИ В АДАПТАЦИИ ПОПУЛЯЦИЙ A. ARTEMISIIFOLIA К ЛОКАЛЬНЫМ УСЛОВИЯМ ТЕПЛООБЕСПЕЧЕННОСТИ
Пыление амброзии растянуто во времени и начинается, как правило, с раскрывания нескольких цветков самых нижних корзинок терминальной кисти. В последующие дни последовательно раскрываются и пылят остальные цветки корзинки. Одновременно, по мере созревания верхних корзинок кисти, их цветки также переходят к последовательному пылению. Боковые кисти дифференцируются и переходят к цветению позднее, чем терминальная. Все это в итоге приводит к растянутости периода пыления особи.
Отдельные особи популяции несколько различаются по порогу чувствительности к фотопериодическому сигналу, что приводит к неодновременности их зацветания и еще большей растянутости периода пыления в целом. Например, по наблюдениям на Славянском стационаре (45.2° с.ш.), самое раннее из отмеченных растений местной популяции начало пылить 7 августа при 14.5-часовом дне (т.е. при длине дня 14 ч 30 мин) и завершило пыление 4 сентября. Самые поздние из отмеченных в Славянске растений начинали пыление в начале сентября (13.1-часовой день) и закончили пылить в конце сентября. Таким образом, пыление особи может быть растянуто на месяц, а для популяции в целом этот период может длиться до двух месяцев и более. Средняя для всех изученных растений местной славянской популяции дата начала пыления пришлась на 20 августа. Длина дня на эту дату составила 13.9 ч. Эту длину дня можно считать фотопериодическим порогом пыления местной популяции амброзии, хотя понятно, что фотопериодический сигнал был принят растениями еще раньше, и переход в пылящее соцветие потребовал некоторого времени.
В Славянске подобная настройка механизма фотопериодического порога пыления приводит к тому, что первые завязи семян у растений местной популяции начинают образовываться в среднем с 20 августа, и до окончания периода активной вегетации на созревание семян популяция имеет САТ10 порядка 1158°С за среднемноголетний период (рис. 1).
Рис. 1.
Теплообеспеченность (50-процентная) периода созревания семян местной популяции A. artemisiifolia в Славянске-на-Кубани за период 1989–2018 гг. Ориентация начала пыления на 13.9-часовой день в условиях Славянска-на-Кубани за среднемноголетний период предоставляет местной популяции сумму активных температур 1158°C с порогом выше 10°C на созревание семян от начала их завязывания до окончания периода вегетации. САТ10 периода вегетативного роста при этом составляет 2817°C (соответствует площади фигуры между осью абсцисс, линией графика и отрезками, спроецированными из соответствующих пограничных точек графика на ось абсцисс).
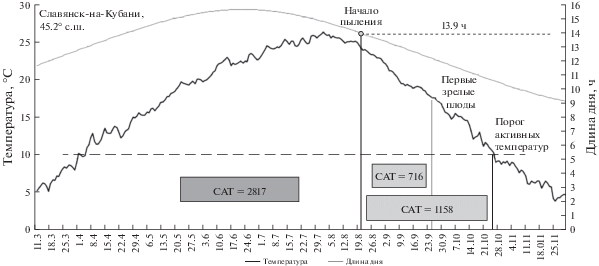
Учитывая, что на юге для формирования зрелых плодов A. artemisiifolia от момента их завязывания достаточно порядка 716°С (табл. 1), а физиологические САТ10 от даты появления первых зрелых плодов до массового плодоношения составляют порядка 300°С, ориентация на 13.9-часовой день гарантирует популяции амброзии полыннолистной в условиях южной зоны стабильную теплообеспеченность периода вызревания не только самых ранних завязей, но и самой массовой когорты семян, завязавшихся в период пика пыления.
Однако подобная настройка механизма фотопериодической чувствительности не универсальна в экологическом отношении для условий других зон. В географическом опыте 2020 г. растения южной славянской популяции, посеянные на более северных стационарах – в Москве (55.6° с.ш.) и Уфе (54.7° с.ш.), – перешли к пылению при длине дня, близкой к той, при которой они начинали пылить в Славянске – 13.6- и 13.5-часовой день соответственно (табл. 2). Но такая длина дня в Москве, в отличие от Славянска, приходится не на вторую декаду августа, а на 4 сентября. По среднемноголетним данным сумма активных температур выше 10°C от этой даты до конца активной вегетации составляет в Москве всего 238°C. Поэтому ресурсов тепла в Москве в среднемноголетнем аспекте недостаточно для созревания семян даже самых раннеспелых растений славянской популяции. Условия осени конкретного 2020 г. в Москве были аномально теплыми, и сумма активных температур от 4 сентября до конца вегетации составила 499°C. Но и этой суммы температур хватило на формирование только сравнительно небольшого количества зрелых семян у самых раннеспелых растений славянской популяции (табл. 3).
Таблица 2.
Длина дня на даты начала пыления южной и северных популяций A. artemisiifolia в местах натурализации и при высаживании их на северных стационарах
| Место | Славянская (Краснодарский край, 45.2° с.ш.) | Воронежская (Воронежская обл., 51.6° с.ш.) | Горшечное (Курская обл., 51.5° с.ш.) | Каплино (Белгородская обл., 51.4° с.ш.) | |
|---|---|---|---|---|---|
| В местных условиях | Дата | 20 августа | 10 августа | – | – |
| Длина дня, ч | 13.9 | 14.9 | – | – | |
| В Москве (55.6° с.ш.) | Дата | 4 сентября | 15 августа | 13 августа | 10 августа |
| Длина дня, ч | 13.6 | 15.1 | 15.2 | 15.4 | |
| В Уфе (54.7° с.ш.) | Дата | 4 сентября | 16 августа | 12 августа | 1 августа |
| Длина дня, ч | 13.5 | 14.8 | 14.9 | 15.8 |
Таблица 3.
Семенная продуктивность растений популяций A. artemisiifolia разного происхождения на Московском стационаре в связи с количеством тепла (САТ10) за период созревания их семян (стационар ВНИИКР, Москва, 55.6° с.ш., 2020 г.)
| Параметры | Славянская (Краснодарский край, 45.2° с.ш.) (n = 10) | Воронежская (Воронежская обл., 51.6° с.ш.) (n = 10) | Каплино (Белгородская обл., 51.4° с.ш.) (n = 10) | Горшечное
(Курская обл., 51.5° с.ш.) (n = 7) |
||||
|---|---|---|---|---|---|---|---|---|
| среднее | SD | среднее | SD | среднее | SD | среднее | SD | |
| САТ10 от начала пыления до конца активной вегетации, °C | 499 | 58.6 | 845 | 51.0 | 927 | 126.9 | 877 | 173.4 |
| Средняя семенная продуктивность на растение | 264 | 458.4 | 1083 | 429.1 | 1399 | 410.3 | 1210 | 410.8 |
Как происходила адаптация A. artemisiifolia к условиям более северной зоны? Продвижение амброзии полыннолистной на север происходило постепенно из южного очага – предположительно, из Предкавказья и/или Крыма (Марьюшкина, 1986; Afonin et al., 2018) с начала 20-го столетия. Возможность продвижения обеспечивалась естественным отбором адаптированных к северным условиям генотипов из первоначальной инвазивной популяции и, возможно, дополнительным внешним заносом генотипов, еще более адаптированных к условиям северных зон (Genton et al., 2005). К настоящему времени некоторые из самых северных натурализовавшихся популяций A. artemisiifolia отмечены на севере Курской и Воронежской областей.
Стационарное изучение местной популяции амброзии в Воронеже показало, что растения с северной границы вторичного ареала отличаются от растений южных популяций прежде всего порогом фотопериодической чувствительности. Самые ранние растения местной популяции начинали пыление в Воронеже 31 июля, т.е. при 15.5-часовом дне, самые поздние 20 августа – при 14.4-часовом дне. Средняя по отмеченным растениям местной воронежской популяции дата начала пыления пришлась в 2020 г. на 10 августа, длина дня на эту дату (фотопериодический порог завязывания семян местной популяции) составила 14.9 ч. В условиях Воронежа среднемноголетняя 50-процентная теплообеспеченность периода от начала пыления местных популяций, если оно ориентировано на 14.9-часовой день, составляет до окончания активной вегетации САТ10 = 819°C (рис. 2).
Рис. 2.
Теплообеспеченность (50%-ная) периода созревания семян местной популяции A. artemisiifolia в Воронеже за период 1998–2018 гг. Ориентация начала пыления на 14.9-часовой день в условиях Воронежа предоставляет местной популяции в среднем за многолетний период сумму активных температур САТ10 = 819°C на созревание семян от начала их завязывания до окончания периода вегетации.
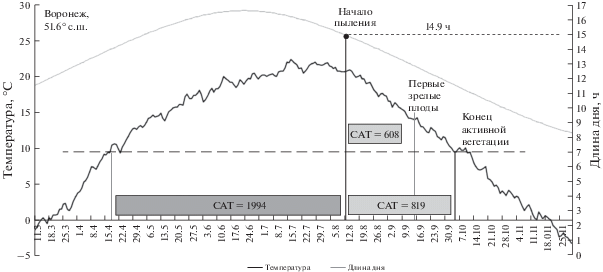
Учитывая, что для созревания первых семян от даты начала пыления воронежской популяции необходима САТ10 = 608°C (табл. 1), а САТ10 от даты начала плодоношения до массового плодоношения составляет порядка 300°C, ориентация на 14.9-часовой день гарантирует воронежской популяции амброзии полыннолистной в местных условиях стабильную теплообеспеченность периода вызревания семян только ранних генотипов. Вероятность вызревания семян, завязавшихся в пик пыления изученной местной популяции, составляет в условиях Воронежа менее 50%. Смещение пика пыления на дату негарантированного вызревания семян может быть обусловлено как недолгой историей распространения A. artemisiifolia в Воронежской области, так и эффективностью механизма твердосемянности, позволяющего надежно депонировать семена более урожайных позднеспелых генотипов в почвенном банке. В такой ситуации механизм адаптации, базирующийся на производстве большего числа семян при меньшей гарантии, может иметь преимущество перед механизмом, основывающимся на гарантированном во всем ряде лет вызревании небольшого числа семян.
Аналогичными фотопериодическими порогами, способствующими раннему началу пыления, характеризуются растения и других популяций с северной границы распространения A. artemisiifolia на европейской территории России: Горшечное (Курская обл.) – 14.9–15.2-часовой день, Каплино (Белгородская обл.) – 15.4–15.8-часовой день (табл. 2).
Отметим, что растения одной популяции в условиях разных зон зацветают при примерно одинаковой длине дня, и что растения северных популяций зацветают при более длинном дне, чем южных.
Отбор генотипов, ориентированных на зацветание при более длинном дне, в условиях северного предела распространения A. artemisiifolia приводит к удлинению репродуктивного периода за счет сокращения периода вегетативного роста. В Воронежской области среднемноголетняя теплообеспеченность периода вегетативного роста для местной натурализовавшейся популяции составляет САТ10 = 1994°C по сравнению с 2817°C для местной популяции, натурализовавшейся в Славянске-на-Кубани (рис. 1 и 2). Сокращение периода вегетативного роста приводит к уменьшению теплообеспеченности этого периода и формированию упрощенного габитуса растений, а, следовательно, к падению конкурентоспособности растений в фитоценозе. Конкурентоспособность может снижаться как непосредственно из-за замедления вегетативного роста, а значит высоты растений, проективного покрытия листьев, мощности корневой системы, так и из-за сокращения потенциала семенной продуктивности, которая может быть связана с уменьшением перечисленных параметров и числа плодоносящих побегов. В Краснодарском крае мощные растения A. artemisiifolia конкурентоспособны в посевах подсолнечника и кукурузы. На севере Воронежской области амброзия уходит с полей и ее значительно более мелкие растения распространены преимущественно в рудеральных местообитаниях с нарушенным растительным покровом – на откосах дорог, участках строительства. Приспосабливаясь к условиям северной зоны увеличением продолжительности репродуктивного периода и, соответственно, повышая его теплообеспеченность, амброзия жертвует конкурентоспособностью в ценозах.
ЭКОЛОГО-ГЕОГРАФИЧЕСКИЙ ПОТЕНЦИАЛ ПРОДВИЖЕНИЯ A. ARTEMISIIFOLIA НА СЕВЕР: ВПЕРЕДИ МОСКВА?
Насколько соответствует фотопериодическая настройка растений A. artemisiifolia с современной северной границы ареала возможности дальнейшего продвижения этого вида на север? Ответ на этот вопрос дает опыт выращивания растений A. artemisiifolia севернее современного предела распространения амброзии.
В проведенном нами в 2020 г. географическом опыте в Москве растения воронежской популяции начинали пыление 15 августа при 15.1-часовом дне – это примерно соответствует пороговой длине дня на дату начала пыления растений этой популяции и под Воронежем (14.9 ч). В условиях теплого 2020 г. сумма активных температур от этой даты до конца вегетации составила 845°C (рис. 3б), примерно столько, сколько характерно в среднем и для Воронежа. Такое количество тепла позволило растениям воронежской популяции образовать в 2020 г. в Москве существенное число зрелых семян – более тысячи на растение. В условиях аномально теплого года аналогичную высокую семенную продукцию в условиях Москвы продемонстрировали растения и других популяций амброзии с северных границ ее распространения – из Белгородской и Курской областей (табл. 3).
Рис. 3.
Теплообеспеченность (50%-ная) периода созревания семян популяций A. artemisiifolia из Воронежа (51.6° с.ш.) и Славянска-на-Кубани (45.2° с.ш.) в Москве за период 1998–2018 гг. (а), и количество тепла за период созревания семян этих популяций в 2020 году (б). Ориентация начала пыления южной славянской популяции на 13.6-часовой день в условиях Подмосковья предоставляет ей в среднем сумму активных температур 238°C с порогом выше 10°C на созревание семян от начала их завязывания – этого недостаточно для созревания семян. Более северная воронежская популяция, ориентированная на зацветание в условиях Подмосковья на 15.1-часовой день, имеет в среднем САТ10 = 554°C, чего также недостаточно для формирования стабильного ежегодного урожая семян.
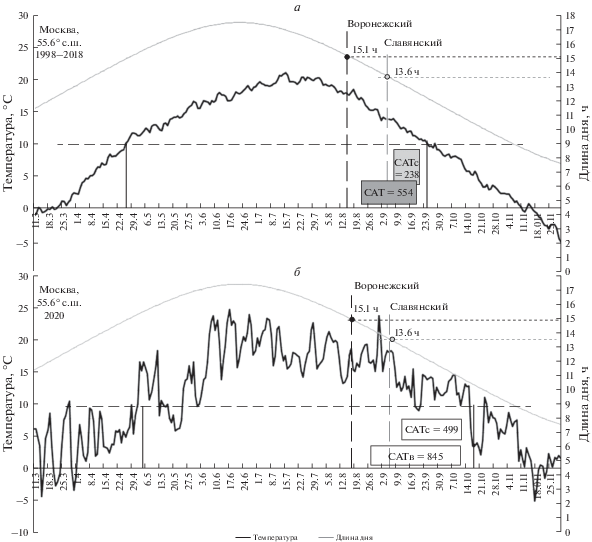
Однако средняя (50%-ная) за последние 30 лет теплообеспеченность в Москве за период от 15 августа до осеннего перехода среднесуточной температуры через 10°C составляет только 554°C (рис. 3а).
Такое количество тепла является недостаточным для стабильного ежегодного вызревания семян воронежских генотипов.
Метеостанционные архивные данные позволяют проанализировать динамику теплообеспеченности периода созревания семян A. artemisiifolia в последние десятилетия. Если считать САТ10 = 608°C (табл. 1) от начала пыления до конца активной вегетации минимумом, достаточным для вызревания необходимого для натурализации амброзии полыннолистной числа семян, то в Москве количество тепла, достаточное для вызревания самых раннеспелых из известных генотипов, приступающих к пылению при 15.5-часовом дне, в период 1989–1998 гг. отмечалось в 5 годах из 10 (табл. 4).
Таблица 4.
Теплообеспеченность периода созревания семян A. artemisiifolia по десятилетиям в Москве и Воронеже с 1989 по 2018 год (выражена в числе лет из 10 с САТ10 ≥ 608°C)
| Метеостанции | Москва (27612) | Воронеж (34123) | ||||||
|---|---|---|---|---|---|---|---|---|
| 14 ч/30.08* | 15 ч/16.08 | 15.5 ч/9.08 | 16 ч/2.08 | 14 ч/24.08 | 15 ч/9.08 | 15.5 ч/31.07 | 16 ч/21.07 | |
| 1989–1998 | 0 | 2 | 5 | 8 | 2 | 10 | 10 | 10 |
| 1999–2008 | 0 | 6 | 10 | 10 | 3 | 10 | 10 | 10 |
| 2009–2018 | 0 | 7 | 10 | 10 | 7 | 10 | 10 | 10 |
В последующие два десятилетия наблюдалось потепление климата, и теплообеспеченность последовательно возрастала. С 1999 г. для генотипов с порогом зацветания 15.5 ч все годы в Москве были достаточно теплыми для формирования того или иного числа семян. То, что такие генотипы до сих пор не получили распространения в Москве, может быть объяснено их редкостью (в популяции Горшечного таких растений примерно 1 из 10, в Каплинской популяции – 4 из 10), пониженной конкурентоспособностью раннеспелых генотипов, эффективной работой карантинных служб – массированные укосы обочин и рудеральных местообитаний в городской черте стали повсеместным в последнее десятилетие. Также это может быть обусловлено длительностью инвазионного процесса: отбора устойчивых генотипов, их размножения и распространения.
КАРТОГРАФИРОВАНИЕ ЭКОЛОГО-ГЕОГРАФИЧЕСКОГО ПОТЕНЦИАЛА ПРОДВИЖЕНИЯ A. ARTEMISIIFOLIA НА СЕВЕР
Биоэкологические исследования предоставляют данные, необходимые для прогноза распространения и сезонного развития видов. Ключевыми параметрами для биоклиматического прогноза распространения короткодневных видов растений являются вариации порогов чувствительности к сигнальным факторам среды и суммы температур, необходимые для прохождения ключевых фаз развития. Количественные параметры, полученные нами в результате стационарных географических исследований и проведенного биоклиматического анализа популяций амброзии разного происхождения, позволяют уточнить эколого-географический потенциал продвижения на север генотипов A. artemisiifolia с разными фотопериодическими порогами пыления и представить потенциал натурализации амброзии в картографическом виде.
Мы рассмотрели перспективы продвижения на север генотипов с порогами начала пыления при 14-, 15-, 15.5- и 16-часовом дне. Были составлены четыре карты сумм активных температур с соответствующими фотопериодическими порогами: САТфп14, САТфп15, САТфп15.5 и САТфп16. Пределом распространения считали изолинии сумм температур 608°C (см. табл. 1). Генотипы, переходящие к пылению при более коротком дне (более позднеспелые), имеют меньший потенциал распространения на север. Самые раннеспелые из известных к настоящему времени генотипы, переходящие к пылению при 15.5-часовом дне, имеют наибольший потенциал распространения и в условиях современного климата способны натурализоваться на европейской территории России и сопредельных стран: до северных границ Литвы, Минской, Могилевской, Брянской, Ульяновской и Тульской областей, юга Московской и севера Рязанской области, Мордовии, юга Татарстана и Башкортостана (рис. 4).
Рис. 4.
Эколого-географический потенциал распространения на север европейской территории России экотипов A. artemisiifolia с разной фотопериодической чувствительностью. Синим цветом показана зона, к условиям теплообеспеченности которой могут адаптироваться растения, начало пыления которых ориентировано на 15–15.5-часовой день. Оранжевая зона оптимальна для экотипов, ориентированных на пыление при более чем 14-часовом дне. Зеленая зона по условиям теплообеспеченности периода созревания семян пригодна для экотипов с порогами фотопериодической чувствительности менее 14 ч. В отмеченной желтым зоне возможна натурализация предполагаемых генотипов с фотопериодической чувствительностью 15.5–16 ч или фотопериодически нейтральных. Следует отметить, что распространение амброзии на юго-восток сдерживается дополнительным лимитирующим фактором – недостаточной влагообеспеченностью. Пунктирной линией показана современная северная граница натурализации A. artemisiifolia в России.

ЗАКЛЮЧЕНИЕ
Проведенное исследование показывает, что основным оперативным механизмом адаптации короткодневных видов растений при продвижении в высокие широты служит изменение порогов их фотопериодической чувствительности. Растения характеризуются широкой вариацией фотопериодической чувствительности как на меж-, так и внутрипопуляционном уровнях. Диапазон вариации фотопериодической чувствительности характеризует адаптивный потенциал видов при продвижении их в высокие широты.
Изучение диапазона порогов фотопериодической чувствительности популяций предоставляет исследователям инструмент для прогноза распространения видов на север.
Исследование фотопериодических порогов перехода к репродуктивной фазе популяций A. artemisiifolia показывает, что вид обладает потенциалом распространения до широты Москвы. Акклиматизация самых раннеспелых генотипов в Москве тем более вероятна, поскольку город является островом тепла. Появление более раннеспелых (с 16-часовым порогом), в том числе фотопериодически нейтральных генотипов, и дальнейшее потепление климата могут продвинуть потенциальную границу распространения A. artemisiifolia еще дальше на север. Сдерживающим экспансию фактором может служить снижение конкурентоспособности раннеспелых генотипов A. artemisiifolia в фитоценозах и эффективная работа служб защиты растений.
Список литературы
Афонин А.Н., Соколова Ю.В., 2018. Эколого-географический анализ и моделирование распространения биологических объектов с использованием ГИС. СПб.: Изд-во ВВМ. 121 с.
Марьюшкина В.Я., 1986 Амброзия полыннолистная и основы биологической борьбы с ней. Киев: Наукова думка. 120 с.
Afonin A.N., Luneva N.N., Fedorova Y.A., Kletchkovskiy Yu.E., Chebanovskaya A.F., 2018. History of introduction and distribution of common ragweed (Ambrosia artemisiifolia L.) in the European part of the Russian Federation and in the Ukraine // EPPO Bull. V. 48. № 2. P. 266–273.
Allard H.A., 1945. Flowering behavior and natural distribution of the Eastern ragweeds (Ambrosia) as affected by length of day // Ecology. V. 26. № 4. P. 387–394.
Deen W., Hunt T., Swanton C.J., 1998. Influence of temperature, photoperiod, and irradiance on the phenological development of common ragweed (Ambrosia artemisiifolia) // Weed Sci. V. 46. P. 555–560.
Dickerson C.T., Sweet R.D., 1971. Common ragweed ecotypes // Weed Sci. V. 19. № 1. P. 64–66.
Genton B.J., Shykoff J.A., Giraud T., 2005. High genetic diversity in French invasive populations of common ragweed, Ambrosia artemisiifolia, as a result of multiple sources of introduction // Mol. Ecol. V. 14. № 14. P. 4275–4285.
Leiblein-Wild M., Tackenberg O., 2014. Phenologic variation of 38 European Ambrosia artemisiifolia populations measured in a common garden experiment // Biol. Invasions. V. 16. № 9. P. 2003–2015.
Li X.-M., She D.-Y., Zhang D.-Y., Liao W.-J., 2015. Life history trait differentiation and local adaptation in invasive populations of Ambrosia artemisiifolia in China // Oecologia. V. 177. № 3. P. 669–677.
Scalone R., Lemke A., Štefanić E., Kolseth A.-K., Rašić S., Andersson L., 2016. Phenological variation in Ambrosia artemisiifolia L. facilitates near future establishment at northern latitudes // PLoS One. V. 11. № 11. https://doi.org/10.1371/journal.pone.0166510
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии