

Журнал общей биологии, 2022, T. 83, № 6, стр. 434-449
Бюджет углерода в экосистемах среднетаежных коренных ельников
К. С. Бобкова 1, М. А. Кузнецов 1, *
1 Институт биологии Коми научного центра УрО РАН
167982 Сыктывкар, Коммунистическая, 28, Россия
* E-mail: kuznetsov_ma@ib.komisc.ru
Поступила в редакцию 06.07.2022
После доработки 08.11.2022
Принята к публикации 16.11.2022
- EDN: TWWHVQ
- DOI: 10.31857/S0044459622060033
Аннотация
В работе приведена характеристика основных составляющих углеродного цикла в экосистемах спелых среднетаежных ельников. Объекты исследования представлены ельниками разнотравно-черничными на подзолистых и чернично-сфагновыми на торфянисто-подзолисто-глееватых почвах. Приходно-расходные статьи углеродного бюджета фитоценозов оценены по показателям, характеризующим продукционные процессы органического вещества. Определены фитомасса и прирост надземных органов древесных растений методом модельных деревьев. Масса корней определена по методу крупных и мелких монолитов. Количество опада растений древесного яруса выявлено с помощью опадоуловителей. Массу травяных растений, кустарничков и мхов определяли методом укосов на 10 площадках размером 0.5 × 0.5 м. Опад растений травяно-кустарничкого яруса оценивали по их приросту. Запасы органического углерода в почвах определяли по его содержанию в различных горизонтах с учетом их мощности и плотности. Эмиссию СО2 измеряли, используя метод темных камер при помощи ИКГ LI-COR 8100 (LI-COR Biosciences, США). Общий пул углерода в экосистемах старовозрастных ельников составляет 184.4–187.7 т га–1, из них в растениях фитоценоза 48–49%, в крупных древесных остатках (сухостой и валеж) – 6–8%. В почве, включая подстилку и минеральный слой, запасается 44–45% от общего пула углерода, примерно третья часть которого концентрируется в органогенном горизонте мощностью 7.7 см в ельнике разнотравном черничном и 13.1 см в ельнике чернично-сфагновом. Выявлено, что в старовозрастных ельниках фитоценоз оказывает существенное влияние на динамику содержания растворенного углерода органического вещества в атмосферных осадках и в почвенных водах. Ежегодное депонирование углерода фитоценозами ельников равно 3.30 в разнотравно-черничном и 3.08 т га–1 – в чернично-сфагновом. В продукции углерода исключительно важная роль принадлежит древостою. Количество углерода, поступающего с опадом, составило 2.48–2.84 т га–1 год–1. В общем потоке углерода в атмосферу доля минерализационных потерь за счет разложения растительных остатков в ельниках составила менее 30%. Установлено влияние на процесс выделения диоксида углерода с поверхности почвы погодных условий сезона, отмечена положительная корреляция эмиссии СО2 с температурой почвы (r = 0.6–0.8). Эмиссия СО2 с поверхности почвы обоих исследуемых ельников составила 2.47 т С га–1 сезон–1. Показано, что коренные среднетаежные ельники служат местом стока углерода в размере 0.36 и 0.87 т га–1 год–1.
Бореальные леса являются крупнейшим в мире депо углерода, однако реакция их на прогнозируемое изменение климата неопределенна (Stocker et al., 2013). С позиций оценки углеродного цикла лесные экосистемы представляют собой систему блоков – растительность, почва, крупные древесные остатки (КДО), атмосфера – характеризующихся определенными запасами углерода органического вещества (ОВ). Они связаны между собой его потоками: фотосинтетической ассимиляцией углерода атмосферы в нетто-продукции (Net Primary Production, NPP) и освобождением, возвратом его в атмосферу в ходе разложения ОВ. Соотношение интенсивностей этих процессов характеризует роль лесного сообщества в углеродном цикле биосферы и определяет, является ли та или иная экосистема стоком или источником углерода (Лесные экосистемы…, 2002; Кудеяров и др., 2007).
Большинство экосистем хвойных сообществ таежной зоны являются стоком углерода (Valentini et al., 2000; Коренные еловые…, 2006; Goulden et al., 2011; Ведрова и др., 2018; Uri et al., 2019, и др.). Однако есть работы, в которых показано, что лесные экосистемы в отдельные периоды могут выступать и источником углерода (Lindroth et al., 1998; Milyukova et al., 2002; Бобкова, 2007; Hadden, Grelle, 2016, и др.). Даже незначительные изменения между запасами и потоками ОВ в лесах бореальной зоны могут оказать существенное влияние на глобальный углеродный бюджет атмосферы (Johnston et al., 2004). Следовательно, важно определить динамику как запасов, так и потоков углерода ОВ с целью оценки функционирования лесных экосистем, разработки политики в области изменений климата, принятия мер по адаптации и смягчению их последствий.
На европейском Северо-Востоке России, где лесная растительность формирует основные природные ландшафты, доминируют еловые леса. Так, лесопокрытая площадь на территории Республики Коми (РК) составляет 30.2 млн га, из них 55% занимают еловые леса, эдификатором в которых является ель сибирская (Picea obovata L.). Они распространены во всех подзонах таежной зоны региона и произрастают в различных условиях, за исключением сухих песчаных почв и торфяников. Возрастная структура ельников РК на 80% представлена спелыми и перестойными древостоями (Лесное хозяйство…, 2000; Коренные еловые леса…, 2006). Известно, что скорость поглощения диоксида углерода из атмосферы еловыми фитоценозами зависит от климатических условий и количества доступного азота (Pregitzer, Euskirchen, 2004; Magnani et al., 2007). Однако данных, характеризующих пул углерода в КДО, потоки его при связывании в NPP, поступление с растительным опадом, атмосферными осадками и выделение в атмосферу при дыхании почвы для старовозрастных ельников недостаточно.
Цель данной работы – оценка запасов и потоков углерода ОВ в экосистемах двух типов старовозрастных ельников средней тайги.
ОБЪЕКТЫ И МЕТОДЫ
Анализ параметров углеродного цикла в экосистемах коренных среднетаежных ельников проведен на основе их количественных оценок, полученных при стационарных исследованиях и полевых экспериментах длительностью 15 лет (2002–2017 гг.). Работа выполнена на территории Ляльского лесоэкологического стационара Института биологии Коми научного центра УрО РАН (62°17′ с.ш., 50°40′ в.д.). Площадь стационара является частью Мезенско-Вычегодской равнины, поверхность которой покрыта четвертичными отложениями мощностью в десятки метров. Отложения включают два горизонта морены, верхний из которых был оставлен льдами Московского оледенения. Верхняя морена большей частью размыта и опесчанена, во многих местах перекрыта озерно-ледниковыми и флювиогляциальными отложениями. Озерно-ледниковые отложения мощностью 0.4–11 м залегают непосредственно под почвенно-растительным слоем. Они состоят из голубовато-серого суглинка, на поверхности коричневого, с прослойками и линзами супеси, песка, местами с галькой и гравием. Пески мелкие, глинистые, редко пылеватые, коричневые, коричневато-желтые, серые. Флювиогляциальные отложения представлены мелкими желтовато-коричневыми кварцевыми слабоглинистыми песками средней плотности. Мощность отложений 1.7 м (Атлас Коми…, 1964). Подземные воды не имеют сплошного развития, расположены спорадически и приурочены к песчаным линзам и прослойкам в толще суглинистых отложений. Глубина залегания подземных вод от 5 до 15.5 м. Мощность водоносных слоев от нескольких сантиметров до 3 м и более (Рубцов и др., 1990).
По агроклиматическому районированию стационар расположен на границе прохладного и умеренно-прохладного районов (Агроклиматические ресурсы…, 1973). Согласно данным наблюдений на метеостанции “Усть-Вымь”, расположенной в 15 км южнее от Ляльского стационара, средняя годовая температура воздуха составляет 0.1°С, средние минимальные и максимальные температуры соответственно равны –4.2 и 4.3°С. Средняя температура воздуха за вегетационный период составляет 11.6°С. Осадки в годовом цикле распределены неравномерно, основная часть их приходится на апрель–октябрь – 442 мм, в том числе 332 мм на вегетационный период. По почвенному районированию территория стационара относится к Нившеро-Вымскому округу Двино-Вычегодской провинции среднетаежной подзоны подзолистых почв. Почвообразующие породы представлены ледниковыми, водно-ледниковыми, в поймах рек – аллювиальными отложениями (Атлас Коми…, 1964).
Древесный ярус ельника разнотравно-черничного (Piceetum herboso-myrtillosum) состоит из ели сибирской, пихты сибирской Abies sibirica Ledeb., березы пушистой Betula pubescens Ehrh., березы повислой B. pendula Roth, сосны обыкновенной Pinus sylvestris L., осины обыкновенной Populus tremula L., имеет состав 7Е3Б + Пх, С ед. Ос (табл. 1). Древостой плотностью 891 экз. га–1 аккумулирует 311 м3 га–1 древесины. Сухостой 200 экз. га–1 представлен елью с запасом древесины 25 м3 га–1. Подрост 8.3 тыс. экз. га–1, в основном из ели. На поверхности почвы довольно много валежа (291 экз. га–1) из ели и березы. В подлеске встречаются единичные кусты можжевельника обыкновенного Juniperus cоmmunis L., шиповника иглистого Rosa acicularis Lindl., рябины обыкновенной Sorbus aucuparia L., ивы Salix sp. Травяно-кустарничковый ярус имеет простое синузиальное строение, состоит из типичных бореальных видов растений: черники Vaccinium myrtillus L., брусники V. vitis-idaea L., майника двулистного Maianthemum bifolium (L.) F.W. Schmidt., линнеи северной Linnaea borealis L., седмичника европейского Trientalis europaea L., голокучника обыкновенного Gymnocarpium dryopteris (L.) Newman и др. Почти сплошной моховой покров образуют Hylocomium splendens (Hedw.) Schimp., Pleurozium schreberi (Brid.) Mitt. с незначительной примесью Dicranum polysetum Sw., Polytrichum commune Hedw. и небольшими пятнами сфагновых мхов Sphagnum sp. Названия растений приведены по С.К. Черепанову (1995). Микрорельеф биогенный из заросших мхами валежа, старых пней. Почва текстурно-дифференцированная, подзолистая (Полевой определитель…, 2008; IUSS Working Group…, 2015). В почвенном профиле горизонт О (подстилочно-торфяной) имеет мощность 7.7 ± 0.4 см, под которым залегает подзолистый горизонт Е (7.7 ± ± 1.1 см), переходящий в горизонт BEL (субэлювиальный) – 13.0 ± 0.7 см, ВТ (текстурный) – 18.0 ± ± 0.7 см.
Таблица 1.
Лесоводственно-таксационная характеристика ельников
| Тип ельника | Состав древостоя | Вид древесного растения | Возраст деревьев, лет | Число деревьев, экз. га–1 | Запас древесины, м3 га–1 |
Среднее | Количество подроста, экз. га–1 | |||
|---|---|---|---|---|---|---|---|---|---|---|
| растущих | сухих | растущих | сухих | высота, м | диаметр, см | |||||
| Разнотравно-черничный | 7Е3Б + Пх, С ед Ос (2006) | Ель | 80–160 | 575 | 167 | 216 | 25.0 | 18 | 22 | 6838 |
| Береза | 60–100 | 258 | 8 | 68 | 0.02 | 18 | 20 | 1450 | ||
| Сосна | 110 | 17 | 8 | 12 | 0.2 | 22 | 29 | – | ||
| Пихта | 110 | 33 | 17 | 10 | 0.02 | 18 | 19 | – | ||
| Осина | 110 | 8 | – | 5 | – | 24 | 26 | – | ||
| Итого | 891 | 200 | 311 | 25.24 | 8288 | |||||
| Чернично-сфагновый | 9Е1Б + С ед Пх (2003) | Ель | 106–200 | 595 | 85 | 177 | 8.30 | 16 | 20 | 5179 |
| Береза | 110 | 15 | – | 8 | – | 23 | 32 | 1395 | ||
| Сосна | 110 | 5 | 10 | 8 | 1.19 | 22 | 40 | – | ||
| Пихта | 50 | 10 | – | 1 | – | 10 | 10 | 66 | ||
| Итого | 625 | 95 | 194 | 9.49 | 6640 | |||||
В ельнике чернично-сфагновом (Piceetum myrtilloso-sphagnosum) состав древостоя 9Е1Б + С ед. Пх. Всего на 1 га насчитывается 625 деревьев, аккумулирующие 194 м3 га–1 древесины. Сухостой – 95 экз. га–1 с запасом древесины 3.4 м3 га–1, валеж различной стадии гниения – 200 экз. га–1, в котором сосредоточено 55 м3 га–1 древесины. Подрост – 6.6 тыс. экз. га–1, из ели, пихты, березы (табл. 1). Подлесок редкий, состоит из шиповника, ивы, можжевельника, рябины. Травяно-кустарничковый ярус имеет проективное покрытие 60–70%, число видов 21: черника, брусника, линнея северная, хвощ (Equisetum sylvaticum L.), осока шаровидная (Carex globularis L.), костяника обыкновенная (Rubus saxatilis L.), костяника хмелелистная (R. humilifolius C.A. Mey), княженика (R. articus L.), майник, седмичник, кислица (Oxalis acetosella L.), голокучник, лютик (Ranunculus sp.), фиалка болотная (Viola palustris L.), мятлик луговой (Poa pratensis L.), бодяк полевой (Cirsium arvense (L.) Scop.), марьянник луговой (Melampyrum pretense L.), луговик извилистый (Avenella flexuosa (L.) Drey.), ожика волосистая (Luzula pilosa (L.) Willd.), золотая розга (Solidago virgaurea L.), дудник (Angelica sp.). Моховой покров с покрытием 80–90% образован сфагновыми мхами, на прикомлевых возвышениях встречаются небольшие пятна Polytrichum commune и Pleurozium schreberi, Hylocomium splendens, Rhytidiadelphus triquetrus. Почва характеризуется отчетливой дифференциацией на генетические горизонты. В ее профиле мощность горизонта О составляет 13.1 ± 2.3 см с четкой дифференциацией на подгоризонты по степени деструкции (сфагновый очес 0–5 см; O(F) 5–11 см; O(H) 11–13 см). Ниже формируется элювиальный горизонт (ELg) с отчетливо выраженными признаками оглеения в виде сизых и сизовато-бурых пятен. Тяжелый гранулометрический состав нижележащих горизонтов (BT1g–BT2g) затрудняет внутрипочвенный отток атмосферных осадков и определяет застойно-промывной тип водного режима. С 90 см сочится вода. Средняя и нижняя часть почвенного профиля также имеют признаки оглеения в виде сизых и ржавых морфонов, с редкими Mn–Fe конкрециями и примазками, что позволяет отнести данную почву к торфянисто-подзолисто глееватой (Полевой определитель…, 2008; Zhangurov et al., 2011).
Постоянные пробные площади (ППП) в ельниках заложены согласно ОСТ 56-69-83 (1983). На них проведен сплошной перечет деревьев древостоя, подроста, валежа. Обработка материала выполнена по “Лесотаксационному справочнику…” (1986). Возраст древостоя определяли с помощью кернов, взятых с 25 деревьев. Массу и продукцию надземных органов древесных растений определяли методом модельных деревьев (Уткин, 1975; Усольцев, 2007). Массу корней древесных растений оценивали по методу крупных и мелких монолитов (Орлов, 1967). Прирост корней определяли по формуле
Опад растений древесного яруса учитывали с помощью 18–20 опадоулавливателей (ОУ) размером 0.5 × 0.5 м. Растительные остатки собирали дважды в год – летом и поздней осенью. Массу растений травяно-кустарничкого яруса, мхов в ельнике чернично-сфагновом оценивали методом укосов на уровне поверхности подстилки на 10 площадках размером 0.5 × 0.5 м, в ельнике разнотравно-черничном – буром диаметром 10 см в 35-кратной повторности. Опад растений этого яруса определяли по их приросту, для чего проводили срезания побегов текущего года у 80–100 растений на тех же площадках. Ежегодно отмирающую массу брусники принимали равной 30%, черники, травянистых растений и мхов – 100% прироста (Родин и др., 1968). Прирост и опад корней растений травяно-кустарничкого яруса был принят равным ¼ от их массы (Dahlman, Kuceera, 1965; Бобкова и др., 1982). Массу подстилки определяли с помощью металлических шаблонов площадью 98 см2 в 30–35-кратной повторности. Каждый образец подстилки разбирали вручную на подгоризонты. Пересчет запасов массы растений, опада и подстилки на массу углерода осуществляли дифференцированно по компонентам с применением коэффициентов (0.45–0.53), свойственных отдельным фракциям (Бобкова, Тужилкина, 2001). Для оценки скорости разложения растительных остатков в органогенном горизонте на год закладывали в пятикратной повторности компоненты растительного опада и образцы лесной подстилки в капроновых мешочках (Heath et al., 1964).
Установка пробоотборников и сбор образцов осадков и лизиметрических вод проводили согласно общепринятым международным методикам (Manual on methods…, 2011). Атмосферные осадки учитывали ежемесячно в течение года. Дождевые осадки собирали с июня по октябрь. Для их сбора использовали осадкоприемники с диаметром приемной поверхности 18 см в 15-кратной повторности, которые размещались на расстоянии 5 м друг от друга под кронами елей, берез и в межкроновых пространствах (“окнах”). Снег отбирали с декабря по март. Для сбора твердых осадков использовали снегоприемники (S = 1017.4 см2), которые были установлены в трех–четырех повторностях под кронами елей, берез и в “окнах”. Образец из каждого снегоприемника взвешивали и растаивали, затем на химический анализ отбирали средний образец из-под крон елей, берез и “окон”. Воды, свободно стекающие по почвенному профилю, собирали в лизиметры с диаметром приемной поверхности 20 см, расположенные под каждым из горизонтов: О (подстилочно-торфяной) на глубине 7 см, Е (подзолистым) на глубине 15 см, BEL (субэлювиальный) на глубине 28 см в 3-кратной повторности. Содержание водорастворимого органического углерода в воде определяли в единицах массовых концентраций. Для более корректной интерпретации результаты эксперимента пересчитывали в соответствующие значения уровней поступления веществ в месяц на поверхность в кг га–1. Углерод в атмосферных и почвенных водах рассчитывали по данным, опубликованным ранее (Робакидзе и др., 2013, 2015).
Закладку почвенных разрезов, описание почвы, отбор образцов для анализа проводили в соответствии с рекомендациями (Агрохимические методы…, 1975). Физико-химические анализы растительных и почвенных образцов, проб воды проведены в аккредитованной экоаналитической лаборатории Института биологии Коми научного центра УрО РАН по метрологически аттестованным методикам количественного химического анализа (№ РОСС RU.0001.511257). Содержание органического углерода в образцах подстилки и почвы определяли методом газовой хроматографии на анализаторе элементном ЕА 1110 (CHNS–O) (фирма CE Instruments, Италия).
Запас органического углерода в почвах еловых фитоценозов рассчитывали по формуле А.В. Смагина и соавт. (2001):
где С% – процентное содержание органического вещества относительно массы сухой почвы, ρb – плотность почвы (г см–3), h – мощность слоя почвы, в котором сосредоточен запас углерода (см).При измерении эмиссии СО2 с поверхности почвы использовали две разновидности метода камер: закрытая статическая (накопительная) с использованием инфракрасного газового анализатора ПГА-6 и открытая динамическая (принудительной продувки) с применением анализатора LI-COR 8100 (LI-COR Biosciences, США) с почвенной камерой Survey chamber 8100-103 диаметром 20 см. Учет факторов, влияющих на эмиссию СО2 из почвы, как изменение давления внутри измерительной камеры при ее установке, способ перемешивания анализируемой воздушной смеси, рост концентрации СО2 в камере и эффект растворения газа в водном паре, позволяет считать ее эталонной. Измерение эмиссии на ИКГ LI-COR проведены по методику Хили с соавт. (Healy et al., 1996). Для изучения почвенного дыхания камерным методом предварительно срезали зеленую часть растений напочвенного покрова и проводили измерения с поверхности подстилки. Температуру почвы измеряли при помощи автономных термисторов Logger Hobo (США), установленных на глубину 5, 10, 15 см. Измерения вели в непрерывном почасовом режиме в течение всего вегетационного периода. В сроки измерений определяли влажность почвы весовым методом (Роде, 1960). Расчет эмиссии углекислого газа методом динамической продувки осуществляли при помощи программ LI-8100(M) PC Client v.2.0.0 и File Viewer v.2.0.0. Для оценки выделения СО2 из древесины валежа различной стадии гниения использовали образцы КДО: ель от 1-й до 5-й, сосна 2-й и береза 2-й стадии разложения. Делали спилы стволовой части валежа и исследовали эмиссию углекислого газа методом темных камер на газовом анализаторе Li-COR 8100 в трехкратной повторности. Стадии гниения древесины определяли по В.Г. Стороженко (1990). Статистический анализ материалов выполнен по И.И. Гусеву (2002).
РЕЗУЛЬТАТЫ
Пул углерода в древостое. Данные о запасах и потоках ОВ в исследуемых старовозрастных ельниках нами приведены ранее (Кузнецов, Бобкова, 2014; Бобкова и др., 2020). В ельниках разнотравно-черничном аккумулировано 219.8, чернично-сфагновом – 216.28 т га–1 растительного ОВ. В данной работе рассмотрим пул и потоки углерода ОВ в этих сообществах. Так, в ельнике разнотравно-черничном в массе растущих деревьев сосредоточено 90.04 т С га–1 (табл. 2). Его формируют в основном ель (74.3%) и береза (18.2%). Участие сосны, пихты, осины составляет 3.5, 2.9, 1.1% соответственно. В фитомассе древостоя ельника чернично-сфагнового аккумулировано 85.48 т С га–1. В его содержании доминирует ель (95.1%). Небольшое количество углерода концентрируют береза (2.9%), сосна (1.8%) и пихта (0.2%). Полученные данные по запасам углерода в древостоях ельников вписываются в пределы колебаний, приведенных в литературе. Так, спелые и перестойные древостои среднетаежных ельников Европейско-Уральской провинции в фитомассе аккумулируют в среднем 57.2 т С га–1 (Уткин и др., 2004). В условиях РК в массе растущих органов древостоев среднетаежных ельников черничных концентрируется 84.9–89.7 т С га–1 (Коренные еловые леса…, 2006). Н.Н. Кошурникова и С.В. Верховец (2011) выявили, что в фитомассе 170-летнего древостоя ельника разнотравно-осочкового Западной Сибири содержится 107.5 т С га–1.
Таблица 2.
Запас, продукция, опад углерода органического вещества (среднее за три года) в фитоценозах ельников
| Состав ОВ | Разнотравно-черничный | Чернично-сфагновый | ||||
|---|---|---|---|---|---|---|
| фитомасса, т га–1 | продукция, т га–1 год–1 |
опад, т га–1 год–1 |
фитомасса, т га–1 |
продукция, т га–1 год–1 |
опад, т га–1 год–1 |
|
| Древостой | 90.04 | 2.77 | 2.23 | 85.48 | 2.49 | 2.05 |
| Подрост + подлесок | 0.66 | 0.05 | 0.002 | 0.69 | 0.07 | 0.003 |
| Напочвенный покров | 1.67 | 0.53 | 0.61 | 1.95 | 0.52 | 0.50 |
| КДО | 10.44 | – | – | 15.7 | – | – |
Распределение углерода фитомассы по компонентам в древостоях исследуемых коренных ельников довольно близкое. На долю стволовой древесины приходится 54.8–59.9%, стволовой коры – 6.2–6.5%, ветвей – 7.5–7.7%, листьев (хвои) – 5.7–7.7%, корней – 20.7–23.0% от общего содержания углерода в древостоях. А.И. Уткин с соавт. (2004) приводили сопоставимые с нашими показатели соотношения отдельных компонентов для ельников Европейско-Уральской провинции. По их данным, от углерода фитомассы древостоя на стволовую древесину приходится 60.1%, ветвей – 11.3%, листьев (хвои) – 7.1%, корней – 21.3%.
Углерод КДО. В хвойных экосистемах, особенно в спелых и перестойных, в бюджете углерода существенную роль выполняют КДО (Замолодчиков, 2009; Pan et al., 2011). Запас последнего складывается из сухостоя, отмерших, но прикрепленных к стволу ветвей, корней, а также древесины остолопа (дерева со сломанной вершиной) и валежа различной стадии гниения. В ельнике разнотравно-черничном запасы углерода КДО равны 10.44 т га–1, из них 37.3% сосредоточено в древесине сухостойных деревьев, 22.5% – в сухих ветвях, 21.6% – в древесине ствола валежа, представленного елью и березой, 12.3% – в корнях сухостойных деревьев, 6.3% – в древесине остолопа. В КДО ельника чернично-сфагнового аккумулировано 15.72 т С га–1, половина которого приходится на валеж. Стволовая древесина, ветви и корни сухостойных деревьев содержат соответственно 29.4, 7.4 и 10.2% углерода, накопленного в КДО. Вклад остолопа в запасе углерода КДО невелик (3%). В валеже чернично-сфагнового ельника, состоящего из ели, сосредоточено 8.23 т С га–1, из них 19.7% приходится на древесину первой, 40.2% – второй, 29.0% – третьей и 11.0% – четвертой стадии гниения.
Углерод растений нижних ярусов. Подрост ельника разнотравно-черничного (8.3 тыс. экз. га–1) представлен преимущественно здоровой елью мелкой и средней категорий крупности. Фитомасса деревьев этого яруса содержит 0.26 т С га–1. Вклад растений подлеска в накоплении углерода – 0.40 т га–1. В фитомассе растений напочвенного покрова аккумулировано 1.67 т С га–1, из них в кустарничках – 10.2%, травяных растениях – 4.7%, мхах – 34.7%, корнях кустарничков и трав – 50.4%.
В ельнике чернично-сфагновом в древесных растениях подроста запасается 0.67 и подлеска – 0.02 т С га–1. Растения напочвенного покрова в этом ельнике содержат 1.95 т С га–1. Большую часть запаса углерода формируют мхи, преимущественно сфагновые, на долю которых приходится 24.8%. Углерод наземной фитомассы кустарничков и трав занимает 7.5 и 7.2% соответственно. 60.5% от запасов углерода растений напочвенного покрова сосредоточено в корнях кустарничков и трав.
Углерод почв находится в состоянии динамического равновесия. С одной стороны, его количество пополняется за счет притока растительных остатков опада и углерода атмосферных осадков, с другой – убывает, подвергаясь окислению, микробному разложению и выносу за пределы почвенной толщи (Смагин и др., 2001; Аккумуляция углерода…, 2018). Исследуемые ельники являются коренными типами сообществ, не испытавших заметного антропогенного воздействия. Как было отмечено, ельник разнотравно-черничный развивается на типичных подзолистных, а чернично-сфагновый – на торфянисто-подзолисто-глееватых почвах. Следует отметить, что почвы близки по гранулометрическому составу (супесчаные на суглинках) и химическим свойствам (кислые, бедные поглощенными основаниями). Почвы отличаются по их водному режиму. Подзолистая почва ельника разнотравно-черничного характеризуется относительно благоприятными условиями влажности. Влагозапас теплого периода в ней держится основном в доступной форме. Почва умеренно дренированная, отмечается периодическое увлажнение в весенний, редко в летний период. Ельник чернично-сфагновый входит в динамический ряд заболачивания ельников черничных. Торфянисто-подзолисто-глееватая почва этого сообщества в весеннее-летнее время почти постоянно переувлажнена (Коренные еловые леса…, 2006).
При оценке пула органического углерода в почвах среднетаежных ельников РК выявлена его значительная вариабельность: от 57 т га–1 в типичной подзолистой почве до 151 т га–1 в глееподзолистой почве (Продуктивность…, 1975; Осипов, Кузнецов, 2010; Бобкова и др., 2014). Такой широкий диапазон запасов органического углерода является одной из характерных черт накопления ОВ в почвах еловых сообществ средней тайги, отображающих разнообразие лесорастительных условий при слабой степени гумификации растительных остатков.
Согласно результатам наших исследований, пул углерода в метровом слое почвы, включая лесную подстилку, в подзолистой почве ельника разнотравно-черничного составляют 84.7 ± 8.3 т га–1, а в торфянисто-подзолисто-глееватой ельника чернично-сфагнового – 81.3 ± 8.3 т га–1 (табл. 3). Для почв рассматриваемых еловых экосистем характерно накопление ОВ на поверхности почвы в виде лесной подстилки. Органогенный слой почвы ельника разнотравно-черничного мощностью 7.7 ± 0.4 см аккумулирует 15.9 ± 1.5 т С га–1, а ельника чернично-сфагнового мощностью 13.1 ± 2.3 см – 26.6 ± 0.3 т С га–1. Следует отметить, что в слое почвы (0–50 см) исследуемых ельников концентрируется 68–73% углерода ОВ от общего его количества в метровом слое.
Таблица 3.
Запасы органического углерода в почвах ельников
| Запас углерода, т га–1 | Разнотравно-черничный | Чернично-сфагновый |
|---|---|---|
| В слое 0–20 см | 28.5 ± 3.4 | 10.8 ± 1.6 |
| В слое 0–50 см | 47.1 ± 5.2 | 32.9 ± 8.2 |
| В слое 0–100 см | 68.8 ± 8.2 | 54.7 ± 13.2 |
| Лесная подстилка | 15.9 ± 1.5 | 26.6 ± 2.5 |
| В слое 0–100 см, в том числе лесной подстилки | 84.7 ± 8.3 | 81.3 ± 8.3 |
Таким образом, анализ аккумуляции углерода в естественно развивающихся экосистемах коренных среднетаежных ельников разнотравно-черничного на подзолистых и чернично-сфагнового на торфяно-подзолисто-глеевых почвах показал, что общий пул углерода в этих экосистемах составляет 184.4–187.9 т га–1, из них 47.8–49.1% сконцентрировано в живой фитомассе, 5.6–8.1% – в КДО, 44.1–45.3% – в метровом слое почвы, включая лесную подстилку (рис. 1).
Рис. 1.
Запасы углерода ОВ в экосистемах ельников: разнотравно-черничного (а), чернично-сфагнового (б). Углерод: 1 – фитомассы, 2 – КДО, 3 – почвы, включая подстилку. Общий запас углерода в экосистемах приведен в рамке.

Углерод в NPP (Net Primary Production). Потоки углерода ОВ в экосистемах рассматриваемых ельников представлены на рис. 2. Интенсивность продуцирования органического вещества, во многом определяющая ход процессов развития фитоценозов в экосистемах, используется в целях оценки углероддепонирующей способности лесов. Годичная продукция углерода древостоя в ельнике разнотравно-черничном составляет 2.77 т га–1, из них ель накапливает 74.6%, сосна – 3.0%, береза – 18.0%, пихта – 3.2% и осина – 1.2%. Вклад в продукцию углерода растений подроста и подлеска составляет 0.05 т га–1. Растения напочвенного покрова за год накапливают 0.53 т С га–1. В депонировании углерода в растениях данного яруса основную роль выполняют корни кустарничков, трав (44.4%) и мхи (30.5%). Участие в годичной продукции углерода наземных органов кустарничков и трав составляет 9.1 и 16.0% соответственно. В целом, NPP фитоценоза ельника разнотравно-черничного достигает 3.30 ± 0.56 т С га–1 год–1 (табл. 2).
Рис. 2.
Запасы (т С га–1) и потоки (т С га–1 год–1) углерода в ельниках: разнотравно-черничном (а), чернично-сфагновом (б).
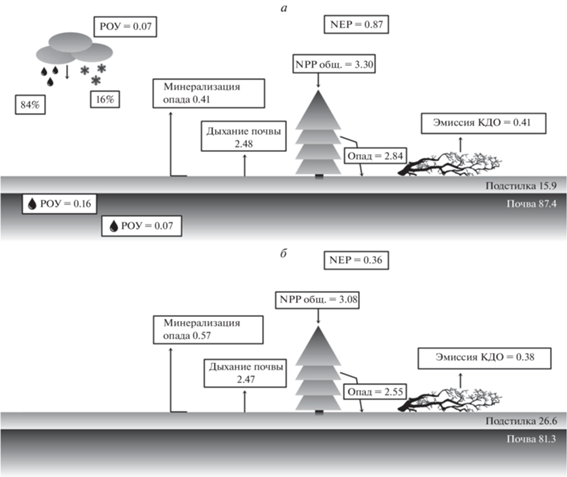
Годичная продукция углерода фитомассы древесных растений ельника чернично-сфагнового на полугидроморфных почвах равна 2.56 т га–1 в год. Существенный вклад в накоплении углерода этого яруса вносят листья (хвоя) 34.3% и корни 31.0%. Прирост углерода ОВ стволовой древесины в исследуемом ельнике составляет 17.7%, ветвей – 12.3%, коры – 4.7% от прироста фитомассы древостоя. В продукции растений травяно-кустарничкого яруса данного ельника накапливается 0.52 т С га–1 год–1. Вклад кустарничков и мхов составляет по 26.9%, трав – 13.5%, корней кустарничков и трав – 32.7%. Таким образом, в NPP фитоценоза ельника чернично-сфагнового накапливается 3.08 ± 0.29 т С га–1 год–1 (табл. 2).
Лесной опад обеспечивает переход углерода из фитомассы в почву. Он состоит из опавших растительных остатков древесных растений, растений напочвенного покрова и корнепада. С растительными остатками в ельнике разнотравно-черничном поступает 2.84 ± 0.11, а в чернично-сфагновом – 2.55 ± 0.17 т С га–1 год–1 (табл. 2). С наземным древесным опадом в ельники разнотравно-черничный и чернично-сфагновый поступает 1.59 ± 0.12 и 1.26 ± 0.13 т С га–1 год–1 соответственно. В обоих рассматриваемых фитоценозах доминируют растительные остатки ели (71–79%) и березы (12–24%). В ельнике разнотравно-черничном участие корней древесных растений в опаде составляет 24.5%, а в ельнике чернично-сфагновом – 31.0% от углерода всего опада. Доля участия растений напочвенного покрова в структуре массы опада составляет 19.6–21.5%. Масса углерода ОВ опада в рассматриваемых ельниках составляет 82–86% показателей его ежегодного прироста.
Поступивший опад в результате действия почвенных беспозвоночных, микроорганизмов и грибов разлагается. В результате часть ОВ опада превращается в гумус, другая часть окисляется до СО2 и воды с выделением энергии. Результаты изучения почвенной микрофауны в исследуемых и близких к ним по структуре насаждениях исследуемого региона (Смоленцева, 1979; Мелехина, 2001) показали, что почвенные беспозвоночные распространены в основном в органогенном слое почвы. Среди изученных групп мелких членистоногих преобладали сапрофаги (клещи и ногохвостки) – около 90%. Почвенное население более крупных размеров в изучаемых ельниках (мезофауна – 1.28–10 мм) представлено всеми основными группами постоянных обитателей таежных почв: дождевыми червями, многоножками, наземными моллюсками, жужелицами и стафилинидами. Согласно Ф.М. Хабибуллиной (2009), в старовозрастных ельниках рассматриваемого региона в микробной биомассе почв доминируют грибы. На их долю приходится более 99% общей биомассы, из которых 1–6% составляет биомасса спор, а более 94% – мицелия. В подзолистой почве ельника насчитывается 58, а в болотно-подзолистой – 34 вида микромицетов. В подзолистой почве преобладали представители родов Penicillium, Mucor, Mortierella. Целлюлозоразрушающие грибы представлены 16 видами. Доминируют в этой группе Trichoderma и Chaetomium. Межсезонные колебания численности грибов наблюдаются по всему профилю подзолистой почвы, но их максимум регистрируется в гумусовом горизонте и подстилке. Согласно исследованиям этого автора, в торфянисто-подзолисто-глееватой почве по численности и видовому разнообразию преобладают пенициллы (11 видов), единичны аспергиллы, темноцветные гифомицеты, многочисленны виды р. Trichoderma (6 видов), светло- и темноокрашенные формы стерильного мицелия, а также мицелий базидиальных грибов с пряжками. К доминирующим видам грибов в торфянисто-подзолисто-глееватых почвах относятся Trichoderma sympodianum, T. viride, Penicillium camemberti, Mortierella ramanniana, а также мицелий базидиальных грибов. Глубина распространения микроорганизмов вниз по профилю небольшая и составляет 20–30 см.
Исследования разложения опада, проведенные в еловом сообществе средней тайги РК, показали, что размер годичной деструкции древесного опада составляет 32.9% (Продуктивность…, 1975), в сосново-еловом – 30% (Смоленцева, 1979). Согласно К.С. Бобковой с соавт. (1989), скорость разложения надземной части опада хвойных насаждений средней тайги в первый год составляет от 15 до 30%. Показано, что в результате таких малых темпов деструкционных процессов происходит образование грубогумусовой подстилки мощностью от 5 до 20 см. Так, подстилка ельника разнотравно-черничного мощностью 7.7 ± 0.4 см аккумулирует 15.9 ± 1.5 т С га–1, а ельника чернично-сфагнового мощностью 13.1 ± 2.3 см – 26.6 ± 0.3 т С га–1. В ранее проведенных исследованиях по разложению подстилки в ельниках средней тайги выявлено, что за год минерализуется от 7 до 15.6% от исходной массы (Фролова, 1965; Продуктивность…, 1975; Бобкова и др., 2014).
Деструкция растительных остатков в рассматриваемых ельниках исследовалась на компонентах опада, играющих значимую роль в его общей массе. Интенсивность разложения отдельных компонентов растительных остатков опада за год в ельниках колеблется от 2.5 до 63%. По убыванию скорости разложения фракции опада располагаются следующим образом: черника > травяные растения > листья березы > брусника > мхи > > хвоя сосны > хвоя ели > ветви ели > шишки ели и сосны > кора ели (табл. 4).
Таблица 4.
Потеря массы основных компонентов растительных остатков опада и подстилки при деструкции (средние за три года), %
| Компонент опада | Потеря массы | |
|---|---|---|
| ельник разнотравно-черничный | ельник чернично-сфагновый | |
| Древесные растения | ||
| Ель | ||
| Хвоя | 22.0* ± 0.9** | 19.2 ± 3.6 |
| Кора | 2.5 ± 0.1 | 2.6 ± 0.6 |
| Ветви | 9.1 ± 1.0 | 9.2 ± 1.4 |
| Шишки | 3.9 ± 0.7 | 3.9 ± 1.0 |
| Береза | ||
| Листья | 35.7 ± 7.1 | 30.8 ± 6.6 |
| Ветви | 5.6 ± 0.8 | – |
| Сосна | ||
| Хвоя | 26.0 ± 1.3 | 25.2 ± 2.0 |
| Кора | 6.9 ± 2.4 | – |
| Шишки | 6.0 ± 3.3 | – |
| Осина | ||
| Листья | 24.2 ± 1.1 | 50.1 ± 13.5 |
| Ветви | 5.6 ± 0.8 | – |
| Растения напочвенного покрова | ||
| Черника | 51.0 ± 4.1 | 63.0 ± 16.3 |
| Брусника | 29.8 ± 1.4 | 28.9 ± 5.9 |
| Разнотравье | 56.0 ± 3.8 | 48.1 ± 8.7 |
| Мхи зеленые | 28.0 ± 3.1 | 30.8 ± 4.0 |
| Мхи политриховые | 26.0 ± 2.6 | – |
| Мхи сфагновые | – | 32.2 ± 1.2 |
| Лесная подстилка | 15.6 ± 3.0 | 14.4 ± 2.2 |
| Подгоризонт L | – | 4.6 ± 1.2 |
| Подгоризонт F | – | 9.6 ± 2.5 |
| Подгоризонт H | – | 0.2 ± 0.05 |
Растворенный органический углерод (РОУ). В углеродном бюджете лесных сообществ важную роль выполняет РОУ (Никонов, Лукина, 1994; Лесные экосистемы…, 2002; Горбачева, Лукина, 2004, и др.). Он является главным пищевым субстратом для микробных сообществ, определяет реакцию почвенных вод, контролирует доступность питательных элементов для растений. Состав РОУ лизиметрических вод отражает биологическую продуктивность ценозов, интенсивность разложения, физико-химическую адсорбцию и транспортные процессы в почве. Нами ранее (Робакидзе и др., 2013, 2015) была проведена оценка миграционных потоков РОУ с осадками в трех типах среднетаежных еловых экосистем на подзолистых почвах. Показано, что в составе атмосферных осадков, поступающих под полог ельников, содержание РОУ подвержено значительным колебаниям. На основе экспериментальных исследований определено количество поступающего РОУ в исследуемом ельнике разнотравно-черничном. Поступление РОУ со снегом под полог древостоя варьирует по месяцам зимнего сезона от 1.3 до 8.0 кг га–1. Более высокие выпадения его отмечены в декабре (рис. 3). В феврале происходит снижение в снеговой воде, а в марте увеличение содержания РОУ. Таким образом, за зимний период в ельнике разнотравно-черничном в разные годы выпало от 9.4 до 15.2 кг га–1 РОУ. В составе дождевых осадков содержание данного компонента в зависимости от периода вегетации под пологом древостоя ельника варьирует от 3 до 28 кг га–1. Более высокие показания его отмечены в июне и июле. В августе наблюдали снижение содержания РОУ в дождевой воде в исследуемом ельнике в среднем в 2.3 раза по сравнению с июлем. В октябре его концентрация увеличивается. За вегетационный период под полог ельника поступает 58.1–64.8 кг га–1 РОУ. Итого с осадками под полог ельника в разные годы поступает от 69.2 до 79.9 кг га–1 РОУ, из них 94% в подкроновое пространство. Содержание РОУ в лизиметрических водах снижается в процессе продвижения вниз по почвенному профилю. В течение вегетации концентрации РОУ в почвенных водах под разными горизонтами варьируют, так же как и в дождевых осадках, поступающих к поверхности почвы ельника. Более высокое содержание РОУ в почвенных водах отмечается в мае–июне, в июле резко снижается. В сентябре наблюдали повышенное содержание РОУ, а в октябре оно снова падает. Сентябрьское увеличение концентрации РОУ, видимо, связано с сезонным накоплением на поверхности почвы опада, который является источником ОВ. В сентябре колебания температуры способствуют разложению опада и вымыванию из него углерода ОВ в нижележащие горизонты, тогда как к октябрю ОВ в почве закрепляется в нерастворимом виде. За вегетационный сезон (май–октябрь) из подстилочно-торфяного горизонта ельника разнотравно-черничного вымывается в среднем 156.9 кг га–1, из подзолистого – 95.4 кг га–1, из субэлювиального – 68.6 кг га–1 РОУ. Для сравнения, в северотаежном ельнике зеленомошном Мурманской области вынос углерода из лесной подстилки составляет 95 кг га–1, из минерального профиля – 35 кг га–1 (Аккумуляция углерода…., 2018). Анализ потоков РОУ в осадках и лизиметрических водах ельника в системе атмосфера–древостой–почва свидетельствует о том, что атмосферные выпадения существенно обогащаются органическим углеродом.
Рис. 3.
Поступление РОУ с осадками (а) и вынос с лизиметрическими водами (б) в ельнике разнотравно-черничном (среднее за три года). 1 – подстилочно-торфяной, 2 – подзолистый, 3 – субэлювиальный горизонты почвы.

Эмиссия СО2 с поверхности почвы служит важнейшей составляющей общего поступления парниковых газов в атмосферу. На основе регистрации потока диоксида углерода с поверхности почв исследуемых ельников выявлены суточные и сезонные изменения его показателей. В течение всего вегетационного периода отмечается понижение интенсивности СО2-газообмена почвы в утренние и дневные, повышение в вечерние и ночные часы. Количество выделяющегося СО2 с поверхности подзолистой почвы ельника разнотравно-черничного в середине мая, после схода снега составляло от 0.03 до 0.06 г С м–2 ч–1, в июле–августе постепенно увеличивалось от 0.29 до 0.44 г С м–2 ч–1 и постепенно снижалось к осени, составляя в конце октября 0.01–0.03 г С м–2 ч–1. С поверхности почвы ельника разнотравно-черничного выделяется в разные годы от 2.26 до 2.69, в среднем 2.48 т С га–1 сезон–1 (Машика, 2006). Согласно результатам наших наблюдений (Кузнецов, 2014), в конце мая эмиссия диоксида углерода с поверхности полугидроморфной почвы ельника чернично-сфагнового составляла 0.0003–0.01 г С м–2 ч–1, более высокого значения достигала в июле 0.11–0.15, и в октябре снижалась до 0.02 г С м–2 ч–1. Средняя удельная скорость выделения СО2 с поверхности почвы ельника чернично-сфагнового с мая по октябрь в зависимости от условий вегетационного сезона составила от 2.44 до 2.50, в среднем 2.47 т С га–1 сезон–1.
В исследуемых ельниках выявлена положительная корреляционная взаимосвязь (r = 0.34–0.95, при р < 0.05) скорости эмиссии СО2 с температурой верхних горизонтов почвы. Влияние влажности почвы на выделение диоксида углерода проследить довольно сложно из-за небольших ее колебаний. При средних значениях влажности и среднесуточной эмиссии СО2 линейный корреляционный анализ показал отрицательную связь (r = –0.44–0.86, при р < 0.05) для ельника разнотравно-черничного (Машика, 2006) и недостоверную (r = 0.13–0.36, при р > 0.05) для ельника чернично-сфагнового (Кузнецов, 2014).
В ельнике чернично-сфагновом были проведены наблюдения эмиссии СО2 с поверхности почвы в зимний период – с января по апрель 2009 г. Выделение диоксида углерода с января по март незначительное, в среднем 0.003 г С м–2 ч–1. Близкие к полученным данным показатели эмиссии СО2 с поверхности почвы в зимний период приведены для хвойных лесов Западной Сибири (Чебакова и др., 2002) и Канады (Mahli et al., 1999). В апреле выделение диоксида углерода из почвы среднетаежного ельника значительно увеличилось и равнялось 0.02 г С м–2 ч–1. Увеличение эмиссии СО2 в начале весны Н.М. Чебакова с соавт. (2002) объясняют усилением стволового дыхания, обусловленным, в свою очередь, увеличением теплообмена в почве в весенний период. Согласно нашим наблюдениям, поток диоксида углерода с января по апрель в ельнике чернично-сфагновом составил 0.23 т С га–1 или 8.5% от эмиссии СО2 с поверхности почвы за вегетационный период.
При изучении цикла углерода в лесных экосистемах следует учитывать роль потоков углерода из КДО. Известно, что процессы биологического разложения древесины протекают при участии широкого круга организмов, образующих ксилофильные сообщества, где ведущая роль принадлежит дереворазрушающим грибам. Показано, что скорость разложения древесины в естественных условиях зависит от многих факторов, среди которых наиболее важным является влажность воздуха приземного слоя и почвы в период вегетации. Эти показатели, в свою очередь, определяют влажность древесины КДО, рост мицелия грибов, их видоспецифичность. В скорости разложения значима также роль линейных параметров стволов, положения их относительно земли, характеристики экотопа и другие (Krankina, Harmon, 1995; Шорохова, Шорохов, 1999; Mukhin et al., 2021). Показано также, что выделение СО2 с древесины валежа определяется стадией ее гниения (Сафонов и др., 2012; Иванов и др., 2018; Гитарский и др., 2020). Согласно нашим исследованиям, у древесины ели первой стадии гниения эмиссия СО2 составила 0.01 г С м–2 ч–1. Затем она постепенно возрастала по мере увеличения деструкции и достигала 0.38 г С м–2 ч–1 у древесины пятой стадии гниения. Измерения показали, что эмиссия углерода из древесины сухостойных деревьев и валежа составила 0.41 ± 0.11 и 0.38 ± 0.07 т га–1 год–1 для ельника разнотравно-черничного и чернично сфагнового соответственно.
ОБСУЖДЕНИЕ
В исследуемых среднетаежных старовозрастных ельниках эдификатором экосистем является разновозрастный еловый древостой, представленный деревьями четырех–пяти классов возраста. В фитоценозах выражен успешный возобновительный процесс. Подрост 6–8 тыс. экз. га–1 состоит в основном из ели, здоровый. Следовательно, в древостоях ельников происходит непрерывная смена поколений. В ельнике разнотравно-черничном на подзолистой почве и в ельнике чернично-сфагновом на торфянисто-подзолисто-глееватой почве пул углерода в экосистеме довольно близок и составляет 187.68 и 184.40 т га–1 соответственно. Из его запасов 47.8–49.1% аккумулировано в растительном ОВ, включающем древостой, подрост, подлесок и растения напочвенного покрова. Углерод ОВ почвы занимает 44.1–45.3%, КДО – 5.6–8.1% от его общего пула. Небольшие различия аккумуляции углерода ОВ в ельниках объясняется тем, что исследуемые еловые сообщества произрастают на близких по гранулометрическому составу и химическим свойствам почвах. Почвы супесчаные, подстилаемые суглинками, обладают высокой кислотностью. В минеральных горизонтах почвы с глубиной резко падает содержание гумуса и азота (Коренные еловые леса…, 2006). Продукция ОВ ельника разнотравно-черничного составляет 3.3 т С га–1 год–1, ельника чернично-сфагнового – 3.08 т С га–1 год–1. За год с опадом в почву старовозрастных ельников поступает 2.84 и 2.55 т С га–1 соответственно. Эмиссия диоксида углерода с поверхности почв ельников в разные годы составляет 2.3–2.7 т га–1 сезон–1. Вклад углерода КДО в потоке углекислого газа составляет 0.41 для ельника разнотравно-черничного и 0.38 т га–1 год–1 для ельника чернично-сфагнового.
Оценка бюджета углерода в экосистеме проведена на основании потоков углерода, т.е. по разности между поступлением и потерями углерода по уравнению (Кудеяров и др., 2007)
Чистая экосистемная продукция (Net Ecosystem Production, NEP) = NPP – Дыхание гетеротрофов
Дыхание гетеротрофных организмов является одним из ключевых звеньев при оценке углеродного цикла как в почве, так и в экосистеме. Оно определяется по разности между эмиссией СО2 с поверхности почвы и дыханием корней. В исследованиях ряда авторов (Bond-Lamberty et al., 2004; Кудеяров и др., 2007; Евдокимов и др., 2010, и др.) показано, что дыхание корней древостоев занимает от 7 до 36% от общей эмиссии углерода из почвы. В наших расчетах доля дыхания корней была принята 27% от общего дыхания с поверхности почвы. Данный показатель является средним согласно вышеприведенным работам.
Так, в ельнике разнотравно-черничном бюджет углерода (т С га–1 год–1) в экосистеме равен 3.30 – 2.43 = 0.87 (с учетом выноса РОУ из почвы 0.07 он составит 0.80), а в чернично-сфагновом 3.08 – 2.72 = 0.36. Для сравнения, старовозрастные еловые экосистемы черничных типов в условиях РК служат местом стока для 0.2–0.3 в северной и 0.3–0.4 т С га–1 год–1 в средней подзоне тайги (Коренные еловые леса…, 2006).
Спелые и перестойные ельники рассматриваются преимущественно как сток углерода и реже как углерод-нейтральные экосистемы (Valentini et al., 2000; Luyssaert et al., 2008; Ueyama et al., 2014, и др.) Количественная характеристика пулов и потоков углерода, а также величина поглощения или выделения углерода в лесной экосистеме зависит от возраста древостоя (Law et al., 2003). В работе Крииска с соавт. (Kriiska et al., 2019) показано, что NEP хвойных экосистем снижается с увеличением возраста древостоя, смещаясь в сторону умеренного источника углерода в насаждениях возрастом более 100 лет. Спелые и перестойные лесные фитоценозы, приближаясь к устойчивому равновесному состоянию (стадии климакса), в отдельные периоды могут стать источником углерода вследствие снижения их продуктивности, накоплению больших запасов почвенного углерода и отпада деревьев (Бобкова, 2007; Luyssaert et al., 2008; Goulden et al., 2011). Приведенный в данной статье материал показывает, что рассматриваемые среднетаежные коренные ельники являются стоком углерода. Несмотря на то, что общие запасы углерода фитомассы и почвы исследуемых экосистем близки, показатели NPP и NEP ельника разнотравно-черничного на подзолистых почвах несколько выше, чем ельника чернично-сфагнового на торфянисто-подзолисто-глееватых. Это, видимо, следует объяснить прежде всего неблагоприятными условиями влажности полугидроморфной почвы ельника чернично-сфагнового, для которой характерно постоянное переувлажнение (Коренные еловые леса…, 2006). Избыток влаги в почве способствует снижению скорости деструкции растительных остатков и аккумуляции углерода в лесной подстилке, почве и КДО. Запасы углерода мертвого органического вещества (КДО + лесная подстилка) в исследуемых нами лесных сообществах значительны. В ельнике разнотравно-черничном они равны 25.54, в ельнике чернично-сфагновом 41.97 т га–1, что составляет 13.6 и 22.0% соответственно от общих запасов углерода в экосистемах. Следует отметить, что накапливая мортмассу, старовозрастные ельники не обязательно становятся источником углерода. Они сохраняют аккумулирующие функции. Медленно разлагаясь и накапливаясь на поверхности и в толще почвы, фитодетрит исполняет роль хранителя углерода в экосистемах и субстрата для появления новых поколений древесных растений. Как показали исследования, экосистема коренного среднетаежного ельника чернично-сфагнового на полугидроморфных почвах является слабым, но все-таки стоком углерода, в то время как сфагново-черничный ельник на торфянисто-подзолисто-глееватой почве в условиях южной тайги – его источником. Согласно А.В. Варлагину с соавт. (2012), по результатам 12-летних наблюдений в южнотаежном ельнике величина NEР варьирует от –55 до –318 г С м–2 год–1. В то же время старовозрастный сложный неморальный ельник подзоны южной тайги характеризуется положительным (NEE = = 166 г С м–2 год–1) углеродным балансом (Ольчев и др., 2009).
ЗАКЛЮЧЕНИЕ
В целом анализ входного (расход атмосферного углерода на формирование продукции) и выходного (возврат С-СО2 в атмосферу) потоков углерода показывает, что старовозрастные ельники разнотравно-черничный на подзолистых почвах и чернично-сфагновый на торфянисто-подзолисто-глееватых служат местом стока углерода. NEP равен 0.87 и 0.36 т С га–1 год–1 соответственно, что находится в соответствии с приведенными оценками (0.66 т С га–1 год–1) для покрытых лесом земель средней тайги России (Швиденко, Щепаченко, 2014), выше оценок (0.26 т С га–1 год–1) для бореальных лесов Сибири (Ведрова, Ваганов, 2009). Вызывают сомнение данные С.В. Загировой с соавт. (2020), характеризующие положительный баланс углерода (NEE) 327 г С м–2 в течение вегетационного периода (апрель–август) для среднетаежного перестойного ельника черничного на подзолистых почвах. Приведенный в статье материал может быть использован при оценке роли старовозрастных хвойных лесов в углеродном балансе биосферы Северных регионов таежной зоны.
Список литературы
Агроклиматические ресурсы Коми АССР, 1973. Л.: Гидрометеоиздат. 134 с.
Агрохимические методы исследования почв, 1975 / Отв. ред. Соколов А.В. М.: Наука. 656 с.
Аккумуляция углерода в лесных почвах и сукцессионный статус лесов, 2018 / Под ред. Лукиной Н.В. М.: Т-во науч. изд. КМК. 232 с.
Атлас Коми АССР, 1964. М.: УГК. 112 с.
Бобкова К.С., 2007. Биологическая продуктивность и компоненты баланса углерода в заболоченных коренных ельниках Севера // Лесоведение. № 6. С. 45–54.
Бобкова К.С., Тужилкина В.В., 2001. Содержание углерода и калорийность органического вещества в лесных экосистемах Севера // Экология. № 1. С. 69–71.
Бобкова К.С., Машика А.В., Смагин А., 2014. Динамика содержания углерода органического вещества в среднетаежных ельниках на автоморфных почвах / Отв. ред. Лукина Н.В. СПб.: Наука. 270 с.
Бобкова К.С., Робакидзе Е.А., Торлопова Н.В., 2020. Круговорот элементов минерального питания в экосистеме коренного разнотравно-черничного ельника средней тайги (республика Коми) // Сиб. лесн. журн. № 2. С. 40–54. https://doi.org/10.15372/SJFS20200205
Бобкова К.С., Смоленцева Н.Л., Тужилкина В.В., Артемов В.А., 1982. Круговорот азота и зольных элементов в сосново-еловом насаждении средней тайги // Лесоведение. № 5. С. 3–11.
Бобкова К.С., Галенко Э.П., Тужилкина В.В., Сенькина С.Н., 1989. Эколого-физиологические основы продукционного процесса хвойных фитоценозов на Севере. Серия препринтов “Научные доклады”. Вып. 213. Сыктывкар: Коми НЦ УрО АН СССР. 28 с.
Варлагин А.В., Выгодская Н.Н., Курбатова Ю.А., Широня И.В., Широня Н.В., Шульце Э.-Д., 2012. Многолетняя динамика составляющих энерго-массообмена между атмосферой и лесными экосистемами // Динамика многолетних процессов в экосистемах Центрально-Лесного заповедника. Тр. ЦЛГПБЗ. Вып. 6. Великие Луки: ООО “Великолукская городская типография”. С. 71–81.
Ведрова Э.Ф., Ваганов Е.А., 2009. Углеродный бюджет бореальных лесов Средней Сибири // ДАН. № 5. С. 678–682.
Ведрова Э.Ф., Мухортова Л.В., Трефилова О.В., 2018. Участие старовозрастных лесов в бюджете углерода бореальной зоны Центральной Сибири // Изв. РАН. Сер. биол. № 3. С. 326–336. https://doi.org/10.7868/S0002332918030104
Гитарский М.Л., Замолодчиков Д.Г., Мухин В.А., Диярова Д.К., Грабар В.А. и др., 2020. Сезонная изменчивость эмиссии диоксида углерода при разложении елового валежа южной тайги Валдая // Лесоведение. № 3. С. 239–249. https://doi.org/10.31857/S0024114820030055
Горбачева Т.Т., Лукина Н.В., 2004. Органический углерод в водах подзолов ельников зеленомошных Кольского полуострова // Лесоведение. № 4. С. 43–50.
Гусев И.И., 2002. Моделирование экосистем. Архангельск: Изд-во Арханг. гос. техн. ун-та. 112 с.
Евдокимов И.В., Ларионова А.А., Шмитт М., Лопес де Гереню В.О., Бан М., 2010. Экспериментальная оценка вклада дыхания корней растений в эмиссию углекислого газа из почвы // Почвоведение. № 12. С. 1479–1488.
Загирова С.В., Михайлов О.А., Елсаков В.В., 2020. Потоки диоксида углерода, тепла и влаги между еловым насаждением и атмосферой на европейском Северо-Востоке России // Изв. РАН. Сер. биол. № 3. С. 325–336.
Замолодчиков Д.Г., 2009. Оценка пула углерода крупных древесных остатков в лесах России с учетом влияния пожаров и рубок // Лесоведение. № 4. С. 3–15.
Иванов А.В., Браун М., Замолодчиков Д.Г., Лошаков С.Ю., Потоцкий О.В., 2018. Эмиссия углерода с поверхности валежа в кедровых лесах южного Приморья // Экология. № 4. С. 275–281. https://doi.org/10.7868/S0367059718040042
Коренные еловые леса Севера: биоразнообразие, структура, функции, 2006 / Отв. ред. Бобкова К.С., Галенко Э.П. СПб.: Наука. 337 с.
Кошурникова H.Н., Верховец С.В., 2011. Продукция углерода фитомассы в южно-таежных темнохвойных лесах Западной Сибири (Кеть-Чулымский лесорастительный округ) // Растительные ресурсы. Т. 47. № 3. С. 8–21.
Кудеяров В.Н., Заварзин Г.А., Благодатский С.А., Борисов А.В., Воронин П.Ю. и др., 2007. Пулы и потоки углерода в наземных экосистемах России. М.: Наука. 315 с.
Кузнецов М.А., 2014. Выделение СО2 с поверхности почвы в ельнике чернично-сфагновом // Углерод в лесных и болотных экосистемах особо охраняемых природных территорий Республики Коми / Отв. ред. Бобкова К.С., Загирова С.В. Сыктывкар: ИБ Коми НЦ УрО РАН. С. 87–94.
Кузнецов М.А., Бобкова К.С., 2014. Потоки органического углерода в системе почва-фитоценоз ельника чернично-сфагнового средней тайги Республики Коми // Экология. № 5. С. 338–345.
Лесное хозяйство и лесные ресурсы Республики Коми, 2000 / Под ред. Козубова Г.М., Таскаева А.И. М.: ИПЦ ДИК. 512 с.
Лесные экосистемы Енисейского меридиана, 2002 / Под ред. Плешикова Ф.И., Ваганова Е.А., Ведровой Э.Ф. и др. Новосибирск: Изд-во СО РАН. 356 с.
Лесотаксационный справочник для северо-востока европейской части СССР, 1986 / Под ред. Войнова Г.С. Архангельск: Правда Севера. 357 с.
Машика А.В., 2006. Эмиссия диоксида углерода с поверхности подзолистой почвы // Почвоведение. № 12. С. 1457–1464.
Мелехина Е.Н., 2001. Почвенная микрофауна // Биопродукционный процесс в лесных экосистемах Севера / Под ред. Бобковой К.С., Галенко Э.П. СПб.: Наука. С. 234–250.
Методы изучения лесных сообществ, 2002. СПб.: НИИ Химки СПбГУ. 240 с.
Никонов В.В., Лукина Н.В., 1994. Биогеохимические функции лесов на северном пределе распространения. Апатиты: Изд-во КНЦ РАН. 315 с.
Ольчев А.В., Курбатова Ю.А., Татаринов Ф.А., Молчанов А.Г., Варлагин А.В. и др., 2009. Оценка первичной валовой и чистой продуктивности еловых лесов центрально-европейской части России с помощью полевых измерений и математической модели // Успехи соврем. биологии. Т. 129. № 6. С. 565–577.
Орлов А.Я., 1967. Метод определения массы корней деревьев в лесу и возможность учета годичного прироста органической массы в толще лесной почвы // Лесоведение. № 1. С. 64–69.
Осипов А.Ф., Кузнецов М.А., 2010. Содержание органического углерода в болотно-подзолистых почвах хвойных лесов средней тайги // Лесоведение. № 6. С. 65–70.
ОСТ 56-69-83., 1983. Пробные площади лесоустроительные. Метод закладки. М.: ЦБНТИ Гослесхоза СССР. 60 с.
Полевой определитель почв России, 2008. М.: Почв. ин-т им. В.В. Докучаева. 88 с.
Продуктивность и круговорот элементов в фитоценозах Севера, 1975 / Отв. ред. Пономарева В.В. Л.: Наука. 130 с.
Робакидзе Е.А., Торлопова Н.В., Бобкова К.С., 2013. Химический состав жидких атмосферных осадков в старовозрастных ельниках средней тайги // Геохимия. № 1. С. 72–83.
Робакидзе Е.А., Торлопова Н.В., Бобкова К.С., 2015. Химический состав снеговых вод под пологом старовозрастных еловых древостоев средней тайги Республики Коми // Лесоведение. № 6. С. 458–469.
Роде А.А., 1960. Методы изучения водного режима почв. М.: АН СССР. 244 с.
Родин Л.Е., Ремезов Н.П., Базилевич Н.И., 1968. Методические указания к изучению динамики и биологического круговорота в фитоценозах. Л.: Наука. 143 с.
Рубцов М.В., Дерюгин А.А., Соломина Ю.Н., Гурцев В.И., 1990. Водорегулирующая роль таежных лесов. М.: Агропромиздат. 221 с.
Сафонов С.С., Карелин Д.В., Грабар В.А., Латышев Б.А., Грабовский В.И. и др., 2012. Эмиссия углерода от разложения валежа в южнотаежном ельнике // Лесоведение. № 5. С. 44–49.
Смагин А.В., Садовникова Н.Б., Смагина М.В., Глаголев М.В., Шевченко Е.М. и др., 2001. Моделирование динамики органического вещества почв. М.: МГУ. 120 с.
Смоленцева Н.Л., 1979. Роль почвенных животных и микроорганизмов в разложении опада сосново-елового насаждения средней тайги // Экология роста и развития сосны и ели на Северо-Востоке Европейской части СССР. Сыктывкар: Коми филиал АН СССР. С. 104–116.
Стороженко В.Г., 1990. Датировка разложения валежа ели // Экология. № 6. С. 66–69.
Усольцев В.А., 2007. Биологическая продуктивность лесов Северной Евразии: методы, база данных и ее приложения. Екатеринбург: УрО РАН. 636 с.
Уткин А.И., 1975. Биологическая продуктивность лесов (методы изучения и результаты) / Лесоведение и лесоводство // Итоги науки и техники. М.: ВИНИТИ АН СССР. Т. 1. С. 9–189.
Уткин А.И., Замолодчиков Д.Г., Честных О.В., 2004. Углеродные пулы фитомассы, почв и депонирование углерода в еловых лесах России // Хвойные бореальной зоны. № 2. С. 21–30.
Фролова Л.Н., 1965. Особенности почвообразования в еловых лесах в связи со сменой пород в условиях Коми АССР. Автореф. дис. … канд. биол. наук. Сыктывкар. 18 с.
Хабибуллина Ф.М., 2009. Почвенная микробиота естественных и антропогенно нарушенных экосистем Северо-Востока европейской части России. Автореф. дис. … докт. биол. наук. Сыктывкар. 40 с.
Чебакова Н.М., Колле О., Золотухин Д.А., Ллойд Дж., Арнет А., Парфенова Е.И., 2002. Годичная и сезонная динамика энерго- и массоообмена в сосновом лесу средней тайги // Лесные экосистемы Енисейского Меридиана. Новосибирск: Изд-во СО РАН. С. 52–64.
Черепанов С.К., 1995. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб.: Мир и семья. 990 с.
Швиденко А.З., Щепаченко Д.Г., 2014. Углеродный бюджет лесов России // Сиб. лесн. журн. № 1. С. 69–92.
Шорохова Е.В., Шорохов А.Л., 1999. Характеристика классов разложения древесного детрита ели, березы и осины в ельниках подзоны средней тайги // Тр. СПб НИИ лесного хоз-ва. Вып. 1. С. 17–23.
Bond-Lamberty B., Wang C., Gower S.T., 2004. Contribution of root respiration to soil surface CO2 flux in a boreal black spruce chronosequence // Tree Physiol. V. 24. P. 1387–1395.
Dahlman R.C., Kuceera C.L., 1965. Root productivity and turnover in native prairie // Ecology. V. 46. № 1–2. P. 102–105.
Goulden M.L., McMillan A.M.S., Winston G.C., Rocha A.V., Manies K.L. et al., 2011. Patterns of NPP, GPP, respiration, and NEP during boreal forest succession // Glob. Change Biol. V. 17. № 2. P. 855–871. https://doi.org/10.1111/j.1365-2486.2010.02274.x
Hadden D., Grelle A., 2016. Changing temperature response of respiration turns boreal forest from carbon sink into carbon source // Agric. For. Meteorol. V. 223. P. 30–38. https://doi.org/10.1016/j.agrformet.2016.03.020
Healy R.W., Striegl R.G., Russell T.F., Hutchinson G.L., Livingston G., 1996. Numerical evaluation of static-chamber measurements of soil-atmosphere gas exchange: Identification of physical processes // Soil Sci. Soc. Am. J. V. 60. P. 740–747.
Heath G.W., Edwards C.A., Arnold M.K., 1964. Some methods for assessing the activity of soil animals in the breakdown of leaves // Pedobiologia. V. 4. № 1–2. P. 80–87.
IUSS Working Group WRB, 2015. World Reference Base for Soil Resources 2014, update 2015. International soil classification system for naming soil and creating legends for soil map. World Soil Resources Reports № 106. Rome: FAO. 192 p.
Johnston C.A., Groffman P., Breshears D.D., Cardon Z.G., Currie W. et al., 2004. Carbon cycling in soil // Front. Ecol. Environ. № 2. P. 522–528.
Krankina O.N., Harmon M.E., 1995. Dynamics of the dead wood carbon pool in northern-western Russian boreal forests // Water Air Soil Pollut. V. 82. P. 227–238.
Kriiska K., Frey J., Asi E., Kabral N., Uri V. et al., 2019. Variation in annual carbon fluxes affecting the SOC pool in hemiboreal coniferous forests in Estonia // For. Ecol. Manag. V. 433. P. 419–430. https://doi.org/10.1016/j.foreco.2018.11.026
Law B.E., Sun O.J., Campbell J., Tuyl S., van, Thornton P.E., 2003. Changes in carbon storage and fluxes in a chronosequnece of ponderosa pine // Glob. Change Biol. V. 9. P. 510–524. https://doi.org/10.1046/j.1365-2486.2003.00624.x
Lindroth A., Grelle A., Moren A.-S., 1998. Long-term measurements of boreal forest carbon balance reveal large temperature sensitivity // Glob. Change Biol. V. 4. P. 443–450.
Luyssaert S., Schulze E.-D., Börner A., Khohl A., Hessenmöller D. et al., 2008. Old-growth forests as global carbon sinks // Nature. V. 455. P. 213–215. https://doi.org/10.1038/nature07276
Magnani F., Mencuccini M., Borghetti M., Berbigier P., Berninger F. et al., 2007. The human footprint in the carbon cycle of temperate and boreal forests // Nature. V. 447. P. 848–850. https://doi.org/10.1038/nature05847
Mahli Y., Baldocchi D., Jarvis P., 1999. The carbon balance of tropical, temperate and boreal forests // Plant Cell Environ. V. 22. P. 715–740.
Manual on methods and criteria for harmonized sampling, assessment, monitoring and analysis of the effects of air pollution on forest, 2011. Part XI. 30 p. http://icp-forests.net/page/icp-forests-manual
Milyukova I.M., Kolle O., Varlagin A.V., Vygodskaya N.N., Schulze E.D., Lloyd J., 2002. Carbon balance of a southern taiga spruce stand in European Russia // Tellus B. Chem. Phys. Meteorol. V. 54. P. 429–442. https://doi.org/10.3402/tellusb.v54i5.16679
Mukhin V.A., Diyarova D.K., Gitarskiy M.L., Zamolodchikov D.G., 2021. Carbon and oxygen gas exchange in woody debris: The process and climate-related drivers // Forests. V. 12. № 9. https://doi.org/10.3390/f12091156
Pan Y., Birdsey R.A., Fang J., Houghton R., Kauppi P.E., Kurz W.A., 2011. A large and persistent carbon sink in the world’s forests // Science. V. 333. P. 988–993.
Pregitzer K.S., Euskirchen E.S., 2004. Carbon cycling and storage in world forests: Biome patterns related to forest age // Glob. Change Biol. V. 10. P. 2052–2077. https://doi.org/10.1111/j.1365-2486.2004.00866.x
Stocker T.F., Qin D., Plattner G.-K., Alexander L.V., Allen S.K., et al., 2013. Technical summary // Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge Univ. Press. P. 33–115. https://doi.org/10.1017/CBO9781107415324.005
Ueyama M., Harazono Y., Kim Y., Tanaka N., 2009. Response of the carbon cycle in sub-arctic black spruce forests to climate change: Reduction of a carbon sink related to the sensitivity of heterotrophic respiration // Agric. For. Meteorol. V. 149. P. 582–602
Uri V., Kukumägi M., Aosaar J., Varik M., Becker H. et al., 2019. The carbon balance of a six-year-old Scots pine (Pinus sylvestris L.) ecosystem estimated by different methods // For. Ecol. Manag. V. 433. P. 248–262. https://doi.org/10.1016/j.foreco.2018.11.012
Valentini R., Matteucci G., Dolman A.J., Schulze E.-D., Rebmann C. et al., 2000. Respiration as the main determinant of carbon balance in European forests // Nature. V. 404. P. 861–865.
Zhangurov E.V., Lebedeva M.P., Zaboeva I.V., 2011. Microstructure of genetic horizons in automorphic soils of the Timan Ridge // Eurasian Soil Sci. V. 44. P. 261–271. https://doi.org/10.1134/S1064229311030203
Дополнительные материалы отсутствуют.
Инструменты
Журнал общей биологии