

Океанология, 2020, T. 60, № 2, стр. 250-258
Закономерности использования берега ларгой в Японском море
В. А. Нестеренко 1, *, И. О. Катин 2
1 Федеральный научный центр биоразнообразия наземной биоты
Восточной Азии ДВО РАН
Владивосток, Россия
2 Национальный научный центр морской биологии ДВО РАН
Владивосток, Россия
* E-mail: vanester@mail.ru
Поступила в редакцию 15.04.2019
После доработки 23.10.2019
Принята к публикации 16.12.2019
Аннотация
Рассмотрены закономерности использования тюленями берега в зависимости от местоположения лежбищ, их геоморфологических параметров и влияния океанических условий. Установлено, что при характерной для ларги (пятнистой нерпы) на юге Японского моря береговой репродукции для родов и выкармливания детенышей используются только примкнувшие пляжи с косами, пересыпи и прислоненные пляжи в карманообразных абразионных бухтах малых и особо малых островов, обеспечивающие необходимый в репродуктивный период уровень укрытости.
В пределах обширного ареала, протянувшегося от Чукотского до Желтого моря, ларга (Phoca largha Pallas, 1811) распределена неравномерно и выделено несколько участков тихоокеанской акватории, в пределах которых тюлени концентрируются в период размножения [21]. Группировкам тюленей в этих зонах придавался ранг самостоятельных популяций [8, 18] или даже подвидов [1]. Данные молекулярно-генетических исследований [25, 26, 27, и др.] не позволили однозначно трактовать внутриродовую таксономическую структуру и территориальные группировки ларги определяются пока как “репродуктивные концентрации” [15, 24, 29]. Так называемый Южный популяционный сегмент [21], объединяющий генетически близкие [26] территориальные группировки ларги Японского и Желтого морей, насчитывает 5 тыс. особей, из которых численность первой составляет около 3 тыс. особей. В отличие от всех других видов ластоногих, которые размножаются либо на льду, либо на берегу [20], для ларги известна как ледовая, так и береговая репродукция, причем северные группировки являются преимущественно пагофильными, а для южных группировок исторически сложившейся нормой является береговое размножение [14]. Репродукция ларги на льдах описана ранее [17, 18, 21, 22, 28 и др.]. Что касается берегового размножения, то хотя на примере ларги в заливе Петра Великого Японского моря доказано, что из-за невозможности синхронизации процессов репродукции с динамикой и свойствами ледового субстрата, роды и выкармливание детенышей у этого вида происходит здесь на островных лежбищах [6], особенности использования берега тюленями остаются недостаточно изученными. Немаловажно, что в приконтинентальной части Японского моря ларга связана с берегом круглый год и в зависимости от фазы популяционного цикла предъявляет различные требования к выбору участков нахождения на суше. Соотношения между местоположением лежбищ, их геоморфологическими параметрами и влиянием океанических условий играют решающее влияние на характер использования берега тюленями на разных фазах жизненного цикла и особенно в репродуктивный период, и данная статья посвящена выявлению этих закономерностей.
РАЙОН ИССЛЕДОВАНИЙ. МАТЕРИАЛ И МЕТОДИКА
Японское море является окраинным морем Тихого океана, сообщающимся с Восточно-Китайским и Желтым морями через Корейский пролив, с Охотским морем – через проливы Татарский и Лаперуза и с Тихим океаном через Сангарский пролив. Крупнейшим заливом (площадь около 10.2 тыс. км2) северо-западной части Японского моря является зал. Петра Великого, на акватории которого расположено до 50 больших (1000 га и более), малых (не более 10 га) и особо малых (около 1 га) островов. Территориально острова объединены в несколько групп (рис. 1). Южную группу составляют шесть кекуров и островов, самым крупным из которых является о. Фуругельма. В центральной части расположен архипелаг Римского-Корсакова, состоящий из 11 островов, из которых девять – малых и особо малых. Архипелаг Евгении, представляющий собой вытянутую на 30 км на юго-запад от п-ова Муравьева-Амурского гряду из 25 островов, включающих такие большие острова как Русский и Попова, составляет Северную островную группу. Самостоятельную группу в восточной части залива формируют пять кекуров и островов, основными из которых являются о. Путятина и о. Аскольд. Поскольку все лежбища ларги зарегистрированы на островах [6] в соответствии с вышеназванными группами выделено четыре лежбищных района: Южный, Центральный, Северный и Восточный, в которых сосредоточено 95% всех зарегистрированных в заливе лежбищ ларги [15].
Рис. 1.
Лежбищные районы в зал. Петра Великого Японского моря: 1 – Южный, 2 – Центральный, 3 – Северный, 4 – Восточный.
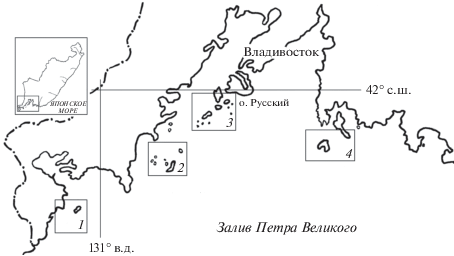
Зал. Петра Великого находится в зоне муссонной циркуляции, при которой зимний муссон с ветром северных направлений устанавливается в октябре и длится до апреля. Условия в заливе детально описаны в специальных публикациях [2, 9, 11, 16, 19 и др.]
Исследования по изучению особенностей распределения ларги на акватории западной части Японского моря проводятся с 1982 г. С 1997 по 2014 гг. осуществлялся круглогодичный мониторинг использования лежбищ тюленями в зал. Петра Великого.
Данные по динамике использования берега тюленями были получены во время маршрутов на моторных лодках. В ходе исследований существующие методики учета были модернизированы и разработан ряд новых, включая методику дистанционного определения возраста детенышей, методику описания лежбищ и методику накопительного учета приплода [5, 6, 13, 14 ]. Кроме системы регулярных маршрутных учетов на лежбищах и обследования акватории с лодки велись скрытые (с использованием длиннофокусной оптики) наблюдения с берега, позволяющие определить детали размещения тюленей, специфику их распределения по лежбищу и другие характеристики, доступные только при отсутствии беспокойства животных. Продолжительность прямых наблюдений за тюленями составила более 10 тыс. ч.
Со времени рождения детенышей до их перехода к самостоятельному образу жизни ежегодно осуществлялось регулярное слежение за семейными группами и альянсами тюленей. Идентификация особей велась по кормящим самкам, имеющим характерные особенности окраски, шрамы и другие естественные метки.
С 2009 по 2013 гг. проводилось мечение ларги методом горячего таврения [23] и помечена 951 особь. Суммарно накоплено 836 фактов повторных регистраций меченых животных, на основании анализа которых оценивали особенности использования пространства животными. В теплые сезоны велось регулярное слежение за распределением тюленей, как в заливе, так и вдоль побережья Приморья.
Представленные в статье океанографические данные основываются на оперативном и ретроспективном анализе спутниковой информации высокого и среднего пространственного разрешения (15–250 м), полученной с различных космических аппаратов. В 2004–2012 гг. со спутниковыми измерениями синхронизировались визуальные и инструментальные наблюдения в зал. Петра Великого [3, 4].
В данной статье приняты следующие термины и понятия. Под лежбищем мы понимаем топографически ограниченный участок супралиторали с установленным характером его использования тем или иным видом ластоногих. Береговые объединения ларги – это открытые социально структурированные группировки тюленей, функционирующие на лежбищах на определенных фазах популяционного цикла. Характерный уровень численности береговых объединений является биологической характеристикой лежбища, это количественный показатель, выведенный в результате многолетних наблюдений и отражающий биологическую емкость лежбища [13].
Характеристики льда соответствуют международной номенклатуре [12]. Названия грунтов даны по классификации, предложенной Лопатиным с соавторами [10]. Для статистического анализа использован пакет программ Statistica 10.0.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЯ
Ларга в зал. Петра Великого формирует береговые объединения четырех типов [14]. В прелиминарных объединениях концентрируются вернувшиеся из миграций животные. Репродуктивные объединения представляют собой сложные социальные структуры [15], после рождения детенышей состоящие из коалиции нескольких семейных триад (самка–самец–детеныш). По мере завершения периода репродукции в апреле формируются пострепродуктивные, предмиграционные береговые объединения, названные линными только поскольку время их функционирования совпадает с периодом линьки тюленей. Распад группировок этого типа обусловлен началом миграций и оставшиеся в заливе резиденты широко распределяются по заливу, формируя восстановительные объединения. Смена береговых объединений складывается в ежегодный цикл, причем в ходе трансформации последовательных типов объединений, осуществляющегося путем распада одних с одновременным образованием других, происходит перераспределение животных по лежбищам [14].
На разных фазах жизненного цикла тюлени предъявляют различные требования к используемому участку берега. К первой группе требований (неизменные) относятся местоположение и геоморфология лежбища, включая его площадь и характеристику субстрата, ко второй группе (изменчивые) – орография лежбища и формы его трансформации под влиянием океанологических условий. Основными критериями для выбора участка берега для нахождения на суше являются степень укрытости (т.е. подверженности конкретного участка берега воздействию океанологических условий, ведущих к гибели животных или снижению успешности протекания биологических процессов) и безопасности (возможности контролировать обстановку, чтобы избежать опасности неблагоприятного воздействия). “Идеальным” лежбищем должен был бы быть такой участок островного берега, при котором его укрытость и безопасность являлись максимальными. Однако таких участков в заливе нет, и на всех пригодных для функционирования береговых объединений ларги лежбищах тюлени подвергаются той или иной степени риска.
Использование берега в репродуктивный период. Репродуктивная фаза годового популяционного цикла ларги, во время которой в зал. Петра Великого проходят массовые роды детенышей и их выкармливание, является наиболее уязвимым периодом. После родов кормящие самки продолжительное время находятся на берегу, а детеныши проводят на пляже большую часть суток. В этот период фактор беспокойства должен быть сведен к минимуму. Кроме того, количество пригодных для выхаживания новорожденных островных участков должно быть достаточным для того, чтобы животные имели возможность их выбора, как на начальных этапах формирования репродуктивных объединений, так и при изменении океанологических условий.
В Южном лежбищном районе нет береговых участков, подходящих для прохождения процессов репродукции, и лежбища здесь не используются для родов и выкармливания детенышей. В Северном лежбищном районе есть береговые участки, по геоморфологическим характеристикам потенциально пригодные для проведения процессов репродукции, но в условиях максимального по заливу антропогенного прессинга на островах архипелага Евгении нет условий для функционирования репродуктивных объединений ларги. В Восточном лежбищном районе лежбища также используются только для формирования восстановительных береговых объединений (табл. 1). Причины отсутствия здесь объединений других типов те же: сильное антропогенное давление и более чем 70-километровая удаленность от расположенного на островах архипелага Римского-Корсакова репродуктивного популяционного ядра.
Таблица 1.
Лежбища ларги в зал. Петра Великого (кроме Центрального района): характеристика и особенности использования тюленями
| Лежбище | Геоморфоло- гический тип |
Площадь* | Тип берегового объединения** (уровень численности) | ||
|---|---|---|---|---|---|
| № | местоположение | название | |||
| Южный лежбищный район | |||||
| 1 | кам. Бутакова | Камни Бутакова | Рифовое | 20 000 | В (30) |
| 2 | о. Веры | Остров Веры | Пляжное | 600 | В (10) |
| 3 | о. Фуругельма | Мыс Свиньина | Рифовое | 2500 | В настоящее время не используется |
| 4 | кам. Михельсона | Камни Михельсона | Рифовое | 7000 | В (40) |
| Северный лежбищный район | |||||
| 5 | о. Желтухина | Желтухинское | Рифовое | 15 000 | В (20) |
| 6 | о. Моисеева | Моисеевское | Пляжное | 1000 | В (15) |
| 7 | о. Сергеева | Сергеевское | Пляжное | 750 | В (10) |
| 8 | о-ва Верховского | Верховского | Рифовое | 20 000 | В (10) |
| Восточный лежбищный район | |||||
| 9 | кек. Пять пальцев | Пять пальцев | Рифовое | 25 000 | В (20) |
| 10 | о-ва Унковского | Унковского | Рифовое | 20 000 | В (40) |
| Лежбища вне лежбищных районов | |||||
| 11 | о-ва Гаккеля | Гаккель | Рифовое | 10 000 | В (15) |
| 12 | кам. Алексеева | Камни Алексеева | Рифовое | 3000 | В (20) |
| 13 | кекуры Бакланьи | Бакланье | Рифовое | 45 000 | В (80) |
| 14 | кекур Колонна | Колонна | Рифовое | 21 000 | В (15) |
В противоположность рассмотренным выше районам Центральный лежбищный район имеет очевидные достоинства. Малые острова здесь расположены достаточно компактно и в удалении от материка. Береговые условия на этих островах разнообразны и оказываются подходящими для протекания разных биологических процессов, поэтому на них образуются объединения самого разного функционального назначения. Решающим фактором, является то, что острова расположены в районе с существенно сниженным прямым антропогенным воздействием из-за нахождения на акватории Дальневосточного морского заповедника, где исключена ведущая к трансформации среды деятельность и обеспечивается низкий уровень беспокойства на всех лежбищах. Именно здесь и только здесь сосредоточены все лежбища, на которых формируются репродуктивные объединения и соответственно рождаются практически все детеныши ларги рассматриваемой территориальной группировки (табл. 2).
Таблица 2.
Лежбища ларги в Центральном лежбищном районе зал. Петра Великого: характеристика и особенности использования тюленями
| Лежбище | Геоморфологический тип | Площадь, в м2 | Экспозиция, в ° | Тип берегового объединения* (уровень численности**) |
|
|---|---|---|---|---|---|
| о. Большой Пелис | Тельняшка | Бухтовое | 350 | 195 | Р (15) |
| Каблук | Пляжное (пересыпь), с рифовым выделом | 2500 | 90 и 270 | Р (20), в (40) | |
| Антимолчанского | Пляжное | 3500 | 90 | Р (10), Л (100) | |
| о. Кентавр | Кентавр | Пляжное (пересыпь), с рифовым выделом | 3000 | 45 и 220 | П (280), Р (15), Л (200), В (5) |
| о. Матвеева | Восточное | Пляжное | 6300 | 210 | П (450), Р (50), Л (400), |
| Водопадное | Бухтовое | 200 | 220 | Р (7) | |
| Глухарь | Бухтовое | 1300 | 220 | Р (30) | |
| Малый Глухарь | Бухтовое | 300 | 270 | Р (8) | |
| Северо-западное | Пляжное, пересыпь | 6400 | 90 и 270 | П (550), Р (30), Л (600) | |
| о. 2-й Камень | Бельгийское | Бухтовое | 700 | 30 | Р (15) |
| Таблеточное | Пляжное (пересыпь), с рифовым выделом | 2900 | 270 | П (180), Р (30), Л (300), В (20) | |
| Сверчковое | Бухтовое | 900 | 220 | Р (15) | |
| Малое | Бухтовое | 150 | 45 | Р (8) | |
| о. 3-й Камень | 3-й Камень | Пляжное (коса) | 700 | 30 и 160 | П (80), Р (15), Л (60) |
| о. 4-й Камень | 4-й Камень | Пляжное (пересыпь), с рифовым выделом | 500 | 45 и 220 | Р (15), Л (40), В (25) |
| о. Дурново | Купол | Бухтовое | 250 | 90 | Р (12) |
| Сапсан | Бухтовое | 1000 | 165 | Р (50), Л (100) | |
| Филин | Бухтовое | 350 | 230 | Р (14) | |
| Минное | Пляжное | 3000 | Р (50), Л (250) | ||
| о. Гильде-брандта | Кекурное | Бухтовое | 250 | 180 | Р (15) |
| Оленье | Пляжное (коса) | 1000 | 90 и 270 | Р (20), Л (100), В (10) | |
| о. Де-Ливрона | Южное | Бухтовое | 1800 | 160 | Р (90) |
| Кабанье | Пляжное | 3500 | 0 и 270 | Р (50) | |
| Сосновое | Бухтовое | 400 | 155 | Р (22) | |
| Тригопункт | Бухтовое | 700 | 60 | Р (33) | |
| о. Стенина | Подвершинное | Рифовое | 100 | 215 | В (15) |
| Голова быка | Рифовое | 350 | 70 | В (15) | |
| Бычий хвост | Пляжное (коса) | 200 | 0 и 220 | Р (5), В (5) | |
Репродуктивные береговые объединения функционируют только на лежбищах пляжного и бухтового типа, субстратом на которых всегда является мелкофракционный пляж и береговой лед.
К бухтовым на островах арх. Римского-Корсакова относятся 14 лежбищ (табл. 2), которые обычно представляют собой галечно-булыжный пляж с валунами и глыбами, среди которых аккумулируется галечник. Ледовые новообразования на прилегающих акваториях не развиваются больше стадии темного ниласа. Дрейфующий лед обычно проходит мористее, но на некоторых лежбищах возможно поджатие льда к берегу и его сплачивание с образованием кратковременного припая и даже торошением льда на берегу, при котором полностью перекрывается берег (рис. 2а).
Рис. 2.
Ледяной покров в районе архипелага Римского-Корсакова на изображении (http://pacificinfo.ru/eisp/piter), полученном 30.01.2018 г. со спутника Sentinel-2B (а) и торошение льда на лежбище Восточное (а, на врезке); лежбище Северо-Западное (б): до (на врезке) и после перемыва пересыпи.

Укрытость бухтового лежбища может быть очень высокой, в том случае, когда оно находится в волновой тени, либо очень низкой, когда волновое воздействие прямое. Безопасность этих лежбищ находится в обратной зависимости от укрытости. Из-за орографических особенностей находящиеся на таких лежбищах тюлени не могут уверенно контролировать обстановку.
При внезапной резкой смене силы и направления волн детеныши ларги могут оказаться в “ловушке” и погибнуть. Так, в 2005 г. при специальном осмотре всех лежбищ с репродуктивными объединениями зарегистрировано 12 мертвых детенышей недельного возраста, которые были вморожены в ледяные торосы, что, вероятно, свидетельствует об их гибели во время сильного волнового наката. Однако при постепенном изменении условий самки имеют достаточно времени, чтобы переместиться на соседние лежбища, перетаскивая, или сопровождая детеныша. Именно поэтому важно, что на небольшой территории расположено более десятка лежбищ с разными условиями.
Таким образом, предпочтения, связанные с отношением укрытость/безопасность в период репродукции, смещены в пользу необходимости большей укрытости и суммарный вклад в воспроизводство ларги в бухтовых лежбищах в отдельные годы составляет более 60%.
Между размером лежбища и максимально зарегистрированном на нем количеством новорожденных существует корреляция (rs = 0.7561, при p < 0.05), но эта зависимость характеризует скорее емкость лежбища, т.е. предельно возможное количество семейных триад на определенном береговом участке при условиях близких к оптимальным. При выборе лежбища беременными самками в каждый конкретный год площадь лежбища не имеет существенного значения. В зависимости от складывающейся ситуации различия “вклада” в репродукцию того или иного лежбища может отличаться год от года более чем на 90%. Так, за период 2000–2014 гг. число ежегодно рожденных детенышей колебалось на лежбище Кабанье от 2 до 27, на Сосновом – от 6 до 22, Тригопункт – от 8 до 33 и т.д. Такой разброс обусловлен влиянием океанологических условий. Пляж лежбища Кентавр, укрытого от волнений южного, юго-восточного и восточного направлений, регулярно промывается северными ветровыми волнами, в результате чего площадь субстрата существенно меняется, что отражается на количестве триад в репродуктивных объединениях. Наибольшее трансформирующее воздействие на это лежбище оказывают волны зыби, приходящие с юго-запада и, если это случается в период выкармливания детенышей, гибель некоторых из них весьма вероятна. Размер репродуктивных береговых объединений на пересыпях связан с высокой подвижностью мелкофракционного аккумулированного грунта. Так, например, пляж протяженностью 320 м между о. Матвеева и 1-м Камнем Матвеева, на котором расположено лежбище Северо-западное, регулярно перемывается с образованием пролива шириной до 100 м (рис. 2б). Немаловажным фактором изменения количества детенышей на том или ином лежбище является перекрывающее берег торошение льда, препятствующее доступу тюленей на лежбище. Примером подверженных поджатию дрейфующего льда с севера и его сплачиванию являются имеющие северо-восточную экспозицию лежбища Малое и Бельгийское. Кроме того, выбор участка для родов, вероятно, обусловлен также социальными причинами [15] и историей его использования особями.
Из 15 лежбищ пляжного типа 12 также расположены в Центральном лежбищном районе (табл. 2). Грунт пляжей на лежбищах этого типа не больше мелкого булыжника. Рельеф простой, слабонаклонный, но с периодическим образованием крутого волнового набоя и террас. Примкнувшие пляжи образуются на самых малых островах, а на более крупных островах образование таких пляжей возможно только в том случае, если коренной массив острова оказался вдоль вектора волнового воздействия и береговая линия его слабо изрезана. Непременный мысообразный излом береговой линии продолжается в виде подводной косы. Площадь субстрата обычно изменяется несущественно, но могут происходить перемещения грунта с изменением рельефа и береговой черты. В случае, когда аккумулятивный берег соединяет два коренных массива, образуется пересыпь с оппозитными выделами условно восточной и западной экспозиции. Используя эту особенность, тюлени в зависимости от внешней обстановки занимают тот или иной выдел лежбища, причем переходить животные могут и по суше. В противоположность примкнувшим пляжам, у пересыпей площадь субстрата может изменяться многократно: грунт пляжа мелкофракционный, нескелетезированный, очень подвижный и поэтому даже в течение года он неоднократно смывается с образованием широких проливов (рис. 2б). Подвижность грунта пересыпей обусловливает регулярное изменение площади таких лежбищ. Следует принять во внимание, что, несмотря на значительную общую площадь, в период репродукции используется лишь отдельные участки лежбища.
В ледовый период пляжные лежбища могут забиваться дрейфующим льдом, который на некоторое время сплачивается и смерзается. При обледенении камней и валунов пространство между ними заполняется льдом, неровности рельефа сглаживаются с образованием плоской поверхности, и происходит улучшение свойств субстрата, позволяющее использовать его тюленями. В большинстве случаев изменение ледовой обстановки на лежбищах пляжного типа не ведет к блокированию к ним доступа для тюленей. Исключением является лишь лежбище Бычий хвост, которое редко используется для формирования репродуктивных объединений в силу особенности расположения на северо-западе о. Стенина. Здесь происходит постоянное выдавливание льда на берег с образованием торосов высотой до трех метров и такой лед, перекрывающий грунт пляжа, не пригоден для процессов репродукции.
Таким образом, пляжные лежбища обладают высокой степенью безопасности, но слабой укрытостью.
Использование берега в пост- и предмиграционный периоды. Прелиминарные береговые объединения ларги формируются только на 5 пляжных лежбищах, компактно расположенных на островах Матвеевской гряды (рис. 1, табл. 2) в Центральном лежбищном районе. Все эти лежбища позже используются для репродукции.
Линные береговые объединения функционируют на 9 лежбищах пляжного типа, включая те, на которых в свое время функционируют прелиминарные объединения, а также на крупноразмерных пляжах островов Б. Пелис, Дурново и Гильдебрандта. Из бухтовых лежбищ на начальных этапах формирования линных береговых объединений крупные агрегации тюленей наблюдаются только на пляже лежбища Сапсан.
В отличие от репродуктивной фазы годового цикла, в пред- и постмиграционный периоды площадь лежбища является крайне важным требованием для формирования береговых объединений ларги. Это обусловлено необходимостью выстраивания отношений между мигрантами из разных потоков и их социализации с резидентами, а также между образующими ядра репродуктивных береговых объединений беременными самками в прелиминарных объединениях и для формирования миграционных групп в преддверии очередного миграционного периода в линных объединениях, что оптимально в составе крупных скоплений.
Хотя линька является индивидуальным процессом и не требует агрегированного нахождения тюленей на берегу, в этот период ларги особо требовательны к субстрату, в первую очередь к гранулометрической структуре грунта, обеспечивающей комфортное вытирание меха и достаточно сильное прогревание. Кроме того, из-за специфики поведения особей в эти периоды и необходимости много времени проводить на берегу они более требовательны к безопасности, чем к укрытости.
Использование берега в миграционный период. К рифовому типу относятся три из четырех лежбищ в Южном лежбищном районе, по два лежбища в Центральном, Северном и Восточном лежбищных районах (табл. 1, 2), а также четыре лежбища, не входящие в состав лежбищных районов. Кроме того, в состав трех лежбищ Центрального лежбищного района входят выделы рифового типа.
Субстрат на рифовых лежбищах представлен двумя типами: скальными образованиями (рифы, глыбы и валуны) или мелкофракционным пляжем. В зависимости от приливно-отливных процессов площадь субстрата меняется незначительно. При намораживании площадь не увеличивается, а образующиеся при этом ледяные карнизы и козырьки являются препятствием в использовании рифов тюленями зимой.
Основным преимуществом лежбищ рифового типа является повышенная безопасность находящихся на нем тюленей и при неблагоприятных условиях они могут быстро покинуть лежбище. Недостатком лежбищ рассматриваемого типа является практически полное отсутствие укрытости. По этим причинам использование лежбищ рифового типа невозможно для родов и выкармливания детенышей и на них регистрируется функционирование только восстановительных объединений.
Восстановительные береговые объединения ларги образуются на лежбищах всех четырех лежбищных районов. На лежбищах бухтового типа такие объединения не формируются, а на пляжных лежбищах тюлени чаще выходят не на сам пляж, а на рифы, расположенные на прилегающей к лежбищу акватории. Во время исследовательских перемещений сеголетков или трофических перемещений особей-резидентов по заливу одиночные тюлени или их небольшие группы могут использовать для нахождения на берегу для отдыха так называемые “транзитные пункты”. Их количество в зал. Петра Великого ограничено [15] и резидентные ларги, по-видимому, хорошо их знают и временно используют по мере необходимости.
Большая часть тюленей в летний период покидает зал. Петра Великого. Миграции идут в нескольких направлениях [7], но в независимости от направленности перемещений ларга в летний период использует участки берега аналогичные тем, которые используются в зал. Петра Великого, а именно рифовые участки побережий материка и островов.
ЗАКЛЮЧЕНИЕ
Таким образом, использование берега ларгой по мере цикличной трансформации ее береговых объединений в зал. Петра Великого существенно различается. Для родов и выкармливания детенышей используются только примкнувшие пляжи с косами и пересыпи(лежбища пляжного типа) и прислоненные пляжи в карманообразных абразионных бухтах (лежбища бухтового типа). При береговом размножении ларги именно такие участки обеспечивают необходимый в репродуктивный период уровень укрытости. Основным риском в этот период является вероятность смерти детенышей при изменении океанических условий, когда животные оказываются в “ловушке” и погибают. Несмотря на наличие геоморфологически пригодных для проведения процессов репродукции островных участков по всей акватории залива, массовые роды проходят только на архипелаге Римского-Корсакова. Именно эта островная система обеспечивает, с одной стороны, необходимую степень концентрации пригодных для успешного проведения процессов репродукции береговых участков, с другой стороны, высокий уровень безопасности тюленей из-за нахождения в зоне охраны акватории Морским заповедником. В постмиграционный и совпадающий у ларги с линькой предмиграционный период тюленям для выстраивания социальных отношений между разными популяционными стратами необходимы большие береговые площади, способные вместить сотни особей и для этого пригодными оказываются только пляжи с косами и пересыпи (лежбища пляжного типа). В летний период, когда мотивация группового нахождения на берегу практически отсутствует, и нет необходимости в субстрате мелкофрационного пляжа для проведения процессов линьки, репродукции и социализации, ларги как в зал. Петра Великого, так и вдоль материкового побережья Японского моря используют рифовые участки (лежбища рифового типа), характеризующиеся максимальным уровнем безопасности.
Список литературы
Волошина И.В. Береговые тюлени Японского моря. Владивосток: Русский остров, 2007. 304 с.
Гидрометеорология и гидрохимия морей. Т. 3. Японское море // Гидрометеорологические условия / Под ред. Васильева А.С. и др. СПб.: Гидрометеоиздат, 2003. 398 с.
Дубина В.А., Митник Л.М., Катин И.О. Особенности циркуляции вод залива Петра Великого на основе спутниковых мультисенсорных данных // Современное состояние и тенденции изменения природной среды залива Петра Великого Японского моря. М.: ГЕОС, 2008. С. 82–96.
Ростов И.Д., Плотников В.В., Дубина В.А. и др. Информационно-справочная система “Режим и изменчивость ледяного покрова залива Петра Великого (Японское море)” // Метеорология и гидрология. 2017. № 11. С. 113–118.
Катин И.О. Ларга залива Петра Великого Японского моря (характеристика и использование лежбищ, состояние популяции): Дис. … канд. биол. наук: 03.00.08. Владивосток, 2006. 146 с.
Катин И.О., Нестеренко В.А. Океанологические условия и репродукция пятнистой нерпы (Phoca largha) в заливе Петра Великого Японского моря // Океанология. 2010. Т. 50. № 1. С. 82–88.
Катин И.О., Нестеренко В.А. Хоминг неполовозрелых особей ларги (Phoca largha) по результатам мечения // Зоол. журн. 2012. Т. 91. № 3. С. 1273–1276.
Косыгин Г.М., Тихомиров Э.А. Ларга (Phoca largha Pallas) залива Петра Великого // Изв. ТИНРО. 1970. Т. 70. С. 114–137.
Ластовецкий, Е.И., Якунин, Л.П. Гидрометеорологическая характеристика Дальневосточного государственного морского заповедника // Цветковые растения островов Дальневосточного морского заповедника. Владивосток: ДВНЦ АН СССР, 1981. С. 18–33.
Лопатин Б.Г., Борщева П.А., Власова Н.П. и др. Методические рекомендации по изучению донных образований Арктического шельфа. Л.: НИИГА, 1981. 80 с.
Лоция Японского моря. Ч. I. Изд. ГУНИО, 1972. 287 с.
Международная символика для морских ледовых карт и номенклатура морских льдов. Л.: Гидрометеоиздат, 1984. 56 с.
Нестеренко В.А., Катин И.О. Лежбище: объем понятия, порядок установления // Экология. 2009. № 1. С. 53–59.
Нестеренко В.А., Катин И.О. Цикл трансформации береговых объединений ларги (Phoca largha Pallas, 1811) в заливе Петра Великого Японского моря // Биол. моря. 2010. Т. 36. № 1. С. 48–55.
Нестеренко, В.А, Катин, И.О. Ларга (Phoca largha) в заливе Петра Великого. Владивосток: Дальнаука, 2014. 219 с.
Справочник по волнению: Прибрежная зона Приморья / Под ред. Якунина Л.П. Владивосток: ДВНИГМИ, 1976. 62 с.
Тихомиров Э.А. Некоторые данные о распределении и биологии ларги в Охотском море в летне-осенний период и организация ее промысла // Изв. ТИНРО. 1966. Т. 58. С. 105–115.
Трухин А.М. Ларга. Владивосток: Дальнаука, 2005. 246 с.
Юрасов Г.И., Яричин В.Г. Течения Японского моря. Владивосток: ДВО РАН, 1991. 176 с.
Boeus B.J. Sexual strategies of seals and walruses // New scientist. 1986. № 49. P. 35–39.
Boveng P.L., Bengtson J.L., Buckley T.W. et al. Status review of the spotted seal (Phoca largha) / U.S. Dep. Comm. NOAA Tech. Memo. NMFS-AFSC-200, 2009. 146 p.
Lowry L.F., Burkanov V.N., Frost K.J. et al. Habitat use and habitat selection by spotted seals (Phoca largha) in the Bering Sea // Can. J. Zool. 2000. V. 78. P. 1959–1971.
Merrick R.L., Loughlin T.R., Calkins D.G. Hot branding: a technique for long-term marking of pinnipeds / US. Dep. Commer. NOAA Tech. Memo. NMFS-AFSC, 1996. 21 p.
Mizuno A.W., Wada A., Ishinazaka T. et al. Distribution and abundance of spotted seals Phoca largha and ribbon seals Phoca fasciata in the southern Sea of Okhotsk // Ecological Research. 2002. V. 17. P. 79–96.
Mizuno A.W., Onuma M., Takahashi M. et al. Population genetic structure of the spotted seal Phoca largha along the coast of Hokkaido, based on mitochondrial DNA sequences // Zoological Science. 2003. V. 20. P. 783–788.
O’Corry-Crowe G., Lowry L.F., Burkanov V.N. et al. The biogeography and population structure of spotted seals (Phoca largha) using mitochondrial DNA // 14-th Biennial Conference on the Biology of Marine Mammals. Canada, Vankuver, 2001. P. 159.
O’Corry-Crowe G.M., Westlake R.L. Molecular investigation of spotted seals (Phoca largha) and harbour seals (P. vitulina) and their relationship in areas of sympatry // Molecular genetics of marine mammals: Special Publication N. 3 / Eds. Dizon A.E. et al. Lawrence: The Society for Marine Mammalogy, 1997. P. 291–304.
Rugh D.J., Shelden K.E.W., Withrow D.E. Spotted seal, Phoca largha, in Alaska // Mar. Fish. Rev. 1997. V. 59. № 1. P. 1–18.
Shaughnessy P.D., Fay F.H. A review of the taxonomy and nomenclature of North Pacific harbour seals // J. Zool. 1977. V. 182. P. 385–419.
Дополнительные материалы отсутствуют.