

Океанология, 2020, T. 60, № 4, стр. 576-594
Потоки осадочного вещества в Лофотенской котловине Норвежского моря: сезонная динамика и роль зоопланктона
А. В. Дриц 1, *, А. А. Клювиткин 1, **, М. Д. Кравчишина 1, В. А. Карманов 2, А. Н. Новигатский 1
1 Институт океанологии им. П.П. Ширшова РАН
Москва, Россия
2 Московский государственный университет им. М.В. Ломоносова,
факультет почвоведения
Москва, Россия
* E-mail: adrits@mail.ru
** E-mail: klyuvitkin@ocean.ru
Поступила в редакцию 06.04.2020
После доработки 06.04.2020
Принята к публикации 08.04.2020
Аннотация
По материалам годовой (август 2017–июль 2018 гг.) экспозиции автоматической глубоководной седиментационной обсерватории (АГОС) проведено исследование процессов седиментации в Лофотенской котловине Норвежского моря. Получены оценки вертикальных потоков осадочного вещества, органического углерода (Сорг), карбоната кальция (CaCO3), аморфного кремнезема (SiO2ам), алюминия и основных частиц планктонного происхождения (птероподы, фораминиферы, фекальные пеллеты, некрозоопланктон). В поверхностном 0–500 м слое годовой ход вертикальных потоков характеризуется унимодальным распределением с максимумом в сентябре 2017 г. (191 мг/м2/сут). Начиная с ноября, величина потока осадочного вещества заметно снижается, достигая минимальных значений (2.6 мг/м2/сут) в марте–апреле 2018 г. Максимальные величины потока Сорг (5.2 мг/м2/сут), CaCO3 (178 мг/м2/сут) и SiO2био (9.1 мг/м2/сут) наблюдаются в осенний период. На глубине 2950 м, из-за переосаждения из нефелоидного слоя, величина потоков и вклад литогенного материала выше, а амплитуда изменений потоков в течение года заметно ниже. Наиболее высокие величины потока обусловлены массовым осаждением молоди птеропод Limacina retroversa. Вклад птеропод в поток CaCO3 в осенний период составлял 60–100%, в поток Сорг – 56–73%. Вклад фораминифер в общий поток СaCO3 не превышал 6%, и только в октябре на 550 м достигал 60%. Поток пеллетного материала, в котором преобладали пеллеты остракод, изменялся от <0.01 до 2.1 мгС/м2/сут, составляя от 0.1 до 90% потока Сорг. Поток некрозоопланктона (НЗ) в слое 0–550 м изменялся от 0 до 0.45 мгС/м2/сут, с максимумом в феврале. Относительно суммарного (с учетом НЗ) потока Сорг, поток НЗ составлял 0–33% в летне-осенний сезон, 12–52% зимой и 23–41% весной. В слое 550–2950 м поток НЗ изменялся от 0.04 до 0.22 мгС/м2/сут, что составляло 0.7–2% потока Сорг. Суммарный вклад всех исследованных компонентов планктонного происхождения в годовой поток карбоната кальция составлял 16–47%, в годовой поток Сорг – 37–45%. Анализ результатов наших исследований и данных литературы, полученных в 90-е годы прошлого столетия, показал, что наблюдаемые климатические изменения (повышение температуры и солености, увеличение толщины слоя атлантической воды) в Лофотенской котловине Норвежского моря не привели к кардинальным изменениям количественных характеристик процесса седиментации.
ВВЕДЕНИЕ
Изучение осадочного вещества от его формирования до отложения на дно – важнейшая задача современной седиментологии. Рассеянное осадочное вещество, взвешенное в многокилометровой толще вод океанов, является тем материалом, из которого, в конечном счете, образуются донные осадки и осадочные породы [8]. Потоки осадочного вещества в океане – это одна из основных количественных характеристик осадкообразования, которая позволяет в динамике изучать процессы седиментации осадочного вещества, преобразование его при прохождении водной толщи, оценивать количество и состав вещества, поступающего на дно. Согласно биоседиментационной концепции “живого океана” [2, 9] осадочный материал, поступивший в океан из разных источников (с речным стоком, аэрозолями, эндогенным веществом), трансформируется под влиянием морской биоты. Формируя вертикальные потоки осадочного материала в морях и океанах, живые организмы являются важнейшими участниками седиментационных процессов. Количественная характеристика этих процессов, контролирующих вертикальный транспорт взвешенного вещества, имеет фундаментальное значение для понимания глобального цикла углерода в океане и процессов современного осадкообразования. Особый интерес представляют исследования потока и состава осадочного материала в течение года, поскольку дают возможность получить представление не только об изменчивости интенсивности седиментации, но и о сезонной динамике структуры и функционировании экосистем эпипелагиали.
Норвежское море является транзитной зоной, через которую осуществляется перенос теплой и соленой Атлантической воды (АВ) из Северной Атлантики в Северный Ледовитый океан [31, 48, 55]. Лофотенская котловина служит основным тепловым резервуаром субарктических морей и одновременно энергоактивной зоной, которая напрямую влияет на климат Европы и дальнейшее поступление АВ в Баренцево море и пролив Фрама [41, 59].
Неудивительно, что изучению процессов седиментации и механизмов его формирования в этом районе посвящено значительное число работ [22, 30, 39, 44, 52, 60], большая часть которых была выполнена в 80–90-е годы прошлого столетия. Вместе с тем, с начала 2000-х годов усиливается “атлантификация” Северного Ледовитого океана, что способствует переносу тепла через Норвежское море в северном и северо-восточном направлениях [38, 46]. По данным [42] в Норвежском море наблюдается тенденция к увеличению температуры воды, как в поверхностном, так и в более глубоком слое, за счет увеличения теплосодержания АВ. Для оценки возможного влияния климатических изменений на количественные характеристики процессов седиментации в этом районе необходимы современные данные о потоках осадочного вещества, его составе и масштабе сезонной изменчивости.
В связи с этим целью нашей работы было исследование вертикальных потоков осадочного вещества в Лофотенской котловине Норвежского моря по материалам годовой экспозиции автоматической глубоководной седиментационной обсерватории (АГОС).
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЙ
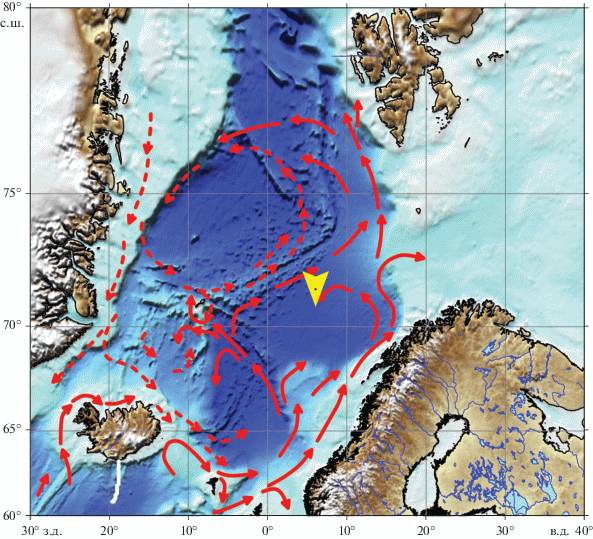
Материал для исследования собирался в Лофотенской котловине Норвежского моря в течение года с 22 июля 2017 г. (68-й рейс НИС “Академик Мстислав Келдыш” [6]) по 17 июля 2018 г. (71-й рейс НИС “Академик Мстислав Келдыш” [13]) с помощью автоматической глубоководной седиментационной обсерватории (АГОС [10, 11]) (рис. 1).
Рис. 1.
Расположение АГОС (ст. 5522), 71°33.773′ с.ш. и 06°04.864′ в.д., глубина 3000 м на карто-схеме субполярной Северной Атлантики. Стрелками показана общая циркуляция поверхностных вод [21].
Гидрологические исследования и отбор проб воды. Для изучения водной толщи при постановке и подъеме АГОС использовался зондирующий комплекс SBE911p/SBE-32 с 10-литровыми батометрами Нискина (24 шт.), датчиками температуры, электропроводности, давления и мутности.
Было отобрано 13 проб взвеси. Концентрацию взвеси определяли стандартным методом фильтрации под вакуумом 400 мбар через мембранные фильтры (0.45 мкм размер пор, 47 мм диаметр фильтра). Концентрацию взвеси определяли взвешиванием фильтров с точностью до ±0.01 мг, соотнося с объемом профильтрованной воды. Для определения концентрации хлорофилла “а” (хл “а”) пробы воды фильтровали через предварительно прокаленные (при температуре 450°С) стекловолокнистые фильтры Whatman GF/F под вакуумом не более 200 мбар. Определение концентрации хл “a” проводилось флуориметрическим методом (с экстрагированием в 90%-м ацетоне) на флуориметре Trilogy 1.1 (Turner Designs, США) в судовой лаборатории по методике [15]. Калибровка прибора выполнена при помощи стандарта хл “а” фирмы “Sigma” (C6144–1MG).
Сбор осадочного вещества. Осадочное вещество собиралось седиментационными ловушками на двух горизонтах: 550 м для оценки потока из верхнего квазиоднородного слоя атлантической воды; 2950 м для оценки потока, достигающего дна и участвующего в формировании осадков. Для сбора осаждающегося материала использовались 12-стаканные конические седиментационные ловушки “Лотос-3” с площадью сбора 0.5 м2 производства ОКБ ОТ РАН, которые позволяют получить непрерывный во времени (до года и более) ряд проб осаждающего вещества с заданной экспозицией (30 суток). Для исключения биологической трансформации собираемого вещества флаконы-пробосборники ловушек заполнялись раствором HgCl2 (1% от насыщенного раствора). Соленость фиксирующего раствора доводилась добавлением NaCl до двухкратной солености морской воды на горизонте постановки ловушки (~70 епс) для исключения вымывания фиксирующего раствора из пробосборников. Каждую ловушечную пробу предварительно фильтровали через сито с размером ячеи 1 мм, после чего отбирали по 30 мл для анализа состава ловушечного материала под бинокулярным микроскопом. Смыв с сита (частицы крупнее 1 мм) собирали в отдельный флакон. Оставшуюся часть пробы фильтровали аналогично пробам взвеси для определения величины суммарных потоков и дальнейших исследований состава вещества. Одновременно вели фильтрацию через прокаленные при температуре 450°С стекловолокнистые фильтры Whatman GF/F для определения содержания общего (Собщ), органического (Сорг) и карбонатного (Скарб) углерода.
Анализ осадочного вещества. Определение валового Si и Al проводилось фотометрическим методом по методике ГЕОХИ РАН [3] с дополнениями А.Б. Исаевой (ИО РАН) для работы с малыми навесками вещества (точность 2–5%). Аморфный кремнезем (SiO2ам) определялся методом терригенной матрицы по разнице содержаний валового и литогенного кремнезема.
Углерод общий и органический определяли методом кулонометрического титрования на анализаторе АН-7560 (Россия) до и после подкисления 1 н HCl, точность 5–10%. Содержание Скарб оценивали по разнице содержаний Собщ и Сорг. Содержание CaCO3 рассчитывали по стехиометрическому уравнению: 100/12 × Скарб.
Изучение микроморфологии и состава осаждающегося вещества выполнено с помощью сканирующего электронного микроскопа TESCAN VEGA 3 с микрозондовой приставкой Oxford INCA Energy 350.
Концентрацию хлорофилла в поверхностном слое моря во время экспозиции АГОС рассчитывали с использованием поисковой системы Ocean Color Web (https://oceancolor.gsfc.nasa.gov/) и программного обеспечения SeaSAS 7.5.3 (https:// seadas.gsfc.nasa.gov/). Данные спутникового сканера цвета моря MODIS-Aqua [54] LAC (Local Area Coverage) 3-го уровня (Level 3) осреднялись для квадратов 20 × 20 и 50 × 50 км.
Анализ компонентов планктонного происхождения в материале ловушек. Предварительный просмотр под бинокуляром проб осадочного материала, полученных после префильтрации через 1 мм сито, показал, что наиболее многочисленными компонентами размерной фракции <1 мм были раковины пелагических крылоногих моллюсков Limacina retroversa и планктонных фораминифер, и фекальные пеллеты (ФП). Кроме того, в отдельных пробах были встречены радиолярии отряда Spumellaria, яйца планктонных животных и объекты, по внешнему виду и размерам соответствующие сперматофорам кальмаров [37] (в дальнейшем “сперматофоры”, рис. 2).
Подсчет и измерение диаметра раковин лимацин и фораминифер проводили под бинокулярным микроскопом Leica при увеличении ×100 либо во всей пробе, либо в 1/10–1/200 пробы (в зависимости от численности раковин). Подсчет и классификацию пеллет проводили в 1/10 пробы. Подсчет в подпробах проводили в 3 повторностях. Для оценки содержания органического углерода в ФП (Сфп), радиоляриях, сперматофорах и яйцах отбирали 100–200 пеллет одной формы, 3–5 радиолярий, 2–3 сперматофора и 30–50 яиц и после троекратной промывки в дистиллированной воде отфильтровывали через прокаленный стекловолокнистый фильтр GF/F диаметром 25 мм. Сфп измеряли на анализаторе Shimadzu TOC-VCPH. Для оценки содержания органического и неорганического углерода в лимацинах из проб с их высокой численностью случайным образом отбирали 100–300 раковин и после троекратной промывки в дистиллированной воде отфильтровывали через прокаленные стекловолокнистые фильтры GF/F диаметром 25 мм. Содержание общего и органического углерода измеряли на анализаторе Shimadzu TOC-VCPH до и после подкисления 1н HCl, содержание неорганического (карбонатного) углерода рассчитывали по разнице Собщ и Сорг. Аналогичным методом измеряли эти параметры в фораминиферах размером 0.2–0.4 мм, для мелких фораминифер (диаметр <0.1 мм) измеряли только Собщ. Содержание органического и неорганического углерода в этих фораминиферах рассчитывали, исходя из соотношений Сорг/Собщ и Скарб/Собщ, полученных для крупных форм.
Подсчет и определение видового состава некрозоопланктона (НЗ) проводили в камере Богорова при просмотре под бинокулярным микроскопом Leica смыва с сита после префильтрации проб ловушек. Учитывали только особей с хорошо выраженным разрушением тканей тела и отслоением тканей от хитиновых покровов тела [4]. Для определения содержания органического углерода в НЗ, отобранных из ловушек животных каждого вида (1–5 экз.) после промывания в дистиллированной воде помещали на прокаленный стекловолокнистый фильтр GF/F. Измерения содержания Сорг в НЗ проводили аналогично определению Сорг в ФП.
Отбор проб донных осадков. Пробы донных осадков были отобраны с помощью мультикорера Mini Muc K/MT 410, KUM GmbH (Германия). Из верхнего слоя осадка были изготовлены препараты с помощью стандартных предметных стекол методом смер-слайда, которые изучались на судне под поляризационным микроскопом Ломо ПОЛАМ Л-213М с камерой МС-5 (USB 2.0). Смер-слайд представляет собой тонкий слой (“мазок”) несцементированного осадка, закрепленный на предметном стекле с помощью оптической жидкости (канадского бальзама с показателем преломления n = 1.55). Смер-слайд является надежным экспресс-методом для полуколичественной диагностики минерального и гранулометрического состава осадков, а также определения микро(нано)фоссилий. Изучение смер-слайдов дает весьма точное представление о вещественном составе отложений [12]. Карбонатность осадков определяли эспресс-методом, основанным на измерении давления СО2↑ при реакции HCl с карбонатом кальция, с помощью кальциметра.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Гидрологические условия, концентрация взвеси и хлорофилла “а”
Вертикальное распределение температуры, солености, концентрации взвеси и хл “а” в водной толще на ст. 5522 показано на рис. 3.
Рис. 3.
Вертикальное распределение параметров среды (температура, соленость, кислород и мутность) по данным CTD-зондирования и концентрации водной взвеси и хл “а” на станции АГОС в момент постановки (а) и подъема (б) ловушек. 1 – температура, 2 – соленость, 3 – мутность, 4 – концентрация взвеси, 5 – концентрация хлорофилла.
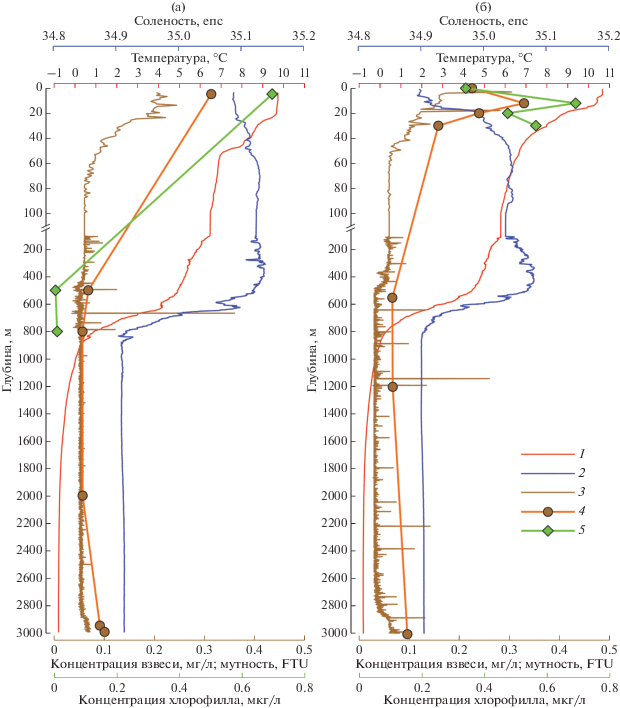
Относительно теплая и соленая атлантическая водная масса занимала в Лофотенской котловине верхний слой толщиной 625–695 м. Соленость АВ в этом слое варьировала от 34.95 до 35.13 епс. Приповерхностный слой (около 0–20 м) обычно несколько опреснен, что ярче было выражено в августе 2018 г. (до 34.9 епс). В слое 0–50 м и в тот, и в другой год выделялся двухступенчатый сезонный термоклин, который слабо проявлялся в августе 2018 г. Температура воды в верхнем опресненном слое достигала 10.7°С в августе 2018 г. и была примерно на 1°С ниже в июле 2017 г. В целом температура АВ массы в изученный период варьировала от 9.7–10.7°С вблизи поверхности до 2.0°С на границе с холодной промежуточной водной массой. В качестве нижней границы АВ принимаем изопикническую поверхность σ = 27.9 кг/м3 вслед за работой [56].
Глубинная водная масса занимает слой от 625–695 м до дна. Температура воды здесь достигала отрицательных величин (до –0.76°С), а соленость составляла около 34.9 епс (рис. 3). Максимальная плотность в придонном слое достигала 28.08 кг/м3.
Концентрация взвеси в слое 0–5 м составила 0.31 мг/л в июле 2017 г. и 0.23 мг/л в августе 2018 г. Концентрация хл “а” варьировала от 0.69 до 0.34 мкг/л соответственно. В августе 2018 г. зафиксирован подповерхностный максимум взвеси (0.33 мг/л на глубине 15 м), который совпадал с максимумом хл “а” (0.69 мкг/л).
Концентрация взвеси уменьшалась с глубиной в несколько раз и возрастала в придонном нефелоидном слое. Мощность нефелоидного слоя по данным CTD-зондирований с использованием нефелометра варьировала от 150 м в июле 2017 г. до 350 м в августе 2018 г. (рис. 3).
Анализ спутниковых данных показал, что максимум концентрации хлорофилла в поверхностном слое воды Лофотенской котловины приходился на конец лета–осень 2017 г. (рис. 4). Осенью 2018 г. концентрации хлорофилла в этом районе были значительно ниже.
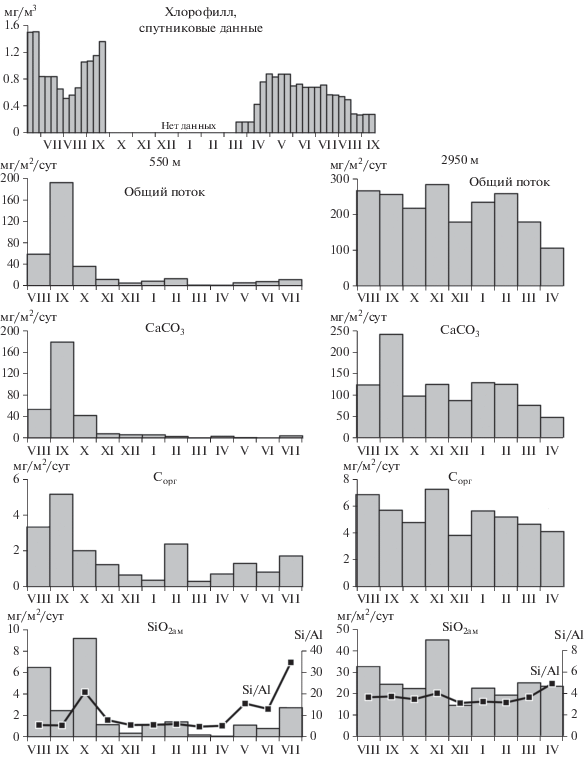
Общий поток осадочного вещества
Анализ материала ловушек показал значительную изменчивость общих потоков вещества в течение года в подповерхностном слое (горизонт 550 м) и относительную однородность потоков в придонном слое (горизонт 2950 м) (рис. 4).
В подповерхностном слое годовой ход потоков характеризуется унимодальным распределением с максимумом в сентябре 2017 г. (191 мг/м2/сут). Начиная с ноября, величина потока заметно снижается, достигая минимальных значений (2.6 мг/м2/сут) в марте–апреле 2018 г. С мая по июль поток возрастает с 7.0 до 12.9 мг/м2/сут.
В придонном слое средняя величина вертикального потока в семь раз выше, чем в подповерхностном слое (221.0 и 30.6 мг/м2/сут соответственно). При этом изменчивость потоков в течение года менее выражена по сравнению с верхним слоем. Самые низкие величины потока в придонном слое (107 мг/м2/сут) приходятся на апрель, что совпадает с минимумом потока в подповерхностном слое.
Поток основных биогенных компонентов
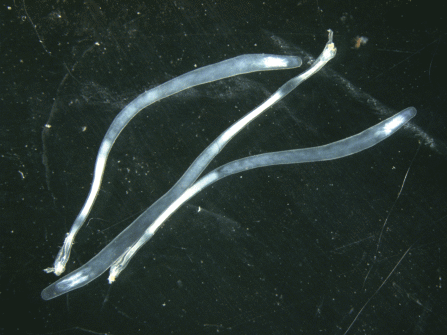
При изучении проб под сканирующим электронным микроскопом в составе осадочного вещества подповерхностного и придонного горизонтов в августе–октябре было обнаружено большое количество крупных (до 700 мкм) раковин птеропод Limacina retroversa (рис. 5). Именно в эти месяцы в подповерхностном слое отмечены высокие величины потока СаСО3 с максимумом (178 мг/м2/сут) в сентябре (рис. 4). В этот период поток СаСО3 составлял более 90% от всего осадочного вещества. В остальное время года поток CaCO3 варьировал от 0.3 до 9.3 мг/м2/сут. В придонном слое максимальный поток СаСО3 наблюдался также в сентябре (240 мг/м2/сут), оставаясь достаточно высоким (до 129 мг/м2/сут) с августа по февраль.
Рис. 5.
Микрофотографии ловушечного вещества в августе, иллюстрирующие преобладание раковин Limacina retroversa: (а) горизонт 550 м и (б) горизонт 2950 м.

Наибольший поток Сорг в подповерхностном слое (2.0–5.2 мг/м2/сут) наблюдался в августе–октябре (рис. 4). В остальное время года поток Сорг не превышал 1–2 мг/м2/сут. В придонном слое поток Сорг на протяжении всего года изменялся незначительно от 3.8 до 7.2 мг/м2/сут.
Поток SiO2ам в подповерхностном слое достигал наибольших величин (9.1 мг/м2/сут в октябре) в октябре. В остальное время года поток не превышал 2.7 мг/м2/сут. В придонном слое поток SiO2ам был заметно выше и изменялся в течение года в диапазоне от 14.5 до 45.0 мг/м2/сут.
Отношение Si/Al в подповерхностном слое большую часть года колебалось в интервале 5.0–6.2, резко возрастая в октябре (20.9) и в мае–июле (13.2–34.6). Обогащение осадочного вещества валовым Si по отношению к Al указывает на увеличение доли биогенного кремния в составе потока. Содержание Si в осадочном веществе придонного слоя (12.4–19.9%) приближается его среднему содержанию в пелагических донных осадках океана (19.6% [7]). Отношение Si/Al в придонном слое составляет 3.1–4.0, что близко к среднему значению отношения этих элементов в земной коре [49, 61]. Лишь в июле Si/Al возрастает до 4.9, незначительно обогащая осадочное вещество придонного слоя биогенным Si.
Седиментация компонентов планктонного происхождения
Птероподы Limacina retroversa. Вся популяция лимацин, обнаруженная в материале ловушек, была представлена молодью со средним диаметром раковины 0.49 ± 0.19 мм (550 м) и 0.48 ± ± 0.12 мм (2950 м). Практически все раковины были пустыми или с небольшим количеством органического вещества. Сезонный ход седиментации этих моллюсков характеризуется выраженным максимумом (13 000–15 000 раковин/м2/сут) в сентябре на глубине 550 м и в сентябре–октябре на 2950 м (рис. 6). В ноябре вертикальный поток птеропод резко снижается и в оставшийся период практически равен нулю.
Рис. 6.
Внутригодовая изменчивость вертикальных потоков разных компонентов планктонного происхождения.
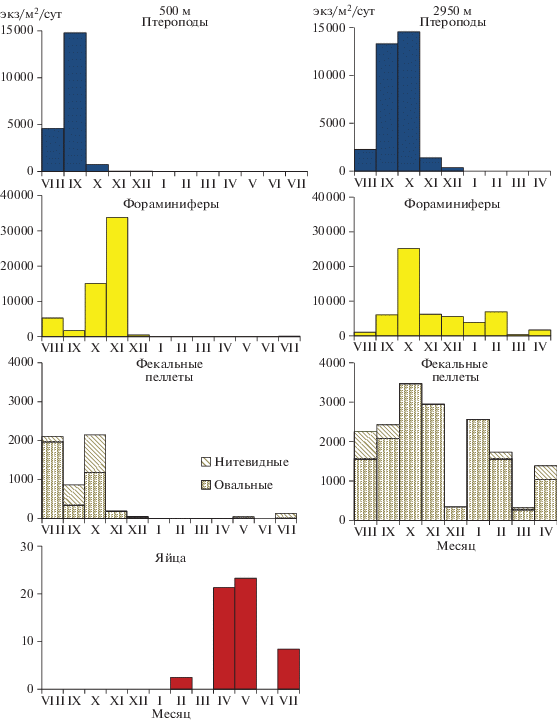
Планктонные фораминиферы. Среди раковин фораминифер наиболее многочисленными были мелкие экземпляры <0.1 мм, доля более крупных раковин (0.2–0.4 мм) не превышала 5%. Изменения потока этих организмов в течение года носят унимодальный характер с максимумом в октябре–ноябре (рис. 6). В зимние месяцы фораминиферы обнаружены только в пробах глубоководной ловушки.
Фекальные пеллеты (ФП). В пеллетном материале по численности преобладали овальные пеллеты, нитевидные пеллеты в заметном количестве были встречены только в сентябре–октябре в ловушках на глубине 550 м (рис. 6). Основной поток пеллет в верхнем 550 метровом слое (900–2000 пел/м2/сут) был приурочен к концу вегетационного периода, в остальное время года поток был почти на порядок ниже и не превышал 200 пел/м2/сут. В слое 550–2950 м минимальный поток пеллет (200–300 пел/м2/сут) был отмечен в декабре и в марте, в остальное время поток изменялся от 1500 до 3500 пел/м2/сут.
Яйца планктонных животных были встречены только в пробах ловушек на 550 м в весенне-летний период. Максимальный поток в мае–июне составлял 21–23 экз/м2/сут.
Данные о содержании Сорг и Скарб в рассмотренных выше планктонных компонентах осадочного вещества приведены в табл. 1.
Таблица 1.
Содержание общего (Собщ), органического (Сорг ) и карбонатного (Скарб) углерода (мкг/экз) в разных компонентах осадочного вещества
| Собщ | Сорг | Скарб | |
|---|---|---|---|
| Лимацины | 1.07 ± 0.52 (6) | 0.24 ± 0.09 (3) | 0.83 |
| Фораминиферы <0.1 мм | 0.028 ± 0.010 (9) | 0.0078 | 0.02 |
| Фораминиферы 0.2–0.4 мм | 0.14 ± 0.08 (3) | 0.04 | 0.10 |
| Пеллеты овальные | 0.48 ± 0.12 (6) | ||
| Пеллеты нитевидные | 1.34 | ||
| Яйца 0.25 мм | 1.65 ± 0.09 (2) | ||
| Яйца 0.5 мм | 7.46 |
Сезонные изменения потока Сорг и СaCO3, обусловленные седиментацией разных компонентов, показаны на рис. 7. В слое 0–550 м высокие величины потока Сорг (2.2–4.4 мгС/м2/сут) характерны для позднелетнего–осеннего периода. Основу потока в августе составляли птероподы и ФП (по 1.1 мгСорг/м2/сут), в сентябре – птероподы (3.6 мгС/м2/сут), в октябре – ФП (1.9 мгСорг/м2/сут). В другие сезоны поток Сорг в этом слое существенно ниже (0.001–0.4 мгСорг/м2/сут). Вклад всех компонентов в общий поток Сорг максимален в позднелетний–осенний сезон, достигая в октябре 100%. В январе–марте вклад в общий поток Сорг не превышает 1%, в апреле возрастает до 22%, затем снижается до 1% в июне и вновь увеличивается до 12% в июле. Увеличение доли планктоногенных компонентов в первом случае обусловлено седиментацией яиц, во втором – нитевидных ФП. Сезонные изменения потока Сорг и его состава в слое 550 м–дно сходны с описанным для верхнего слоя с высокими значениями (2.4–5.2 мгСорг/м2/сут) в сентябре–октябре и заметным снижением потока в другие сезоны. В осенний период поток Сорг определялся, главным образом, птероподами, в остальное время – ФП. Суммарный вклад этих компонентов в общий поток Сорг изменялся от 80–100% в августе–сентябре до 4–35% в зимне-весенний период.
Рис. 7.
Внутригодовая изменчивость потока Сорг и СaCO3, обусловленные седиментацией разных компонентов планктонного происхождения (столбцы) и их суммарный вклад в поток Сорг и СaCO3 (линия).
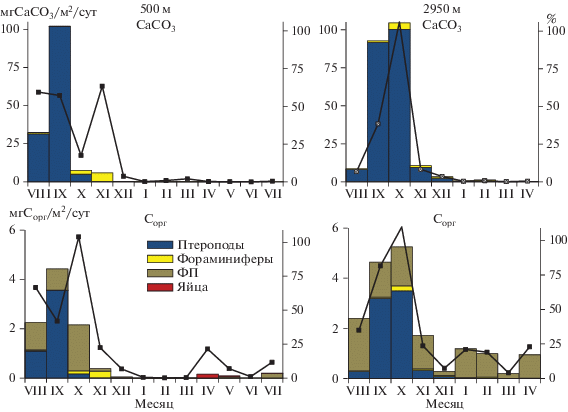
Поток СaCO3 во всем столбе воды, обусловленный, главным образом, седиментацией птеропод изменялся от <1 до 104 мг/м2/сут (рис. 7). Максимальные величины были получены в сентябре–октябре. В это время от 40 до 100% общего потока карбоната кальция определялось седиментацией раковин Limacina retroversa. Вклад фораминифер в общий поток СaCO3 не превышал 6%, и только в одном случае (октябрь, 550 м) достигал 60%.
Некрозоопланктон (НЗ). Количество и состав НЗ, обнаруженного в материале ловушек, существенно различались на разных глубинах (рис. 8). В ловушках на 500 м наиболее многочисленными в течение всего года были копеподы Metridia longa, в отдельные периоды отмечалась высокая численность глубоководных копепод Heterorhabdus norvegicus и Paraeuchaeta sp. В придонном слое только в трех пробах были встречены копеподы Gaetanus sp., поток трупов этих копепод составлял 0.3, 1.7 и 1.3 экз/м2/сут в октябре, ноябре и январе соответственно. Кроме того, в двух пробах (январь и апрель) присутствовали радиолярии отряда Spumellaria (поток – 3.4 и 3.9 экз/м2/сут), в одной (февраль) – “сперматофоры” кальмаров (поток – 1.2 экз/м2/сут). Поток Сорг в слое 0–550 м, рассчитанный с учетом данных о содержании органического углерода в НЗ (табл. 2), изменялся от 0 до 0.45 мгСорг/м2/сут, с максимумом в феврале (рис. 9). В зависимости от сезона менялся вклад разных видов: в ноябре–декабре более половины суммарного потока было обусловлено седиментацией трупов Paraeuchaeta sp., в январе–марте – Amphipoda, в апреле–мае – M. longa. Относительно суммарного (с учетом НЗ) потока Сорг, поток НЗ составлял 0–33% в летне-осенний сезон, 12–52% зимой, 23–41% весной.
Рис. 8.
Внутригодовая изменчивость вертикальных потоков разных видов некрозоопланктона на глубине 550 м. M. longa – Metridia longa, H. norvegica – Heterorhabdus norvegica, P. sp. – Paraeuchaeta sp., G. sp. – Gaetanus sp., Euph. – Euphausiidae, Amph. – Amphipoda.
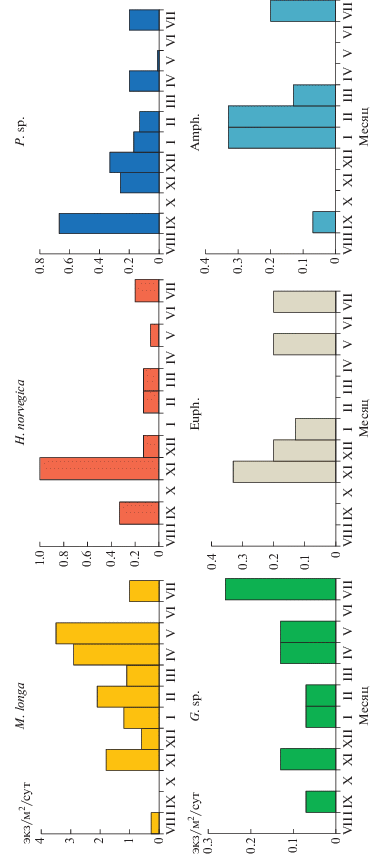
Таблица 2.
Содержание органического углерода (мкгС/экз) в разных видах некрозоопланктона
| Месяц | Metridia longa | Heterorhabdus norvegica | Gaetanus sp. | Paraeuchaeta sp. | Calanus sp. | Euphausiidae | Amphipoda | |||
|---|---|---|---|---|---|---|---|---|---|---|
| самки | самцы | самки | самки | самки | CV | 5–6 мм | 9–10 мм | 10–12 мм | 12–14 мм | |
| IX | – | – | 36.5 | – | 312 ± 214 (3) | – | – | – | – | – |
| XI | 44 ± 19 (2) | 27 ± 5 (3) | 40 ± 6 (3) | 73 ± 32 (2) | 689 ± 181 (2) | – | 62 ± 13 (3) | – | – | – |
| XII | 84 | 24 ± 2 (2) | 73 | – | 565 ± 27 (3) | – | 61 | 326 | – | |
| I | 42 ± 3 (3) | 23 ± 5 (3) | – | – | 468 ± 50 (3) | – | – | – | – | 567 ± 235 (4) |
| II | 40 ± 6 (2) | 14 ± 3 (4) | 12 | – | 313 ± 96 (2) | – | – | – | – | – |
| III | 55 ± 7 (3) | 20 | – | 31 | – | – | – | – | – | 770 |
| IV | 72 ± 40 (2) | – | 13 | 433 ± 191 (3) | 30 | – | – | – | 724 | |
| V | 54 ± 3 (3) | 13 ± 4 (3) | – | 82 | 182 | 70 | – | – | 419 | – |
| VII | 62 ± 10 (2) | 19 ± 2 (2) | – | 101 ± 20 (2) | 215 ± 3 (2) | – | – | – | – | 857 ± 126 (3) |
Рис. 9.
Внутригодовая изменчивость потока Сорг, обусловленная седиментацией разных видов некрозоопланктона (столбцы) и их суммарный вклад в поток Сорг (линия). M. longa – Metridia longa, H. norvegica – Heterorhabdus norvegica, P. sp. – Paraeuchaeta sp., G. sp. – Gaetanus sp., Euph. – Euphausiidae, Amph. – Amphipoda., Rad. – радиолярии, Sper. – сперматофоры кальмаров.
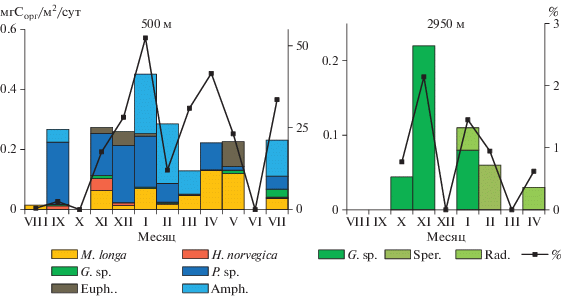
Поток Сорг в слое 550–2950 м, рассчитанный с учетом содержания Сорг в копеподах Gaetanus sp. (139.5 ± 23.0 мгСорг/экз., n = 3), радиоляриях (8.4 ± ± 7.2 мгСорг/экз, n = 3) и “сперматофорах” (46.1 ± ± 3 мгСорг/экз., n = 3), изменялся от 0.04 до 0.22 мгСорг/м2/сут, что составляло 0.7–2% потока Сорг (рис. 9).
Биогенные остатки в донных осадках. Вскрытая толща голоценовых донных осадков (0–26 см) была представлена карбонатным (более 30% СаСО3) пелитовым илом с содержанием пелитовой фракции (<0.01 мм) до ~80%. В составе верхнего слоя (0–10 см) осадков преобладали (более 80%) остатки планктонных и бентосных организмов: фораминиферы, кокколиты, диатомовые, радиолярии, спикулы губок. Среди мирофоссилий, обнаруженных в осадке, абсолютно преобладали кокколитофориды (~65%), фораминиферы составляли 10–15%, а остальные были представлены единичными экземплярами. Среди кокколитофорид преобладали два вида: крупноклеточный Coccolithus pelagicus и мелкоклеточный Emiliania huxleyi. Следует подчеркнуть, что птероподы и их фрагменты в изученных осадках не наблюдались, несмотря на преобладание этих организмов в составе осадочного вещества в водной толще. В более глубоких слоях осадка содержание биогенных остатков снижалось.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Лофотенская котловина Норвежского моря играет важную роль в поддержании глобальной меридиональной циркуляции в области, где атлантические водные массы отдают тепло в атмосферу, перемешиваются с окружающими водами и претерпевают трансформацию, формируя глубинные воды в соседнем Гренландском море [1]. По данным многолетних наблюдений [24] влияние потепления АВ в этом районе в период с 1993 по 2017 г. наиболее выражено. Это проявляется в том, что даже при уменьшении притока АВ за этот период на 1 Sv (данные норвежской Ocean Weather Station М, OWSM, 66° с.ш. и 2° в.д.) [34], толщина слоя АВ увеличивается за счет повышения солености и температуры в этом слое [62].
Для оценки степени влияния этих изменений на характеристики процессов седиментации мы сравнили наши результаты с данными годовых седиментационных ловушек, полученных в Лофотенской котловине в 80–90-х годах прошлого века (табл. 3). Как видно из таблицы, наши оценки годового потока потоков осадочного вещества и Сорг на глубине 550 м (11 г/м2 и 0.8 гСорг/м2) в 2–2.5 раза ниже по сравнению с опубликованными данными. Эти различия могут быть проявлением межгодовой изменчивости, которая отмечалась в предыдущих исследованиях [22, 44]. Другой причиной снижения потока осадочного вещества и Сорг в период наших работ может быть уменьшение притока АВ в Норвежское море и, как следствие, уменьшение количества приносимой взвеси. В придонном слое различия менее выражены: по нашим данным годовой поток осадочного вещества составляет 79 г/м2, по данным [35, 44] – 23–62 г/м2, годовой поток Сорг составил 1.9 гСорг/м2, а по данным [35, 44] – 1.4–2.7 гСорг /м2. Годовой поток CaCO3 и SiO2ам в верхнем слое сопоставим с величинами, измеренными в 80–90-х годах прошлого века, а в придонном слое – в несколько раз выше.
Таблица 3.
Годовой поток (г/м2/год) осадочного вещества (общий поток), взвешенного органического углерода (Сорг), карбоната кальция (CaCO3) и биогенного кремния (Siбио) в Лофотенской котловине по данным годовых седиментационных ловушек 1983–1992 гг.
Характерная особенность процессов седиментации в глубоководных районах Норвежского моря в конце прошлого столетия – значительное увеличение потока осадочного вещества и всех его компонентов в придонном слое [22, 23, 44]. По мнению цитируемых авторов, это связано с латеральной адвекцией взвешенного вещества из районов континентального склона и его переотложением. Согласно их расчетам более 50% общего потока не может быть объяснено вертикальным осаждением из вышележащих слоев, а, скорее, обусловлено переосаждением из нефелоидного слоя. Многократное увеличение потока осадочного вещества на фоне высокой мутности в придонном нефелоидном слое, отмеченные в нашей работе, также указывают на значительную роль процессов ресуспензии и переотложения взвешенного вещества в Лофотенской котловине.
Характер сезонной динамики вертикальных потоков в Лофотенской котловине практически не изменился за период с 1988–1992 гг. по 2017–2018 гг.: по данным [35, 44, 60] максимальные величины потока осадочного вещества, Сорг и СаСО3 размерной фракции <1 мм наблюдались в августе–октябре, по нашим данным (рис. 4) – в сентябре. Так же как в настоящей работе, сезонный ход вертикальных потоков в более ранних исследованиях [44] наиболее выражен на глубине 500 м. В придонном слое амплитуда изменений потоков в течение года заметно ниже. По мнению [60], сдвиг сезонного максимума вертикального потока в летне-осенний период типичен для океанических районов Норвежского моря и обусловлен, главным образом, массовым осаждением птеропод Limacina retroversa [19, 39]. В районе плато Воринг (Норвежское море) поток этих организмов достигал в сентябре 8000 экз/м2/сут [19]. По расчетам [17] их вклад в годовой поток CaCO3 составлял 15%. В Лофотенской котловине максимальный поток лимацин в августе–октябре 1983 г. составлял 150–550 экз/м2/сут или 8.4–18.4 мгCaCO3/м2/сут, а их вклад в поток СаСО3 в этот период – 15–24%, или 11.5% годового потока [39]. По нашим данным, максимальные величины потока Limacina retroversa (13 000–15 000 экз/м2/сут или 60–100% потока CaCO3) также наблюдались в сентябре–октябре, а их вклад в годовой поток СаСО3 составлял 44 и 15% на глубинах 550 и 2950 м соответственно. Вклад лимацин в годовой поток Сорг (11–18%) сопоставим с оценками (8% [17]), полученными в районе плато Воринг. Согласно [39], ведущая роль птеропод в формировании потока биогенного материала в осенний период обусловлена массовым отмиранием молоди, появляющейся в Норвежском море в августе, и высокой скоростью оседания особей (1–2.5 см/с [26]). При такой скорости оседания отмершие лимацины могут достигать дна всего за несколько суток. Неудивительно, что по нашим данным, их количество в ловушках подповерхностного и придонного горизонтов практически одинаково. Очевидно, что и в настоящее время птероподы играют заметную роль в вертикальном переносе Сорг и карбоната кальция в Лофотенской котловине. Эти результаты подтверждают выводы [17, 19, 20, 25] о значительной роли птеропод в функционировании биологического углеродного насоса и формировании карбонатной системы в Норвежском море и в отдельных районах арктического бассейна.
Планктонные фораминиферы, являясь постоянным и многочисленным компонентом осадочного вещества, играют важную роль в вертикальном переносе СаСО3 [29, 33, 53]. Годовой поток CaCO3 фораминифер в разных районах Мирового океана составляет 0.2–22 гCaCO3/м2, а вклад в общий поток СаСО3 – 2–100% [53]. Наши оценки (0.3 гCaCO3/м2/год, 3% годового потока) приближаются к минимальным значениям, характерным для больших глубин океана. Вместе с тем, величины потоков в разные сезоны (0.1–5.6 мгCaCO3/м2/сут), полученные в настоящей работе, сопоставимы с величинами, рассчитанными [53] для Гренландского и Норвежского морей (0.5–6.3 мгCaCO3/м2/сут) на глубинах больше 500 м, а их вклад в поток СаСО3 в отдельные месяцы достигал 60%.
С 90-х гг. прошлого века отмечается общая тенденция увеличения вклада кальцифицирующего планктона в вертикальный поток осадочного вещества в Северной Атлантике и, наоборот, уменьшения роли диатомей и другого кремнистого планктона [14]. Высокая растворимость арагонитовых раковинок птеропод на больших глубинах по сравнению с кальцитовыми раковинами фораминифер и оболочками кокколитофорид [20, 40] объясняет отсутствие ископаемых остатков птеропод в донных осадках Лофотенской котловины. В донных осадках субполярной Северной Атлантики повсеместно распространены раковины фораминифер и оболочки кокколитофорид [5]. Согласно нашим данным верхний слой донных осадков Лофотенской котловины представлен преимущественно кокколитовыми пелитовыми илами. Аналогичные данные о составе донных осадков в Лофотенской котловине приведены в работе [52]. Доминирование кокколитофорид в осадках обусловлено высокой численностью осаждающихся оболочек этих водорослей. Вертикальный поток кокколитофорид в Лофотенской котловине по оценкам [52] достигал более 100 × × 106 экз/м2/сут. Роль фораминифер в вертикальном потоке и в формировании осадков не столь значительна.
Важнейшим компонентом вертикального потока Сорг являются ФП зоопланктона [58]. По современным оценкам вклад ФП в разных районах Мирового океана составляет от 1 до >90% потока Сорг [58]. В последние годы показано, что значительная часть ФП, продуцируемых в верхнем продуктивном слое, реминерализируется и не достигает глубоких слоев [28, 43, 47, 63]. Накопление ФП в промежуточных и глубоких слоях связано либо с питанием глубоководных видов зоопланктона [36], либо с суточными вертикальными миграциями зоопланктеров [28]. В пеллетном материале ловушек в Лофотенской котловине мы не обнаружили ФП копепод. Незначительный вклад ФП копепод в поток Сорг глубже 500 м в Норвежском море отмечался также в работах [18, 22]. В большинстве проб преобладали овальные ФП. Значительное количество овальных пеллет в материале седиментационных ловушек, установленных в районе плато Воринга в Норвежском море, также отмечалось в работе [19]. По мнению этих авторов такие пеллеты, вероятнее всего, продуцируются планктонными остракодами, численность которых в глубоководных слоях в Норвежском море достаточно велика. Интересно, что по нашим данным, сезонная изменчивость потока овальных ФП в слое 550–2950 м выражена меньше, чем в верхнем 500 м слое. Годовой поток Сорг ФП в исследуемом районе увеличивался с глубиной с 0.13 гСорг/м2 (500 м) до 0.40 гСорг/м2 (2950 м), а их вклад составлял 16–21% годового потока. Эти результаты согласуются с представлениями о том, что в промежуточных и глубоких слоях основную роль в вертикальном транспорте Сорг играют ФП, продуцируемые глубоководными видами зоопланктона.
Исследования потока НЗ в разных районах Мирового, обобщенные в обзоре [57] свидетельствуют о том, что НЗ составляет заметную часть потока Сорг и может играть ключевую роль в вертикальном потоке углерода прибрежных областей и над континентальным шельфом вне периода “цветения” фитопланктона. В Норвежском море исследований подобного рода не проводилось. По нашим оценкам поток Сорг НЗ в Лофотенской котловине изменялся от 0 до 0.45 мг гСорг/м2/м2/сут, с максимумом на глубине 550 м в январе. В это время поток НЗ составлял 52% общего потока Сорг (поток Сорг < 1 мм + поток Сорг НЗ). Значительное увеличение потока НЗ в зимний период было также показано для моря Бофорта и залива Амундсена [50, 51]. По данным этих авторов, на фоне снижения общего потока Сорг в этот сезон, вклад НЗ достигал 86–91%. Годовой поток Сорг НЗ в море Бофорта в верхнем 70 м слое (2.5 г гСорг/м2/м2) более чем в 20 раз выше величины потока, рассчитанного на основании наших данных для слоя 0–550 м (0.11 гСорг/м2). В придонном слое годовой поток Сорг НЗ снижался на порядок до 0.01 г гСорг/м2. Заметно различался и состав НЗ в зависимости от глубины постановки седиментационных ловушек. На глубинах 70 м в море Бофорта и 112 м в заливе Амундсена в потоке НЗ по численности доминировали старшие возрастные стадии Calanus glacialis [50, 51], так же как и на глубинах 50–60 и 130–140 м в Карском море [4]. В Лофотенской котловине на больших глубинах среди НЗ особи доминирующего по биомассе в зоопланктоне Норвежского моря Calanus finmarchicus [27, 45], были встречены в небольшом количестве (3 экз.) только в одной пробе на глубине 550 м. Отсутствие трупов калянуса в материале ловушек на протяжении всего года, вероятнее всего, связано с особенностями вертикального распределения этого вида на разных этапах его жизненного цикла. В весенне-летний период, когда популяция C. finmarchicus в Норвежском море концентрируется в верхнем 100 м слое [16], поток трупов этого вида не достигает мезопелагиали. С июля по февраль большая часть популяции населяет глубинные слои, где зимует в состоянии диапаузы [16]. По данным [32] зимующая часть популяции наиболее многочисленна в слое 600–1300 м, т.е. глубже горизонта постановки верхней ловушки. Очевидно, что и в этот период отмершие особи C. finmarchicus не оседают на дно, а, вероятнее всего, разлагаются в процессе осаждения под действием бактерий или потребляются глубоководными зоопланктерами. Можно предположить, что незначительный поток трупов калянуса – одна из причин невысоких значений потока НЗ в глубоководных районах Норвежского моря по сравнению с оценками, полученными в других Арктических морях на основании данных мелководных седиментационных ловушек. Низкие величины потока НЗ в придонном слое Лофотенской котловины свидетельствуют о незначительной роли НЗ в качестве источника органического вещества для глубоководных бентосных сообществ.
ЗАКЛЮЧЕНИЕ
Анализ результатов наших исследований и данных, полученных в 90-е годы прошлого столетия, показал, что наблюдаемые климатические изменения (повышение температуры и солености АВ, увеличение толщины слоя АВ) в Лофотенской котловине Норвежского моря не привели к кардинальным изменениям количественных характеристик процесса седиментации. В целом, характер и масштаб сезонных изменений вертикального потока осадочного вещества и его основных компонентов в современных условиях аналогичен описанному в работах тремя десятилетиями ранее. Отмеченные различия в величинах валового потока и потока Сорг в верхнем 500 м слое могут быть проявлением межгодовой изменчивости, либо следствием наблюдаемого в период с 1993 по 2017 г. уменьшения притока АВ в Норвежское море. Данные по составу компонентов планктонного происхождения в осадочном материале и их вкладу в поток Сорг и CaCO3 во многом согласуются с результатами, полученными ранее. Так же как в предыдущих исследованиях, заметную роль в вертикальном переносе Сорг и карбоната кальция в Лофотенской котловине играют птероподы Limacina retroversa. Отмирая, эти планктонные животные, во многом определяют цикл углерода в водной толще, но не участвуют непосредственно в формировании осадков. Преобладание в материале наших ловушек ФП остракод, населяющих преимущественно глубинные слои, отмечалось и в исследованиях, выполненных в 90-х годах.
Полученные впервые в Норвежском море оценки потока НЗ свидетельствуют о незначительной роли этого компонента осадочного вещества в вертикальном транспорте Сорг в глубинные слои. Одной из причин низких величин потока НЗ по сравнению с другими Арктическими морями может быть отсутствие в осадочном материале трупов доминирующего в зоопланктоне вида копепод Calanus finmarchicus, что, вероятнее всего связано с особенностями вертикального распределения этого вида на разных этапах жизненного цикла.
Результаты проведенного исследования показывают, что процесс седиментации в глубоководных районах Норвежского моря в значительной степени контролируется зоопланктоном. Суммарный вклад всех исследованных компонентов планктонного происхождения (птероподы, фораминиферы, яйца, ФП и НЗ) в годовой поток карбоната кальция составлял 16–47%, в годовой поток Сорг – 37–45%. В придонном слое существенно возрастает роль процессов ресуспензии и переотложения вещества верхнего слоя донных осадков.
Благодарности. Авторы благодарят экипаж НИС “Академик Мстислав Келдыш”, А.И. Коченкову, Е.О. Золотых, А.В. Булохова и Г.В. Малафеева за помощь в сборе и обработке проб ловушечного вещества, Д.В. Ерошенко и С.М. Исаченко за помощь при отборе проб осадков.
Источники финансирования. Экспедиционные исследования выполнены при поддержке госбюджета (финансирование морских экспедиций) и РНФ (проект № 14-50-00095). Геохимические и биологические исследования поддержаны РФФИ (проекты № 18-05-60069 Арктика, № 19-05-00787 и № 19-05-00022). Обработка материала частично выполнена в рамках государственного задания Минобрнауки России (тема № 0149-2019-0007). Анализ результатов и подготовка публикации выполнены при поддержке РНФ (проект № 20-17-00157).
Список литературы
Белоненко Т.В., Колдунов А.В., Сентябов Е.В., Карсаков А.Л. Термохалинная структура Лофотенского вихря Норвежского моря на основе экспедиционных исследований и по данным гидродинамического моделирования // Вестник СПбГУ. Науки о Земле. 2018. Т. 63. Вып. 4. С. 502–519.
Виноградов М.Е., Лисицын А.П. Глобальные закономерности распределения жизни в океане и их отражение в составе донных осадков. Закономерности распределения планктона и бентоса в океане // Изв. АН СССР. Сер. геол. 1981. № 3. С. 5–25.
Гельман Е.М., Старобина И.З. Фотометрические методы определения породообразующих элементов в рудах, горных породах и минералах. ГЕОХИ АН СССР, Центральная лаборатория, Сектор химических методов анализа. М.: ГЕОХИ АН СССР, 1976. 69 с.
Дриц А.В., Кравчишина М.Д., Пастернак А.Ф. и др. Роль зоопланктона в вертикальном потоке вещества в Карском море и море Лаптевых в осенний сезон // Океанология. 2017. Т. 57. № 6. С. 934–948.
Емельянов Е.М. Седиментогенез в бассейне Атлантического океана. М: Наука, 1982. 190 с.
Кравчишина М.Д., Новигатский А.Н., Саввичев А.С. и др. Исследование седиментосистем Баренцева моря и Норвежско-Гренландского бассейна в 68-м рейсе научно-исследовательского судна “Академик Мстислав Келдыш” // Океанология. 2019. Т. 59. № 1. С. 173–176.
Лисицын А.П. Процессы океанской седиментации. Литология и геохимия. М.: Наука, 1978. 392 с.
Лисицын А.П. Современные представления об осадкообразовании в океанах и морях. Океан как природный самописец взаимодействия геосфер Земли // Мировой океан. Т. II. Физика, химия и биология океана. Осадкообразование в океане и взаимодействие геосфер Земли / Под общ. ред. Лобковского Л.И. и Нигматулина Р.И. М.: Научный мир, 2014. С. 331–571.
Лисицын А.П., Виноградов Е.М. Глобальные закономерности распределения жизни в океане и их отражение в составе осадков: Образование и распределение биогенных осадков // Изв. АН СССР. Сер. геол. 1982. № 4. С. 5–24.
Лисицын А.П., Лукашин В.Н., Новигатский А.Н. и др. Глубоководные обсерватории на транскаспийском разрезе – непрерывные исследования потоков рассеянного осадочного вещества // Докл. РАН. 2014а. Т. 456. № 4. С. 485–489.
Лисицын А.П., Новигатский А.Н., Шевченко В.П. и др. Рассеянные формы осадочного вещества и их потоки в океанах и морях на примере Белого моря (результаты 12 лет исследований) // Докл. РАН. 2014б. Т. 456. № 3. С. 355–359.
Лисицын А.П., Петелин В.П. К методике предварительной обработки проб морских осадков в судовых условиях // Тр. Института океанологии. 1956. Т. 19. С. 240–251.
Новигатский А.Н., Гладышев С.В., Клювиткин А.А. и др. Мультидисциплинарные исследования в С-еверной Атлантике и прилегающей Арктике в 71-м рейсе научно-исследовательского судна “Академик Мстислав Келдыш” // Океанология. 2019. Т. 59. № 3. С. 510–512.
Antia A., Koeve N., Fischer W. et al. Basin-wide particulate carbon flux in the Atlantic Ocean: regional export patterns and potential for atmospheric CO2 sequestration // Global Biogeochem. Cycles. 2001. V. 15. P. 845–862.
Arar E.J., Collins G.B. Method 445.0. In vitro determination of chlorophyll “a” and pheophytin “a” in marine and freshwater algae by fluorescence. Revision 1.2. Cincinnati: U.S. Environmental Protection Agency, 1997. 22 p.
Arashkevich E.G., Tande K.S., Pasternak A.F., Ellertsen B. Seasonal moulting patterns and the generation cycle of Calanus finmarchicus in the NE Norwegian Sea, as inferred from gnathobase structures, and the size of gonads and oil sacs // Marine Biology. 2004. V. 146. № 1. P. 119–132.
Bathmann U.V., Noji T.T., Bodungen B. Sedimentation of pteropods in the Norwegian Sea in autumn // Deep–Sea Res. Part A. Oceanographic Research Papers. 1991. V. 38. № 10. P. 1341–1360.
Bathmann U.V., Noji T.T., Voss M., Peinert R. Copepod fecal pellets: abundance, sedimentation and content at a permanent station in the Norwegian Sea in May/June 1986 // Marine Ecology Progress Series. 1987. V. 38. P. 45–51.
Bathmann U.V., Peinert R., Noji T.T., Bodungen B.V. Pelagic origin and fate of sedimenting particles in the Norwegian Sea // Progress in Oceanography. 1990. V. 24. № 1–4. P. 117–125.
Bauerfeind E., Nöthig E.-M., Pauls B. et al. Variability in pteropod sedimentation and corresponding aragonite flux at the Arctic deep-sea long-term observatory HAUSGARTEN in the eastern Fram Strait from 2000 to 2009 // J. Mar. Systems. 2014. V. 132. P. 95–105.
Blindheim J., Rey F. Water-mass formation and distribution in the Nordic Seas during the 1990s // ICES J. of Marine Science. 2004. 61. P. 846–863.
Bodungen B., Antia A., Bauerfeind E. et al. Pelagic processes and vertical flux of particles: an overview of a long-term comparative study in the Norwegian Sea and Greenland Sea // Geologische Rundschau. 1995. 84. P. 11–27.
Bodungen B., Bathmann U., Voss M., Wunsch M. Vertical particle flux in the Norwegian Sea – resuspension and interannual variability // Sediment trap studies in the Nordic countries: Proceedings of the Symposium on Sediment Traps in Marine Ecological Research and Monitoring, Fiskebäckskil, Sweden, 1990 / Eds. Wassmann P. et al. 1991. V. 2. P. 116–136.
Broomé S., Chafik L., Nilsson J. Mechanisms of the time-varying sea surface height and heat content trends in the eastern Nordic Seas // Ocean Sci. Discussions. 2019. In review.https://doi.org/10.5194/os-2019-109
Busch K., Bauerfeind E., Nöthig E.-M. Pteropod sedimentation patterns in different water depths observed with moored sediment traps over a 4-year period at the LTER station HAUSGARTEN in eastern Fram Strait // Polar Biology. 2015. V. 38. № 6. P. 845–859.
Byrne R.H., Acker J.G., Betzer P.R., Feely R.A., Cates M.H. Water column dissolution of aragonite in the Pacific Ocean // Nature. 1984. V. 312. № 5992. P. 321–326.
Carstensen J., Weydmann A., Olszewska A., Kwaśniewski S. Effects of environmental conditions on the biomass of Calanus spp. in the Nordic Seas // J. of Plankton Res. 2012. V. 34. № 11. P. 951–966.
Cavan E.L., Le Moigne F.A., Poulton A.J. et al. Attenuation of particulate organic carbon flux in the Scotia Sea, Southern Ocean, is controlled by zooplankton fecal pellets // Geophys. Res. Lett. 2015. V. 42. № 3. P. 821–830.
Chernihovsky N., Torfstein A., Almogi-Labin A. Seasonal flux patterns of planktonic foraminifera in a deep, oligotrophic, marginal sea: Sediment trap time series from the Gulf of Aqaba, northern Red Sea // Deep Sea Research Part I: Oceanographic Research Papers. 2018. V. 140. P. 78–94.
Dall’Olmo G., Mork K.A. Carbon export by small particles in the Norwegian Sea // Geophys. Res. Lett. 2014. V. 41. P. 2921–2927.
Dugstad J., Fer I., LaCasce J. et al. Lateral heat transport in the Lofoten Basin: near-surface pathways and subsurface exchange // J. of Geophys. Res.: Oceans. 2019. V. 124. № 5. P. 2992–3006.
Edvardsen A., Pedersen J.M., Slagstad D. et al. Distribution of overwintering Calanus in the north Norwegian Sea // Ocean. Sci. 2006. V. 2. P. 87–96.
Eguchi N.O., Kawahata H., Taira A. Seasonal response of planktonic foraminifera to surface ocean condition: Sediment trap results from the central North Pacific Ocean // J.l of Oceanography. 1999. V. 55. № 6. P. 681–691.
Eldevik T., Risebrobakken B., Bjune A. et al. A brief history of climate e the northern seas from the Last Glacial Maximum to global warming // Quaternary Science Reviews. 2014. V. 106. P. 225–246.
Honjo S., Manganini S. J., Wefer G. Annual particle flux and a winter outburst of sedimentation in the northern Norwegian Sea // Deep - Sea Res. Part A. Oceanographic Research Papers. 1988. V. 35. № 8. P. 1223–1234.
Honjo S., Manganini S.J., Krishfield R.A., Francois R. Particulate organic carbon fluxes to the ocean interior and factors controlling the biological pump: A synthesis of global sediment trap programs since 1983 // Progress in Oceanography. 2008. V. 76. P. 217–285.
Hoving H-J.T., Perez J.A.A., Bolstad K.S.R. The Study of Deep-Sea Cephalopods // Advances in Marine Biology. 2014. V. 67 P. 235–359.
Lind S., Ingvaldsen R.B., Furevik T. Arctic warming hotspot in the northern Barents Sea linked to declining sea-ice import // Nature Climate Change. 2018. V. 8. P. 634–639.
Meinecke G., Wefer G. Seasonal pteropod sedimentation in the Norwegian Sea // Palaeogeography, Palaeoclimatology, Palaeoecology. 1990. V. 79. № 1–2. P. 129–147.
Mucci A. The solubility of Calcite and Aragonite in seawater at various salinities, temperatures, and one atmosphere total pressure // Am. J. Sci. 1983. V. 283 (7). P. 780–799.
Nilsen J.E.Ø., Falck E. Variations of mixed layer properties in the Norwegian Sea for the period 1948–1999 // Progress in Oceanography. 2006. V. 70. № 1. P. 58–90.
Norwegian Sea ecoregion – Ecosystem overview // ICES Ecosystem Overviews. Norwegian Sea ecoregion. 2017. P. 1–15.
Olli K., Wexels Riser C.W., Wassmann P. et al. Seasonal variation in vertical flux of biogenic matter in the marginal ice zone and the central Barents Sea // J. of Marine Systems. 2002. V. 38. № 1–2. P. 189–204.
Peinert R., Antia A., Bauerfeind E. et al. Particle flux variability in the Polar and Atlantic Biogeochemical Provinces of the Nordic Seas // The Northern North Atlantic: A Changing Environment / Eds. Schafer P. et al. Berlin: Springer. 2001. P. 53–68.
Planque B., Batten S.D. Calanus finmarchicus in the North Atlantic: the year of Calanus in the context of interdecadal change // ICES J. of Marine Science. 2000. M. 57. № 6. P. 1528–1535.
Polyakov I.V., Pnyushkov A.V., Alkire M.B. et al. Greater role for Atlantic inflows on sea-ice loss in the Eurasian Basin of the Arctic Ocean // Science. 2017. V. 365. Iss-ue 6335. P. 285–291.
Reigstad M., Wexels Riser C.W., Wassmann P., Ratkova T. Vertical export of particulate organic carbon: attenuation, composition and loss rates in the northern Barents Sea // Deep–Sea Res. Part II: Topical Studies in Oceanography. 2008. V. 55. № 20–21. P. 2308–2319.
Rossby T., Prater M. D., Søiland H. Pathways of inflow and dispersion of warm waters in the Nordic seas // J. Geophys. Res. 2009. V. 114. P. C04011.
Rudnick R. L., Gao S. Composition of the continental crust // Treatise on geochemistry. 2014. V. 4. P. 1–51.
Sampei M., Sasaki H., Hattori H. et al. Significant contribution of passively sinking copepods to the downward export flux in Arctic waters // Limnology and Oceanography. 2009. V. 54. P. 1894–1900.
Sampei M., Sasaki H., Forest A., Fortier L. A substantial export flux of particulate organic carbon linked to sinking dead copepods during winter 2007–2008 in the Amundsen Gulf (southeastern Beaufort Sea, Arctic Ocean) // Limnology and oceanography. 2012. V. 57. № 1. P. 90–96.
Samtleben C., Bickert T. Coccoliths in sediment traps from the Norwegian Sea // Marine Micropaleontology. 1990. V. 16. № 1–2. P. 39–64.
Schiebel R. Planktic foraminiferal sedimentation and the marine calcite budget // Global Biogeochemical Cycles. 2002. V. 16. № 4. P. 13-1–13-21.
SeaWiFS Project, NASA/Goddard Space Flight Center and ORBIMAGE. http://oceancolor.gsfc.nasa.gov
Segtnan O.H., Furevik T., Jenkins A.D. Heat and freshwater budgets of the Nordic seas computed from atmospheric reanalysis and ocean observations // J. of Geophys. Res. Oceans. 2011. V. 116. № C11003.
Skagseth Ø., Mork K.A. Heat content in the Norwegian Sea, 1995–2010 // ICES J. of Marine Science. 2012. V. 69. № 5. P. 826–832.
Tang K.W., Elliott D.T. Copepod carcasses: occurrence, fate and ecological importance // Copepods: Diversity, Habitat and Behavior / Ed. Seuront L. Hauppauge: Nova Science Publishers, 2014. P. 255–278.
Turner J.T. Zooplankton fecal pellets, marine snow, phytodetritus and the ocean’s biological pump // Progress in Oceanography. 2015. V. 130. P. 205–248.
Volkov D.L., Belonenko T.V., Foux V.R. Puzzling over the dynamics of the Lofoten Basin – a sub-Arctic hot spot of ocean variability // Geophys. Res. Letters. 2013. V. 40. № 4. P. 738–743.
Wassmann P., Peinert R., Smetacek V. Patterns of production and sedimentation in the boreal and polar Northeast Atlantic // Polar Res. 1991. V. 10. № 1. P. 209–228.
Wedepohl K.H. The composition of the continental crust // Geochimica et Cosmochimica Acta. 1995. V. 59. № 7. P. 1217–1232.
Weidberg N., Basedow S.L. Long-term variability in overwintering copepod populations in the Lofoten Basin: The role of the North Atlantic oscillation and trophic effects // Limnology and Oceanography. 2019. V. 64. № 5. P. 2044–2058.
Wexels Riser C.W., Wassmann P., Olli K. et al. Seasonal variation in production, retention and export of zooplankton faecal pellets in the marginal ice zone and central Barents Sea // J. of Marine Systems. 2002. V. 38. № 1–2. P. 175–188.
Дополнительные материалы отсутствуют.