

Океанология, 2020, T. 60, № 6, стр. 858-875
Структура фитоценозов Енисейского эстуария и прилежащего Карского шельфа в поздневесенний период
И. Н. Суханова 1, М. В. Флинт 1, *, Е. Г. Сахарова 2, А. В. Федоров 3, П. Н. Маккавеев 1, А. А. Недоспасов 1
1 Институт океанологии им. П.П. Ширшова РАН
Москва, Россия
2 Институт биологии внутренних вод им. И.Д. Папанина РАН
Борок, Ярославской обл., Россия
3 Южное отделение Института океанологии им. П.П. Ширшова РАН
Геленджик, Россия
* E-mail: m_flint@ocean.ru
Поступила в редакцию 27.12.2019
После доработки 27.03.2020
Принята к публикации 15.08.2020
Аннотация
Исследования выполнены на материале, собранном в 66-м рейсе НИС “Академик Мстислав Келдыш” в области Енисейского эстуария и на прилежащем Карском шельфе в широтном диапазоне от 71°50.5′ до 75°55.0′ с.ш. Наблюдения проведены через 3 недели после окончания половодья с 25 по 28 июля 2016 г. Зарегистрировано около 250 видов планктонных водорослей. Установлено существование выраженной широтной зональности в структуре сообществ фитопланктона, связанной с изменениями гидрофизических и гидрохимических условий пелагической среды. Наибольшее видовое разнообразие в южном опресненном (<1 PSU) районе эстуария формировали диатомовые, зеленые и синезеленые водоросли, в мористых районах – диатомеи и динофлагелляты. Самые высокие значения численности и биомассы водорослей связаны с внутренней опресненной областью эстуария – 2.7–2.9 × 106 кл/л и 1.1–1.5 г/м3 соответственно, где количественно преобладали пресноводные диатомовые рода Aulacoseira. Распространение пресноводных видов водорослей на север было ограничено соленостью 12–15 PSU (74°20′ c.ш.). Наибольшее снижение численности и биомассы фитопланктона наблюдалось во внешней части эстуария севернее ~73° с.ш. На срединном шельфе севернее эстуария Енисея максимальные численность (до 740 × 103 кл/л) и биомасса (до 240 мг/м3) фитопланктона были приурочены к глубинам 30–40 м при низкой освещенности и высоких концентрациях биогенных элементов, что было типично и для Обского района в весенне-летний сезон.
Эстуарные области крупных арктических рек, где происходит интенсивное взаимодействие пресноводного континентального стока и морских вод, играют огромную роль в трансформации и осаждении приносимого речным стоком аллохтонного материала, регулировании воздействия континентальных процессов на морские экосистемы и формировании биологической продукции [5, 8, 9, 11, 15–17, 19, 21, 22, 24–26]. Смешение морских и речных вод в эстуарных районах порождает мощнейшую эстуарную фронтальную зону (ЭФЗ), внутри которой формируются специфические высокоградиентные по горизонтали и вертикали условия пелагической среды [2, 6, 8, 10, 19, 25]. Уникальное пространственное биотопическое разнообразие в эстуариях и на прилежащем шельфе определяет важнейшие для эпиконтинентальных морей Арктики биологические процессы и четко выраженную широтную (кросс-шельфовую) зональность в структуре пелагических экосистем [5, 14–17, 19, 22, 23]. Эта широтная зональность во многом отражает влияние пресноводного стока на эстуарные экосистемы и прилежащие шельфовые районы морей, коренным образом влияет на биологическую продуктивность, горизонтальные и вертикальные потоки вещества. ЭФЗ формируют южное звено макромасштабной широтной зональности в структуре пелагических экосистем, которая во многом определяет процессы взаимодействия в глобальной системе Сибирский континент – Арктический бассейн. Роль ЭФЗ в формировании широтной зональности и отдельных ее элементов показана для целого ряда высокоширотных морских районов [1, 15, 28, 30], в том числе и для Карского моря.
Процессы в крупных арктических эстуариях находятся под воздействием климатических факторов, определяющих объемы и режим пресноводного стока, а также факторов антропогенной природы, тем или иным путем влияющих на химический состав и загрязнение речных вод. Таким образом, долговременная изменчивость явлений в экосистемах эстуарных зон крупных арктических рек может служить показателем климатической и антропогенной изменчивости процессов взаимодействия в системе река–море. В сильнейшей степени влияют на эстуарные экосистемы и сезонные процессы, меняя физические и химические свойства среды, состав пелагической биоты, ее распределение и продуктивность [15, 16, 18, 23].
Карское море принимает в себя крупнейший в Арктике годовой речной сток, объем которого в среднем составляет около 1200 км3/год и условно увеличивает уровень бассейна на 1 м при его средней глубине 111 м [27, 31, 33]. Эта величина почти на два порядка выше, чем в среднем для Мирового океана, что говорит о принципиальной важности процессов, связанных с взаимодействием речных и морских вод в эстуариях двух Сибирских рек – Оби и Енисея, определяющих более 95% поступления пресных вод, для функционирования природной системы Карского бассейна.
При существующей исследованности экосистем эстуариев крупных Сибирских рек, впадающих в Карское море, и прилежащего к эстуариям арктического шельфа, весенние процессы в планктонных сообществах, связанные с максимальным сезонным стоком, остаются практически не изученными. Экспедиционные работы, выполненные во второй половине июля 2016 г. позволили впервые характеризовать состав, количественные параметры фитоценозов и зоопланктона и характер их пространственного распределения в связи с особенностями пелагической среды в эстуарии Оби и на прилежащем Карском шельфе в поздневесенний сезон [16, 23].
Енисей дает самый большой пресноводный сток в Арктику (в среднем 624 км3), при этом меридиональная протяженность его эстуария существенно меньше, чем Обского и составляет всего 224 км против ~800 км [11, 25]. Эстуарий Енисея существенно отличается от Обского и по конфигурации береговой линии, а сами реки разнятся по режиму и составу стока и характеру влияния на прилежащий Карский шельф [4, 10, 11, 25], а также структуре планктонных сообществ. Пик весеннего половодья в Енисее более поздний и более консолидированный во времени, чем в Оби [3, 11], что определяется особенностями водосборного бассейна. На июнь–начало июля приходится 36% годового объема стока, и уже в следующем месяце он падает до 12%. Именно половодье, когда в эстуарии поступает основные объемы пресной воды и аллохтонных материалов, включая биогенные элементы, и непосредственно следующий за половодьем период в значительной степени определяют важнейшие процессы в эстуарных планктонных сообществах и дальнейший ход сукцессии региональных фитоценозов в районах Карского шельфа, в сильной степени подверженных воздействию континентального стока.
Несмотря на огромное значение Енисея для функционирования природной системы Карского бассейна, планктонные сообщества Енисейского эстуария исследованы существенно меньше, чем Обского [9, 17, 22]. Немногие исследования фитоценозов Енисейского эстуария были сделаны в позднелетний и осенний сезоны при низкой интенсивности стока, а данные о структуре и продуктивности фитопланктонных сообществ в весенний сезон до настоящего времени отсутствовали.
Задачей настоящих исследований был анализ состава и пространственной структуры фитопланктонных сообществ эстуарной области Енисея и прилежащего Карского шельфа в поздневесенний сезон непосредственно за пиком весеннего половодья; оценка связей структурных характеристик фитоценозов с особенностями высокоградиентной среды в зоне взаимодействия енисейских и морских вод; выявление биологических явлений и экологических границ, ассоциированных с ЭФЗ и ее отдельными фронтальными разделами и оценка их пространственных масштабов; оценка роли ЭФЗ в формировании широтной зональности фитоценозов Карского бассейна.
МАТЕРИАЛ И МЕТОДЫ
Материал, положенный в основу настоящей работы, был получен на квазимеридиональном разрезе между 71°50′5 и 75°55′0 c.ш. протяженностью 526 км, охватывающем область с поверхностной соленостью от 0.437 PSU до 31.432 PSU (рис. 1, 2). Год выполнения исследований – 2016 г. – относится к маловодным – общий сток Енисея составлял 531 км3 при среднемноголетней величине 624 км3. Наши исследования выполнены с 25 по 28 июля 2016 г., практически через три недели после окончания мощного сезонного половодья, во время которого в течение месяца в эстуарий поступает более трети годового стока.
Рис. 1.
Район работ в Карском море (а) и схема расположения станций (б). ⚫ – гидрофизические, гидрохимические наблюдения и отбор проб фитопланктона; ⚪ – гидрофизические и гидрохимические наблюдения.
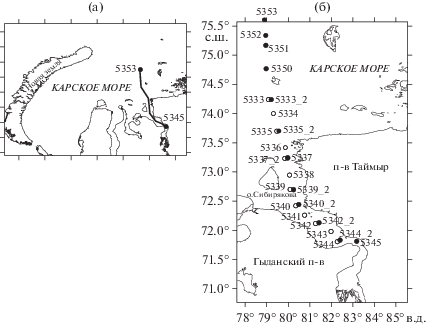
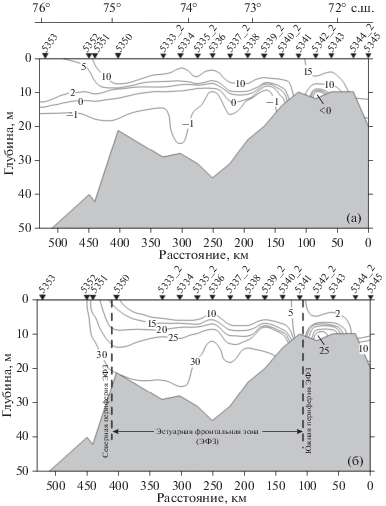
Рис. 2.
Распределение (а) – температуры (T°C) и (б) – солености (S, PSU), положение и границы эстуарной фронтальной зоны (ЭФЗ) на Енисейском разрезе в июле 2016 г.
Пробы фитопланктона и сопутствующие гидрофизические и гидрохимические материалы получены в ходе 66-го рейса НИС “Академик Мстислав Келдыш”, выполнявшегося в рамках многолетней программы “Морские экосистемы Сибирской Арктики”. На квазимеридиональном разрезе, упомянутом выше, было сделано 12 станций.
Пробы воды для анализа фитопланктона в объеме 2 л отбирали из 5-литровых батометров Нискина комплекса “Розетта” SBE 32. Выбор горизонтов отбора проб проводили на основании вертикального распределения температуры, солености и флуоресценции, полученного при предварительном зондировании CTD-зондом SeaBird-911, оснащенным датчиком флуоресценции WetLab ECO AFL-FL. Пробы отбирали с 3–5 горизонтов: в верхнем перемешанном слое (1–2 горизонта), в слое скачка плотности и/или максимума флюоресценции (1–2 горизонта) и в слое под пикноклином (1–2 горизонта). Всего было отобрано и обработано 46 проб фитопланктона. Пробы воды для анализа фитопланктона и гидрохимических определений, характеризующих абиотическую составляющую экосистем, брали из одних и тех же батометров.
Концентрирование фитопланктона в пробах перед обработкой проводили методом мягкой обратной фильтрации. Для этого два литра морской воды профильтровывали через лавсановые ядерные фильтры с размером ячеи 1 мкм на установках для обратной фильтрации [13]. Объем полученного концентрата составлял 40–90 мл. Все пробы были обработаны в живом состоянии без предварительной фиксации сразу же после отбора или в течение одного–двух дней. До обработки пробы хранили в холодильнике в темноте при температуре 2–3°С. Обработку проб вели в камерах Ножотта (объем 0.085 мл) при увеличении ×400 и Наумана (объем 1 мл) при увеличении ×200 с использованием микроскопов Jena Lumar, Leica и МБИ-3. Клетки были просчитаны в 1 или 2 камерах в зависимости от численности вида. Объем клеток (сырую биомассу) рассчитывали на основе их линейных размеров и принципа подобия соответствующим стереометрическим фигурам. Биомасса водорослей в углеродном эквиваленте была определена исходя из их объемов с использованием соответствующих коэффициентов для разных таксономических и размерных групп [29, 32]. Для оценки состояния популяций массовых видов диатомей нами были учтены мертвые клетки и клетки с видоизмененными хлоропластами.
РЕЗУЛЬТАТЫ
В исследованном районе нами было зарегистрировано около 250 видов планктонных водорослей. Наибольшим видовым разнообразием характеризовались диатомеи, динофлагелляты и в опресненном районе – диатомовые, зеленые и синезеленые водоросли. Небольшое число видов было отмечено в классах Prymnesiophyceae, Dictyophyceae, Prasinophyceae, Chrysophyceаe, Cryptophyceae. Часть водорослей была определена только до рода. Жгутиковые (диаметром 4–8 мкм) и трудно определяемые ювенильные стадии в жизненном цикле динофлагеллят были просчитаны, измерены и распределены по размерным группам для расчета общей биомассы фитопланктона. Определенную трудность представлял количественный учет синезеленых водорослей рода Anabaena из-за образования клубков спутанных нитей и ряда других родов из-за трудно различимых клеток в трихомах.
Следует отметить, что клетки многих синезеленых водорослей характеризуются присутствием газовых вакуолей, что определяет их плавучесть и преимущественное обитание в самом поверхностном слое моря. Верхний горизонт отбора проб, из-за конструкции пробоотборника, как правило, располагался на глубине 1–2 м. Это, вероятно, привело к недоучету синезеленых и, соответственно, занижению общей численности и биомассы фитопланктона на станциях разреза, где были встречены эти водоросли.
В опресненной части эстуария на станциях 5345, 5344_2 и 5342_2 в верхних 6–10 м соленость была <1 PSU, температура – 16–17°С, содержание нитратов и фосфатов не превышало 0.25 и 0.12 мкг-ат/л соответственно (рис. 2, 3а). Концентрация кремния была высокой – >70 мкг-ат/л (рис. 3б). Пресноводные диатомовые водоросли составляли >90% общей численности фитопланктона, при этом преобладали виды рода Aulacoseira (>80%). Кроме них многочисленными были диатомеи Asterionella formosa и Fragilaria crotonensis. От 2 до 5% численности составляли зеленые водоросли. В поверхностном слое наблюдались скопления сине-зеленых водорослей. Основные части популяций всех доминирующих видов находились в верхних 6–10 м. В этом же слое зарегистрированы максимальные для исследованного района численность и биомасса фитопланктона (2700–2850 × 103 кл/л и 1.1–1.5 г/м3, табл. 1, рис. 4) и содержание хлорофилла “а”: от 3.7 до 5.8 мг Сhl a/м3 [20].
Рис. 3.
Распределение нитратного азота – NO3 и кремния – Si (мкг-ат/л) на Енисейском разрезе в июле 2016 г.
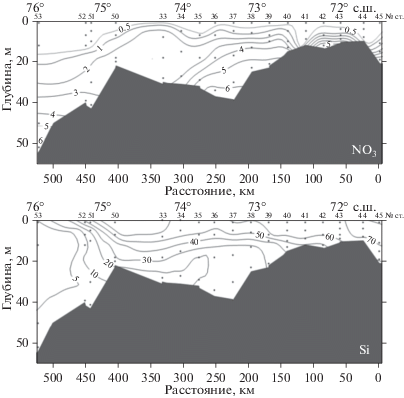
Таблица 1.
Численность (N × 103 кл/л), сырая биомасса (В, мг/м3), биомасса в углероде (ВС, мкг/л) всего фитопланктона и отдельных групп водорослей и их вклад в общую численность и биомассу (%)
| № станции слой, м |
Общая | Диатомеи | Динофлагеляты | Зеленые | Сине-зеленые | Флагелляты | Хризофиции | Криптофиции | Остальные | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 5345 | N; % | 2698 | 2607; | 96.6 | 0.4; | + | 85.0; | 3.2 | 1.2; | + | 4.1; | 0.1 | 0.3; | + | ||||
| B; % | 1515 | 1427; | 94.2 | 6.1; | 0.4 | 68.0; | 4.5 | 9.5; | 0.6 | 2.7; | 0.2 | 1.7; | 0.1 | |||||
| 0–20 | ВС; % | 218.2 | 206.0; | 94.4 | 0.3; | 0.1 | 10.2; | 4.7 | 1.0; | 0.5 | 0.5; | 0.2 | 0.2; | 0.1 | ||||
| 5344 | N; % | 2866 | 2757; | 96.2 | 0.5; | + | 52.5; | 1.8 | 28.9; | 1.0 | 21.5; | 0.7 | 5.6; | 0.2 | ||||
| B; % | 1073 | 1020; | 95.0 | 2.9; | 0.3 | 11.0; | 1.0 | 26.6; | 2.5 | 10.6; | 1.0 | 1.9; | 0.2 | |||||
| 0–10 | BС; % | 154.8 | 149.2; | 92.6 | 0.4; | 0.2 | 1.4; | 0.9 | 1.4; | 4.8 | 2.1; | 1.3 | 0.3; | 0.2 | ||||
| 5342 | N; % | 1014 | 941.0; | 93.0 | 1.1; | 0.1 | 52.0; | 4.9 | 11.5; | 1.1 | 8.4; | 0.9 | ||||||
| B; % | 424.7 | 389.4; | 91.9 | 7.0; | 1.6 | 14.1; | 3.2 | 12.6; | 2.8 | 1.6; | 0.5 | |||||||
| 0–11 | BС; % | 57.2 | 53.4; | 93.4 | 0.7; | 1.2 | 1.5; | 2.6 | 1.3; | 2.2 | 0.3; | 0.6 | ||||||
| 5340 | N; % | 293 | 230.8; | 78.9 | 6.6; | 2.2 | 34.1; | 11.6 | 12.2; | 4.2 | 7.0; | 2.4 | ||||||
| B; % | 191.7 | 123.5; | 64.4 | 43.0; | 22.4 | 1.6; | 0.8 | 9.4; | 4.9 | 2.3; | 1.2 | |||||||
| 0–9 | BС; % | 20.2 | 11.3; | 55.9 | 4.5; | 22.3 | 0.2; | 1.0 | 1.5; | 7.4 | 0.2; | 1.0 | 2.5; | 12.4 | ||||
| 5339 | N; % | 504 | 473; | 93.8 | 0.9; | 0.2 | 20.8; | 4.1 | 0.8; | 0.2 | 3.3; | 0.6 | 4.6; | 0.9 | 0.3; | + | ||
| B; % | 307.6 | 292.8; | 95.2 | 6.1; | 2.0 | 1.6; | 0.5 | 5.5; | 1.8 | 1.3; | 0.4 | + | + | 0.2; | 0.1 | |||
| 0–19 | BС; % | 39.5 | 37.6; | 95.2 | 0.7; | 1.8 | 0.2; | 0.6 | 0.6; | 1.5 | 0.3; | 0.7 | + | + | + | 0.1 | ||
| 5337 | N; % | 50.4 | 39.0; | 77.4 | 4.6; | 9.1 | 0.4; | 0.8 | 3.8; | 7.5 | 2.6; | 5.2 | ||||||
| B; % | 56.5 | 23.3; | 41.2 | 29.7; | 2.6 | 0.4; | 0.7 | 1.1; | 1.9 | 2.0; | 3.5 | |||||||
| 0–28 | BС; % | 7.0 | 3.3; | 47.1 | 2.9; | 41.4 | 0.1; | 1.4 | 0.5; | 7.1 | 0.2; | 2.9 | ||||||
| 5335 | N; % | 138.6 | 92.6; | 66.8 | 3.1; | 2.2 | 0.1; | 0.1 | 0.1; | 0.1 | 0.2; | 0.1 | 38.0; | 27.4 | 4.5; | 3.2 | ||
| B; % | 144.8 | 70.9; | 49.1 | 27.2; | 18.8 | + | + | 0.3; | 0.2 | 0.2; | 0.1 | 1.0; | 0.7 | 44.2; | 30.5 | |||
| 0–29 | BС; % | 19.0 | 9.4; | 49.5 | 2.6; | 13.7 | + | + | + | 0.1 | 0.3; | 3.2 | 0.1; | 0.5 | 6.5; | 34.2 | ||
| 5333 | N; % | 121.1 | 99.9; | 82.5 | 13.7; | 1.3 | 0.6; | 0.5 | 0.3; | 0.2 | 2.5; | 2.1 | 0.8; | 0.7 | 3.3; | 2.7 | ||
| B; % | 126.8 | 60.7; | 47.8 | 62.2; | 49.0 | 0.2; | 0.2 | 0.2; | 0.1 | 1.5; | 1.2 | 0.4; | 0.3 | 1.7; | 1.8 | |||
| 0–28 | BС; % | 15.0 | 7.4; | 49.4 | 6.9; | 46.2 | + | 0.1 | + | 0.1 | 0.3; | 2.0 | + | 0.2 | 0.3; | 2.0 | ||
| 5350 | N; % | 183.8 | 125.7; | 68.4 | 26.1; | 14.2 | 13.5; | 7.3 | 12.8; | 7.0 | 5.7; | 3.1 | ||||||
| B; % | 170.0 | 73.0; | 42.9 | 87.6; | 51.5 | 2.3; | 1.3 | 0.8; | 0.5 | 6.3; | 3.7 | |||||||
| 0–20 | BС; % | 18.0 | 8.5; | 47.2 | 8.5; | 47.2 | 0.2; | 1.1 | 0.1; | 0.5 | 0.7; | 3.9 | ||||||
| 5351 | N; % | 354.5 | 277.7; | 78.3 | 11.8; | 3.3 | 58.2; | 16.4 | 4.0; | 1.1 | 0.7; | 0.2 | 2.1; | 0.6 | ||||
| B; % | 106.8 | 68.1; | 63.7 | 31.0; | 29.0 | 5.8; | 5.4 | 0.3; | 0.3 | + | + | 1.6; | 1.5 | |||||
| 0–40 | BС; % | 12.9 | 8.3; | 64.4 | 3.6; | 27.9 | 0.7; | 5.4 | + | 0.2 | + | + | 0.2; | 1.5 | ||||
| 5352 | N; % | 257.0 | 235.2; | 91.5 | 2.7; | 1.0 | 0.6; | 0.2 | 14.9; | 5.8 | + | + | 3.5; | 1.4 | ||||
| B; % | 127.1 | 94.4; | 74.3 | 23.7; | 18.6 | 0.2; | 0.2 | 0.5; | 0.4 | + | + | 8.3; | 6.5 | |||||
| 0–38 | BС; % | 12.8 | 8.8; | 68.0 | 2.3; | 18.0 | + | + | 0.1; | 0.8 | + | + | 1.4 | 10.9 | ||||
| 5353 | N; % | 119.3 | 100.6; | 84.3 | 4.3; | 3.6 | 0.6; | 0.5 | 12.2; | 10.2 | 0.7; | 0.6 | 0.9; | 0.8 | ||||
| B; % | 62.7 | 43.8; | 69.3 | 16.5; | 26.3 | 0.1; | 0.2 | 1.3; | 2.5 | + | 0.1 | 1.0; | 1.6 | |||||
| 0–56 | BС % | 7.3 | 5.4; | 74.2 | 1.5; | 20.6 | + | 0.4 | 0.3; | 4.1 | + | + | + | 0.3 | ||||
Рис. 4.
Вертикальное распределение солености (S, PSU), нитратного азота (NO3, мкг-ат/л), флуоресценции (Fl), численности (N × 103 кл/л) и биомассы (B, мг/м3) фитопланктона на станциях Енисейского разреза в июле 2016 г.
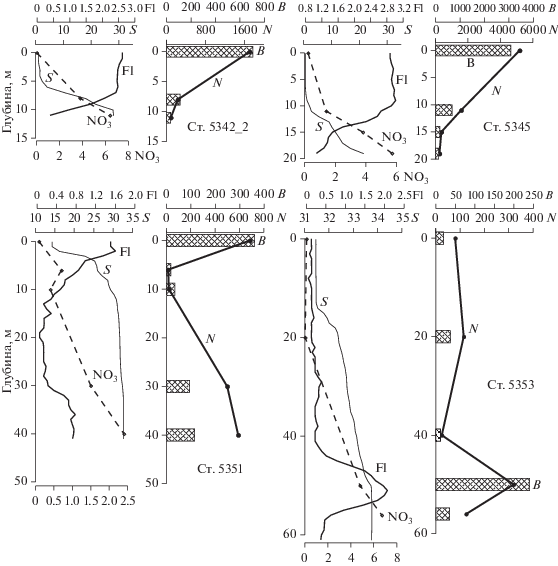
На самой южной ст. 5345 (рис. 1, глубина 21 м) ниже 10 м располагался пикноклин с градиентом солености 2.2 PSU/м (рис. 2). В пикноклине резко увеличилось содержание биогенных элементов (рис. 3, 4). Эти изменения не сказались на качественном составе фитопланктона, однако с глубиной существенно менялись численность, биомасса фитопланктона и концентрация хлорофилла “а” (рис. 4). На горизонте 11 м общее число клеток водорослей уменьшились в 3.3, биомасса – в 4.5 раза. На горизонтах 15 и 20 м изменения достигали порядка величин по сравнению с поверхностным слоем. Концентрация хлорофилла на 11 м снизилась в 2.5 раза по сравнению верхним слоем водной толщи, на 15 и 20 м – в 7.5 раз. На нижних горизонтах резко возрастало число мертвых клеток пресноводных диатомей, которые составляли от 82% на 15 м до 90% на 20 м от общего числа живых и мертвых клеток (табл. 2).
Таблица 2.
Общая численность (N × 103 кл/л) диатомовых водорослей, численность живых и мертвых клеток пресноводных и морских видов диатомовых, доля (%) живых и мертвых клеток на разных горизонтах
| № станции | Горизонт, м | N, общая живые и мертвые | N, живые клетки | %, живых кл. | N, мертвые клетки | %, мертвых кл. | ||
|---|---|---|---|---|---|---|---|---|
| пресно- водные |
морские | пресно- водные |
морские | |||||
| 5345 | 0 | 5502.2 | 5014.6 | – | 91.1 | 487.6 | – | 8.9 |
| 11 | 2386.6 | 1499.0 | – | 62.8 | 887.6 | – | 37.2 | |
| 15 | 1925.3 | 352.3 | – | 18.3 | 1573.0 | – | 81.7 | |
| 20 | 2318.4 | 235.3 | – | 10.1 | 2083.1 | – | 89.9 | |
| 5344 | 0 | 4450.2 | 3374.5 | – | 75.8 | 1075.7 | – | 24.2 |
| 5 | 4126.5 | 3376.0 | 5.7 | 81.9 | 750.4 | – | 18.2 | |
| 10 | 1130.1 | 892.6 | – | 79.0 | 237.5 | – | 21.0 | |
| 5342 | 0 | 2030.8 | 1569.2 | 3.9 | 77.7 | 452.7 | – | 22.3 |
| 8 | 278.4 | 211.9 | 12.3 | 80.5 | 54.2 | – | 19.5 | |
| 11 | 2696.0 | 86.4 | – | 3.2 | 2609.6 | – | 96.8 | |
| 5340 | 0 | 306.1 | 88.7 | 181.8 | 88.3 | 35.6 | – | 11.7 |
| 5 | 353.6 | 110.1 | 182.5 | 82.7 | 26.1 | 35.1 | 17.3 | |
| 9 | 523.2 | 35.2 | 27.3 | 11.9 | 459.4 | 1.3 | 88.1 | |
| 5339 | 0 | 1252.0 | 342.7 | 800.4 | 91.3 | 108.9 | – | 8.7 |
| 4 | 1510.0 | 433.6 | 858.5 | 85.6 | 211.3 | 6.6 | 14.4 | |
| 9 | 105.7 | 25.3 | 0.6 | 24.5 | 78.8 | 1.0 | 75.5 | |
| 19 | 179.5 | 7.8 | 5.2 | 7.2 | 166.5 | – | 92.8 | |
| 5337 | 0 | 173.5 | – | 126.6 | 73.0 | – | 46.9 | 27.0 |
| 17 | 35.4 | 2.9 | 2.2 | 14.4 | 26.7 | 3.6 | 85.6 | |
| 28 | 45..3 | 1.9 | – | 4.2 | 43.4 | – | 95.8 | |
| 5335 | 0 | 286.3 | 7.7 | 235.0 | 84.7 | – | 43.6 | 15.3 |
| 10 | 199.5 | 79.0 | 56.4 | 67.9 | 64.1 | – | 32.1 | |
| 20 | 21.7 | – | 2.6 | 12.0 | 6.1 | 13.0 | 88.0 | |
| 29 | 96.3 | 1.2 | 4.8 | 6.2 | 65.3 | 25.0 | 93.8 | |
| 5333 | 0 | 304.8 | – | 232.8 | 76.6 | – | 71.0 | 23.4 |
| 6 | 342.9 | 24.8 | 266.7 | 85.4 | 42.5 | 8.9 | 14.6 | |
| 12 | 40.7 | 4.1 | 17.9 | 54.0 | 18.7 | – | 46.0 | |
| 28 | 16.6 | 1.4 | – | 8.4 | 13.8 | 1.4 | 91.6 | |
| 5350 | 0 | 80.5 | – | 64.8 | 80.5 | – | 15.7 | 19.5 |
| 18 | 384.7 | – | 179.7 | 46.7 | – | 205.0 | 53.3 | |
| 20 | 352.2 | 6.8 | 213.0 | 64.4 | 40.5 | 91.9 | 35.6 | |
| 5351 | 0 | 90.7 | – | 88.2 | 97.2 | 2.5 | 2.8 | |
| 6 | 22.5 | 0.8 | 8.3 | 40.4 | 0.7 | 12.7 | 59.6 | |
| 10 | 46.5 | – | 17.2 | 36.9 | – | 29.3 | 63.1 | |
| 30 | 529.8 | 1.6 | 497.9 | 94.3 | 14.6 | 15.7 | 5.7 | |
| 40 | 684.3 | – | 588.6 | 86.0 | – | 95.7 | 14.0 | |
| 5352 | 0 | 152.4 | – | 148.4 | 97.4 | – | 4.0 | 2.6 |
| 7 | 32.7 | – | 27.0 | 82.6 | – | 5.7 | 17.4 | |
| 12–25 | 230.8 | 1.7 | 225.7 | 98.5 | 3.3 | 0.1 | 1.5 | |
| 38 | 818.3 | 0.1 | 726.4 | 88.8 | 0.2 | 91.6 | 11.2 | |
| 5353 | 0 | 80.5 | – | 78.1 | 97.0 | – | 2.4 | 3.0 |
| 20 | 140.0 | 7.8 | 106.2 | 81.5 | 26.0 | – | 18.5 | |
| 40 | 26.8 | – | 24.5 | 91.4 | – | 2.3 | 8.6 | |
| 52 | 229.6 | – | 214.7 | 94.8 | 0.5 | 14.4 | 6.5 | |
На ст. 5344_2 при глубине ~10 м гидрофизические и гидрохимические характеристики во всей толще воды практически не различались (рис. 2, 3), при этом количество фитопланктона существенно менялось с глубиной. На нижнем десятиметровом горизонте численность и биомасса фитопланктона сократились в 3.5 раза по сравнению с верхним слоем, концентрация хлорофилла – в полтора раза. Число мертвых клеток диатомовых на всех горизонтах колебалось от 18 до 24% от общего числа живых и мертвых клеток (табл. 2).
На более северной ст. 5342_2 (рис. 1, глубина 12 м) почти в 3 раза сократились общие численность и биомасса фитопланктона в столбе воды (табл. 1, рис. 5). Верхний однородный слой составлял 6 м, ниже – между 6 и 10 метрами лежал резкий пикноклин с градиентом солености 5.5 PSU/м и температуры 3°С/м (рис. 2, 4). В градиентном слое больше, чем на порядок, возрастала концентрация нитратов (рис. 3, 4), в несколько раз увеличилось содержание аммония и фосфатов. При этом численность фитопланктона на глубине 8 м была в 7.5 раз, биомасса 6.5 раз ниже, чем в верхнем перемешенном слое (соответственно, 1704 × 103 и 228 × 103 кл/л, 705 и 110 мг/м3), концентрация хлорофилла сократилась в 6 раз – с 2.6 до 0.4 мг Сhl a/м3. На глубине 11 м при солености 26 PSU и высоких концентрациях биогенных элементов численность и биомасса фитопланктона снижались более, чем на порядок по сравнению поверхностным слоем (рис. 4). Во всем столбе воды в фитоцене доминировали пресноводные виды диатомовых водорослей. Мертвые диатомеи в верхнем однородном слое составляли 22%, в градиентном слое и под пикноклином – 97%, от общего числа живых и мертвых клеток (табл. 2).
Рис. 5.
Распределение поверхностной и придонной солености (S, PSU), численности (N × 103 кл/л) и биомассы (В, мг/м3) фитопланктона на Енисейском разрезе в июле 2016 г. 1 – пресноводный фитопланктон; 2 – морской фитопланктон. N и В – средние для столба воды.
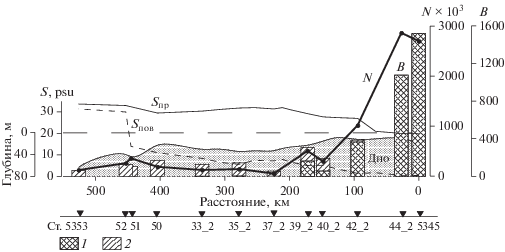
В целом во внутренней опресненной части эстуария на станциях 5345, 5344_2, 5342_2 зарегистрированы самые высокие для исследованного района численность и биомасса фитопланктона (табл. 1, рис. 5).
Севернее ст. 5342_2 во внешней области эстуария и на прилежащем шельфе (рис. 1, станции 5340_2 – 5351) при глубинах от 15 до 43 м водная толща характеризовалась наличием жесткого пикноклина с градиентами от 2.2 до 6.5 PSU/м (рис. 2, 4, 6). Концентрация нитратов в верхнем перемешенном слое, глубина которого составляла 4–8 м, была повсеместно низкой и увеличивалась в несколько раз в слое пикноклина и под ним (рис. 3). Концентрация кремния была высокой с максимумом до 50–60 мкг-ат/л в верхнем перемешенном слое (рис. 3), что вместе с низкой соленостью указывало на сильное действие речного стока.
Рис. 6.
Вертикальное распределение солености, нитратного азота, флуоресценции, численности и биомассы фитопланктона на станциях Енисейского разреза в июле 2016 и 2011 (ст. 5020) годов. Обозначения как на рис. 4.
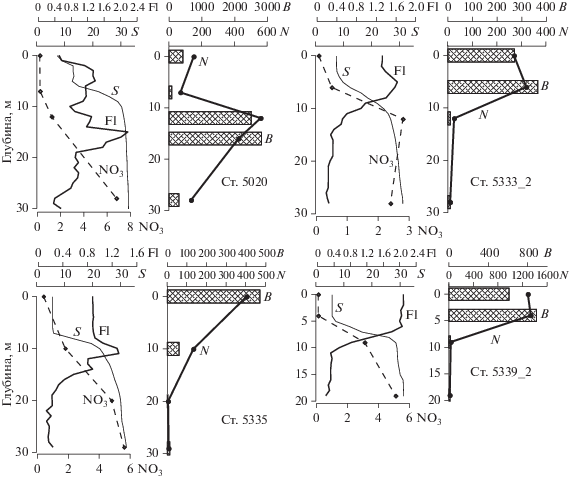
Выраженные изменения качественного состава и обилия водорослей по сравнению с внутренней частью эстуария наблюдались на ст. 5340_2 на 40 км севернее ст. 5342_2. Соленость в верхнем однородном шестиметровом слое возросла до 6 PSU, численность и биомасса фитопланктона в столбе воды снизились в несколько раз по сравнению со ст. 5342_2 (табл. 1, рис. 2, 5). В фитопланктоне верхнего однородного слоя (0–7 м), помимо пресноводного комплекса диатомей, зеленых и синезеленых водорослей, появились морские неритические виды. Диатомовые в общей численности фитопланктона верхнего однородного слоя составляли более 80%, из них на долю морских неритических видов, представленных, главным образом, Skeletonema costatum и Chaetoceros wighamii, приходилось ~55%. Около 50% биомассы фитопланктона создавали солоноватоводная Thalassiosira cf. вaltica и морские динофлагелляты: Peridinella catenata, Akashiwo sanquinea и Dinophysis acuminata. В слое пикноклина, где градиент солености достигал 5.2 PSU/м, и под пикноклином численно преобладали пресноводные виды (58%). Концентрация хлорофилла в верхнем перемешенном слое была ~2.5 мг Сhl a/м3, доля мертвых клеток диатомей составляла около 15% (табл. 2). Содержание хлорофилла под пикноклином снижалось до 0.9 мг Сhl a/м3, мертвые клетки составляли 88% от общего числа диатомей; на 97% это были пресноводные виды (табл. 2).
Распределение солености, температуры, пространственные изменения вертикальной структуры водной толщи (рис. 2) и изменения структуры фитоценоза (табл. 1, рис. 2, 5) позволяют определить область между станциями 5342_2 и 5340_2 как южную границу (южный фронтальный раздел) ЭФЗ.
На ст. 5339_2 в 26 км севернее ст. 5340_2 в верхнем однородном 7-метровом слое наблюдалось почти четырехкратное увеличение численности и биомассы фитопланктона, в полтора раза увеличилась концентрация хлорофилла. При этом значимых изменений в гидрофизических и гидрохимических условиях по сравнению со ст. 5340_2 отмечено не было. В верхнем перемешанном слое соленость составляла 6 PSU, температура – 13°С (рис. 2). В верхнем слое более 90% численности и биомассы фитопланктона формировали диатомовые водоросли, из них ~60% было представлено морскими видами. 42% численности и 46% биомассы приходилось на долю рода Chaetoceros, 20% численности и 6.8% биомассы – на долю Skeletonema costatum, 2% численности и 22% биомассы составляла Thalassiosira nordenskioeldii. В пикноклине с градиентом солености 5.1 PSU/м и под ним численность и биомасса фитопланктона были в 50 раз ниже, чем в верхнем однородном слое (рис. 6). Под пикноклином преобладали пресноводные диатомеи. Содержание хлорофилла в верхних 7 метрах составляло 2.7 мг Сhl a/м3, на 9 м снижалось до 0.2 мг Сhl a/м3. Как и на более южной станции ниже пикноклина, доля мертвых клеток пресноводных диатомей была в 8 раз выше, чем в верхнем однородном слое (табл. 2).
Самое низкое для исследованного района обилие водорослей было зарегистрировано на ст. 5337_2. Средняя численность фитопланктона в столбе воды здесь составляла 50.4 × 103 кл/л, средняя биомасса – 56.5 мг/м3 (табл. 1, рис. 5). Численность и биомасса фитоцена в верхнем однородном слое более чем на порядок превосходила значения для нижележащих слоев. По числу клеток в верхнем слое доминировали несколько видов рода Chaetoceros (62.5% общей численности), при этом вклад мелкоразмерного Chaetoceros wighamii составлял 42.3%. Существенную часть общей биомассы водорослей (52.0%) формировали динофлагелляты (Dinophysis acuminate, Peridinella catenata, Protoperidinium pellucidum). На станции наблюдалось резкое снижение доли пресноводных видов в фитоцене – до 10% от общей численности и 2.6% от биомассы (табл. 2). Изменения в фитопланктонном сообществе позволяют говорить, что в области между станциями 5339 и 5337 (~73° c.ш.) проходила важная граница между фитоценозами, различающимися по составу и количественным характеристикам, при том, что резкие широтные изменения свойств среды в этом районе не были отмечены (рис. 2, 3). Присутствие пресноводных диатомей и зеленых водорослей, связанное с поверхностным слоем значительно опресненной воды, прослеживалось и севернее, и эти формы исчезали из планктона при солености 11–12 PSU.
Во внешней части ЭФЗ на станциях 5335_2, 5333_2 (рис. 1) в верхнем перемешенном слое по численности доминировали виды рода Chaetoceros. Максимальный вклад в численность – 70–80% формировал C. wighamii. Фитопланктон на станциях 5335_2 и 5333_2 концентрировался в верхнем перемешенном слое (рис. 6). На ст. 5333_2 различия в численности и в биомассе фитопланктона между верхним слоем и слоем под пикноклином ниже 6–10 м превышали порядок величин. Содержание хлорофилла в верхнем слое составляло 1.2–1.6 мг Chl a/м3 и почти на порядок превосходило концентрацию в слое под пикноклином. Пресноводные диатомеи присутствовали в верхнем слое, но составляли всего 4.8% общей численности и 2.7% общей биомассы фитоцена. При этом на ст. 5335_2 на глубине 10 м они доминировали, и на их долю приходилось 55.8% общей численности и 43.6% общей биомассы фитопланктона. Число мертвых клеток диатомей в нижнем слое, как и на более южных станциях, существенно превышало число живых (табл. 2). Ст. 5335_2, выполненная на 74°45′ с.ш., была последней, на которой в значимом количестве были встречены представители пресноводной флоры.
На ст. 5350 (глубина 22 м), сделанной в ~180 км севернее внешней орографической границы эстуария, соленость в верхнем перемешанном слое и общий характер вертикальной стратификации практически не отличались от более южной группы станций (рис. 2, 3). Численность и биомасса фитопланктона были распределены в столбе воды относительно равномерно по сравнению с более южными районами. Максимальная биомасса фитопланктона (230 мг/м3) наблюдалась в верхнем девятиметровом слое и определялась высокой численностью динофлагеллят с большим объемом клеток 3500 мкм3. На долю динофлагеллят в этом слое приходилось 27.0% численности и 64.5% общей биомассы фитопланктона. Доминировала Peridinella catenata (47.7%) с объемом клеток от 5500 до 8000 мкм3. Отмечено небольшое увеличение общего числа клеток водорослей с глубиной. На глубинах 16–20 м ~80% общей численности фитопланктона составлял опустившийся в нижние слои Сhaetoceros wighamii. Концентрация хлорофилла в поверхностном слое была 1.7 мг Chl a/м3, на глубине 20 м – 1.3 мг Chl a/м3. Доля мертвых клеток водорослей в нижнем слое существенно снизилась по сравнению с более южными станциями (табл. 2).
На ст. 5351, выполненной в 220 км севернее створа эстуария над глубиной 43 м, изменилась структура полей солености и температуры (рис. 2, 4). В узком верхнем трехметровом слое соленость составляла 14.7 PSU, двумя метрами ниже увеличивалась до 26.5 PSU, глубже 10 м достигла 31 PSU. Температура снижалась с 11°С в верхнем слое до 0°С, характерного для подпикноклинных вод шельфовой области, ниже 10 м (рис. 2а). Концентрация нитратов в верхнем слое составляла <0.1 мкг-ат/л, кремния – 66.7 мкг-ат/л и с глубиной резко уменьшалась до 18 мкг-ат/л. (рис. 3, 4). По сравнению с более южными районами существенно изменился состав фитопланктона. В верхнем трехметровом слое в массе встречены мелкие жгутиковые диаметром 4–6 мкм, которые формировали 70% общей численности фитопланктона. Основу биомассы составляли динофлагелляты, доля которых достигала 58%. Наибольший вклад вносили Karenia mikimotoi (27.2% общей биомассы), Peridiniella catenata (12.7%), Dinophysis acuminatа (4.0%). Концентрация хлорофилла “а” составляла 1.6 мг Chl a/м3. В слое от 3 до 20 м численность и биомасса фитопланктона были на порядок ниже, чем в узком поверхностном слое, содержание хлорофилла уменьшалось до 0.1 мг Chl a/м3 (рис. 4). Ниже 20 метров на горизонтах 30 и 40 м численность водорослей возрастала до уровня близкого к наблюдавшемуся в поверхностном слое. Это определялось массовым скоплением спор Chaetoceros socialis (460 × 103 и 550 × 103кл/л соответственно), которые составляли 91.5 и 93.5% общей численности фитопланктона. Из-за малого размера спор биомасса на этих горизонтах увеличивалась не столь значительно и была в 3.5 раза меньше, чем в верхнем слое, содержание хлорофилла возрастало до 0.8 мг Chl a/м3 (рис. 4). Мертвых клеток морских диатомей было больше всего в обедненном фитопланктоном слое от 3 до 20 м – до 63% (табл. 2). В поверхностном слое и в слое 30–40 м процент мертвых клеток диатомей был невысоким и составлял 2.8% в слое 0–3 м, 5.7% на 30 м и 14% на 40 м.
Самые северные станции разреза 5352 (глубина 30 м) и 5353 (глубина 65 м) выполнены в пределах пелагического биотопа, существенно отличающегося по свойствам среды от более южных районов. Соленость в верхнем перемешенном слое здесь возрастала до 31 PSU, температура снижалась до 4–6°С. (рис. 2, 4). Ниже пикноклина соответствующие значения составляли 32.0–33.7 PSU и <–1.0°С, что характерно для большинства районов карского шельфа. Водная толща характеризовалась относительно глубоким – 12–14 м верхнем перемешанным слоем и растянутым по вертикали пикноклином со слабыми градиентами (рис. 2, 4). Концентрация нитратного азота в перемешанном слое составляла от 0.05 до 0.15 мкг-ат/л, кремния – ~3 мкг-ат/л (рис. 3, 4). Максимальная численность фитопланктона была зарегистрирована на ст. 5352 на глубинах 30–40 м (730 × 103 кл/л), на ст. 5353 – на 45–52 м (320 × × 103 кл/л) (рис. 4). На обеих станциях в нижних слоях высокую численность и значительную часть биомассы фитопланктона формировали споры Chaetoceros socialis, на ст. 5353 в сочетании со спорами Chaetoceros debilis. На ст. 5352 высокая биомасса в верхнем пятиметровом слое была сформирована диатомовыми – 79 мг/м3 (42.2%) и динофлагеллятами – 70 мг/м3 (37.2%). В нижних слоях биомасса фитопланктона определялась скоплением спор C. socialis (121.4 мг/м3, 89% общей биомассы). Повышенные концентрации хлорофилла отмечены в поверхностном слое – 0.6 мг Chl a/м3 и на 38 м – 0.63 мг Chl a/м3. На станции 5353 слой максимальной биомассы фитопланктона на глубине 45–52 м совпадал со слоем максимальной численности клеток (рис. 4). В этом слое доля диатомей и динофлагеллят в численности составляла соответственно 67.5 и 7.2%, в биомассе – 62 и 32%. Доминирующими видами были Chaetoceros socialis и C. debilis, представленные, в основном, в виде спор. Среди динофлагеллят наибольший вклад в биомассу вносили Peridiniella catenata, Karenia mikimotoi и Gymnodinium sp. В этом же слое была отмечена максимальная концентрация хлорофилла – 0.7 мг Chl a/м3.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Характер широтного распределения солености, температуры и гидрохимических характеристик позволил четко выделить в исследованном районе несколько различающихся пелагических биотопов:
Опресненная зона эстуария (станции 5345, 5344-2, 5342-2; 71°50.4′–72°12.4′ с.ш.) с соленостью <1 PSU в верхней части водной толщи, высоким содержанием кремния (>70 мкг-ат/л), низкой концентрацией нитратов (<0.5 мкг-ат/л) (рис. 2, 3) и фосфатов (<0.05 мкг-ат/л). Для районов с небольшими глубинами в пределах этого биотопа (ст. 5344, рис. 2, 3) типично равномерное распределение в толще воды всех гидрофизических и гидрохимических характеристик. Биотоп населен пресноводным фитопланктоном, выделяется самыми высокими во всем исследованном районе численностью и биомассой водорослей и содержанием хлорофилла “а” (табл. 1, рис. 5). Фитопланктон концентрируется в верхнем слое водной толщи (рис. 4).
Мористая (шельфовая) зона (станции 5352, 5353; севернее 75°55.0′ с.ш.) с соленостью в верхнем перемешанном слое >30 PSU, растянутым по вертикали пикноклином со слабыми градиентами (0.2–0.4 PSU/м), низкими концентрацияим нитратов, фосфатов и кремния в верхних 20–30 м и увеличением содержания биогенных элементов в несколько раз на глубине 30–50 м (рис. 2, 3, 4). Биотоп населен морскими видами водорослей. Концентрация фитопланктона достигала максимума на 30–50 м в слоях с высоким содержанием биогенных элементов (рис. 4). В поздневесенний сезон доминирующим видом был Chaetoceros socialis, представленный, в основном, спорами. В слоях максимальной концентрации фитопланктона вид составлял >90% общей численности клеток. В биомассу существенный вклад вносили динофлагелляты. Повышенная концентрация хлорофилла “а”, так же, как и обилие фитопланктона, наблюдалась в слоях ниже пикноклина (рис. 4).
Область, находящаяся между опресненным и морским биотопами – ЭФЗ, имела широтную протяженность 340–350 км (рис. 1, 2, станции 5342_2 – 5351; 72°12.4′–75°12.2′ с.ш.) и характеризовалась жесткой стратификацией водной толщи. Слой над пикноклином был обеднен нитратами (<0.5 мкг-ат/л), фосфатами (<0.05 мкг-ат/л), на фоне высокого содержания кремния >50 мкг-ат/л (рис. 3, 4, 6). В пикноклине и под пикноклином (глубже 5–7 м) наблюдалось резкое увеличение концентрации нитратов до 2.5–6.0 мкг-ат/л и фосфатов до 0.5 мкг-ат/л, а содержание кремния снижалось в 2–3 раза. Фитопланктон концентрировался в верхнем перемешенном слое, в котором зарегистрировано и максимальное для толщи воды содержание хлорофилла “а” (рис. 4, 6). Различия и в численности и в биомассе фитопланктона между верхним перемешенным слоем и слоем под пикноклином превышали порядок величин (рис. 4, 6).
У южной границы ЭФЗ (рис. 2, 3; станции 5342-5340_2; 72°12.4′–72°35.8′ с.ш.) формируется южный фронтальный раздел, где происходит наиболее интенсивное взаимодействие речных и морских вод. На формирование этого фронтального раздела оказывают влияние и приливно-отливные течения, действие которых усугубляется резким уменьшение глубины дна до 11 м (рис. 2). Соленость в верхнем перемешенном 5–7-метровом слое здесь резко возрастает до 6 PSU, что объясняет появление морских эвригалинных видов. В морской составляющей фитоцена по численности доминировали мелкоклеточные Chaetoceros wighamii и Skeletonema costatum. В верхнем слое ~30% биомассы фитопланктона приходилось на долю видов рода Chaetoceros и ~25% – на долю Thalassiosira. Вблизи 73° с.ш. между станциями 5339 и 5337 проходила хорошо выраженная граница между фитопланктонными сообществами, различающимися по составу и количественным характеристикам. Здесь наблюдалось резкое снижение обилия фитопланктона, сокращение его пресноводной составляющей, увеличение роли динофлагеллят. К северу от 73° с.ш. небольшое количество пресноводных водорослей встречалось вплоть до практически полной их элиминации в районе 74°15′ с.ш. при солености ~12 PSU.
Соотношение в популяциях диатомей живых и мертвых клеток, вегетативных клеток и спор позволяет судить о состоянии фитопланктона в период исследований. В южном опресненном биотопе эстуария в популяциях пресноводных видов диатомей доля мертвых клеток в верхнем слое водной толщи не превышала 25%, в нижнем слое – возрастала до 80–97%. В области ЭФЗ и на прилежащем к эстуарию мелководном шельфе в верхнем перемешанном слое живые клетки диатомей составляли от 73.0 до 91.3% (табл. 2). Судя по большому числу делящихся клеток морских видов водорослей, фитопланктон в этой части исследованного района находился в состоянии активного роста. По мере увеличения солености в верхнем перемешанном слое (до 7.5–10.0 PSU) на прилежащем к эстуарию шельфе (станции 5335 и 5333) в популяции доминирующего по численности эвригалинного вида Chaetoceros wighamii наблюдалось спорообразование и основная часть спор опускалась в верхнюю часть пикноклина. В мористой зоне скопление спор C. socialis ниже 30 м свидетельствовало об окончании цветения этого вида.
В нашем распоряжении имеются материалы, характеризующие состояние среды и фитоценозов тех же районов в осенний сезон (18–22 сентября 2011 г., [17]). Необходимо отметить, что и в конце июля, и в конце сентября концентрации нитратов и фосфатов на всей исследованной акватории были низкими. Это связано с геохимическими особенностями водосбора Енисея. Среднесибирское плоскогорье и другие участки Сибирской платформы, сложенные кристаллическими породами, устойчивыми к речной эрозии и вымыванию химических элементов стоковыми водами. Об этом говорит и низкая – ~2.0 мг/л концентрация взвеси в пресноводной части эстуарии в оба сезона [7]. Для поздней весны и осени для района исследований были характерны высокие концентрации кремния (рис. 3) [10].
Анализ качественного состава пресноводного фитоцена, населяющего опресненную зону Енисейского эстуария, показал, что в поздневесенний и осенний [17] сезоны основной вклад и в численность, и в биомассу вносили пресноводные диатомовые водоросли, главным образом рода Aulacoseira. Сезонные различия касались представительства зеленых и сине-зеленых водорослей. В поздевесенний период (исследования 2016 г.) в поверхностном слое в значительном количестве были встречены сине-зеленые водоросли родов Anabaena, Aphanocapsa, Aphanothece, Aphanizomenon и др. Даже при очевидном количественном недоучете этих водорослей из-за характера их вертикального распределения (см. выше), их вклад в общую биомассу фитопланктона в столбе воды доходил до 8%. В осенний сезон сине-зеленых водорослей было встречено крайне мало. В этот сезон второй группой по вкладу и в численность, и в биомассу фитоцена опресненной зоны были зеленые водоросли. Их доля в общей численности клеток варьировала от 3 до 18%, а в биомассе – от 2 до 8%.
В оба сезона средняя численность и биомасса фитопланктона в опресненной зоне были самыми высокими (рис. 7). Сложная орография дна, чередование участков эстуария с относительно большими и меньшими глубинами влияет на динамические характеристики потока и действие приливных течений. Этим, вероятнее всего, определяются наблюдаемые отличия в гидрофизической структуре водной толщи, вертикальном распределении биогенных элементов, планктонных водорослей и хлорофилла “а” в южной опресненной части эстуария, “отгороженной” от его более северной части мелководной областью с глубиной ~11 м. В поздневесенний сезон (ст. 5345) при глубине 21 м в верхних 9 м соленость была 0.4 PSU. Ниже лежал пикноклин, в котором соленость возрастала до 19 PSU, температура снижалась с 17.5°С до 6.5°С, концентрация нитратов увеличивалась с 0.22 до 5.7 мкг-ат/л, фосфатов с 0.12 до 0.45 мкг-ат/л. На фоне такой стратификации водной толщи численность и биомасса фитопланктона в слое пикноклина и под ним были более чем на порядок ниже, чем в верхнем девятиметровом слое, а концентрация хлорофилла “а” – в 12 раз. Фитопланктон состоял исключительно из пресноводных видов. Осенью в этом районе эстуария (ст. 5013) при глубине 32 м выраженная стратификация водной толщи отсутствовала. Распределение солености, температуры и нитратов мало менялось от поверхности до дна, и соответствующие величины составляли 0.06 PSU, 9.5°С и 0.22 мкг-ат/л [17]. Концентрация хлорофилла в столбе воды варьировала от 2.5 до 2.9 мг Chl a/м3 [20], различия в численности и биомассе фитопланктона верхнего и нижнего слоев не превышали трех раз [17].
Рис. 7.
Распределение поверхностной и придонной солености (S, PSU), (а) – численности (N × 103 кл/л) и (б) – биомассы (В, мг/м3) фитопланктона на Енисейском разрезе в июле 2016 г. и в сентябре 2011 г. N и В – средние для столба воды.
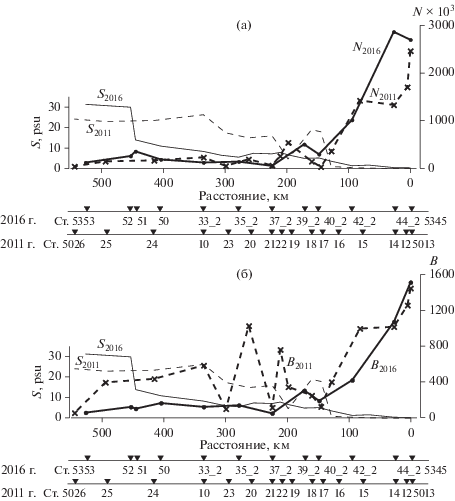
Область ЭФЗ характеризовалась наибольшими сезонными изменениями. В этой зоне наблюдались различия в видовом составе, численности, биомассе и вертикальном распределении фитопланктона (рис. 7). В поздневесенний сезон среди морских видов диатомей в численности преобладали мелкоклеточные Chaetoceros wighamii (средний объем клетки (CV) ~ 1000 мкм3), споры С. socialis (CV ~100 мкм3), Skeletonema costatum (CV ~ ~ 200 мкм3) и споры Chaetoceros spp. (CV ~ 700 мкм3). В биомассу помимо видов рода Chaetoceros существенный вклад вносили Thalassiosira sp. и T. nordenskioeldii. Численность и биомасса водорослей в верхнем перемешенном слое были на порядок выше, чем в слое пикноклина и под ним. Доля пресноводного фитопланктона и в численности и в биомассе увеличивалась с глубиной.
Пресноводный фитопланктон в области ЭФЗ был сконцентрирован в верхнем перемешенном слое, где его доля в численности и в биомассе доходила до 90%. В нижних слоях пресноводная составляющая вносила существенно меньший вклад в численность (5–25%) и в биомассу (1–18%). Здесь доминировали крупноклеточные морские виды диатомей Сhaetoceros compressus (CW ~ 2000 мкм3) и Thalassiosira nordenskioeldii (CW ~ 12 000 мкм3), Leptocylindrus danicus (CW ~ ~ 2500 мкм3). Различия в биомассе между верхними и нижними слоями водной толщи не превышали 3–4 раз.
В поздневесенний и осенний сезоны с районом ~73° с.ш. была связана хорошо выраженная граница фитоценозов разного состава и количественных характеристик. Эта граница между фитоценозами существовала на фоне отсутствия резких изменений условий среды. При этом небольшое количество пресноводных водорослей (до 10% численности и 2% биомассы) было встречено севернее вплоть до северной границы ЭФЗ, где соленость в верхнем перемешанном слое возрастала до 12–16 PSU. В поздневесенний период эта граница лежала на 74°15′ с.ш. В осенний сезон пресноводные виды исчезали на 73°20′ с.ш., где проходила узкая граница шириной ~20 км, разделяющая сильно опресненные поверхностные воды с соленостью 5–6 PSU и воды шельфа с соленостью 15–16 PSU [17].
В северной части ЭФЗ в оба сезона наблюдений доминировали морские виды. При сопоставимой численности водорослей в поздневесенний и осенний сезоны биомасса фитопланктона в осенний период была значительно выше благодаря доминированию крупноклеточных видов Сhaetoceros compressus, Thalassiosira nordenskioeldii, Leptocylindrus danicus. (800 и 160 мг/м3 соответственно).
Существенные различия наблюдались в вертикальном распределении фитопланктона. В поздневесенний сезон водоросли концентрировались в верхнем перемешенном слое. Численность и биомасса в этом слое более чем на порядок превосходили величины, наблюдаемые в пикноклине и под ним (рис. 4, 6). Соответствующие различия в содержании хлорофилла составляли разы. В осенний сезон в условиях не менее жесткого пикноклина максимальные величины численности и биомассы (от 800 до 2300 мг/м3) фитопланктона зарегистрированы в нижних слоях водной толщи на 15–20 м (рис. 6, ст. 5020) [17]. Здесь же зарегистрированы максимальные концентрации хлорофилла “а”. Высокая прозрачность воды (показатель ослабления света 0.50–0.55 м–1) и высокое положение пикноклина (5–7 м), ниже которого в разы возрастало содержание биогенных элементов, создавали условия для успешного развития крупноклеточного морского фитопланктона на этих глубинах.
В фитоцене срединного шельфа (глубины >50 м), так же, как на мелководном шельфе, наблюдались выраженные сезонные различия в составе фитопланктона, которые, в свою очередь, определили существенные различия в биомассе водорослей (рис. 7). В осенний сезон не наблюдались характерные для поздневесеннего периода глубинные максимумы фитопланктона, образованные скоплениями Chaetoceros socialis. Осенью фитопланктон был или равномерно распределен в слое 0–30 м или концентрировался в слое 5–10 м. В оба сезона наблюдений фитопланктон в северной части разрезов был самым бедным и по численности и по биомассе [17].
Почти одновременно (18–24 июля 2016 г.) с поздневесенними работами в Енисейском эстуарии был собран материал в Обском эстуарии и на прилежащем шельфе [16], что позволяет провести сравнение качественных и количественных характеристик фитоценозов в этих районах.
Общим для обоих исследованных районов является хорошо выраженная широтная зональность в распределении гидрофизических и гидрохимических характеристик среды, которая формировала пространственную структуру сообществ фитопланктона. Исследованиями Обского эстуария были также выделены: зона, занятая опресненной водой речного стока, населенная пресноводным комплексом видов; эстуарная фронтальная зона с жестким пикноклином, населенная смешенным пресноводным, морским и солоноватоводным фитопланктоном; зона мелководного шельфа с сохраняющимся жестким пикноклином и морскими неритическими эвригалинными видами водорослей; морская зона со слабым заглубленным пикноклином и морским фитоценом. При общем сходстве картины зональности существуют существенные региональные различия в условиях среды и количественных характеристиках фитоценозов. Обский эстуарий отличался от Енисейского, прежде всего, значительно более высокой концентрацией нитратов в верхнем слое – до 15.2 мкг-ат/л и в 4 раза более высокой численностью и биомассой фитопланктона (рис. 8) [16]. Различия в гидрохимических характеристиках эстуариев Оби и Енисея связаны с особенностями водосборных бассейнов этих рек и различиями в условиях прохождения речного стока через эстуарные районы [2, 11, 12]. Концентрация взвеси в Обском эстуарии была почти на порядок выше, чем в Енисейском – от 10.0 до 14.5 мг/л. Вместе с взвесью с обширной заболоченной территории Западно-Сибирской низменности с водами Оби выносится значительное количество биогенного материала, формирующего основу для регенерации биогенных элементов [2, 11].
Рис. 8.
Распределение поверхностной и придонной солености (S, PSU), (а) – численности (N × 103 кл/л) и (б) – биомассы (В, мг/м3) фитопланктона на Енисейском (SЕн, NЕн, BЕн) и Обском (SОбь, NОбь, BОбь) разрезах в июле 2016 г. N и В – средние для столба воды.
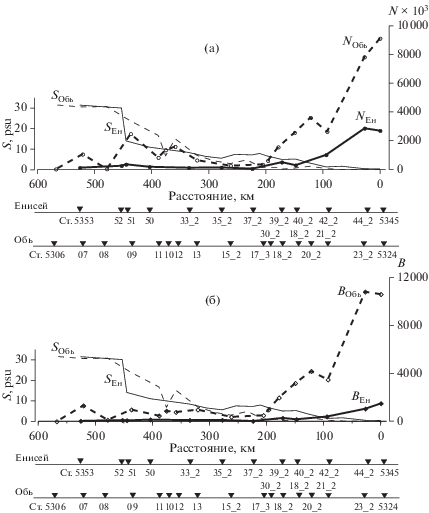
Фитоцен внутренних опресненных зон обоих эстуариев с соленостью <1 PSU состоял из пресноводных видов, главным образом диатомей, на долю которых и в численности и в биомассе приходилось от 63 до 83%. Отличие касалось синезеленых водорослей в значительном количестве встреченных в Енисейском эстуарии и практически отсутствовавших в Обском. Кроме того, во внешней части Обского эстуария (~72°30′ с.ш.) обнаружен небольшой район с кардинально отличающимся фитопланктонным сообществом, связанным, как мы предполагаем, с квазизамкнутой циркуляцией в восточной части Обской губы [16]. В вертикальном распределении фитопланктона во внутренней опресненной части Обского эстуария максимальная концентрация клеток, так же, как и в Енисейском эстуарии, наблюдалась в верхнем слое, однако различия и в численности и в биомассе в верхних и нижних слоях водной толщи были менее выражены и составляли разы, а не порядок величин, как в Енисее (рис. 4) [16].
На южной границе ЭФЗ, где формировался жесткий фронтальный раздел, в обоих эстуариях соленость в поверхностном слое резко возрастала от десятых долей до 4–6 PSU. В Оби эти изменения сопровождались резким падением концентрации нитратов с 4.6 мкг-ат/л до <0.5 мкг-ат/л, т.е. до уровня, характерного для всей исследованной области Енисейского эстуария и прилежащего шельфа. В обоих эстуариях численность и биомасса фитопланктона на южной границе ЭФЗ снижалась в 3–4 раза.
Общим для областей ЭФЗ Оби и Енисея было доминирование в морской составляющей фитоценозов водорослей рода Chaetoceros, наиболее многочисленным был один и тот же вид – Chaetoceros wighamii. В Оби помимо Chaetoceros в биомассу существенный вклад вносили крупные солоноватоводные виды Thalassiosira baltica, Cyclostephanus sp., Stephanopyxis sp., cf. Astasia sp., встреченные в ЭФЗ Енисея лишь изредка. Численность и биомасса фитопланктона в Обской ЭФЗ была в несколько раз выше, чем в Енисейской. Северная граница ЭФЗ Оби проходила в области внутреннего шельфа на 74°39′ с.ш., в енисейском районе – на 75°12′, т.е. на ~60 км севернее.
На Енисейском мелководье максимальные численность и биомасса фитопланктона наблюдались в верхнем перемешенном слое. Здесь большой вклад в биомассу фитопланктона наряду с C. wighamii вносили динофлагелляты – Peridinella catenata, несколько видов Dinophysis и др. На Обском мелководье максимальные скопления фитопланктона зарегистрированы под пикноклином. Доминирующую роль в численности водорослей в этом слое играл С. socialis, биомассу формировали крупноклеточные диатомеи Thalassiosira nordenskioeldii, T. pacifica, Melosira nummuloides и на нескольких станциях – cf. Astasia sp. в виде цист [16].
На cрединном шельфе за пределами ЭФЗ в 180–260 км севернее створов Обского и Енисейского эстуариев максимальная численность фитопланктона наблюдалась в слоях ниже 25 метров и определялась скоплениями спор Chaetoceros socialis. Значительные пространственные колебания численности определялись только пятнистостью распределения этого вида. На шельфе севернее Енисейского эстуария равнозначные вклады в биомассу фитопланктона вносили C. socialis, динофлагелляты, C. debilis, Thalassiosira nordenskioeldii. На Обском шельфе роль динофлагеллят в биомассе была незначительна. От 32 до 76% биомассы формировали T. nordenskioeldii, T. bioculata, T. pacifica и Rhizosolenia setigera [16].
ВЫВОДЫ
1. Общим для районов эстуариев Енисея и Оби и прилежащего шельфа Карского моря является выраженная широтная зональность в пространственной структуре фитоценозов, связанная с особенностями гидрофизической и гидрохимической структуры биотопов.
2. В обоих эстуариях самыми высокими величинами численности и биомассы фитопланктона характеризовались опресненные до <1 PSU внутренние зоны, которые населены пресноводным диатомовым фитопланктоном с преобладанием нескольких видов рода Aulacoseira. Опресненная зона Обского эстуария отличалась в несколько раз более высоким обилием фитопланктона, что связано с благоприятным биогенным режимом.
3. В наибольшей степени снижение численности и биомассы фитопланктона в эстуариях Енисея и Оби связано с южной периферией ЭФЗ – возрастанием солености в верхнем слое до 1–2 PSU.
4. Северная граница распространения пресноводных видов водорослей в Енисейском и Обском районах в поздневесенний сезон лежала в области северной периферии ЭФЗ – на 75°12′ и 74°39′ с.ш. соответственно, где соленость в верхнем перемешенном слое возрастала до 11–15 PSU. Осенью в районе Енисея граница распространения пресноводных видов водорослей была смещена к югу относительно поздневесеннего сезона и проходила на ~73°20′ с.ш., что, вероятнее всего, определялось менее интенсивным стоком.
5. При одинаково низком содержании биогенных элементов в верхнем перемешенном слое ЭФЗ Енисея и Оби в поздневесенний период обилие фитопланктона в Обской ЭФЗ было в несколько раз выше. При этом в фитиоценозах обоих эстуариев по численности доминировал один и тот же мелкоклеточный вид диатомей – Сhaetoceros wighamii.
6. В поздневесенний период в районе северной границы ЭФЗ обоих рек и на срединном шельфе на глубинах от 25 до 50 м наблюдались глубинные скопления фитопланктона с численностью и биомассой, как правило, превышающей эти показатели в вышележащих слоях в 5–7 раз. Скопления были сформированы одним и тем же видом Сhaetoceros socialis главным образом в виде спор. Цветение вида, вероятно, прошло ранней весной.
7. Енисейская ЭФЗ характеризовалось на порядок более высокими численностью и биомассой водорослей в верхнем перемешенном слое по сравнению со слоями под пикноклином. В Обской ЭФЗ максимальные концентрации водорослей были приурочены к слоям глубже 15 м. Эти подпикноклинные скопления формировались крупноклеточными морскими и солоноватоводными диатомеями в условиях высоких концентраций биогенных элементов.
8. В ЭФЗ Енисея в поздневесенний и осенний сезоны фитопланктонные сообщества существенно различались по видовому составу и обилию. Осенний сезон характеризовался в несколько раз более высокой биомассой, что определялось доминированием крупноклеточного фитопланктона с объемом клеток от 2000 до 12 000 мкм3: Сhaetoceros compressus, Thalassiosira nordenskioeldii, Leptocylindrus danicus.
9. Для Енисейской ЭФЗ были характерны существенные сезонные различия в вертикальном распределении водорослей. В поздневесенний период обилие фитопланктона в верхнем перемешенном слое было на порядок выше, чем под пикноклином. В осенний cезон максимальные численность и биомасса фитопланктона были приурочены к слоям воды под пикноклином.
10. Область срединного шельфа к северу от ЭФЗ Енисея в поздневесенний и осенний сезоны характеризовалась близкими значениями численности фитопланктона. Более высокая биомасса водорослей, связанная с преобладанием в сообществе крупноклеточного диатомового фитопланктона, отмечена в осенний период.
Благодарности. Авторы благодарят А.Б. Демидова за предоставленные данные о границе слоя фотосинтеза, содержании хлорофилла и феофитина, Т.А.Кузьмичеву за помощь в отборе и подготовке проб фитопланктона, анонимного рецензента за детальный анализ рукописи.
Источники финансирования. Исследования выполнены в рамках госзадания, тема № 149-2019-0008, обработка проб фитопланктона и компютерная обработка первичного материала поддержаны Проектом РФФИ № 18-05-60 069 Арктика, данные по гидрохимии получены при поддержке Проекта РНФ № 19-17-00196.
Список литературы
Арашкевич Е.Г., Дриц А.В., Флинт М.В. и др. Распределение и питание растительноядного зоопланктона в море Лаптевых // Океанология. 2018. Т. 58. № 3. С. 404–419.
Артамонова К.В., Лапин С.А., Лукьянова О.Н., Маккавеев П.Н., Полухин А.А. Особенности гидрохимического режима Обской губы в период открытой воды // Океанология. 2013. Т. 53. № 3. С. 357–366.
Бабич Д.Б., Большиянов Д.Ю., Зайцев А.А. и др. Региональные особенности формирования эстуарно-дельтовых систем Арктического побережья Сибири. В кн.: Эстуарно-дельтовые системы России и Китая. М.: ГЕОС. 2007. С. 112–240.
Гордеев В.В., Джамалов Р.Г., Зекцер И.С. и др. Оценка выноса биогенных элементов с речным и подземным стоком в окраинные моря Российской Арктики // Водные ресурсы. 1999. Т. 26. № 2. С. 206–211.
Дриц А.В., Арашкевич Е.Г., Недоспасов А.А., Амелина А.Б., Флинт М.В. Структурно-функциональные характеристики зоопланктона обского эстуария и прилегающих районов шельфа Карского моря в летний период // Океанология. 2019. Т. 59. № 3. С. 383–395.
Зацепин А.Г., Завьялов П.О., Кременецкий В.В. и др. Поверхностный опресненный слой вод в Карском море // Океанология. 2010. Т. 50. № 5. С. 698–708.
Кравчишина М.Д., Леин А.Ю., Суханова И.Н. и др. Генезис и пространственное распределение концентрации взвеси в Карском море в период наибольшего сокращения арктической ледовой шапки // Океанология. 2015. Т. 55. № 4. С. 687–708.
Лисицын А.П. Маргинальный фильтр океанов // Океанология. 1994. Т. 34. № 5. С. 735–747.
Макаревич П.Р. Планктонные альгоценозы эстуарных экосистем. М: Наука, 2007. 223 с.
Маккавеев П.Н., Полухин А.А., Налбандов Ю.Р., Хлебопашев П.В. Динамика биогенных элементов в Енисейском заливе в период открытой воды // Арктика: экология и экономика. 2019. № 4 (36). С. 69—82. https://doi.org/10.25283/2223-4594-2019-4-69-82
Никаноров А.М., Брызгало В.А., Косменко Л.С., Даниленко А.О. Реки материковой части Российской Арктики. Ростов-на-Дону: Изд-во Изд-во Южного федерального ун-та, 2016. 275 с.
Стунжас П.А., Маккавеев П.Н. Объем вод Обской губы как фактор формирования гидрохимической неоднородности. // Океанология. 2014. Т. 54. № 5. С. 622–634.
Cуханова И.Н. Концентрирование фитопланктона в пробе // Современные методы количественной оценки распределения морского планктона. М.: Наука. 1983. С. 97–105.
Суханова И.Н., Флинт М.В., Георгиева Е.Ю. и др. Структура сообществ фитопланктона в восточной части моря Лаптевых // Океанология. 2017. Т. 57. № 1. С. 86–103
Суханова И.Н., Флинт М.В., Мошаров С.А. и др. Структура сообществ фитопланктона и первичная продукция в Обском эстуарии и на прилежащем карском шельфе // Океанология. 2010. Т. 50. № 5. С. 785–800.
Суханова И.Н., Флинт М.В., Сахарова Е.Г. и др. Фитоценозы Обского эстуария и Карского шельфа в поздневесенний сезон // Океанология. 2018. Т. 58. № 6. С. 882–898.
Суханова И.Н., Флинт М.В., Сергеева В.М. и др. Структура сообществ фитопланктона Енисейского эстуария и прилежащего Карского шельфа // Океанология. 2015. Т. 55. № 6. С. 935–948.
Стунжас П.А., Маккавеев П.Н. Объем вод Обской губы как фактор формирования гидрохимической неоднородности. // Океанология. 2014. Т. 54. № 5. С. 622–634.
Флинт М.В., Семенова Т.Н., Арашкевич Е.Г. и др. Структура зоопланктонных сообществ в области эстуарной фронтальной зоны реки Обь // Океанология. 2010. Т. 50. № 5. С. 809–822.
Demidov A.B., Gagarin V.I., Vorobieva O.V. et al. Spatial and vertical variability of primary production in the Kara Sea in July and August 2016: The influence of the river plume and subsurface chlorophyll maxima // Pol. Biol. 2018. V. 41. № 3. P. 563–578.
Deubel H., Engel M., Fetzer I. et al. The south Kara Sea ecosystem : Phytoplankton, zooplankton and benthos communities influenced by river run-off. Siberian river run-off in the Kara Sea / Eds. Stain R. et al. Amsterdam: Elsevier Science B.V. 2003. P. 237–266.
Drits, A.V., Pasternak, A.F., Arashkevich E.G. et al. Distribution and grazing of the dominant mesozooplankton species in the Yenisei estuary and adjacent shelf in early summer (July 2016) // Continental Shelf Research 2020. V. 201. P. 104–133.
Drits A., Pasternak A., Flint M. Distribution and grazing of dominant zooplankton species in the Ob Estuary: Influence of the runoff regime //Estuaries and Coasts. 2017. Vol. 40. P. 1082–1095.
Futterer D.K., Galimov E.M. Siberian river run-off in the Kara Sea: Characterization, quantification, variability and environmental significance. Siberian river run-off in the Kara Sea / Eds. Stain R. et al. Amsterdam: Elsevier Science B.V., 2003. P. 1–8.
Gordeev V.V. River input of water, sediment, major ions, nutrients and trace metals from Russian territory to the Arctic Ocean // The freshwater budget of the Arctic Ocean. Ed. E.L.Lewis. Dordrecht et al.: Kluwer. 2000. P. 297–322.
Gordeev V.V., Martin J.M., Sidirov M.V. et al. A reassessment of the Eurasian river input of water, sediment, major elements, and nutrients to the Arctic Ocean // Amer. J. Sci. 1996. № 296.
Johannessen O.M., Borodachev V.E. et al. Polar seas oceanography: An integrated study of the Kara Sea. Berlin: Springer, 2002. 450 p.
Le Fevre J. Aspects of the biology of frontal systems // Advances in Marine Biology. 1986. V. 23. P. 163–299.
Menden-Deuer S., Lessard E.J. Carbon to volume relationships for dinoflagellates, diatoms and other protist plankton // Limnol. Oceanogr. 2000. V. 45. № 3. P. 569–579.
Sorokin Yu.I., Sorokin P.Yu. Plankton and primary production in the Lena river estuary and in the south-eastern Laptev Sea // Estuar. Coastal. Shelf Sci. 1996. V. 43. P. 399–418.
Stein R. Circum Arctic river discharge and its geological record // Int. J. Earth Science. 2000. V. 89. P. 447–449.
Strathmann R.R. Estimating the organic carbon content of phytoplankton from cell volume, cell area or plasma volume // Limnol. Oceanogr. 1967. V. 12. № 3. P. 411–418.
Volkov V.A., Johannessen O.M., Borodachev V.E. et al. Polar seas oceanography: An integrated study of the Kara Sea. Berlin: Springer, 2002. 450 p.
Дополнительные материалы отсутствуют.