

Океанология, 2021, T. 61, № 3, стр. 423-432
Бентосные фораминиферы моря Лаптевых и их связь с гидрологическими условиями моря
Т. А. Хусид 1, *, Н. В. Либина 1, М. М. Доманов 1
1 Институт океанологии им. П.П. Ширшова РАН
Москва, Россия
* E-mail: tkhusid@mail.ru
Поступила в редакцию 26.09.2020
После доработки 10.02.2021
Принята к публикации 17.03.2021
Аннотация
Проанализирована фауна бентосных фораминифер из северной части моря Лаптевых. Численность раковин бентосных фораминифер в море Лаптевых повсеместно низкая, не более 3–9 экз/г. Наибольшее число фораминифер найдено в западном районе моря на шельфе пролива Вилькицкого. Подавляющее большинство экземпляров в море представлено агглютинирующими формами (77–100%). В восточном районе моря комплекс почти полностью состоит из агглютинированных раковин (90–100%). На шельфе и континентальном склоне в западной части моря постоянно встречается известковый вид Melonis barleеanus. Его доля составляет до 10–15%. Часто в исследуемом районе попадаются мелкие окатанные раковины планктонных фораминифер, принадлежащие полярному виду Neogloboquadrina pachyderma sin. В единственной абиссальной пробе из восточного района число раковин этого вида возрастает до нескольких сотен экз/г. Эта масса раковин представлена и мелкими (ювенильными), и крупными (взрослыми) особями, большинство нормальной сохранности. Здесь не найдено ни одной раковины бентосных фораминифер.
Море Лаптевых представляет собой один из наименее изученных бассейнов на северном побережье Азии. Море Лаптевых разделяется на восточную и западную зоны, граница между которыми проходит по Оленецкому грабену на 120° в.д., эта граница рассматривается как продолжение глубинного хребта Гаккеля (центральная рифтовая долина Арктического океана) [1, 3, 18]. Западная и восточная части моря Лаптевых отличаются историей геологического развития, строением земной коры и имеют различные гидрологические и геоморфологические характеристики [3, 5, 8, 12–14, 18]. Для поверхностных вод западной части моря, куда Таймырское течение приносит воды Арктического бассейна, характерны низкие температуры от +2 до –3°С. В восточной части моря, где распространяется основная масса теплых речных вод, температура на поверхности может достигать +6°С [5].
В подповерхностном слое в западную часть моря, куда с севера вдается глубоководный желоб, проникают соленые и несколько более теплые атлантические воды. В ядре слоя 200–300 м температура воды от +0.5 до –1.5°С, а соленость 34.7–34.9‰ [5]. В восточную часть моря с реверсивными течениями, типичными для подводных реликтовых речных долин, из Центрального Арктического бассейна также поступают трансформированные атлантические воды [4]. Однако поступление атлантических вод в западную часть моря Лаптевых более интенсивное, чем в восточную [5].
Для придонного слоя вод шельфовой зоны моря Лаптевых в пределах изобаты 100 м наиболее достоверные сведения о среднегодовых значениях температуры приведены в работе [4] (табл. 1). Среднегодовая температура придонного слоя изменяется от –0.8 до –1.8°С. Западная часть моря характеризуется низкими температурами от –1.8 до –1.6°С. Эта холодная зона простирается до центральной части моря. В юго-восточной части моря температура воды увеличивается до –0.8°С, но с увеличением глубин более 35 м температура уменьшается от –0.8 до –1.5°С.
Таблица 1.
Характеристики придонного слоя воды и осадка
| Станция | Глубина, м | Пелит, % | Сорг, % | T,°С придонного слоя воды | Соленость, ‰ | Кислород, мл/л |
|---|---|---|---|---|---|---|
| 5947 | 72 | 77 | 0.81 | –1.8175 | 34.34 | 7.328237 |
| 5949 | 547 | 96.7 | 0.59 | 0.742 | 34.80 | 7.000651 |
| 5950 | 1058 | 78.3 | 0.49 | 0.3642 | 34.90 | 6.81769 |
| 5954 | 1545 | 98.1 | 1.04 | –0.6163 | 34.92 | 6.749048 |
| 5956 | 2370 | 99.95 | 0.89 | –0.7677 | 34.94 | 6.652918 |
| 5958 | 2996 | 91.82 | 0.95 | –0.7387 | 34.94 | 6.732713 |
| 5960 | 357 | 95 | 0.77 | 0.2693 | 34.78 | 6.939865 |
| 5961 | 765 | 98.1 | 0.86 | 0.3739 | 34.87 | 6.959608 |
| 5962 | 1090 | 90 | 0.81 | –0.0777 | 34.91 | 6.790204 |
| 5963 | 1484 | 92.5 | 0.98 | –0.6646 | 34.92 | 6.762173 |
| 5964 | 2446 | 98.6 | 1.13 | –0.768 | 34.94 | 6.752661 |
| 5965 | 1994 | 99.9 | 1.24 | –0.7504 | 34.93 | 6.802533 |
| 5966 | 715 | 98.4 | 0.71 | 0.1984 | 34.85 | 7.135468 |
| 5967 | 426 | 96.1 | 0.67 | 0.3918 | 34.82 | 7.009918 |
| 5968 | 350 | 96.3 | 0.77 | 0.2459 | 34.78 | 7.028406 |
| 5969 | 277 | 93.5 | 0.79 | –0.0274 | 34.74 | 6.98 |
| 5970 | 239 | 67.3 | 0.93 | –0.511 | 34.67 | 7.160106 |
| 5972 | 210 | 98.61 | 0.91 | –0.7392 | 34.65 | 7.028711 |
| 5973 | 209 | 51.2 | 0.85 | –0.8568 | 34.35 | 7.028711 |
| 5944 | 217 | 99 | 0.85 | –0.864 | 34.63 | 7.111064 |
Большая часть дна моря покрыта глинистыми илами, содержащими пелитовую фракцию (<0.01 мм) в количестве более 70%. Эти илы в основном распространены в восточной части моря. Скорость седиментации в верхней части континентального склона (глубина 270 м) составляет 5 см/тыс. лет [11].
В северной части моря скорость седиментации в позднем голоцене низкая – около 2 см/тыс. лет [23]. Глубоководная фауна моря отличается бóльшим качественным разнообразием в западной части моря по сравнению с восточной. В северо-западной части моря Лаптевых (200–400 м) выявлено особенно интенсивное поднятие представителей абиссальной фауны Арктического бассейна по идущим от больших глубин желобам в сравнительно мелководную зону моря [6]. О видовом составе фораминифер и их распределении в море известно очень мало. Из-за трудной доступности к глубоководным районам моря Лаптевых исследования удается проводить большей частью в шельфовых районах моря [9–11, 16, 19–21, 37, 38].
В работе [16] прослежена последовательная смена комплексов фораминифер в голоцене, происходящая вслед за подъемом уровня моря и затоплением шельфа, от эстуарных к современным морским. Распределение современных комплексов фораминифер подчинено изменениям параметров окружающей среды – глубины, придонной солености, удаленности от устьев рек, гидродинамического режима [37]. Уровень моря Лаптевых стабилизировался и стал близок к современному около 5–6 тыс. л. н. [2, 7, 12, 13, 23]. После этого гидрологические условия в Арктике также стабилизировались [21]. В настоящее время проводятся совместные многодисциплинарные исследования российскими и немецкими учеными в рамках российско-германских программ по изучению Арктики [15]. Задачей настоящего исследования было проследить пространственное распределение численности и видового состава основных бентосных фораминифер в сублиторальной, батиальной и абиссальной зонах северного региона моря Лаптевых в поверхностном слое осадка и сопоставить эти данные с условиями среды.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Распределение бентосных фораминифер исследовали в северной части моря Лаптевых на большом пространстве от 105° в.д. и до 125° в.д.; от шельфа (72–210 м) и до предельной, абиссальной глубины 2996 м. Материалом для анализа послужили 40 проб, отобранных бокс-корером 50 × × 50 × 60 см в 72-м рейсе НИС “Академик Мстислав Келдыш” в августе–сентябре 2018 г. (рис. 1).
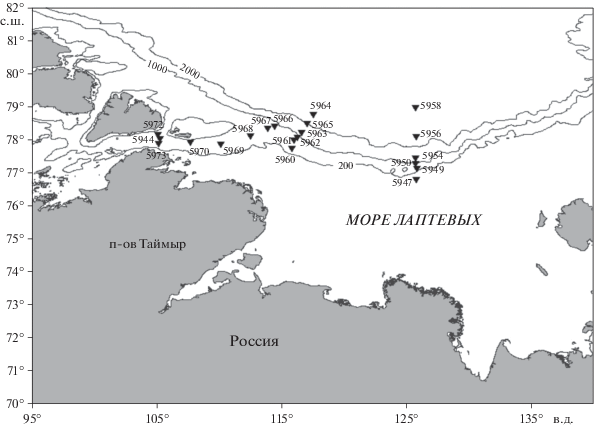
Так как состав фораминифер в этой части моря очень бедный, а скорости седиментации чрезвычайно низкие, то пробы отбирали из горизонта 0–2 и 0–5 см поверхностного слоя осадка (табл. 2, 3). Этот слой охватывает время 2–3 тыс. лет [22]. Полученные результаты показывают, что состав видов в верхнем слое 0–2 см и в слое 0–5 см сходные.
Таблица 2.
Распределение основных видов фораминифер в слое 0–5 см
| Станция | Глубина, м | Число бентосных форам., экз/г | Число видов бентосных форамин. | % агглютини- рующих форм | Reophax spp., % | C. subglobosa | A. glomeratum | T. nana | N. pseudonodulosa | M. barleeanus | Число N. pachyderma sin., экз/г |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 5947 | 72 | 19.5 | 12 | 65 | 12 | 35 | 0 | 0 | 0 | 0 | 0 |
| 5949 | 547 | 35 | 13 | 80 | 5 | 1 | 4 | 0 | 0 | 0 | 2 |
| 5950 | 1058 | 7 | 5 | 100 | 41 | 41 | 0 | 0 | 0 | 1 | 0 |
| 5954 | 1545 | 1 | 3 | 100 | 43 | 21 | 0 | 0 | 0 | 0 | 0.2 |
| 5956 | 2370 | 0.5 | 2 | 100 | 75 | 25 | 0 | 0 | 0 | 0 | 5 |
| 5958 | 2996 | 0.2 | 1 | 100 | 0 | 25 | 0 | 0 | 0 | 0 | 302 |
| 5960 | 357 | 8.5 | 10 | 82 | 37 | 2 | 10 | 14 | 2 | 14 | 0.1 |
| 5961 | 765 | 7 | 12 | 88 | 30 | 19 | 0 | 3 | 4 | 4 | 0.5 |
| 5962 | 1090 | 21 | 10 | 83 | 12 | 58 | 4 | 2 | 8 | 2 | 0.5 |
| 5963 | 1484 | 1.4 | 5 | 100 | 6 | 6 | 0 | 17 | 55 | 0 | 0.2 |
| 5964 | 2446 | 0.2 | 1 | 100 | 0 | 100 | 0 | 0 | 0 | 0 | 2.5 |
| 5965 | 1994 | 0.1 | 2 | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 0.1 |
| 5966 | 715 | 2.6 | 8 | 77 | 30 | 8 | 0 | 0 | 14 | 2 | 0.1 |
| 5967 | 426 | 1.4 | 8 | 88 | 18 | 0 | 0 | 21 | 34 | 6 | 0.1 |
| 5968 | 350 | 1 | 7 | 84 | 37 | 14 | 16 | 5 | 0 | 14 | 0 |
| 5973 | 209 | 9 | 9 | 84 | 18 | 29 | 0 | 10 | 0 | 10 | 0 |
| 5972 | 210 | 5 | 14 | 80 | 12 | 6 | 43 | 8 | 0 | 8 | 0 |
| 5944 | 217 | 8 | 12 | 67 | 6 | 6 | 42 | 3 | 0 | 8 | 1 |
| 5970 | 239 | 8 | 10 | 90 | 32 | 34 | 12 | 0 | 0 | 3 | 0 |
| 5969 | 277 | 23 | 14 | 58 | 5 | 22 | 20 | 5 | 0 | 3 | 0.3 |
Таблица 3.
Распределение основных видов фораминифер в слое 0–2 см
| Станция | Глубина, м | Число бентосных форам., экз/г | Число бентосных видов | % агглютини- рующих форм | Reophax spp., % | C. subglobosus | A. glomeratum | T. nana | N. pseudonodulosa | M. barleeanus | N. labradoricum | Число N. pachyderma sin., экз/ г |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 5947 | 72 | 35 | 10 | 46 | 16 | 22 | 0 | 0 | 0 | 0 | 0 | 0 |
| 5949 | 547 | 13.5 | 8 | 99 | 23 | 7 | 15 | 47 | 0 | 1 | 0 | 0.2 |
| 5950 | 1058 | 5 | 5 | 100 | 10 | 40 | 0 | 0 | 10 | 0 | 0 | 0 |
| 5954 | 1545 | 1 | 2 | 100 | 40 | 60 | 0 | 0 | 0 | 0 | 0 | 0.05 |
| 5956 | 2370 | 0.2 | 2 | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1.5 |
| 5958 | 2996 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 266 |
| 5960 | 357 | 8.5 | 11 | 96 | 26 | 3 | 9 | 13 | 17 | 3 | 0 | 0.1 |
| 5961 | 765 | 4.6 | 9 | 82 | 30 | 5 | 4 | 8 | 42 | 3 | 0 | 0.2 |
| 5962 | 1090 | 8 | 7 | 92 | 13 | 67 | 0 | 0 | 8 | 0 | 0 | 0 |
| 5963 | 1484 | 0.5 | 6 | 90 | 10 | 10 | 20 | 10 | 40 | 0 | 0 | 0.2 |
| 5964 | 2446 | 0.3 | 3 | 80 | 0 | 0 | 0 | 0 | 20 | 0 | 0 | 2 |
| 5965 | 1994 | 0.3 | 1 | 100 | 0 | 100 | 0 | 0 | 0 | 0 | 0 | 0.2 |
| 5968 | 350 | 3 | 10 | 83 | 17 | 7 | 4 | 22 | 0 | 15 | 0 | 0 |
| 5967 | 426 | 0.4 | 2 | 100 | 57 | 0 | 0 | 0 | 43 | 0 | 0 | 6 |
| 5966 | 715 | 1 | 6 | 7 | 27 | 22 | 0 | 0 | 27 | 6 | 0 | 0.1 |
| 5973 | 209 | 1 | 6 | 94 | 25 | 37 | 12 | 0 | 0 | 0 | 6 | 0 |
| 5972 | 210 | 50 | 10 | 89 | 28 | 11 | 12 | 27 | 0 | 0 | 0 | 0 |
| 5944 | 217 | 2.6 | 12 | 62 | 9 | 22 | 30 | 1 | 0 | 7 | 3 | 0.1 |
| 5970 | 239 | 16 | 6 | 94 | 45 | 20 | 9 | 0 | 0 | 4 | 0 | 2 |
| 5969 | 277 | 0.5 | 6 | 75 | 0 | 54 | 0 | 0 | 0 | 8 | 0 | 0 |
Пробы промыты через сито с размером ячейки 0.05 мм. Обработка проб велась под микроскопом Leika WILD M3C с увеличением в 70 раз. При этом подсчитывали общее количество раковин без отдельного определения живых особей.
Для более глубокого понимания закономерностей распределения фауны фораминифер мы рассматривали отдельно фауну в западном (пролив Вилькицкого и район восточнее пролива, до 120° в.д: cтанции 5960, 5961, 5962, 5963, 5964, 5965, 5966, 5967, 5968, 5973, 5972, 5944, 5970, 5969) и восточном регионах моря (восточнее 120° в.д: cтанции 5947, 5949, 5950, 5954, 5956, 5958), которые отличаются по геоморфологическим и гидрологическим характеристикам. В западном регионе проанализировано двадцать восемь проб с глубины от 209 до 2446 м. В восточном, характеризующемся более разнообразным рельефом, проанализировано двенадцать проб, эти пробы охватывают больший диапазон глубин от 72 до 2996 м (рис. 1).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Бентосные фораминиферы. Численность раковин бентосных фораминифер в море Лаптевых повсеместно низкая (рис. 2), наибольших величин число фораминифер достигает в западном районе моря на шельфе пролива Вилькицкого, куда проникают соленые и несколько более теплые атлантические воды. Численность фораминифер местами поднимается до 23 экз/г на глубине 277 м в слое 0–5 см, а в слое 0–2 см – до 50 экз/г на глубине 210 м, но обычно численность фораминифер составляет не более 3–9 экз/г (табл. 2, 3). На континентальном склоне в западном районе моря максимальное число фораминифер местами достигает 8.5 экз/г (слой 0–2 см) и до 21 экз/г (слой 0–5 см), но обычно не превышает 3 экз/г. Число видов в западном районе (13 видов) больше, чем в восточном (8 видов).
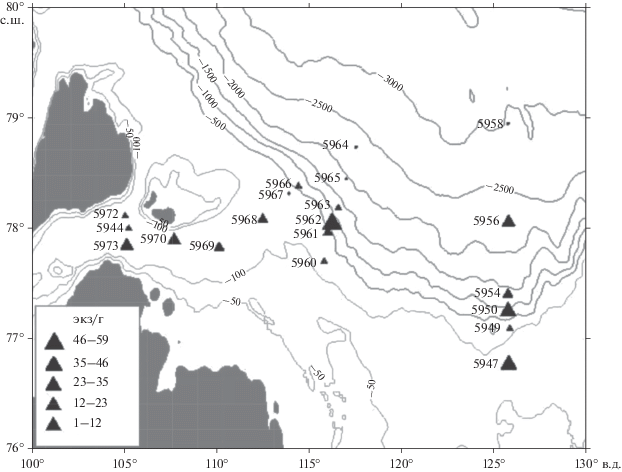
В восточном районе моря на внутреннем шельфе число фораминифер – 19.5–35 экз/г, на континентальном склоне число фораминифер низкое – не более 5–7 экз/г, обычно менее 1 экз/г, глубже, на абиссальной глубине, в интервале 2500–2996 м бентосные фораминиферы исчезают (табл. 2, 3). Сопоставление численности фораминифер показывает, что как в шельфовой зоне, так и на континентальном склоне и абиссали численность фораминифер выше в западной части моря Лаптевых по сравнению с восточной. Согласно зональной классификации, предложенной Тамановой [17], в районе наших исследований развивается комплекс фораминифер, связанный с трансформированными атлантическими водами. При этом поступление атлантических вод в западную часть моря более интенсивное, чем в восточную [5].
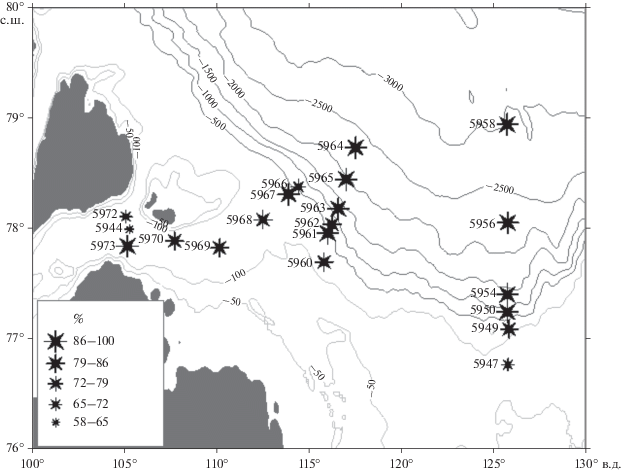
Всего в исследованном районе найдено 50 бентосных видов: 27 агглютинирующих и 23 секреционных известковых. Учитывали только виды, которые представлены в количестве более одного процента. Подавляющее большинство экземпляров представлено агглютинирующими формами (рис. 3). На их долю обычно приходится в суммарном числе экземпляров от 77 до 100%, только в пробе 5947 на внутреннем шельфе восточного района моря (глубина 72 м, слой 0–2 см доля агглютинирующих форм составляет 46%, в слое 0–5 см немного выше – 65%). Глубже и вплоть до 2996 м в восточном районе комплекс почти полностью состоит из агглютинированных раковин (90–100%). И в восточном, и в западных районах моря наибольшая масса фораминифер приурочена к шельфовым глубинам, местами относительно высокое число раковин обнаруживается на континентальном склоне.
Рис. 3.
Распределение агглютинирующих форм бентосных фораминифер в поверхностном слое донных осадков, %.
Видовое разнообразие бентосных фораминифер в море Лаптевых невысокое. На шельфе в одной пробе обычно встречается от 6 до 14 видов, на континентальном склоне число снижается до 2–8, а на глубине абиссали – 1–2 вида. Об экологии агглютинирующих видов известно немного [27, 28, 30]. Некоторые из них обитают на дне моря и питаются сестоном, другие обитают внутри осадка на большей или меньшей глубине и выбирают из него нужную еду. Но достоверных сведений об агглютинирующих видах моря Лаптевых пока мало. В большинстве статей, посвященных морю Лаптевых, сообщается только о том, в каких районах и на каких глубинах найден тот или иной вид фораминифер. В нашем материале постоянно преобладают агглютинирующие виды: представители рода Reophax, в основном R. scorpiurus, R. сurtus и Cribrostomoides subglobosus. По форме их раковин можно предположить, что они принадлежат инфауне, обитают внутри осадка на глубине 2–18 см и питаются детритом и бактериями [24–26, 29]. Реже в море Лаптевых встречаются агглютинирующие виды Adecotrima glomeratum, Nodosinella pseudonodulosa, Trochammina nana, но иногда они составляют заметную долю, особенно в комплексах западного района.
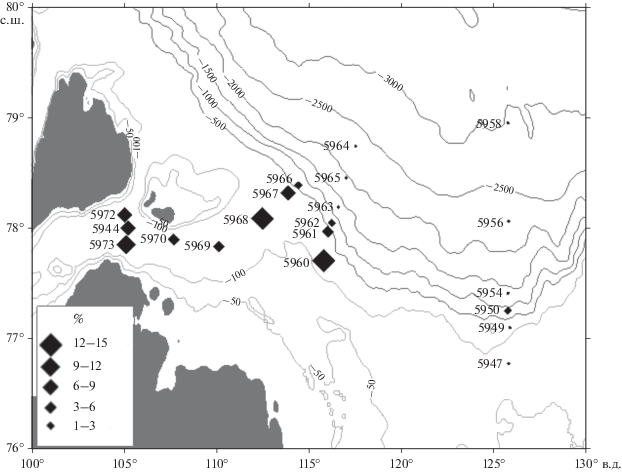
На шельфе и континентальном склоне в западной части моря постоянно встречается известковый вид Melonis barleеanus (рис. 4), который составляет до 10–15%, в восточном районе он отсутствует. Об этом виде известно, что он активно развивается в условиях, когда придонные воды имеют нормальную морскую соленость. Melonis barleеanus часто аккумулируется в шельфовых депрессиях и на континентальном склоне, как и на более глубоких участках, которые отличаются относительно высокой скоростью седиментации и где накапливается тонкий органический осадок с повышенным содержанием сравнительно старого, частично разложившегося морского органического вещества [10]. Эти участки характеризуются устойчивым поступлением пищи. Вид относится к группе инфауны, обитает в поверхностном слое осадка, питается детритом. Он широко распространен в Cеверной Атлантике, в Баренцевом и Карском морях [27–39].
Рис. 4.
Распределение вида бентосных фораминифер Melonis barleеanus в поверхностном слое донных осадков, %.
Значительно реже в море встречаются раковины других известковых видов: Elphidium clavatum, E. incertum, Cassidulina reniforme – эти виды обитают на глубинах внутреннего шельфа и предпочитают слегка опресненные воды, они изредка встречаются и на внешнем шельфе моря Лаптевых. В исследованном районе они составляют 1–3%, редко попадаются и на внешнем шельфе, где их присутствие, возможно, объясняется ледовым переносом. Следует отметить находки в шельфовом комплексе и на континентальном склоне единичных раковин известкового вида Cassidulina teretis, составляющего в некоторых пробах комплекса от 3 до 8%. Редкое присутствие этого вида свидетельствует о неблагоприятных условиях для его жизни.
Планктонные фораминиферы. Часто в исследуемом районе попадаются мелкие окатанные раковины, принадлежащие полярному виду Neogloboquadrina pachyderma sin. Возможно, они занесены течениями из Атлантического океана или льдами из Арктики. В единственной пробе восточного района 5958 на абиссальной глубине 2996 м число раковин этого полярного вида возрастает до нескольких сотен экз/г в слое 0–5 см и до двух с половиной сотен экз/г в слое 0–2 см. Эта масса раковин представлена и мелкими (ювенильными), и крупными (взрослыми) особями обычно хорошей сохранности, и мы считаем, что на этих глубинах (около 3000 м) раковины вида поступают на дно моря из поверхностных вод именно этого района. Очень редко вместе с полярным видом в тех же ассоциациях находятся раковины субполярных видов Globigerina bulloides d′Orbigny и Turborotalia quinqueloba (Natland), они, скорее всего, принесены течениями из Атлантического океана.
ЗАКЛЮЧЕНИЕ
Проанализирована фауна бентосных фораминифер из северного региона моря Лаптевых, одного из самых суровых по климату водоемов Арктики. Численность раковин бентосных фораминифер в море Лаптевых повсеместно низкая.
Наибольших величин численность фораминифер достигает в западном районе моря на шельфе пролива Вилькицкого. Подавляющее большинство экземпляров и на западе, и на востоке моря представлено агглютинирующими формами. На их долю обычно приходится от 77 до 100% от общего числа экземпляров. В восточном районе комплекс почти полностью состоит из агглютинированных раковин (90–100%). На шельфе и континентальном склоне в западной части моря постоянно встречается единственный известковый вид Melonis barleеanus, который составляет до 10–15% в комплексе. Часто в исследуемом районе попадаются единичные мелкие окатанные раковины, принадлежащие полярному виду Neogloboquadrina pachyderma sin. Но только в одной пробе восточного района 5958 на абиссальной глубине (около 3000 м) число раковин этого вида достигает 300 экз/г в слое 0–5 см и 266 экз/г в слое 0–2 см. Эта масса раковин представлена и мелкими (ювенильными), и крупными (взрослыми) особями обычно хорошей сохранности, принадлежащими к этому же полярному виду. Массовое присутствие полярного планктонного вида Neogloboquadrina pachyderma sin на абиссальной глубине восточного района указывает на доминирующее влияние арктических вод в данном регионе.
Благодарности. Авторы благодарят заведующего Лабораторией донной фауны океана Института океанологии им. П.П. Ширшова РАН (ИО РАН) А.В. Гебрука и ведущего научного сотрудника ИО РАН С.А. Корсуна за ценные советы при подготовке рукописи. Мы благодарны научному сотруднику ИО РАН В.И. Гагарину за собранные пробы и ведущему научному сотруднику ИО РАН С.Г. Пояркову за консультацию по гидрологии региона.
Источники финансирования. Работа поддержана бюджетными темами Госзадания Института океанология РАН №№ 0149-2019-0007 (Т.А. Хусид), 0149-2019-0006 (Н.В. Либина), 0149-2019-0008 (М.М. Доманов).
Таксономический список видов, встреченных в море Лаптевых
Планктонные фораминиферы
Neogloboquadrina pachyderma sin. Ehrenberg
Globigerina bulloides d′Orbigny
Turborotalia qunrnqueloba (Natland)
Бентосные фораминиферы
Ammobaculites sp.
Adercotryma glomerata (Brady)
Astrononion gallowayi Loblich and Tappan
Ammodiscus incertum (d′Orbigny)
Ammotium cassis (Parker)
Ammolagena clavatum (Parker and Jones)
Bathysiphon rufus d′Folin
Buсcella frigida (Cushman)
Buccella inusitata Andersen
Bucella tenerrima (Brady)
Bulimina exilis Brady
Cassidulina reniforme Norvang
Cassidulina teretis Tappan = Cassidulina neoteretis Seidenkrantz
Cibicides lobatulus (Walker and Jones)
Cribrostoides subglobosus (Cushman)
Dentalina frobisherensis Loeblich and Tappan
Eggerella advena (Cushman)
Elphidium clavatum Loeblich and Tappan
Elphidium incertum (Williamson)
Epistominella arctica Green
Fissurina orbignynyana Seguenza
Globocassidulina subglobosa (Brady)
Haynesina orbiculare (Brady)
Hormosina normani Brady
Islandiella norcrossi (Cushman)
Lagena strata (d′Orbigny)
Melonis barleeanus (Williamson)
Nodosaria emphysiocuta Loeblich and Tappan
Nodosinella pseudonodulosa Shtschedrina
Nonionionella labradoricum (Dawson)
Quinqueloculina seminulum (Linne)
Parafissurina fusuliformis Loeblich and Tappan
Proteonella sp.
Recurvoides contortus Earland
Reophax curtus Cushman
Reophax scorpiurus d′Montfort
Rosalina vilardeboana d′Orbigny
Saccorhiza ramosa (Brady)
Stainforthia loeblichi Feyling-Hansen
Trifarina fluens (Todd)
Trochammina nana (Brady)
Список литературы
Атлас океанов. Северный Ледовитый океан. Л.: ГУНИО, 1980. 82 с.
Аксенов А.А., Дунаев Н.Н., Ионин А.С. и др. Арктический шельф Евразии в позднечетвертичное время. М.: Наука, 1992. 272 с.
Грачев А.Ф., Деменицкая Р.М., Карасик А.М. Срединный Арктический хребет и его материковое продолжение // Геоморфология. 1970. № 1. С. 42–45.
Дмитренко И.А., Хьюлеманн Й.А., Кириллов С.А. и др. Термический режим придонного слоя моря Лаптевых и процессы, его определяющие // Криосфера Земли. 2001. № 3. С. 40–55.
Добровольский А.Д., Залогин Б.С. Моря СССР. М.: Изд-во МГУ, 1982. 192 с.
Зенкевич Л.А. Биология морей СССР. М.: Изд-во АН СССР, 1963. 739 с.
Каплин П.А. Новейшая история побережий Мирового океана. М.: Изд-во МГУ, 1973. 265 с.
Кордиков А.А. Осадки моря Лаптевых. Л.–М.: Изд-во Главсеврморпути, 1952. Тр. НИИГА. Т. ХI. 132 с.
Лукина Т.Г. Особенности распределения фораминифер в море Лаптевых // Исследования фауны морей. 2004. Т. 54(62). С. 86–123.
Матуль А.Г, Хусид Т.А., Мухина В.В. и др. Современные и позднеголоценовые природные условия на шельфе юго-восточной части моря Лаптевых по данным микрофоссилий // Океанология. 2007. Т. 47. № 1. С. 90–101.
Овсепян Я.С., Талденкова Е.Е., Баух Х.А., Кандиано Е.С. Реконструкция событий позднего плейстоцена–голоцена на континентальном склоне моря Лаптевых по комплексам бентосных и планктонных фораминифер // Стратиграфия. Геологическая корреляция. 2015. Т. 23. № 6. С. 96–112.
Павлидис Ю.А. Шельф Мирового океана в позднечетвертичное время. М.: Наука, 1992. 272 с.
Павлидис Ю.А., Ионин А.С., Щербаков Ф.А. и др. Арктический шельф. Позднечетвертичная история как основа прогноза развития. М.: ГЕОС, 1998. 187 с.
Панов В.В., Шпайхер А.Д. Роль постгляциальных атлантических вод в формировании гидрометеорологического режима Арктических морей // Труды ААНИИ. 1963. 264 с.
Система моря Лаптевых и прилегающих арктических морей: современные условия и палеоклимат / Под ред. Кассенс Х., Лисицына А., Тиде Й. и др. М.: Изд-во моск. ун-та, 2009. 260 с.
Талденкова Е.Е., Баух Х.А., Степанова А.Ю. и др. Изменения палеосреды шельфов морей Лаптевых и Карского в ходе послеледниковой трансгрессии (по ископаемой бентосной фауне) // Система моря Лаптевых и прилегающих арктических морей: современные условия и палеоклимат. М.: Изд-во моск. ун-та, 2009. С. 384–409.
Таманова С.В. Видовой состав современных фораминифер как индикатор гидрологического режима арктических морей // Северный Ледовитый океан и его побережье в кайнозое. Л.: Гидрометеоиздат, 1970. С. 199–203.
Шипилов Э.В. Рифтогеоэкология континентальных окраин Евразиатско-Арктической окраины: Автореф. дисс. ... докт. геогр. наук. М.: МГУ, 1993. 85 с.
Щедрина З.Г. К фауне корненожек Полярных морей СССР // Труды Арктического института. 1936. Т. 33. С. 51–64.
Bauch H.A., Kubisch-Popp M.K., Cronin T.M., Rossak B.A. Study of the calcareous microfauna from Laptev Sea sediments // Berichte zur Polarforschung. 1995. V. 176.
Bauch H.A., Kassens H., Erlenkeuser H. et al. Deposition environments of the Laptev Sea (Arctic Siberia) during the Holocene // Boreas. 1999. V. 28. P. 194–204.
Bauch H.A., Cremer H., Kunc-Pirrung M. Siberian shelf sediments contain clues to paleoclimate forcing // EOS Transactions. 2000. V. 81. P. 233–238.
Bauch H.A., Mueller-Lupp-Lupp T., Taldenkova E. et al. Chronology of Holocene transgression at the North Siberian margin // Global and Planetary Change. 2001. V. 31. P. 125–139.
Corliss B.H. Microhabitats of benthic foraminifera within dep-sea sediment // Nature. 1985. V. 314. № 6010. P. 435–438.
Corliss B.H., Chen C. Morphotype patterns of Norwegian Sea deep-sea benthic foraminifera and ecological implications // Geology. 1988. V. 16. P. 716–719.
Feyling-Hanssen R., Joregensen J., Knudsen K., Andersen A. Late Quaternary from Vendsyssei, Denmark and Sandnes, Norway // Bull. Geol. Soc. Den. 1971. Part 2–3. 316 p.
Hald M., Steinsund P.I. Distribution of surface sediment benthic foraminifera in the southwestern Barents Sea // J. Foraminiferal Res. 1992. V. 22. № 4. P. 347–362.
Korsun S.A. Benthic foraminifera in the Ob estuary, West Siberia // Berichte zur Polarforrschung. 1999. № 300. P. 59–70.
Korsun S.A., Polyak L.V. Distribution of benthic Foraminiferal morphogroups in the Barents Sea // Oceanology. 1989. V. 29. P. 838–844.
Loeblich A., Tappan H. Studier of Arctic Foraminifera // Smithsonian Miscellaneous Collections. 1953. V. 121. 150 p.
Mackensen A., Sejrup H.P., Janssen E. The distribution of living benthic foraminifera on the continental slope and rise of southwest Norway // Marine Micropaleontology. 1985. V. 9. P. 275–306.
Mackensen A., Schumacher S., Radke J., Schmidt D.N. Microhabitat preferences and stable carbon isotopes of epibenthic foraminifera: clue to quantitative reconstruction of oceanic: new production? // Marine Micropaleonology. 2000. V. 40. P. 233–258.
Mudie R.J., Keen C.E., Hardy I.A. Vilks G. Multivariate analysis and quantitative paleoecology of benthic foraminifera in surface and late Quaternary shelf sediments, northern Canada // Marine Micropaleontology. 1983. V. 8. P. 283–313.
Polyak L., Korsun S., Febo L. et al. Benthic foraminiferal assemblages from the Southern Kara Sea, a river-influenced Arctic marine environment // J. Foraminiferal Res. 2002. V. 32. № 3. P. 252–273.
Rasmussen T.L., Thomsen E., Slubovska M.A. et al. Paleoceanographic evolution of the SW Svalbard margin (76° N) since 20 000 14C yr BP // Quaternary Res. 2007. V. 67. P. 100–114.
Rasmussen T.L., Thomsen E. Ecology of deep-sea benthic foraminifera in the North Atlantic during the last glaciation: Food and temperature control // Palaeogeogr., Palaeoclimatol., Palaeoecol. 2017. V. 472. P. 15–32.
Stepanova A., Taldenkova E., Bauch H.A. Recent Ostracoda of the Laptev Sea (Arctic Siberia): taxonomic composition and some environmental implications // Marine Micropaleontology. 2003. V. 48(1–2). P. 23–48.
Volkmann R. Planktic foraminifers in the outer Laptev Sea and the Fram Strait – Modern distribution and ecology // J. Foraminiferal Res. 2000. V. 30. P. 157–176.
Wollenburg J., Mackensen A. Living benthic foraminifers from the central Arctic Ocean faunal composition, standing, stock, and diversity // Marine Micropaleontol. 1998. V. 34(3–4). P. 153–185.
Дополнительные материалы отсутствуют.