

Океанология, 2021, T. 61, № 5, стр. 759-768
Оценка продуктивности плантаций двустворчатых моллюсков на основе результатов моделирования
С. В. Катрасов 1, *, А. Н. Бугаец 1, В. В. Жариков 1, С. И. Масленников 2, В. Н. Лысенко 3, Ю. А. Барабанщиков 3, П. Я. Тищенко 3
1 Тихоокеанский институт географии ДВО РАН
Владивосток, Россия
2 Национальный научный центр морской биологии им. А.В. Жирмунского ДВО РАН
Владивосток, Россия
3 Тихоокеанский океанологический институт им. В.И. Ильичева ДВО РАН
Владивосток, Россия
* E-mail: sergey_katrasov@mail.ru
Поступила в редакцию 26.02.2021
После доработки 10.03.2021
Принята к публикации 08.04.2021
Аннотация
С использованием модели FARM, основанной на учете биологических и гидрохимических параметров, выполнена оценка потенциальной продуктивности плантаций устрицы и мидии в бухте Воевода (о. Русский, Приморский край). Основным источником первичной продукции в районе исследования являются заросли морских трав (Zostera marina) и фитопланктон. Параметры продуцирования органического вещества для каждой расчетной ячейки смоделированы с помощью модуля ECO BLOOM II Algae модели DELWAQ программного пакета Delft3D. Перераспределение органического вещества по акватории бухты оценивалось по результатам моделирования гидродинамики средствами DELFT3D Flow. Полученные результаты были использованы в качестве исходных данных в FARM для расчета обеспеченных значений продуктивности плантаций гигантской устрицы (Crassostrea gigas) и тихоокеанской мидии (Mytilus trossulus) в бухте Воевода. Построены карты пространственного распределения обеспеченных значений модельной продуктивности придонных и садковых плантаций.
ВВЕДЕНИЕ
Среди всех прибрежных районов Дальнего Востока природные условия для культивирования гидробионтов наиболее оптимальны в заливе Петра Великого и южном Приморье. Именно здесь с конца 60-х гг. прошлого века началось развитие отечественной промышленной марикультуры. В настоящее время происходит процесс формирования и распределения рыбоводных участков, и приоритетными задачами развития отрасли становятся разработка методов определения наиболее подходящих районов для создания плантаций, оценка площадей акваторий, пригодных для выращивания того или иного вида, и прогнозирование реальных объемов продукции [20, 24, 25]. При этом основным направлением, способным обеспечить существенный рост продукции марикультуры, является культивирование двустворчатых моллюсков [7].
До настоящего времени при планировании марикультурной деятельности выполнялись лишь экспертные оценки потенциала районов, основанные на анализе физико-географических условий выращивания, самых общих продукционных характеристиках вод и представлениях о трофических потребностях выращиваемых объектов [4, 6]. При этом очевидно, что продуктивность марикультуры тесно связана с функционированием вмещающих экосистем и зависит от комплекса природных факторов. В этом контексте серьезную проблему представляет учет гидро- и геодинамическими параметров, процессов производства первичной продукции, трансформации и транспорта органического вещества, определяющих трофическую базу выращивания гидробионтов в районах размещения плантаций [34].
В масштабе экосистемы производство автохтонного и поступление аллохтонного органического вещества – доминирующие факторы устойчивого функционирования плантаций двустворчатых моллюсков [36, 41]. В локальном масштабе состояние поселений зависит от физических ограничений, таких как присутствие подходящих субстратов и местообитаний, наличие течений, транспортирующих пищу, концентрации взвешенного органического углерода в водной массе и поверхностном слое осадков. Соответственно, выживаемость и смертность культивируемых гидробионтов, связанные с динамикой условий выращивания, также являются ключевыми факторами урожайности плантаций и экономической конкурентоспособности продукции [34, 35, 40].
При определении эксплуатационной емкости района для марикультуры двустворчатых моллюсков принято выделять: (1) физическую емкость – общую площадь морской акватории, пригодную для размещения мариферм по абиотическим показателям (температура, соленость, концентрация растворенного кислорода и т.п.) и (2) продуктивную емкость, определяемую через потенциальную продуктивность с учетом биологических показателей (концентрация органического углерода, хлорофилла и т.п.). Физическая емкость может быть оценена с помощью комбинации гидродинамических моделей и эколого-ландшафтной информации, представленной в ГИС и обработанной средствами пространственного анализа. Для оценки продуктивной емкости разработаны и успешно применяются численные модели, созданные на основе эмпирических данных и экспериментальных подходов [39, 42].
Потенциальная продуктивность выделенных районов рассчитывается на основе оценки доступности питательных веществ как функции количества первичной продукции и физиологических подмоделей, имитирующих возможности роста в зависимости от количества пищи и физиологических характеристик выращиваемых видов. Эксплуатационная емкость с точки зрения размера и качества товарных особей может зависеть от принятых критериев для товарных классов и размеров регионального рынка моллюсков и может значительно отличаться в различных районах выращивания.
На Дальнем Востоке России проблемы оптимального размещения объектов марикультуры в зависимости от гидрологических факторов среды остаются недостаточно изученными. Актуальной задачей является создание соответствующей информационной базы как основы региональной программы комплексного управления прибрежной зоной, включающей, в том числе, развитие марикультуры [6]. При выборе объектов культивирования предпочтение отдается моллюскам, обладающим высокой потенциальной продуктивностью, незначительной смертностью на разных стадиях развития, высоким содержанием мяса в особи [26] и при этом способным переносить резкие колебания выростной среды [2, 27]. Всем этим требованиям отвечают гигантская устрица (Crassostrea gigas) и тихоокеанская мидия (Mytilus trossulus).
В данном исследовании, для оценки продуктивности придонных и садковых плантаций устрицы и мидии в бухте Воевода, средствами программного комплекса DELWAQ (DELft WAter Quality) [33] проведено численное моделирование продуцирования и перераспределения первичной продукции в результате воздействия гидродинамических факторов. Результаты моделирования использовались в модели FARM (Farm Aquaculture Resource Management) [35] для расчета продуктивности для садковых и придонных плантаций C. gigas и мидии M. trossulus. C учетом региональных рекомендаций по выращиванию гидробионтов построены карты пространственного распределения обеспеченных значений продуктивности плантаций.
МАТЕРИАЛЫ И МЕТОДЫ
Среди приостровных акваторий залива Петра Великого бухта Воевода считается одним из наиболее перспективных районов для марикультурной деятельности. Она расположена на западном побережье о. Русский (рис. 1), имеет площадь около 4 км2 и включает в себя две бухты второго порядка – Круглую (1.6 км2) и Мелководную (2.4 км2). Климат в районе исследований умеренный муссонный, зимой преобладают северные ветра, обуславливающие холодную, ясную погоду. В теплый период года преобладают южные ветра с Тихого океана. По данным Приморского УГМС [31] за период моделирования (1989–2019) по данным метеостанции “Владивосток” (31 960) безморозный период длится в среднем 190 дней, среднегодовое количество осадков составляет 848 мм. Среднемесячная относительная влажность воздуха составляет 58–91%. Минимальная, максимальная и средняя температура воздуха составляют –25.2, 32.8 и 9.2°С соответственно. Преобладающим направлением ветра является северное (37%). Средняя скорость ветра 5.8, максимальная 36.0 м/с.

Берега, прилегающие к входным мысам бухты Воевода, возвышенные и приглубые, берега внутренней части бухты низкие, песчаные и отмелые [16]. Водообмен в бухтах Воевода определяется циклонической циркуляцией течений [3], средняя величина прилива составляет 0.2 м с максимальной амплитудой 0.43 м [10]. Стационарные гидрологические наблюдения в бухте не велись. Сезонные гидролого-гидрохимические исследования проводились в 2011–2012 гг. [3]. Имеющиеся в литературе данные по распределению гидробионтов и условиям их обитания в бухте Воевода малочисленны [8, 18].
C. gigas обладает высокой экологической пластичностью и высокими темпами роста [23]. Это эвригалинный вид, живущий при солености не ниже 12 PSU [12], но выдерживающий краткосрочное опреснение до 5 PSU. При этом границы оптимального для устрицы диапазона солености находятся в интервале 23–28 PSU [13, 29]. В заливе Петра Великого устрицы обычно обитают на глубинах 0.5–7 м, местами образуя сплошные поселения (банки, устричники) на илисто-песчаных и скалистых грунтах [22]. По данным водолазного обследования бухты Воевода установлено, что основные устричники находятся в кутовой, хорошо прогреваемой части (б. Мелководная) [14]. Вне плотных скоплений C. gigas формирует небольшие друзы, состоящие из одной крупной особи, обросшей более мелкими, плотность которых колеблется от 0.01 до 1 экз./м2.
M. trossulus – широко распространенный вид мидий, обычный в дальневосточных морях России [19, 43]. Хотя поселения мидий обнаружены до глубины 40 м [15], преимущественно вид обитает в литорально-сублиторальной зоне на скалистых и каменистых грунтах, до глубины 2.5–3 м [21]. Значительно реже мидия селится на песчаных, галечно-гравийных и песчано-илистых грунтах, где прикрепляется к камням и к раковинам других моллюсков, образуя друзы. В заливе Петра Великого наиболее благоприятная температура воды для роста и развития мидий 10–18°С, соленость – 27–33 PSU [11]. Поскольку основной способ питания двустворчатых моллюсков – фильтрация, среди важных абиотических условий выращивания этих видов указывается наличие незначительных скоростей течений (0.02–0.05 м/с) [13, 28].
Модель управления ресурсами аквакультуры на фермах FARM (Farm Aquaculture Resource Management, www.farmscale.org [34]) предназначена для решения задач, связанных с предварительным анализом размещения плантаций и выбором культивируемых видов, экономической оптимизации практики культивирования, а также экологической оценкой воздействия хозяйств на окружающую среду. В рамках FARM для определения продукции и оценки эвтрофикации за периоды культивирования применяется комбинация физических, биогеохимических моделей и моделей роста моллюсков. FARM позволяет рассчитать урожайность гидробионтов на основе данных о геометрических размерах марифермы, плотности моллюсков, локальных трофических условиях выращивания и параметрах окружающей среды.
Входные данные задаются с суточным или большим временным разрешением (в последнем случае производится линейная интерполяция данных в суточные интервалы). Элиминация гидробионтов, связанная с не учитываемыми в модели факторами, задается как параметр. Результатом расчетов модели являются временные ряды концентраций хлорофилла-а (Chl-a), растворенного кислорода (dissolved oxygen, DO), концентрации взвешенных твердых частиц (total particulate matter, TPM) и твердых частиц органического вещества (particulate organic matter, POM), биомассы гидробионтов на каждый расчетный день.
Продукционные характеристики (Chl-a, POM, TPM) рассчитаны с помощью биогеохимической модели DELWAQ (DELft WAter Quality) [33], входящей в программный комплекс Delft3D. Библиотека DELWAQ ECO позволяет моделировать процессы эвтрофикации, динамику биогенов, содержание кислорода, взвешенных веществ и наносов, а также фитопланктона в поверхностных водах. При моделировании водного баланса и гидродинамического переноса веществ поля динамики задаются отдельно с помощью импорта результатов расчетов гидродинамических моделей (в данном случае, Delft3D Flow).
Временной шаг моделирования DELWAQ составляет 24 ч. Водоросли (источник первичной продукции) поглощают питательные вещества из воды для производства клеточного материала. В результате фотосинтеза водоросли производят кислород. Концентрация Chl-a пропорциональна концентрации биомассы планктонных водорослей. Значительная часть биомассы расположенных в толще воды микроводорослей потребляется зоопланктоном (скорость поедания водорослей учитывается через показатель смертности) и переходит в детрит, оседающий на дно в верхний слой отложений, где происходит бактериальное разложение. Для моделирования первичной продукции фитопланктона в ECO используется модуль BLOOM [30, 37, 38]. BLOOM вычисляет биомассу фитопланктона определенных пользователем групп водорослей в зависимости от ограничивающих факторов.
МОДЕЛИРОВАНИЕ ПРОДУКЦИОННЫХ ХАРАКТЕРИСТИК
В работе использованы поля гидродинамики, рассчитанные ранее с помощью трехмерной модели Delft3D Flow за период 01.01.1988–01.01.2019 гг. [14]. Метеорологическое воздействие на поверхность акватории бухты задано на основе архивных данных гидрометеорологических наблюдений на метеостанции “Владивосток” (WMO 31960) [31]. Горизонтальное разрешение модельной сетки 20 м. Вертикальное разрешение сетки составляет 3 σ-слоя, границы которых установлены в 10% от поверхности и от дна бухты. Скорость и направление ветра заданы с дискретностью 3 ч, остальные метеопараметры с суточным разрешением. На открытой границе, расположенной на входе в бухту, заданы параметры гармонических постоянных прилива по данным поста Владивосток [10], средняя температура и соленость воды – по данным сезонных измерений ТОИ ДВО РАН [3], содержание кислорода – как постоянное значение (9.21 мл/л) [1].
Основным источником первичной продукции в районе исследования являются поля морских трав (Zostera marina) и планктонные диатомовые водоросли [3]. При этом концентрация фитопланктона в зарослях трав значительно выше, чем в толще воды [17]. С учетом этого на модельной расчетной сетке были выделены ячейки, входящие в области произрастания зостеры, оконтуренные на основании результатов водолазного обследования бухты [14].
Перечисленные ниже входные параметры модели, необходимые для включения первичной продукции зостеры в блок расчета концентрации и переноса компонентов производства первичной продукции от дна (зостеры) в верхние слои, откуда они дальше перераспределяются по акватории бухты в результате гидродинамического воздействия, были заданы по литературным данным [5, 17, 32, 37, 38] как постоянные значения на каждый расчетный шаг (сутки) для всего периода моделирования. Средний вертикальный водообмен ячеек расчетной сетки, расположенных в районах произрастания морских трав – 0.0017 м3/с. Продуцируемый зостерой кислород – 6 мг/л [32]. Коэффициенты стехиометрии морских диатомовых водорослей заданы для трех, определенных в модуле ECO фенотипов: энергетический (MDIATOMS_E) – 0.24 гC/м3, азотный (MDIATOMS_N) – 0.21 гC/м3 и фосфорный (MDIATOMS_P) – 0.21 гC/м3 по данным работ [38]. Концентрации: (1) частиц твердого неорганического вещества в воде IM1 – 7.9 г/м3; (2) твердых частиц органического углерода в воде POC1 – 33 гC/ м3; (3) биомасса подводной растительности SM1 – 0.003412 гC [5].
Расчет концентраций Chl-a и детрита выполнен с использованием модулей DELWAQ ECO: “Macrophyte submerged 01” и “Marine Diatoms”. Для каждой расчетной ячейки получены суточные значения продуцирования органического вещества зостерой и фитопланктоном в единицах органического углерода (гC), растворенного кислорода (DO, мг/л) и суммарной концентрации твердого неорганического (TPM, гC) и органического вещества (POM, гC) с учетом их перераспределения по району исследования под воздействием гидродинамических факторов.
Выходные данные DELWAQ ECO были осреднены по площади 100 × 100 м (5 × 5 расчетных ячеек), что соответствует оптимальному пространственному масштабу, моделирования FARM [34], и подготовлены в виде суточных рядов в формате входных данных модели. При расчете потенциальной продукции гидробионтов были заданы следующие товарные характеристики для устрицы: вес – 100 г, товарный размер – 12 см; для мидии вес – 12 г, размер – 6 см. Периоды начала культивирования мидии и устрицы заданы в соответствии с региональными рекомендациями [11, 12] – 01.06 и 01.08 соответственно. Продолжительность периода культивирования для обоих видов задана одинаково – 22 мес. Плотность посадки спата: (1) донное выращивание мидии и устрицы – 100 экз/м2, (2) садковое выращивание: мидия – 420 экз/м2, устрица – 200 экз/м2.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Полученные в результате моделирования продукционных характеристик данные позволили с помощью FARM выполнить расчет для 28 рекомендованных региональной биотехнологией культивирования 22-месячных периодов выращивания рассматриваемых гидробионтов [11, 12]. Для расчета продуктивности донного выращивания использованы данные придонного слоя, для расчета продуктивности садкового выращивания были заданы параметры среднего слоя модели Delft3D Flow.
Пространственное распределение значений 75% обеспеченности концентрации модельных значений Chl-a, POM и DO представлены на (рис. 2). Анализ распределения смоделированных продукционных характеристик позволяет сделать заключение, что в районе исследований содержание растворенного кислорода не является лимитирующим фактором для производства первичной продукции и культивирования как для донного, так и для садкового способа выращивания выбранных гидробионтов. Концентрация Chl-a и POM в кутовой части (бухты Мелководная и Круглая) находятся в диапазоне оптимальных значений, ближе к центральной части бухты концентрации уменьшаются и далее в направлении входа в бухту Воевода достигают минимальных значений.
Рис. 2.
Содержание 75% обеспеченности в среднем слое: Chl-a (1), DO (2), POM (3), рассчитанные с помощью DELWAQ.

Для демонстрации и анализа результатов моделирования выделен расчетный блок 100 × 100 м, находящийся в центральной части зоны оптимального выращивания гидробионтов в б. Мелководная. Графики моделирования прироста суммарного веса гидробионтов за каждый из 28 периодов и их осредненная кривая представлены на рис. 3. Модельные данные указывают на то, что при указанных параметрах культивирования для выращивания в б. Воевода в среднем оптимальный период составляет 18–20 мес. для мидии и 12–13 мес. для устрицы. Минимальные значения прироста продукции соответствуют началу периодов выращивания в 1994, 2000, 2012 и 2016 гг. Основной причиной слабого роста является пониженные за период культивирования средние значения солености (29.6–30.5 PSU), а также воздействие сильного кратковременного распреснения (18.4–22.9 PSU), вызванное влиянием значительного притока пресных вод в акваторию бухты, вызванных тайфунами (Мелисса – 1994, Боллавен, Проперун, Саомэй – 2000, Санба, Боллавен – 2012, Лайнрок – 2016) и активным циклогенезом (2000 г.).
Рис. 3.
Графики прироста суммарного веса устрицы (а, в) и мидии (б, г) в соответствии с заданными товарными характеристиками, рассчитанными с помощью моделей прироста суммарного веса гидробионтов FARM за расчетные периоды 1989–2019 гг. в соответствии с заданными товарными характеристиками, и среднее по всем расчетным периодам, (а, б) – донное, (в, г) – садковое культивирование.
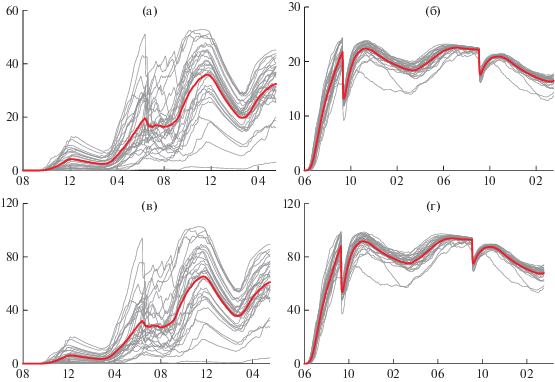
Мидии в целом дают более стабильный урожай как для донного, так и для садкового выращивания. Все 28 модельных графиков прироста биомассы показывают, что к концу 12–13-месячного периода выращивания урожайность практически не зависит от абиотических условий выращивания. Графики прироста биомассы устриц, напротив, демонстрируют значительную реакцию на отклонение условий выращивания от оптимальных. Достаточно сильные кратковременные распреснения бухты, связанные с обильными атмосферными осадками, выпадающими на акваторию и водосбор бухты, существенно влияют на продуктивность как садкового, так и донного их выращивания.
Графики прироста суммарного веса мидии в соответствии с заданными товарными характеристиками, рассчитанными с помощью модели FARM, хорошо интерпретируются сезонными особенностями роста моллюсков. Первый малый пик наблюдется в период роста моллюсков после оседания, который заканчивается с наступлением минимальных температур воды в феврале следующего года. Далее, с прогревом воды наблюдается максимальный прирост массы моллюсков, который продолжается до июня. В этот период происходит первый нерест моллюсков, с последующей потерей массы мягких тканей. Посленерестовая пауза в росте продолжается весь период высоких температур в летний период.
С охлаждением водной массы в октябре начинается период максимального роста, который продолжается также до конца февраля, то есть фактически до окончания периода низких температур морской воды, и начала весеннего прогрева. До декабря наблюдается постоянный прирост мягких тканей, что объясняется оптимальными условиями для развития моллюсков. В период осеннего охлаждения водной массы у моллюсков падают затраты на основной обмен с падением температуры воды. При этом начинается осеннее цветение фитопланктона (которое заканчивается только в конце декабря). Осеннее цветение микроводорослей меньше ограничивается выеданием зоопланктерами, которые также снижают свою активность в период снижения температуры воды.
Прирост второго года продолжается с марта по июнь, когда вновь начинается нерестовая активность и период максимального прогрева. Августовский резкий спад приростов можно объяснить усилением конкуренции за место среди второгодних моллюсков и за счет оседания на коллекторы сеголеток. Конкуренция за место особенно остро ощущается в период максимальных температур, которые наступают в конце августа. Этот же период также известен минимальной продукцией фитопланктона. На графиках мы видим резкое падение, что объясняется свалом с коллекторов на дно и гибелью наиболее слабых особей моллюсков. Потери урожая частично компенсируются последующим осенним ростом, но усиливающаяся при этом конкуренция за место на субстрате ведет к дальнейшему снижению товарного урожая.
Для выделенных расчетных элементов (100 × × 100 м) за каждый период культивирования были выбраны максимальные значения продуктивности гидробионтов на второй год выращивания. Указанные значения были ранжированы и вычислены их эмпирические обеспеченности. На рис. 4 представлены данные о пространственном распределении 50, 75 и 95% значений модельной продуктивности для донного и садкового выращивания гидробионтов в акватории бухты с учетом рекомендованных для выращивания глубин (1 и 5 м для придонного и садкового выращивания соответственно) [28].
Рис. 4.
Значение модельной продуктивности (т./га) 50, 75 и 95% обеспеченности для донного (а, с, е) и садкового (б, д, ж) выращивания устрицы (1) и мидии (2) в бухте Воевода. Выделены районы, ограниченные изобатами 1 и 5 м.
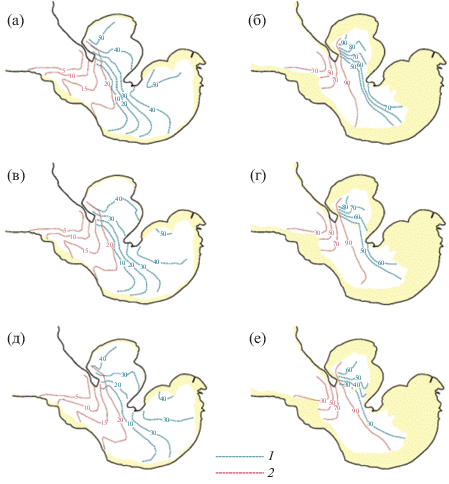
Как было показано выше, практически продуктивность мидии как для садкового, так и для придонного выращивания мало зависит от условий культивирования, изолинии обеспеченных значений продуктивности лишь немного смещаются по направлению ко входу в бухту. Для устрицы пространственное распределение обеспеченных значений продуктивности (50, 75 и 95%) существенно меняется (рис. 4). Если предположить, что значения продуктивности 95% обеспеченности можно рассматривать в качестве гарантированного объема продукции, то разница между ним и значениями обеспеченной продуктивности 75 и 50% для устрицы может достигать 10 т/га для придонного и 10–40 т/га для садкового выращивания, соответственно. Обеспеченные значения (50, 75 и 95%) продуктивности бухты по исследуемым видам двустворчатых моллюсков представлены в табл. 1.
Таблица 1.
Обеспеченные значения общей продуктивности устрицы и мидии. В числителе среднее, т/га, в знаменателе суммарное, т
| Тип выращивания | Мидия | Устрица | ||||
|---|---|---|---|---|---|---|
| 50% | 75% | 95% | 50% | 75% | 95% | |
| Придонное | $\frac{{19.27}}{{1907.5}}$ | $\frac{{18.79}}{{1860.04}}$ | $\frac{{18.1}}{{1787.4}}$ | $\frac{{30.7}}{{2483.9}}$ | $\frac{{27.0}}{{2187.5}}$ | $\frac{{18.9}}{{1530.6}}$ |
| Садковое | $\frac{{69.5}}{{3821.5}}$ | $\frac{{67.7}}{{3723.5}}$ | $\frac{{64.1}}{{3523.2}}$ | $\frac{{36.1}}{{1445.9}}$ | $\frac{{30.1}}{{1203.4}}$ | $\frac{{18.7}}{{747.6}}$ |
На основе выполненных расчетов можно заключить, что гидрологические и биологические условия бухт Мелководная и Круглая больше подходят для выращивания гидробионтов. Центральная часть бухты Воевода обладает меньшим потенциалом для культивирования двустворчатых, модельная продуктивность для обоих видов снижается по направлению ко входу в бухту, что связано с особенностями производства первичной продукции и гидродинамического режима бухты. Несмотря на оптимальные концентрации пищевых ресурсов в кутовой части, условия для выращивания устрицы хуже. Это определяется жестким гидрологическим режимом, характеризующимся резкими перепадами солености. По направлению к открытой части бухты, несмотря на меньшее количество пищевых ресурсов, наблюдаются более стабильные гидрологические условия, с меньшими сезонными и суточными амплитудами базовых параметров.
Полученные значения находятся в хорошем соответствии с интервалами значений урожайности, которые приводятся в литературных источниках по условиям культивирования и продуктивности гидробионтов в заливе Петра Великого. По данным [9, 12] средняя урожайность при двухгодичном выращивании для устрицы составляет 45 т/га, для мидии – 33–60 т/га [9, 11, 28]. В работах [6–8] приводятся данные значений продуктивности культивирования мидий в интервале 80–100 т/га. Максимальная урожайность сырца при подвесном выращивании в толще воды может достигать до 150 т/га [28].
Представленные результаты моделирования обеспеченности продукцией основаны только на учете факторов питания и гидрологического режима, которые наиболее важны на стадии выбора места установки марифермы, культивируемых видов, а также на начальном этапе выращивания гидробионтов. После строительства гидробиотехнических установок могут быть выполнены более точные оценки продуктивности с учетом вторичной эвтрофикации акватории и взаимного ограничения водообмена, обусловленного типом и пространственной конфигурацией самих установок.
Представленные картосхемы обеспеченных значений модельной продуктивности гидробионтов могут быть использованы при планировании размещения садковых и придонных плантаций, выбора видов культивирования с учетом особенностей производства первичной продукции, гидродинамическим режимом бухты, оптимальных условий по абиотическим факторам и обеспеченностью первичной продукцией. Представленные расчеты выполнены с учетом региональных рекомендаций по культивированию гидробионтов, использование отличных от указанных региональных параметров товарной продукции может так же привести к существенно отличающимся от приведенных в работе результатов моделирования.
ЗАКЛЮЧЕНИЕ
Комплексное моделирование продуктивности мариферм, учитывающее влияние притока пресных вод с водосбора водоемов, производство и перераспределение первичной продукции под влиянием гидродинамических факторов, в отечественной практике культивирования марикультуры пока применяется редко. Необходимые компьютерные технологии и информационная поддержка подобных исследований еще слабо развиты. В данной статье с помощью широко применяемых в мировой практике гидродинамической и биогеохимической моделей и модели управления ресурсами аквакультуры продемонстрирован пример технического решения, направленного на обеспечение развития региональной марикультурной деятельности. Выполнен расчет обеспеченных значений модельной продуктивности плантаций гигантской устрицы C. gigas и тихоокеанской мидии M. trossulus в бухте Воевода с учетом региональных рекомендаций по культивированию указанных гидробионтов. Полученные значения модельной продуктивности моллюсков соответствуют данным, указанным в литературных источниках и инструкциях по региональной биотехнологии культивирования. Построены карты пространственного распределения обеспеченных значений модельной продуктивности для придонных и садковых плантаций. Показано, что, несмотря на оптимальные концентрации пищевых ресурсов в кутовых частях бухты, при выборе объектов культивирования следует учитывать влияние резких перепадов солености на условия выращивания. В средней, более открытой, части бухты, несмотря на меньшее количество пищевых ресурсов, гидрологические условия выращивания более стабильны. Представленные картосхемы обеспеченных значений модельной продуктивности гидробионтов могут быть использованы при планировании размещения садковых и придонных плантаций, выбора видов культивирования с учетом особенностей производства первичной продукции, гидродинамического режима бухты, условий по абиотическим факторам и обеспеченности первичной продукцией, оптимальных с точки зрения допустимых для каждого отдельного производителя рисков.
Источник финансирования. Работа выполнена при частичной финансовой поддержке грантов РФФИ (проект № 20-05-00381-а) и РНФ (проект № 21-74-30004).
Список литературы
Администрация Приморского края. Доклад об экологической ситуации в Приморском крае в 2018 году. Владивосток, 2019, 252 с.
Афейчук Л.С., Мокрецова Н.Д. Биологические основы культивирования тихоокеанской мидии (Mytilus trossulus) в открытых районах залива Петра Великого // Изв. ТИНРО. 2000. Т. 127. С. 642–656.
Барабанщиков Ю.А., Тищенко П.Я., Семкин П.Ю. и др. Сезонные гидролого-гидрохимические исследования бухты Воевода (Амурский залив, Японское море) // Изв. ТИНРО. 2015. С. 180:161–178. https://doi.org/10.26428/1606-9919-2015-180-161-178
Брегман Ю.Э. Биоэнергетика трофической цепи “моллюск–фильтратор–голотурия–детритофаг” в условиях биокультуры. // Известия Тихоокеанского научно-исследовательского института рыбного хозяйства и океанографии. Владивосток, 1994. Т. 113. С. 5–12.
Вышкварцев Д.И. Особенности продукционных процессов в мелководных бухтах залива Посьета (Японское море) / Автореф. дисс. … канд. биол. наук. Владивосток. 1979. 22 стр.
Гаврилова Г.С. Приемная емкость аквакультурной зоны залива Петра Великого (Японское море) / Автореф. дисс. … докт. биол. наук. Владивосток: ТИНРО-Центр. 2012. 37 с.
Гаврилова Г.С. Современное состояние и проблемы развития дальневосточной марикультуры // Природные ресурсы, их современное состояние, охрана, промысловое и техническое использование. Мат. VIII Всероссийской научно-практической конференции, посв. 75-летию рыбохозяйственного образования на Камчатке. 2017. С. 68–71.
Гаврилова Г.С. Продуктивность плантаций двустворчатых моллюсков в Приморье [Электронный ресурс]: [монография] / А.В. Кучерявенко, Тихоокеан. науч.-исслед. рыбохоз. центр (ТИНРО-центр), Г.С. Гаврилова. Владивосток: ТИНРО-Центр, 2011. 113 с. Режим доступа: https://rucont.ru/efd/278345 .
Гайван Е.А., Земцов С.П., Мазурова А.А. Марикультура Приморского края. Потенциал развития отрасли в рамках акватерриториальных систем региона. 2012. LAP LAMBERT Academic Publishing.
Деева Р.А. Каталог гармонических и негармонических постоянных приливов отечественных вод морей Дальнего Востока. Л.: Гидрометеоиздат, 1972. 249 с.
Инструкция по технологии культивирования тихоокеанской мидии, сост. А.В. Кучерявенко, А.П. Жук; Тихоокеанский научно-исследовательский рыбохозяйственный центр. Владивосток: ТИНРО-Центр, 2011. 27 с.
Иструкция по технологии культивирования тихоокеанской устрицы, сост. А.В. Кучерявенко, А.П. Жук; Тихоокеанский научно-исследовательский рыбохозяйственный центр. Владивосток: ТИНРО-Центр, 2011. 27 с.
Карпевич А.Ф. Потенциальные свойства гидробионтов и их реализация в аквакультуре / Биологические основы марикультуры. М.: ВНИРО. 1998. С. 78–100.
Катрасов С.В., Бугаец А.Н., Жариков В.В.и др. Определение районов размещения плантаций марикультры на основе результатов гидродинамического моделирования // Океанология, 2021. Принята в печать.
Кафанов А.И. Двустворчатые моллюски шельфов и континентального склона северной Пацифики: аннотированный указатель. Владивосток: ДВО РАН СССР, 1991. 199 с.
Лоция северо-западного берега Японского моря от реки Туманная до мыса Белкина. ГУНО. 1984. 316 с.
Лысенко В.Н. Продукция макробентоса сообщества Zostera marina в северо-западной части Японского моря / Автореф. дисс. … канд. биол. наук., Владивосток: ДВНЦ АН СССР. 1985. 26 с.
Ляшенко С.А. Особенности воспроизводства тихоокеанской мидии в бухте Воевода (остров Русский) // Изв. ТИНРО. 2005. Т. 140. С. 352–365.
МакДональд Дж.Х., Коуэн Р.К., Балакирев Е.С. и др. Видовая принадлежность “съедобной мидии”, обитающей в приазиатской части Тихого океана // Биология моря. 1990. № 1. С. 13–22.
Отраслевая программа “Развитие товарной аквакультуры (товарного рыбоводства) в Российской Федерации на 2015–2020 годы”, 2014 г.
Селин Н.И. Динамика поселений тихоокеанской мидии в южном Приморье // Биология моря. 1990. № 4. С. 68–69.
Скарлато О.А. Двустворчатые моллюски умеренных широт западной части Тихого океана // Определители по фауне СССР. Л., 1981. Вып. 126. 479 с
Соколенко, Д.А., Калинина М.В. Современное состояние и структура естественных поселений тихоокеанской устрицы в северной части Амурского залива (залив Петра Великого, Японское море) // Изв. ТИНРО. 2018. Т. 195. С. 48–60.
Стратегия развития аквакультуры в Российской Федерации на период до 2020 г. М.: Минсельхоз России, 2007. 19 с.
Стратегия развития рыбохозяйственного комплекса Российской Федерации на период до 2030 года. М.: ФГБНУ “Роинформагротех”, 2019. 68 с.
Супрунович А.В. Аквакультура беспозвоночных. Киев : Наук, думка, 1988. 156 с.
Супрунович А.В., Макаров Ю.Н. Культивируемые беспозвоночные. Пищевые беспозвоночные: мидии, устрицы, гребешки, раки, креветки. Киев: Наук. думка, 1990. 264 с.
Технологические аспекты / Технология культивирования приморского гребешка.: Портал ЕСИМО (Единая государственная система информации об обстановке в Мировом океане) [Электронный ресурс] // Версия 1.6.8 Разработано Центром океанографических данных ФГБУ ВНИИГМИ-МЦД 1999-2020. URL: http://portal.esimo.ferhri.ru/portal/ portal/poi/japan/pacificKISWindowJapan (дата обращения: 20.04.2020)
Яковлев Ю.М., Раков В.А., Долгов Л.В. Размножение и развитие тихоокеанской устрицы Crassostrea gigas Thunb. // В кн.: Организмы обрастания дальневосточных морей. Владивосток: ДВНЦ АН СССР, 1981, с. 79–93.
Blauw A.N., Los F.J., Bokhorst M., Erftemeijer P.L.A. GEM: a generic ecological model for estuaries and coastal waters // Hydrobiologia. 2009. V. 618 P. 175–198.
Bugaets A.N., Gonchukov L.V., Sokolov O.V.et al. Information system to support regional hydrological monitoring and forecasting // Water Resources. 2018. T. 45. № S1. C. S59–S66.
Carr H., Axelsson L. Photosynthetic Utilization of Bicarbonate in Zostera marina Is Reduced by Inhibitors of Mitochondrial ATPase and Electron Transport // Plant Physiology. 2008. V. 147. No. 2. P. 879–885. https://doi.org/10.1104/pp.107.115584
D-WAQ TRM. 2013. D-Water Quality Technical Reference Manual. Deltares, 4.00 ed.
Ferreira J. G., Hawkin, A. J. S., Monteiro P. et al. Integrated assessment of ecosystem-scale carrying capacity in shellfish growing areas // Aquaculture. 2008. V. 275. No. 1–4. P. 138–151. https://doi.org/10.1016/j.aquaculture.2007.12.018
Ferreira J.G., Hawkins A.J.S., Bricker S.B. Management of productivity, environmental effects and profitability of shellfish aquaculture – the Farm Aquaculture Resource Management (FARM) model // Aquaculture. 2007. V. 264. P. 160–174.
Gangnery A., Bacher C., Buestel D. Assessing the production and the impact of cultivated oysters in the Thau lagoon (Méditerranée, France) with a population dynamics model // Can. J. Fish Aquat. Sci. 2001. V. 58. P. 1–9.
Los F.J, Wijsman J.W.M. Application of a validated primary production model (BLOOM) as a screening tool for marine, coastal and transitional waters // J. Marine Syst. 2007. V. 64. P. 201–215.
Los F. Eco-hydrodynamic modelling of primary production in coastal waters and lakes using BLOOM. Amsterdam: Ios Press. 2009. 288 p.
McKindsey C.W., Thetmeyer H., Landry T., Silvert W. Review of recent carrying capacity models for bivalve culture and recommendations for research and management // Aquaculture. 2006. V. 261 № 2. P. 451–462. https://doi.org/10.1016/j.aquaculture.2006.06.044
Newell C.R., Richardson J.R. Shellfish carrying capacity and site optimization // Aquac. Irel. 2004. V. 115 P. 15–19.
Nunes J.P., Ferreira J.G., Gazeau F.et al. A model for sustainable management of shellfish polyculture in coastal bays // Aquaculture. 2003. V. 219. № 1–4. P. 257–277.
Silva C., Ferreira J.G., Bricker S.B. et al. Site selection for shellfish aquaculture by means of GIS and farm-scale models, with an emphasis on data-poor environments // Aquaculture. 2011. V. 318 P. 444–457.
Väinölä R., Strelkov P. Mytilus trossulus in northern Europe // Marine Biology. 2011. V. 158. No. 4. P. 817–833.
Дополнительные материалы отсутствуют.