

Океанология, 2022, T. 62, № 1, стр. 112-116
Склерактиниевые кораллы как индикатор вертикальной мощности циркумантарктического течения
Н. Б. Келлер 1, *, Н. С. Оськина 1, **, Д. М. Ольшанецкий 2, ***
1 Институт океанологии им. П.П. Ширшова РАН
Москва, Россия
2 Геологический Институт РАН
Москва, Россия
* E-mail: keller@ocean.ru
** E-mail: nsoskina@mail.ru
*** E-mail: mitia@list.ru
Поступила в редакцию 04.02.2020
После доработки 01.04.2021
Принята к публикации 19.04.2021
- EDN: VVNRCG
- DOI: 10.31857/S0030157422010063
Аннотация
В статье приводятся аргументы, указывающие на связь батиметрического распространения эндемичных одиночных склерактиниевых кораллов, обитающих на шельфе и континентальном склоне Антарктиды, с вертикальной мощностью Циркумантарктического течения. Исследование проведено на материалах экспедиций Института океанологии и литературных данных. Полученные результаты показывают, что эндемичные виды, появившиеся в конце неогена–плейстоцене, не смогли проникнуть в другие океаны из-за уже существовавшего в то время мощного, как по горизонтали, так и по вертикали, циркумполярного течения.
ВВЕДЕНИЕ
Антарктида и омывающий ее Южный океан вызывают интерес геологов и океанографов как своей геологической историей, так и связанной с ней историей формирования циркумантарктического течения. Это течение, замкнутым кольцом охватывающее в настоящее время всю Антарктиду, формировалось параллельно с прогрессирующим оледенением Антарктического материка. По геологическим данным это оледенение началось в палеогене примерно 45 млн лет назад и закончилось в миоцене, когда мощное циркумантарктическое кольцо в Южном океане полностью замкнулось вокруг Антарктиды и приняло современный вид [1, 8, 9].
В настоящее время это сплошное “кольцо западного дрейфа” вокруг Антарктиды располагается между 40° и 50° ю.ш. Скорость водного потока определяется как 0.4–1 км/час, его ширина до 2500 км, общая длина протягивается до 30 000 км, а конкретные значения его вертикальной мощности в литературе не приводятся, лишь упоминается, что она может составлять несколько километров, и в некоторых районах течение достигает дна [7].
В этой статье приводятся аргументы, указывающие на связь батиметрического распространения эндемиков – одиночных склерактиниевых кораллов, обитающих в Антарктиде – с вертикальной мощностью циркумантарктического кольца.
Гидрологические данные применимы только к современному состоянию Циркумполярного течения и не позволяют судить о его горизонтальной и вертикальной мощности в прошлом – в неогене и плейстоцене. Существуют только косвенные данные. К таким косвенным данным относится и наше исследование.
Напоминаем, что склерактиниевые кораллы – группа коралловых полипов, основной особенностью которых является способность строить кальцитовый или арагонитовый скелет. Одиночные склерактинии, являющиеся предметом наших исследований, живущие (в отличие от образующих рифы) в большом диапазоне глубин (от первых десятков до более 6 км) и температур (от 20 до –1.1°С), широко распространены в океане. Они не создают на дне сплошного покрова, но, тем не менее, в результате многочисленных экспедиций, в том числе и российских, были обнаружены тысячи экземпляров этих организмов. В нашем распоряжении имеется уникальная коллекция глубоководных кораллов, собранная экспедициями Института океанологии им. П.П. Ширшова с разных глубин и широт. Изучение этой коллекции привело нас к выводу, что небольшая компактная группа этих кораллов удивительным образом проливает свет на многие аспекты жизни в океане. Приведем лишь несколько примеров. Оценка морфологических особенностей скелета одиночных склерактиний позволяет судить о количестве зоопланктона в окружающих водах. Изучение кораллового населения гайотов дает возможность оценить сравнительную скорость опускания этих одиночных подводных гор в разных областях Тихого и Атлантического океанов. Температурные диапазоны обитания видов приводят к возможности определения геологического возраста некоторых из них, и позволяют подойти к вопросу о происхождении кораллов [2]. Однако предположение о связи батиметрического распространения кораллов – эндемиков Антарктиды с мощностью циркумантарктического кольца течений по вертикали представлено здесь впервые.
МАТЕРИАЛ
Исследование проведено на материале Института океанологии (табл. 1) собранного в Южном полушарии в двух рейсах дизель-электрохода “Обь” и в рейсе НИС “Дмитрий Менделеев”. Многочисленные экземпляры склерактиниевых глубоководных кораллов были обнаружены в индийском секторе Южного океана на глубинах 270–2000 м (табл. 1). Среди них присутствовали 4 эндемичных вида: Gardineria antarctica Gardiner, 1929 (45 экземпляра), Caryophyllia antarctica Marenzeller, 1904 (33 экземпляра), Javania antarctica (Gravier, 1914) (14 экземпляров) и Flabellum impensum Squires, 1962 (3 экземпляра) [3, 6].
Таблица 1.
Распространение склерактиниевых одиночных кораллов, собранных и обработанных авторами в ходе рейсов НИС “Обь” и “Дмитрий Менделеев” института Океанологии им П.П. Ширшова
| Номер рейса | Станция | Глубина (м) | Координаты | Вид | |
|---|---|---|---|---|---|
| ю.ш. | з.д. | ||||
| “Обь”, 2 рейс, 1957 г. | 264 | 1400 | 35°44′ | 22°33′ | F. apertum F. fragilis |
| “Обь”, 3 рейс, 1958 г. | 328 | 1100 | 20°00′ | 88°09′ | Fl. japonicum bythios |
| 330 | 569 | 65°53′ | 114°07′ | J. antarctica | |
| 4089 | 5063–5470 | 60°46′ | 41°03′ | F. marenzelleri | |
| 4090 | 6145–5550 | 60°52′ | 40°58′ | F. marenzelleri | |
| 4096 | 285–260 | 60°50′ | 55°40′ | Fl. impensum | |
| 4109 | 5225 | 38°39′ | 48°10′ | F. marenzelleri | |
| 929 | 720 | 52°08′ | 57°16′ | C. antarctica F. curvatum B. malouinensis |
|
| 930 | 401 | 51°56′ | 57°35′ | M. oculata D. dianthus A. recidivus |
|
| 460 | 370 | 61°15′ | 57°48′ | F. thouarsii G. antarctica |
|
| 471 | 137 | 53°55′ | 64°10′ | F. thouarsii | |
| 336 | 700 | 69°34′ | 161°55′ | C. antarctica G. antarctica |
|
| 342 | 110 | 37°40′ | 150°00′ | B. bairdiana | |
| 355 | 1200 | 49°10′ | 166°23′ | F. apertum | |
| 377 | 900 | 67°21′ | 179°53′ | G. antarctica | |
| “Дмитрий Менделеев”, 43 рейс, 1989 г. | 4104 | 5120 | 62°36′ | 15°28′ | F. marenzelleri |
Однако главным образом мы использовали данные фундаментальной сводки по распространению глубоководных склерактиний в Антарктическом районе, принадлежащей С. Кернсу [10], который подробно описал и изобразил все ныне известные виды склерактиний Антарктики по собственным и всем доступным ему музейным материалам. Особенную ценность работе Кернса придают карты распространения видов, в том числе и видов-эндемиков.
РЕЗУЛЬТАТЫ
На шельфе Антарктиды поселения одиночных склерактиний разрежены и малочисленны, здесь обитает 17 видов. Среди них 8 эндемиков и 2 космополита. Все эндемики – одиночные кораллы. Из них 3 известны по единичным находкам, а 5 имеют циркумполярное распространение и встречаются часто (они приведены в табл. 2).
Таблица 2.
Глубины распространения часто встречающихся эндемиков
| Вид | Глубина в м |
|---|---|
| Caryophyllia antarctica Marenzeller, 1904 | 87–1435 |
| Gardineria antarctica Gardiner, 1929 | 87–728 |
| Flabellum flexuosum Cairns, 1882 | 101–659 |
| Javania аntarctica Gravier, 1914 | 53–1280 |
| Flabellum impensum Squires, 1962 | 46–2260 |
В этой работе мы ограничились изучением особенностей распределения указанных видов-эндемиков и видов-космополитов.
Из табл. 2 видно, что 4 широко распространенных в Антарктиде вида-эндемика имеют узкий батиметрический интервал, кроме последнего, который встречается и за пределами антарктической зоны. Этот вид, Flabellum impensum, имеет в основном циркумполярное приконтинентальное распространение, однако известна находка его представителей к северу от антарктической конвергенции (у островов Антиподы, на максимальных для вида глубинах). Кернс утверждает, что эта находка согласуется с возможным путем миграции фауны из моря Росса через о-ва Баллени – к островам Маккуори [10].
Согласно данным Кернса [10] и приведенной им карте вид-эндемик Сaryophyllia antarctica имеет циркумантарктическое распространение, но несколько больше находок отмечено в западной части региона.
Flabellum impensum также имеет циркумантарктическое распространение, но с резким преобладанием в западной части Приантарктического района. Что касается вида Flabellum flexuosum, то он тоже имеет циркумантарктическое распространение, но больше экземпляров приурочено к западной части, а в восточной его находки единичны. Gardineria аntarctica распространена достаточно равномерно по всему приантарктическому шельфу, тогда как Javania аntarctica встречена только в его западной части [10].
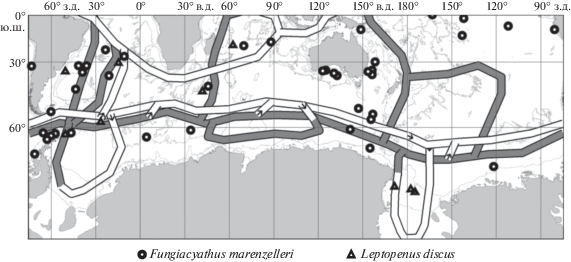
Лишь два вида-космополита, представленных одиночными формами, свободно пересекают антарктическую конвергенцию и встречаются как на континентальном склоне, так и в абиссали. Это Fungiacyathus marenzelleri [11] (300–6380 м) и Leptopenus discus Moseley, 1881 (2000–4870 м) (рис. 1). Кроме этих видов в абиссали Антарктики нет одиночных склерактиний.
Рис. 1.
Карта распространения одиночных склерактиниевых кораллов космополитов Fungiacyathus marenzelleri и Leptopenus discus и глобальный океанический конвейер [7].
 – Fungiacyathus marenzelleri
▲ – Leptopenus discus.
– Fungiacyathus marenzelleri
▲ – Leptopenus discus.
Постараемся ответить на вопрос: отчего в приантарктических водах кораллы представлены в основном эндемичными видами.
ОБСУЖДЕНИЕ
Склерактиниевые кораллы размножаются личинками – планулами, свободно плавающими в толще воды. Большинство личинок оседает в течение 1–2 дней после вымета, но некоторая их часть остается в толще воды в течение нескольких недель и даже месяцев и разносится на значительные расстояния поверхностными и глубинными течениями. Если течения мощные, они могут служить препятствием для проникновения личинок из одной географической области в другую [2].
Теперь поговорим о возрасте склерактиний – эндемиков Антарктиды. Не строящие рифов одиночные кораллы, обитающие в широком диапазоне глубин, от литорали до абиссали, появляются лишь в средней юре. К позднему мелу они уже прочно закрепились в части литорали, на шельфе и в батиальных областях, и к концу мела уже приобрели современный облик. Наиболее мощное их развитие падает на конец третичного времени, и в настоящее время в Мировом океане существует более 1000 их видов. Возраст появления большинства наиболее распространенных в океане родов склерактиний варьирует от средней юры до плейстоцена, но большая их часть возникла в палеогене [11]. Однако возраст появления большей части видов, населяющих современный океан, остается неясным. Для оценки возраста ряда современных видов мы попытались применить палеоклиматический подход, основываясь на ранее установленных нами температурных диапазонах существования различных их форм [2, 5]. Согласно этому подходу эндемики Антарктиды отнесены нами к группе умеренно холодноводных и холодноводных видов. Судя по кривой изменения палеотемператур придонной воды [5], выделенная нами группа умеренно-холодноводных видов, обитающая в диапазоне 0–10°C, смогла сформироваться в середине – конце эоцена (40–32 млн лет назад), а группа холодноводных видов, существующая при температурах ниже 5°С, могла возникнуть в начале олигоцена (32 млн лет назад) и существовать до позднего олигоцена (до 25 млн лет назад). Но затем наступило потепление до 8°С, и холодноводная группа, скорее всего, вымерла. Вторично эта группа могла возникнуть середине– конце миоцена, когда придонная температура начала снижаться до 5–6 °С, но, вероятно, еще позже. Можно предположить, что эндемичные виды молодого возраста–конец неогена или плейстоцен.
В биологии есть понятие – центр видообразования. В плейстоцене одним из таких центров, по-видимому, была Западная Антарктика, где на шельфе и верхней части континентального склона в фауне глубоководных склерактиниевых кораллов возникли новые виды. Появление этих видов было связано с адаптацией к чрезвычайно низкой температуре морской воды, сформировавшейся в плейстоцене [5]. Дальнейшее их распространение определялось гидрологическими условиями. Поскольку личинки кораллов свободно плавают (приблизительно на тех же глубинах, где живут сами кораллы), они стали переноситься Циркумантарктическим течением с запада на восток. Так как скорость течения составляет 0.4–0.9 км/час, то за сутки это такой перенос составляет 10–20 км, за месяц несколько сот км, а за год несколько тысяч км. Т.о. в течение плейстоцена кораллы смогли заселить весь шельф и верхнюю часть континентального склона Антарктиды, и их ареал в четырех случаях из пяти стал циркумантарктическим. А вот за пределы Южного океана виды не смогли проникнуть, так как им мешала не только скорость, но и большая вертикальная мощность течения и его горизонтальная протяженность, поскольку личинки не могут в достаточной мере опускаться на глубину и “подныривать” под течение. В результате появившиеся в плейстоцене виды остались видами-эндемиками.
Исходя из значений глубин вертикального распространения полипов эндемиков, можно предположить, что мощность препятствующего их расселению кольцевого водного потока по вертикали составляет не менее 1.5 км, а, может, и более (эту глубину в одном месте преодолевает эндемичный вид F. impensum).
Основная часть специфических для Антарктики видов склерактиниевых кораллов не пересекает Антарктическую конвергенцию, которая является границей для распространения многих типов фауны.
Лишь два вида-космополита, представленных одиночными формами, обитают как на континентальном склоне, так и в абиссали Антарктики. Это Fungiacyathus marenzelleri (Vaughan, 1906) (глубины 2000–6380 м) и Leptopenus discus Moseley, 1881 (глубины 2000–4870 м). Они принадлежат древним родам Fungiacyathus и Leptopenus, возникшим в глубинных слоях океана, и, нашему заключению [2, 4], являвшимися родоначальниками склерактиний. Эти два вида широко распространены на больших океанских глубинах (везде, кроме Арктики), легко минуют Антарктическую конвергенцию, и один из них даже поднимается по антарктическому шельфу до глубины 300 м (это единственная находка вида Fungiacyathus marenzelleri на такой глубине в Мировом океане). Исходя из нижней границы распространения вида-космополита Leptopenus discus, нижняя граница распространения которого 4870 м, вертикальная мощность течения не могла превышать 4–4.5 км: тогда мощное циркумантарктическое кольцо не может препятствовать его распространению. Следовательно, по вертикали эти течения явно не достигают глубин обитания, обычных для этих космополитных видов, что и согласуется со значениями величин батиметрического распространения шельфовых видов эндемиков Антарктики.
ВЫВОДЫ
1. Исходя из глубины обитания видов-эндемиков одиночных кораллов (шельф и верхняя часть континентального склона) современная вертикальная мощность (1.5–2 км) циркумантарктического кольца течений служит препятствием для их распространения в другие океаны.
2. Поскольку эти виды по нашим данным появились в конце неогена–плейстоцене, то, по всей вероятности, вертикальная мощность циркумантарктического течения в этом геологическом интервале времени уже достигало этих значений.
3. Виды-космополиты [2], обитающие на больших глубинах, свободно проникали в приантарктические воды, что указывает на то, что вертикальная мощность течения “западного дрейфа” не могла превышать 4–4.5 км.
Исследование выполнено в рамках государственного задания тема № 0128-2021-0008 и государственного задания ГИН РАН № 0114-2021-0003 и при поддержке проекта РФФИ № 19-05-00361.
Список литературы
Басов И.А. Океанская и климатическая эволюция в миоцене // Природа. 1999. № 5. С. 18–27.
Келлер Н.Б. Глубоководные склерактиниевые кораллы. М.: Крассанд. 2012. 384 с.
Келлер Н.Б. К вопросу о распространении мадрепоровых кораллов в Антарктике и Субантарктике // Труды ИО АН. 1990. Т. 126. С. 74–79.
Келлер Н.Б., Оськина Н.С., Савилова Т.А. Существует ли связь абиссальных склерактиниевых кораллов с океанским конвейером? // Океанология. 2017. Т. 57. № 4. С. 611–617.
Келлер Н.Б., Оськина Н.С., Николаев С.Д. Новый подход к определению возраста глубоководных видов склерактиний на основании температурных диапазонов их обитания // Докл. РАН. 2009. Т. 425. № 2. С. 218–222.
Келлер Н.Б., Пастернак Ф.А. Фауна кораллов Антарктического шельфа и континентального склона и вопрос о ее роли в процессе формирования современной глубоководной донной фауны океана // Океанология. 1996. Т. 36. № 4. С. 583–587.
Кошляков М.Н., Тараканов Р.Ю. Перенос воды через субантарктический фронт и глобальный океанский конвейер // Океанология. 2011. Т. 51. № 5. С. 773–787.
Baatsen M., van Hinsbergen D.J.J., von der Heydt A.S. et al. Reconstructing geographical boundary conditions for palaeoclimate modelling during the Cenozoic // Clim. Past. 2016. V. 12. Is. 8. P. 1635–1644.
Bice K.L., Scotese C.R., Seidov D., Barron E.J. Quantifying the role of geographic change in Cenozoic ocean heat transport using uncoupled atmosphere and ocean models // Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000. V. 161 P. 295–310. https://doi.org/10.1016/S0031-0182(00)00072-9
Cairns S.D. Antarctic and Subantarctic Scleractinia // Antarctic Research Series. 1982. V. 34. No. 1. P. 1–74.
Vaughan T.W., Wells J.W. Revision of the suborders, families and genera of the Scleractinia // Geol. Soc. Amer. Spec. Pap. 1943. V. 44. 363 p.
Дополнительные материалы отсутствуют.