

Океанология, 2022, T. 62, № 2, стр. 220-234
Количественные и продукционные характеристики фитопланктона северной части моря Уэдделла в летний период
Н. Д. Романова 1, *, С. А. Мошаров 1, 2, О. В. Воробьева 3, 4, Е. В. Бардюкова 1, В. А. Артемьев 1
1 Институт океанологии им. П.П. Ширшова РАН
Москва, Россия
2 Московский государственный технический университет им. Н.Э. Баумана
Москва, Россия
3 Всероссийский научно-исследовательский институт рыбного хозяйства и океанографии
Москва, Россия
4 Московский государственный университет имени М.В. Ломоносова
Москва, Россия
* E-mail: NdRomanova@gmail.com
Поступила в редакцию 13.05.2021
После доработки 10.10.2021
Принята к публикации 16.12.2021
- EDN: QDXPTA
- DOI: 10.31857/S0030157422020149
Аннотация
В работе представлены результаты измерений пространственного распределения основных параметров первичной продуктивности в северной части моря Уэдделла антарктическим летом 2020 г. Интегральное содержание хлорофилла “а” в фотическом слое варьировало от 23.1 до 85.8 мг/м2. Максимальные величины концентрации хлорофилла обычно были приурочены к верхнему 15-метровому слою, в большинстве случаев наблюдался и подповерхностный хлорофильный максимум. Величины интегральной суточной продукции фитопланктона варьировали от 26.7 до 654.6 мг С/м2 в сутки (в среднем 257.5 ± 66.6 мг С/м2 в сутки), снижаясь вдоль северной границы моря Уэдделла в восточном направлении. Наименее продуктивными были области материкового склона Антарктического полуострова. Максимальные величины первичной продукции наблюдались в узком диапазоне температур (–0.2–0.7 °С). Также наблюдалось уменьшение значения интегральной первичной продукции в поверхностных водах при снижении солености. В районах высокой и умеренной продуктивности не наблюдалось более интенсивного развития фитопланктона при уменьшении глубины верхнего перемешанного слоя, описанного для других районов Южного океана.
ВВЕДЕНИЕ
Море Уэдделла расположено в западной части Уэдделльского круговорота (Weddell Gyre), который является одной из главных океанографических особенностей Южного океана. Оно расположено к югу от Антарктического циркумполярного течения, имеющего высокий потенциал первичной продукции [31]. Одной из принципиальных особенностей моря Уэдделла является постоянный обильный ледяной покров и его крайняя сезонная изменчивость: к апрелю следующего года площадь ледового покрытия может сокращаться до трети от своей максимальной величины в сентябре [30].
Высокие летние значения концентрации хлорофилла “а” (хл “а”) как правило наблюдаются в открытом океане (1.5–4 мкг/л), что обычно связано с прикромочной зоной льда [10]. Однако наибольшие описанные летние концентрации (более 10 мкг/л) были приурочены к полыньям моря Уэдделла на северо-западе района [13].
Для значительной части Южного океана, включая и Уэдделльский круговорот, отмечен феномен высокой концентрации биогенных элементов и низкого содержания хлорофилла [16, 23]. Считается, что относительно низкая первичная продукция при высоком содержании биогенных элементов обусловлена в основном ограничениями по доступности света и железа [12]. Причем влияние этих двух факторов на фотосинтетическую активность фитопланктона взаимосвязано на физиологическом уровне. Ограничение количества железа вызывает нарушение синтеза пигментов и неэффективное функционирование системы транспорта электронов, снижая выход фотосинтеза на единицу хлорофилла [11]. Эти нарушения фотосинтетических процессов сильно снижают способность фитопланктона адаптироваться к условиям низкой освещенности [8, 18].
Световые условия развития фитопланктона определяются не только поверхностной освещенностью и длиной светового дня, но и стратификацией водной толщи. В зависимости от глубины простирания верхнего перемешанного слоя и его положения в пределах границ фотического слоя, меняются и условия развития фитопланктона. В области маргинальной ледовой зоны, где основным фактором стратификации выступает талая вода, первичная продукция и концентрация хлорофилла могут быть значительно выше по сравнению с районами дрейфующего льда и открытого моря [20]. На большей части пелагиали Антарктики, мало подверженной влиянию источников талой воды, верхний перемешанный слой имеет относительно большую глубину, выходящую за пределы фотического слоя, и, следовательно, характеризуется низкой первичной продуктивностью и низкими темпами роста фитопланктона [24].
В данной работе рассмотрены пространственное распределения хлорофилла “а” и скорости первичной продукции в северной части моря Уэдделла антарктическим летом 2020 года, а также проанализированы основные факторы, определяющие развитие фитопланктона этого района.
МАТЕРИАЛЫ И МЕТОДЫ
Район исследования и отбор проб. Исследования количественных и продукционных характеристик фитопланктона проводили в ходе второго этапа 79-го рейса НИС “Академик Мстислав Келдыш” в северной части моря Уэдделла в летний период южного полушария c 16 января по 6 февраля 2020 г. Место проведения работ включало районы внешнего восточного шельфа Антарктического полуострова, материковый склон, глубоководную котловину бассейна Пауэлла и подводные поднятия Южного хребта Скотия, отделяющие бассейн на севере от желоба Гесперидес и моря Скотия. Продвижение в центральную часть бассейна Пауэлла было ограничено северной границей распространения кромки дрейфующих льдов (рис. 1). В западной части исследуемого района, над материковым склоном Антарктического полуострова, были также встречены дрейфующие льды.
Рис. 1.
Карта-схема распределения хлорофилла “а” и первичной продукции фитопланктона в фотическом слое в районе работ.
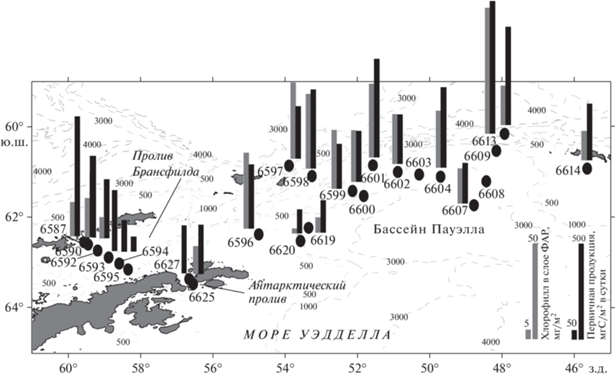
Пробы воды для определения концентрации хлорофилла “а” (хл “а”) и первичной продукции отбирали с помощью пластиковых батометров Нискина в составе комплекса SBE-32 Carousel Water Sampler на 16 станциях в море Уэдделла, а также на 9 станциях в проливе Брансфилда и на трех в Антарктическом проливе. Профили солености и температуры на станциях были получены с помощью CTD-зонда в составе комплекса. Горизонты отбора проб (5–7 горизонтов на станцию) выбирали по следующей схеме: поверхность, слой максимума флуоресценции, нижняя граница фотосинтеза (1% от поверхностной ФАР) и промежуточные горизонты.
Интенсивность надводной фотосинтетически активной радиации (ФАР) была измерена с использованием датчика падающей радиации в диапазоне ФАР LI-190SA (LI-COR). Результаты измерений интегрировались в блоке LI-1400 за пятиминутные интервалы (моль фотонов/м2) в течение всего дня. Профиль ФАР в водной толще измеряли с помощью датчика подводной освещенности LI-193 (LI-COR). Фотическая зона определялась как слой, ограниченный глубиной проникновения 1% от поверхностной ФАР.
Потенциальную плотность воды рассчитывали по данным температуры, солености и давления, полученным по результатам CTD-зондирований. Глубина верхнего перемешанного слоя (ВПС) оценивалась как глубина, на которой градиент плотности был равен или больше, чем 0.02 кг/м3 [7].
Хлорофилл и первичная продукция. Концентрации хл “а” и феофитина определяли флуориметрически [19]. Пробы воды объемом 0.5 л фильтровали через стекловолоконные фильтры Whatman GF/F при разряжении не более 0.3 атм. Далее фильтры помещали в 90% раствор ацетона и в течение суток выдерживали в темноте при температуре +4°С. Затем определяли флуоресценцию экстрактов с использованием флуориметра Smart (МГУ, Россия) до и после подкисления 1 Н HCl. Калибровка флуориметра была выполнена с использованием стандартного раствора хл “а” (Sigma).
Определение скорости первичного продуцирования проводили экспериментально радиоуглеродным методом [28]. Во флаконы c пробами воды объемом 50 мл добавляли раствор NaH14СO3. Экспонирование флаконов осуществляли по методу имитации световых и температурных условий в лабораторном инкубаторе с регулируемой светодиодной подсветкой [2, 4, 6]. Поддержание температуры инкубации, соответствующей температуре в точке отбора проб, осуществляли с помощью лабораторного охладителя HAILEA-100 и помпы для прокачки воды EHEIM. Уровень освещенности для каждого флакона, соответствующий освещенности на горизонте отбора пробы, задавался регулируемой величиной постоянного тока, протекающего через светодиод [4]. Пробы экспонировали в инкубаторе в течение трех часов. После экспозиции пробы фильтровали через мембранные фильтры “Владипор” (Россия) с размером пор 0.45 μ. Определения радиоактивности фильтров проводили на жидкостном сцинтилляционном радиометре Triathler (Hidex, Финляндия).
Ассимиляционное число (АЧ) рассчитывали как отношение величины первичной продукции на разных горизонтах к соответствующей концентрации хл “а”. Долю феофитина определяли как отношение концентрации феофитина к сумме концентраций хл “а” и феофитина. Этот параметр рассматривался как показатель состояния хлорофилла с точки зрения продукционного потенциала. Как показано в работе [6], доля феофитина менее 40% соответствует активному состоянию фитопланктона с высоким продукционным потенциалом.
Необходимые для расчета первичной продукции данные по содержанию в пробах минеральных форм углекислоты в воде были предоставлены А.А. Полухиным (ИО РАН). Интегральные значения первичной продукции и содержания хл “а” в столбе воды рассчитывались по методу трапеции. Статистическая обработка данных выполнялась с помощью программы PAST 3.14.
РЕЗУЛЬТАТЫ
Гидрологические и гидрохимические условия. Глубина ВПС в северной части бассейна Пауэлла варьировала от 8 до 58 м. На всех станциях она была значительно меньше, чем глубина фотического слоя, которая варьировала от 30 до 88 м, превышая на большинстве станций 60 м. Минимальные глубины фотического слоя (30–40 м) наблюдались в северо-восточной части района работ на станциях 6609 и 6613. В проливе Брансфилда глубина ВПС на большинстве станций варьировала от 25 до 43 м, и только на самой южной станции 6595 достигала 100 м. В Антарктическом проливе расчетная величина ВПС на разных станциях составляла 23–55 м. Средняя суточная ФАР в период измерений первичной продукции составляла 23.5 моль фотонов/м2 в сут при длине дня 17 ч.
Хлорофилл. Концентрация хл “а” в северной части бассейна Пауэлла на разных глубинах фотического слоя варьировала от 0.03 до 3.38 мг/м3, составляя в среднем 0.72 ± 0.18 мг/м3 (рис. 1). Максимальные значения были на ст. 6609 (3.38 мг/м3 на глубине 13 м) и в районе над глубоководным каналом хребта Филипп на ст. 6601 и 6613 (1.62 мг/м3 на глубине 45 м и 1.66 мг/м3 на поверхности, соответственно). Значения концентрации хл “а” в поверхностном слое изменялись в пределах от 0.25 до 2.74 мг/м3 (в среднем 0.88 ± ± 0.3 мг/м3).
На большинстве станций максимальные значения концентрации хл “а” наблюдались в верхнем 15-метровом слое. На некоторых станциях значения концентрации хл “а” сохранялись на одном уровне до глубин более 50 м, либо же наблюдались два пика концентрации (ст. 6601 и 6602): в поверхностном слое и на глубине около 40 м (рис. 2). Для большинства станций в море Уэдделла был характерен подповерхностный хлорофилльный максимум (ПХМ) [9, 14] на глубинах от 8 до 75 м, и в преобладающем числе этих случаев его глубина была ниже залегания ВПС (табл. 1).
Рис. 2.
Вертикальное распределение хлорофилла “а” на разрезах над поднятиями Южного хребта Скотия в (а) западном, (б) центральном и (в) восточном районе работ.
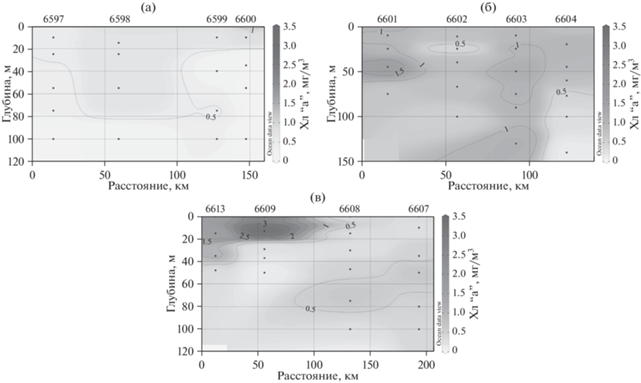
Таблица 1.
Распределение хлорофилла “а” в северной части моря Уэдделла и в проливе Брансфилда в январе–феврале 2020 г.
| Станции | ВПС | Хло | Хлвпс | Хлвод | Хлмакс/Хлср | ГХМ | Хлвпс/Хл∑ | Ф |
|---|---|---|---|---|---|---|---|---|
| Бассейн Пауэлла | ||||||||
| 6596 | 8 | 0.54 | 4.6 | 57.00 | 1.3 | 55 | 8% | 34% |
| 6597 | 22 | 0.54 | 11.8 | 41.30 | 1.3 | 10 | 29% | 32% |
| 6598 | 58 | 0.78 | 43.5 | 64.72 | 1.2 | 0 | 67% | 32% |
| 6599 | 32 | 0.78 | 19.2 | 49.86 | 1.4 | 0 | 38% | 31% |
| 6600 | 17 | 1.03 | 13.0 | 31.09 | 2.2 | 0 | 42% | 47% |
| 6601 | 33 | 1.07 | 35.6 | 85.85 | 1.5 | 45 | 41% | 29% |
| 6602 | 8 | 0.80 | 6.3 | 68.05 | 1.2 | 40 | 9% | 34% |
| 6603 | 8 | 0.90 | 7.2 | 135.74 | 1.2 | 50 | 5% | 40% |
| 6604 | 8 | 0.80 | 7.1 | 70.22 | 1.4 | 20 | 10% | 48% |
| 6607 | 12 | 0.34 | 3.9 | 46.58 | 1.5 | 50 | 8% | 52% |
| 6608 | 22 | 0.37 | 7.3 | 47.47 | 1.7 | 75 | 15% | 38% |
| 6609 | 15 | 2.74 | 45.9 | 72.52 | 2.5 | 13 | 63% | 50% |
| 6613 | 22 | 1.67 | 34.0 | 60.31 | 1.4 | 0 | 56% | 37% |
| 6614 | 22 | 1.15 | 24.5 | 37.57 | 1.5 | 8 | 65% | 38% |
| 6619 | 12 | 0.31 | 23.05 | 1.1 | 0 | 47% | ||
| 6620 | 12 | 0.25 | 3.6 | 26.00 | 1.4 | 15 | 14% | 35% |
| Пролив Брансфилда | ||||||||
| 6587 | 27 | 1.29 | 34.1 | 62.43 | 1.2 | 0 | 55% | 40% |
| 6588 | 39 | 1.11 | 43.7 | 57.22 | 1.4 | 37 | 76% | 37% |
| 6589 | 43 | 0.89 | 25.8 | 32.75 | 1.9 | 0 | 79% | 41% |
| 6590 | 25 | 0.92 | 19.7 | 42.77 | 1.4 | 0 | 46% | 37% |
| 6691 | 31 | 1.28 | 32.6 | 63.50 | 1.6 | 0 | 51% | 38% |
| 6592 | 34 | 0.75 | 22.8 | 41.37 | 1.4 | 0 | 55% | 42% |
| 6593 | 35 | 0.93 | 22.6 | 38.00 | 2.5 | 0 | 59% | 51% |
| 6594 | 25 | 0.41 | 9.4 | 24.55 | 1.3 | 0 | 38% | 31% |
| 6595 | 100 | 0.37 | 22.3 | 22.31 | 1.2 | 0 | 100% | 37% |
| Антарктический пролив | ||||||||
| 6622 | 12 | 0.71 | 9.8 | 23.5 | 1.6 | 0 | 42% | 45% |
| 6625 | 12 | 0.76 | 8.2 | 33.2 | 1.5 | 0 | 25% | 42% |
| 6627 | 32 | 0.61 | 15.2 | 25.1 | 1.5 | 0 | 61% | 42% |
Примечание. Хлo – концентрация хл “а” в поверхностном слое. мг/м3; Хлвпс – содержание хл “а” в слое фотосинтеза. мг/м2; Хлвод – общее содержание хл “а” в водном столбе, мг/м2; Хлмакс/Хлср – степень выраженности хлорофильного максимума, ГХМ – глубина хлорофильного максимума, м; Хлвпс/Хлвод – доля хл “а” в слое фотосинтеза относительно общего его содержания в водном столбе, %; Ф – средняя для слоя фотосинтеза доля феофитина в суммарном количестве хл “а” и феофитина, %.
Доля хл “а”, сосредоточенного в слое максимума, от общего содержания этого пигмента в водной толще существенно варьирует на разных станциях. Для количественной характеристики выраженности пика концентрации хлорофилла использовалось отношение величин концентрации хл “а” в слое максимума и средней концентраций для столба воды на конкретной станции (Хлмакс/Хлср). На рассматриваемой акватории это соотношение изменялось от 1.13 до 2.54. Наиболее ярко хлорофилльный максимум был выражен на ст. 6600 и 6609, где содержание хл “а” в слое максимума превышало среднее для столба значение более чем в два раза. Отсутствие выраженного хлорофилльного максимума, т.е равномерное распределение хл “а” по вертикали наблюдалось на ст. 6598, 6602, 6603 и 6619.
Вертикальное распределение значений концентрации хл “а” на всех рассматриваемых станциях показано на рис. 3а. Максимальные наблюдаемые значения на каждом из горизонтов постепенно снижаются с глубиной. В слое 0–50 м максимальные среди всех станций значения концентрации хл “а” достигали более чем 1 мг/м3. Минимальные, в свою очередь, не опускались ниже 0.25 мг/м3 в слое 0–30 м. На глубинах более 80 м концентрация хл “а” не превышала 0.5 мг/м3. При этом вариабельность значений на разных станциях на каждом горизонте сохранялась на высоком уровне во всей 100-м толще.
Рис. 3.
Вертикальное распределение параметров первичной продуктивности: (а) хлорофилл “а”, (б) первичная продукция в (⚪) море Уэдделла; (◆) проливе Брансфилда; (△) Антарктическом проливе.

Концентрация хл “а” в поверхностном слое воды согласуется с его интегральным содержанием в столбе воды 0–100 м (рис. 4). В северной части моря Уэдделла интегральное содержание хл “а” в столбе воды различалась на разных станциях более чем в четыре раза, варьируя от 23.1 до 85.8 мг/м2 (табл. 1) и составляя в среднем 42.41 ± ± 7.64 мг/м2. Максимальное содержание хл “а” (более 70 мг/м2) было определено в области поднятия южного хребта Скотия на станциях 6601–6604 и 6609. Минимальные величины в бассейне Пауэлла (23–31 мг/м2) наблюдались в западной части района в области материкового склона (станции 6619, 6620 и 6600). Доля хл “а” в фотическом слое (относительно всей водной толщи) варьировала от 38 до 95%, составляя в среднем 64 ± 8%. Более 74% от суммарного содержания хл “а” в столбе воды приходилось на фотический слой в северо-восточной части исследованного района (ст. 6609, 6613 и 6614), а также в области западного склона бассейна Пауэлла (ст. 6619 и 6600). На остальных станциях этот показатель был ниже 70%.
Рис. 4.
Соотношение между поверхностной концентрацией хлорофилла “а” и интегральными значениями для водной толщи в (⚫) море Уэдделла и (△) проливе Брансфилда.
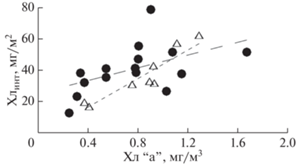
Как уже говорилось выше, глубина ВПС водной толщи была значительно меньше фотического слоя, разделяя последний на две части по вертикали. На большинстве станций, где глубина ВПС была менее 8–12 м, на него приходилось менее 15% от общего содержания хл “а” в водной толще (табл. 1). На станциях с более глубокой нижней границей ВПС (22–33 м) доля хлорофилла в нем составляла 38–65%.
Доля феофитина (неактивной формы хл “а”) в общей сумме хл “а” и феофитина в северной части моря Уэдделла на разных глубинах фотического слоя изменялась от 17 до 62%. Средняя величина этого показателя в фотическом слое варьировала между станциями от 30 до 49%. Значения доли феофитина, превышающие 40% были приурочены к склону бассейна Пауэлла (над глубинами более 1500 м), тогда как в области хребта Скотия, на шельфе Антарктического полуострова и Южно-Оркнейских островов этот параметр составлял 29–38%.
На поперечном разрезе в проливе Брансфилда [1] концентрация хл “а” в водной толще варьировала от 0.04 до 1.29 мг/м3 (в среднем 0.61 ± ± 0.15 мг/м3). Почти на всех станциях разреза максимум хлорофилла находился в поверхностном слое, за исключением ст. 6588 в северной части, ПХМ наблюдался на глубине 37 м. Наиболее выраженный максимум хлорофилла (Хлмакс/Хлср = = 2.5) наблюдался в южной части пролива на ст. 6593. На остальных станциях он был выражен значительно слабее (табл. 1). Среднее интегральное содержание хл “а” в фотическом слое изменялось от 16.7 мг/м2 в южной части пролива до 62.4 мг/м2 на крайней северной его станции. Средняя для фотического слоя доля феофитина в проливе Брансфилда варьировала от 31 до 50%. В Антарктическом проливе концентрация хл “а” варьировала от 0.16 до 0.76 мг/м3 (0.4 ± 0.08 мг/м3). Максимальные концентрации в столбе воды были приурочены к поверхности. Интегральная для фотического слоя величина концентрации хл “а” составляла на разных станциях от 23.5 до 33.3 мг/м2. Средняя для фотического слоя доля феофитина колебалась в пределах 43–47%.
Первичная продукция. Практически на всех станциях в районах исследования максимальная для профиля скорость первичной продукции наблюдалась в поверхностном слое воды, за исключением ст. 6596 и 6597, где максимальные величины продукции были приурочены к горизонту 10 м. График распределения значений скорости первичной продукции по вертикали на разных станциях, представленный на рис. 3б, показывает, что наиболее высокие значения (более 8 мг С/м3 в сут) наблюдались в верхнем 15-м слое воды. В слое глубже 30 м максимальные для горизонта значения не превышали 4 мг С/м3 в сут. Обращает на себя внимание тот факт, что очень низкие значения первичной продукции – менее 1 мг С/м3 в сут – встречались не только вблизи нижней границы фотической зоны, но и в приповерхностных слоях воды. Наблюдалась широкая вариабельность значений скорости первичной продукции на каждом рассматриваемом горизонте, которая снижалась с глубиной за счет уменьшения максимальных для горизонта значений, обусловленных соответствующим снижением освещенности.
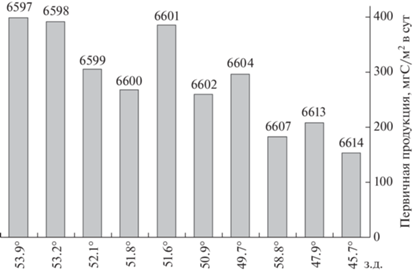
Величины первичной продукции в северной части моря Уэдделла в поверхностном слое воды варьировали от 2.7 до 63.5 мг С/м3 в сутки (табл. 2). Наиболее низкие значения первичной продукции (2.7–9.2 мг С/м3 в сут) наблюдались на глубоководных станциях бассейна Пауэлла. Максимальной величины в поверхностном слое (63.5 мг С/м3 в сут) первичная продукция достигала на станции 6609, также относительно высокие величины были отмечены на станциях 6600 и 6601 (15.9 и 18.1 мг С/м3 в сут соответственно). В остальных точках значения продукции в поверхностном слое варьировали от 11.3 до 14.4 мг С/м3 в сут. Величины интегральной суточной продукции фитопланктона в северной части моря Уэдделла варьировали от 26.7 до 654.6 мг С/м2 в сут, составляя в среднем 257.5 ± 66.6 мг С/м2 в сут. Наименее продуктивными были области материкового склона Антарктического полуострова на станциях 6619 и 6620 (81.3 и 26.7 мг С/м2 в сут соответственно). Максимальная величина первичной продукции (654.6 мг С/м2 в сут) была отмечена на станции 6609, также высокие значения (385.1–398 мг С/м2 в сут) наблюдались на шельфе восточной части пролива Брансфилда (ст. 6596) и на склоне глубоководного канала к востоку от острова Шишкова (ст. 6597, 5598 и 6601). Отдельно стоит выделить тенденцию снижения величин скорости первичной продукции вдоль южного хребта Скотия в восточном направлении с 397.8 мг С/м2 в сут на станции 6597 до 152.5 мгС/м2 в сутки на станции 6614 (r = 0.9; p < 0.01; рис. 5). Исключением стала станция 6609 на которой, как говорилось выше, наблюдалась аномально высокая величина первичной продукции.
Таблица 2.
Продукционные характеристики фитопланктона в северной части моря Уэдделла и в проливе Брансфилда в январе–феврале 2020 г.
| Станции | ППо | ИПП | АЧмакс | АЧср | ФС | ВПС | ППвпс |
|---|---|---|---|---|---|---|---|
| Бассейн Пауэлла | |||||||
| 6596 | 12.7 | 395.8 | 1.38 | 0.85 | 72 | 8 | 32% |
| 6597 | 12.3 | 397.8 | 1.38 | 0.92 | 78 | 22 | 62% |
| 6598 | 11.5 | 391.0 | 0.87 | 0.61 | 58 | 100% | |
| 6599 | 13.1 | 304.6 | 0.98 | 0.77 | 74 | 32 | 85% |
| 6600 | 15.9 | 267.3 | 0.91 | 0.59 | 17 | 65% | |
| 6601 | 18.1 | 385.1 | 0.99 | 0.56 | 58 | 33 | 86% |
| 6602 | 11.6 | 259.5 | 0.99 | 0.63 | 69 | 8 | 28% |
| 6604 | 9.2 | 296.1 | 0.67 | 0.43 | 62 | 8 | 23% |
| 6607 | 5.7 | 182.0 | 1.00 | 0.58 | 87 | 12 | 31% |
| 6609 | 63.5 | 654.6 | 1.36 | 0.61 | 30 | 15 | 91% |
| 6613 | 14.4 | 207.3 | 0.51 | 0.26 | 41 | 22 | 81% |
| 6614 | 11.3 | 152.5 | 0.58 | 0.30 | 54 | 22 | 95% |
| 6619 | 2.7 | 81.4 | 0.51 | 0.28 | 88 | 12 | 40% |
| 6620 | 2.7 | 26.7 | 0.64 | 0.23 | 12 | 67% | |
| Пролив Брансфилда | |||||||
| 6587 | 7.2 | 179.1 | 0.33 | 0.18 | 27 | 70% | |
| 6590 | 6.2 | 205.7 | 0.39 | 0.28 | 25 | 59% | |
| 6592 | 5.8 | 116.0 | 0.45 | 0.24 | 65 | 34 | 89% |
| Антарктический пролив | |||||||
| 6625 | 8.0 | 143.8 | 0.62 | 0.32 | 76 | 12 | 44% |
Примечание. ППо – первичная продукция в поверхностном слое, мг С/м3 в сутки; ИПП – интегральная первичная продукция, мг С/м2 в сутки, АЧмакс – максимальное ассимиляционное число для столба воды, мг С/мг Хл в час; АЧср – среднее для фотического слоя ассимиляционное число, мг С/мг Хл/ч; ФС– глубина фотического слоя, м; ВПС – глубина верхнего перемешанного слоя, м; ППвпс – доля первичной продукции в слое ВПС от общей интегральной первичной продукции, %.
Рис. 5.
Интегральная первичная продукция в области поднятий (хребет Филип и Южно-Оркнейское плато) северной границы бассейна Пауэлла.
В пределах ВПС формировалось от 23 до 100% первичной продукции (в среднем 65 ± 23%) (табл. 2). Наиболее высокий вклад первичной продукции в границах ВПС (более 80%) был отмечен на станциях, где нижняя граница ВПС была глубже 20 м, а отношение толщины ВПС к толщине фотического слоя было выше 0.4.
Максимальные для столба воды значения ассимиляционного числа (АЧмакс), характеризующего удельную фотосинтетическую активность фитопланктона, варьировали на разных станциях от 0.51 до 1.38 мг С/мг Хл в час (0.91 ± 0.16 мг С/мг Хл в час) и также в большинстве случаев были приурочены к поверхностному слою воды. Наибольшие величины ассимиляционного числа (1.38 мг С/мг Хл в час) были отмечены на станциях 6596 и 6597, расположенных в северо-западной части рассматриваемого района. Также высокий показатель ассимиляционного числа наблюдался на станции 6609 (1.36 мг С/мг Хл в час), которая характеризовалась максимальной продукцией фитопланктона. Наиболее низкие величины ассимиляционного числа были приурочены к северо-восточной части рассматриваемого района (ст. 6613 и 6619, 0.51 мг С/мг Хл в час), а также в области материкового склона Антарктического полуострова (ст. 6619 и 6620, 0.51 и 0.64 мг С/мг Хл в час соответственно).
На разрезе поперек пролива Брансфилда [1] интегральная первичная продукция варьировала от 116 до 205.7 мг С/м2 в сутки, при этом значения в южной его части были примерно в полтора раза ниже, чем в северной. По вертикали максимальные величины первичной продукции наблюдались в поверхностном слое воды. Сходная картина вертикального распределения первичной продукции наблюдалась и в Антарктическом проливе, где интегральная величина первичной продукции составляла 143.8 мг С/м2 в сутки. Значения АЧмакс в проливе Брансфилда (0.33–0.45 мгС/мг Хл в час) и Антарктическом проливе (0.62 мг С/мг Хл в час) были в среднем ниже наблюдавшихся в северной части бассейна Пауэлла и также были приурочены к верхнему десяти-метровому слою воды.
ОБСУЖДЕНИЕ
Полученные нами данные о пространственном распределении основных параметров первичной продуктивности в северной части моря Уэдделла и прилегающих районах, относящихся к Атлантическому сектору Южного океана, хорошо согласуются с имеющимися литературными данными. Полученные нами величины ИПП варьировали достаточно широко (27–398 мг С/м2 в сут; табл. 2) и были сопоставимы с предыдущими исследованиями в этом районе Антарктики. Ранее описанные величины первичной продукции в северной части моря Уэдделла в летний период составляли 104–440 мг С/м2 в сут [17, 20], достигая максимальных значений в маргинальной ледовой зоне. Сезонные изменения средних значений первичной продукции в море Уэдделла в районе распространения сезонного льда ярко выражены: весной 1.13 мг С/м2 в сут [20], летом 0.41 мг С/м2 в сут [17] и осенью 0.13 мг С/м2 в сут [27]. Концентрация хл “а” в зоне близости льдов в летний период составляла 28 мг/м2 [21].
Для всего района северной части моря Уэдделла стоит отметить снижение концентрации хл “а” в фотическом слое по мере продвижения на юг (r = –0.75; p < 0.01). При сопоставлении вертикального распределения двух главных параметров первичной продуктивности – концентрации хл “а” и значений скорости первичной продукции (рис. 3) очевидны существенные различия. Максимальные для каждого горизонта концентрации хл “а” достаточно медленно снижаются с глубиной в пределах верхнего 100-метрового слоя (примерно в 3 раза). Максимальные для каждого горизонта скорости первичной продукции экспоненциально снижаются с глубиной в пределах верхнего 50-метрового слоя (примерно в 18 раз), что отражает закономерное снижение освещенности с глубиной. Очевидно, что реализация продукционного потенциала фитопланктона наблюдалась только в верхних хорошо освещенных слоях водной толщи. Сходная ситуация отмечалась нами и в арктических полярных районах (Карское море [5, 22]).
Основываясь на распределении величин интегральной первичной продукции (ИПП) станции в северной части моря Уэдделла можно разделить на 4 группы, приуроченные к определенным районам рассматриваемой области. В анализ не были включены данные для ст. 6609 из-за аномально высокой скорости первичной продукции в поверхностном слое и, следовательно, большого значения ИПП. Станции с относительно высокими значениями ИПП (ст. 6596, 6597, 6598 и 6601; 385–397 мг С/м2 в сут) расположены на северо-западной границе моря Уэдделла (район 1). Предположительно, эти станции находились в районе контакта Антарктического циркумполярного течения и вод моря Уэдделла [3]. Умеренные значения ИПП (259–304 мг С/м2 в сут; ст. 6699, 6600, 6602 и 6604) были определены в области северного склона бассейна Пауэлла (район 2). В северо-восточной части рассматриваемой области (район 3) наблюдались еще более низкие значения ИПП (152–207 мг С/м2 в сут, ст. 6607, 6613, 6614). Минимальные величины ИПП для рассматриваемого района (27–82 мг С/м2 в сут; ст. 6619, 6620) были отмечены в области материкового склона на западной границе бассейна Пауэлла (район 4).
В антарктических водах ПХМ не обязательно совпадает с горизонтом максимальных скоростей первичной продукции. Однако его экологическое значение (вклад в ИПП, вертикальный транспорт углерода, область активного питания зоопланктона) наиболее выражено, когда их положение совпадает. Важную роль играют также процессы фотоакклимации или смены сообществ [9]. Наши исследования показали, что в северной части моря Уэдделла ПХМ совпадал с высокой скоростью первичной продукции только на одной станции (ст. 6597, 10 м). На остальных станциях максимальные значения первичной продукции, наблюдавшиеся в основном на поверхности, либо сопровождались максимумом хл “а” также на поверхности, либо более глубоким ПХМ. На поперечном разрезе в проливе Брансфилда на всех станциях максимумы первичной продукции и хл “а” совпадали, находясь в поверхностном слое воды.
В качестве первичной оценки активности хл “а” можно использовать показатель доли феофитина от общей суммы хлорофилла и феофитина. Ранее в работе [6] для арктических морей было показано, что доля феофитина менее 40% соответствует активному состоянию хлорофилла с высоким продукционным потенциалом. В ходе наших наблюдений потенциальная активность хлорофилла была умеренно высокой, в верхнем 30-метровом слое доля феофитина варьировала от 20 до 46%, что может быть характерно для ранне-летнего фитопланктона, завершившего активную стадию весеннего цветения. Минимальные средние для фотической зоны значения доли феофитина (31–32%) наблюдались в районе 1 с наибольшими значениями ИПП.
По данным гидрохимических исследований, проводившихся одновременно с нашими работами [26], содержание биогенных элементов значительно превышало лимитирующие уровни и не определяло уровень первичной продуктивности в районе исследований.
Показано, что в Антарктических водах при низких величинах первичной продукции на фоне высокого содержания биогенных элементов определяющими ее факторами являются доступность света и железа [12]. Мы проанализировали изменчивость интегральной первичной продукции в северной части моря Уэдделла в летний период в зависимости от глубины ВПС. Именно этот показатель, в случае нахождения ВПС в пределах фотического слоя, определяет световую адаптацию населяющего его фитопланктона.
Северо-западная часть моря Уэдделла в период исследований была покрыта льдом. В рассматриваемой области бассейна Пауэлла сплошного ледового покрова не наблюдалось, но присутствовали айсберги. Максимальное содержание талых вод наблюдалось в южных районах бассейна Пауэлла и в районе Южных Оркнейских о-вов (северо-восточная часть бассейна [3]). Снижение солености в поверхностных водах за счет талой воды сопровождалось уменьшением величин ИПП (r = 0.84; p < 0.01; рис. 6а). Именно в зоне влияния талых вод располагались станции с невысокими значениями ИПП (районы 3 и 4). Ранее для морей Скотия и Уэдделла в весенне-летний период были описаны минимальные величины ИПП в приледной зоне, тогда как максимальных значений этот показатель достигал на расстоянии около половины градуса широты к северу от границы ледового покрова [20].
Рис. 6.
Соотношение между интегральной первичной продукцией и гидрофизическими параметрами поверхностного слоя: (а) соленостью и (б) температурой.
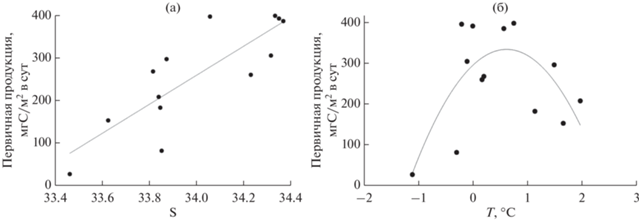
Зависимость величин ИПП от поверхностной температуры воды носила более сложный характер (рис. 6б). ИПП достигала максимальных величин при значениях температуры в диапазоне ‒0.2–0.7°С, и снижалась за пределами этого интервала температур. Считается, что сообщества антарктического фитопланктона представлены преимущественно облигатными психрофиллами, адаптированными к узкому диапазону низких температур и снижающими активность при более высоких температурах [14, 17].
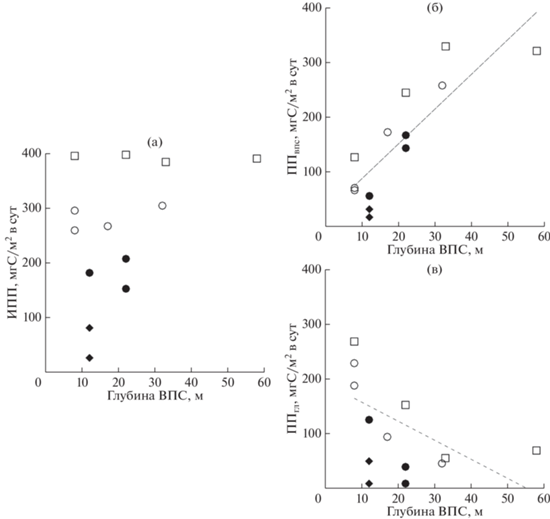
Более ранние исследования в Южном океане [15, 25, 29] показали, что одним из ключевых факторов развития “цветения” фитопланктона и увеличения продуктивности является ослабление световой лимитации фотосинтеза благодаря уменьшению глубины ВПС за счет талых вод. В период наших исследований, как видно из рис. 7, в трех из четырех выделенных на основании величины первичной продукции районов глубина ВПС варьирует в довольно широких пределах, тогда как продуктивность сохраняется примерно на одном уровне. Следовательно, несмотря на то, что фитопланктонные сообщества на станциях этих районов проводят разную по продолжительности часть дня в условиях низкой освещенности в результате перемещения в пределах ВПС, они формируют сходный уровень первичной продуктивности. При рассмотрении соотношения между глубиной ВПС и изменением величины ПП, интегрированной по глубине ВПС (ППвпс) наблюдается рост значений интегральной первичной продукции с увеличением глубины ВПС (рис. 7б). В нижней части фотической зоны (слой глубже ВПС) интегральная для этого слоя первичная продукция (ППгл) значительно увеличивалась с уменьшением глубины ВПС (рис. 7в). При минимальных значениях глубины ВПС (8–12 м) большая часть общей интегральной первичной продукции в фотическом слое (60–77%) формировалась в слое глубже ВПС (табл. 2). В районе 4 (с минимальными величинами ИПП) даже при небольшой глубине ВПС первичная продуктивность была низкой. Эти точки также характеризовались низкими значениями температуры воды (–0.3…–1.12°С) и невысокой по сравнению с соседними районами соленостью (33.46–33.85 psu).
Рис. 7.
Зависимость интегральной первичной продукции от глубины ВПС в фотическом слое (а), в ВПС (б) и в слое глубже ВПС (в) в выделенных районах северной части моря Уэдделла: (◻) – район 1; (⚪) – район 2; (⚫) – район 3; (◆) – район 4. Пунктирной линией обозначена линия тренда для станций районов с высокой и средней продуктивностью (районы 1–3).
На уровень ИПП влияет величина фотосинтетической активности и концентрация хл “а”. Интегральное содержание хл “а” не зависело от глубины ВПС. Фотосинтетическая активность фитопланктона, выражаемая через ассимиляционное число, и в верхней, и в нижней частях фотической зоны (в слое ВПС и глубже ВПС, соответственно) имела тенденцию к снижению с увеличением глубины ВПС (рис. 8). По-видимому, с ростом ВПС клетки фитопланктона периодически оказываются в условиях меньшей освещенности при перемешивании воды и средний уровень освещенности, к которому акклимируется фотосистема фитопланктона, соответственно снижается. Следует отметить более высокий уровень значений АЧ в обоих слоях для станций с высокой и умеренной ИПП (районы 1 и 2). С увеличением глубины ВПС фотосинтетическая активность в этом слое закономерно снижается в связи со снижением среднесуточной освещенности. Однако при этом за счет большей глубины вырастает и содержание хл “а” в слое, что приводит к увеличению продуктивности. Значения ППвпс положительно коррелировали с величиной интегрального содержания хлорофилла в этом слое (r = 0.83, p < 0.05).
Рис. 8.
Зависимость средних для слоя значений АЧ от глубины ВПС в фотическом слое (а), в ВПС (б) и в слое глубже ВПС (в) в выделенных районах северной части моря Уэдделла: (◻) – район 1; (⚪) – район 2; (⚫) – район 3; (◆) – район 4. Пунктирной линией обозначены линия тренда для станций районов с высокой продуктивностью (¦), умеренной и низкой продуктивностью (¦).
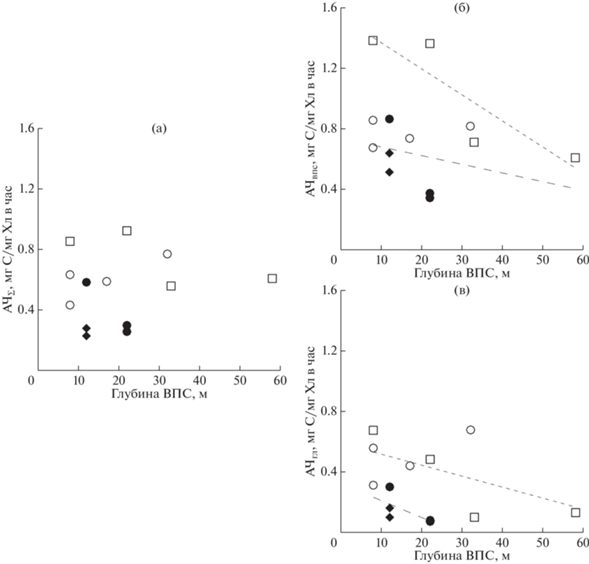
Проведенный анализ позволил оценить роль фактора доступности света, влияющий на продуктивность фитопланктона в изменении интегральной первичной продукции в двух слоях фотической зоны. Нижняя граница ВПС разделяла фотическую зону на две части, и при ее снижении происходило существенное перераспределение суммарного света, проникающего в водную толщу, между двумя слоями. Соотношение между глубиной ВПС и глубиной фотического слоя варьировало на разных станциях от 0.11 до 0.57. Небольшая глубина ВПС (до 12 м) обеспечивала проникновение большого количества света в нижний слой фотической зоны, что определяло благоприятные световые условия для первичной продукции и в этом слое. На нижней границе ВПС в этих случаях было установлено проникновение от 23 до 45% от освещенности на поверхности воды. При увеличении глубины ВПС и расширении светового диапазона, в котором находится фитопланктон в течение дня в результате вертикальной циркуляции в пределах слоя, снижается среднесуточный уровень освещенности, влияющий на фотосинтетическую активность фитопланктона. В результате адаптации фотосистемы фитопланктона к определенному уровню освещенности его фотосинтетическая активность (АЧ) закономерно снижается при увеличении глубины ВПС на всех станциях в обоих слоях фотической зоны (рис. 8).
Сравнивая количественные и продукционные характеристики фитопланктона с ближайшими районами, можно сказать, что величина интегральной для фотического слоя концентрации хл “а” в северной части пролива Брансфилда была близка к показателям, наблюдавшимся в области поднятий северной части моря Уэдделла: 44.3 ± 9.7 и 44.6 ± 8.1 мг/м2 соответственно. Среднее содержание хл “а” в фотическом слое в северной части пролива было близко с показателями для станций над материковым склоном Антарктического полуострова (16.7–19.6 и 12.7–23.1 мг/м2). В Антарктическом проливе величина интегральной концентрации хл “а” в фотическом слое также была относительно невысока, составляя в среднем 24.8 ± 1.4 мг/м2.
По сравнению с проливом Брансфилда и Антарктическим проливом можно говорить о более высоких значениях интегральной первичной продукции в области поднятий на северной границе моря Уэдделла. В среднем этот показатель в проливах был ниже почти в два раза. Однако в области шельфа Южно-Оркнейских островов (ст. 6614) и в центральной части бассейна Пауэлла (ст. 6607) значения интегральной первичной продукции были близки к величинам в проливах, а над материковым склоном Антарктического полуострова (ст. 6619 и 6620) даже ниже более чем в два раза. Несмотря на это, значения АЧ поверхностного слоя воды в проливе Брансфилда (0.39 ± ± 0.07 мкг С/мкг Хл час–1) были минимальными по сравнению с соседними районами: в области поднятий на северной границе моря Уэдделла этот показатель составлял в среднем 0.99 ± ± 0.14 мкг С/мкг Хл час–1, на шельфе Южно-Оркнейских островов и над материковым склоном Антарктического полуострова его среднее значение составляло 0.53 ± 0.07 мкг С/мкг Хл час–1, а в Антарктическом проливе – 0.62 мкг С/мкг Хл час–1.
ЗАКЛЮЧЕНИЕ
Таким образом, вертикальная структура распределения концентрации хлорофилла и активности фитопланктона в северной части моря Уэдделла была более сложной, нежели наблюдавшаяся в прилегающих к морю проливах. Несмотря на то, что на большинстве станций максимальные концентрации хл “а” наблюдались в верхнем 15-метровом слое, в преобладающем количестве случаев был также отмечен и подповерхностный пик концентрации пигмента. Минимальные средние для фотической зоны значения доли феофитина были приурочены к северо-восточной окраине моря, где была отмечена и высокая продукционная активность. В целом для рассматриваемого района потенциальная активность хлорофилла была умеренно высокой, что может быть характерно для ранне-летнего фитопланктона, завершившего активную стадию весеннего цветения.
В областях высокой и умеренной продуктивности наблюдалась разнонаправленная зависимость ПП от глубины ВПС в двух слоях фотической зоны: положительная в ВПС и отрицательная в слое глубже ВПС. Таким образом, в период наших исследований уменьшение глубины ВПС не провоцировало более активный рост фитопланктона верхнего слоя за счет стабильных световых условий, однако сохранение уровня ИПП обеспечивается увеличением величины первичной продукции в фотическом слое под ВПС. Распределение хл “а” также носило консервативный характер: его содержание в слое ВПС и под ним тем больше, чем больше толщина этого слоя. Развитие “цветения” в распресненных из-за таяния льда водах, описанное в других работах, не было отмечено в ходе наших исследований. Это подтверждается и трендом увеличения значений ИПП при росте солености поверхностного слоя воды. В свою очередь, влияние температуры на величину ИПП имело более сложный характер: значения интегральной продукции достигали максимальных величин при температуре в диапазоне –0.2–0.7°С, и снижались за его пределами. Это может косвенно свидетельствовать о доминировании психрофиллов в сообществе фитопланктона. Дальнейший анализ факторов, влияющих на активность фитопланктона, требует как анализа состава сообществ, так и более глубокого рассмотрения гидрологии исследуемого региона в период работ.
Работа выполнена в рамках Госзадания 0128-2019-0008.
Список литературы
Воробьева О.В., Романова Н.Д., Мошаров С.А. и др. Продукционные характеристики фитопланктона пролива Брансфилд в летний период // Труды ВНИРО. 2020. Т. 181. С. 178–186 https://doi.org/10.36038/2307-3497-2020-181-178-186
Гонтарев С.В., Мошаров С.А. Инкубатор и способ инкубации пробы воды: патент на изобретение № 2 547 685 РФ// Бюл. № 10. 2015. 10 с.
Морозов Е.Г., Спиридонов В.А., Молодцова Т.Н. и др. Исследования экосистемы атлантического сектора Антарктики (79-й рейс научно-исследовательского судна “Академик Мстислав Келдыш”) // Океанология. 2020. Т. 60. № 5. С. 823–825. https://doi.org/10.31857/S0030157420050172
Мошаров С.А., Гонтарев С.В., Корсак М.Н. Новый фитоинкубатор с регулированием температуры и освещенности для мониторинга экологических параметров водных экосистем // Безопасность в техносфере. 2015. Т. 4. № 4. С. 3–9. https://doi.org/10.12737/14428
Мошаров С.А., Демидов А.Б., Симакова У.В. Особенности процессов первичного продуцирования в Карском море в конце вегетационного периода // Океанология. 2016. Т. 56. № 1. С. 90–100. https://doi.org/10.7868/S003015741601010X
Мошаров С.А., Сергеева В.М. Оценка функционального состояния морского фитопланктона по флуоресцентным показателям и соотношению концентраций феофитина и хлорофилла а // Вопросы современной альгологии. 2018. № 1 (16). С. 10.
Alderkamp A.-C., Garcon V., de Baar H.J.W., Arrigo K.R. Short-term photoacclimation effects on photoinhibition of phytoplankton in the Drake Passage (Southern Ocean) // Deep Sea Research Part I: Oceanographic Research Papers. 2011. V. 58. P. 943–955. https://doi.org/10.1016/j.dsr.2011.07.001
Alderkamp A.-C., Kulk G., Buma A.G.J. et al. The effect of iron limitation on the photophysiology of Phaeocystis antarctica (Prymnesiophyceae) and Fragilariopsis cylindrus (Bacillariophyceae) under dynamic irradiance1: photophysiology under iron limitation // Journal of Phycology. 2012. V. 48. P. 45–59. https://doi.org/10.1111/j.1529-8817.2011.01098.x
Baldry K., Strutton P.G., Hill N.A., Boyd P.W. Subsurface Chlorophyll-a Maxima in the Southern Ocean // Frontiers in Marine Science. 2020. V. 7. P. 1–19. https://doi.org/10.3389/fmars.2020.00671
Bathmann U.V., Scharek R., Klaas C/ et al. Spring development of phytoplankton biomass and composition in major water masses of the Atlantic sector of the Southern Ocean // Deep Sea Research Part II: Topical Studies in Oceanography 1997. V. 44. P. 51–67. https://doi.org/10.1016/S0967-0645(96)00063-X
Behrenfeld M.J., Bale A.J., Kolber Z.S. et al. 1996. Confirmation of iron limitation of phytoplankton photosynthesis in the equatorial Pacific Ocean // Nature. 1997. V. 383. P. 508–511. https://doi.org/10.1038/383508a0
Boyd P.W., Strzepek R., Fu F., Hutchins D.A. Environmental control of open-ocean phytoplankton groups: Now and in the future // Limnol. Oceanogr. 2010. V. 55. P. 1353–1376. https://doi.org/10.4319/lo.2010.55.3.1353
Cape M.R., Vernet M., Kahru M., Spreen G. Polynya dynamics drive primary production in the Larsen A and B embayments following ice shelf collapse: primary production in the Larsen area // J. Geophys. Res. Oceans. 2014. V. 119. P. 572–594. https://doi.org/10.1002/2013JC009441
Cullen J.J. Subsurface chlorophyll maximum layers: Enduring Enigma or Mystery Solved? // Annu. Rev. Mar. Sci. 2015. V. 7. P. 207–239. https://doi.org/10.1146/annurev-marine-010213-135111
de Baar H.J.W. Synthesis of iron fertilization experiments: From the Iron Age in the Age of Enlightenment // J. Geophys. Res. 2005. V. 110. C09S16. P. 1–24 https://doi.org/10.1029/2004JC002601
de Baar H.J.W., de Jong J.T., Bakker D. C.et al. Importance of iron for plankton blooms and carbon dioxide drawdown in the Southern Ocean //Nature. 1995. V. 373. № 6513. P. 412–415. https://doi.org/10.1038/373412a0
El-Sayed S.Z., Taguchi S. Primary production and standing crop of phytoplankton along the ice-edge in the Weddell Sea // Deep Sea Research Part A. Oceanographic Research Papers. 1981. V. 28. P. 1017–1032. https://doi.org/10.1016/0198-0149(81)90015-7
Galbraith E.D., Gnanadesikan A., Dunne J.P., Hiscock M.R. Regional impacts of iron-light colimitation in a global biogeochemical model //Biogeosciences. 2010. V. 7. P. 1043–1064. https://doi.org/0.5194/bg-7-1043-2010
Knap A., Michaels A., Close A. et al. Protocols for the joint global ocean flux study (JGOFS) core measurements. JGOFS, Reprint of the IOC Manuals and Guides No. 29, UNESCO 1994. 1996. V. 19. 170 p.
Mathot S., Dandois J.-M., Lancelot C. Gross and net primary production in the Scotia-Weddell Sea sector of the Southern Ocean during spring 1988 // Weddell Sea Ecology. Springer Berlin Heidelberg. 1992. P. 321–332. https://doi.org/10.1007/978-3-642-77595-6_37
Morán X., Gasol J., Pedrós-Alió C., Estrada M. Dissolved and particulate primary production and bacterial production in offshore Antarctic waters during austral summer: coupled or uncoupled? // Mar. Ecol. Prog. Ser. 2001. V. 222. P. 25–39. https://doi.org/10.3354/meps222025
Mosharov S.A., Sergeeva V.M., Kremenetskiy V.V. et al. Assessment of phytoplankton photosynthetic efficiency based on measurement of fluorescence parameters and radiocarbon uptake in the Kara Sea // Estuarine, Coastal and Shelf Science. 2019. V. 218. P. 59–69. https://doi.org/10.1016/j.ecss.2018.12.004
Platt T. Sathyendranath S., Edwards A.M. et al. Nitrate supply and demand in the mixed layer of the ocean // Marine Ecology Progress Series. 2003. V. 254. P. 3–9. https://doi.org/10.3354/meps254003
Sakshaug E., Holm-Hansen O. Photoadaptation in Antarctic phytoplankton: variations in growth rate, chemical composition and P versus I curves // J Plankton Res. 1986. V. 8. P. 459–473. https://doi.org/10.1093/plankt/8.3.459
Sambrotto R., Mace B. Coupling of biological and physical regimes across the Antarctic Polar Front as reflected by nitrogen production and recycling // Deep Sea Research Part II: Topical Studies in Oceanography. 2000. V. 47. P. 3339–3367. https://doi.org/10.1016/S0967-0645(00)00071-0
Stepanova S., Polukhin A., Borisenko G. et al. Hydrochemical structure of waters in the northern Weddell Sea in austral summer 2020 // Antarctic Peninsula Region of the Southern Ocean: Oceanography and Ecology. Springer Nature Oceanography. in press
Smith W., Nelson D.M. Phytoplankton growth and new production in the Weddell Sea marginal ice zone in the austral spring and autumn // Limnol. Oceanogr. 1990. V. 35. P. 809–821. https://doi.org/10.4319/lo.1990.35.4.0809
Steemann Nielsen E. The Use of Radio-active Carbon (C14) for measuring organic production in the sea // ICES Journal of Marine Science. 1952. V. 18. P. 117–140. https://doi.org/10.1093/icesjms/18.2.117
Gieskes W.W.C., Bellerby R., Weissing F.J. et al. Light rather than iron controls photosynthate production and allocation in Southern Ocean phytoplankton populations during austral autumn // Journal of Plankton Research. 2004. V. 26. P. 885–900. https://doi.org/10.1093/plankt/fbh088
Vernet M., Geibert W., Hoppema M. et al. The Weddell Gyre, Southern Ocean: Present Knowledge and Future Challenges // Rev. Geophys. 2019. V. 57. P. 623–708. https://doi.org/10.1029/2018RG000604
Whitehouse M.J., Atkinson A., Korb R.E. et al. Substantial primary production in the land-remote region of the central and northern Scotia Sea // Deep Sea Research Part II: Topical Studies in Oceanography. 2012. 59–60, 47–56. https://doi.org/10.1016/j.dsr2.2011.05.010
Дополнительные материалы отсутствуют.