

Океанология, 2022, T. 62, № 5, стр. 726-742
Оценки первичной продукции в северной части Японского моря в различные сезоны по судовым и спутниковым данным
В. И. Звалинский 1, *, П. В. Лобанова 2, П. Я. Тищенко 1, В. Б. Лобанов 1, А. Д. Маховиков 2
1 Тихоокеанский океанологический институт им. В.И. Ильичева Дальневосточного отделения
Российской академии наук
Владивосток, Россия
2 Санкт-Петербургский государственный университет
Санкт-Петербург, Россия
* E-mail: biomar@mail.ru
Поступила в редакцию 14.02.2022
После доработки 21.03.2022
Принята к публикации 14.04.2022
- EDN: EJGKWV
- DOI: 10.31857/S0030157422050215
Аннотация
В работе проанализированы данные десяти экспедиций, выполненных в северной части Японского моря (36°–47° с.ш. и 130°–141° в.д.), включая прибрежные воды России, в период с февраля по ноябрь в 2000–2015 гг., что позволило детально рассмотреть особенности первичного продуцирования в различные сезоны. Анализировались результаты судовых CTД-зондирований, концентрации биогенных веществ (БВ) и хлорофилла-а (Chl), а также спутниковые данные цвета океана. В начале весеннего периода максимальные величины Chl наблюдаются в поверхностном слое. В мае, после установления летней стратификации, основная масса Chl сосредоточена в слое 20–40 м. Осенью, в середине октября, начинается разрушение летней стратификации и происходит увеличение снабжения эвфотического слоя БВ, вследствие чего возрастает содержание Chl. Величина суммарной первичной продукции (P) в слое фотосинтеза по судовым оценкам оказалась примерно в два раза выше спутниковых оценок – 400 и 224 г С/м2 год, соответственно. Проанализированы причины таких различий.
ВВЕДЕНИЕ
Японское море соединяется мелкими проливами с соседними бассейнами – Охотским морем, Тихим океаном и Восточно-Китайским морем. Поскольку глубина проливов не превышает 130 м, формирование промежуточных, глубинных и донных вод происходит непосредственно в море [41]. Другая особенность моря состоит в том, что в нем происходит взаимодействие субарктических вод, которые формируются в северной части, с субтропическими водами, поступающими из Восточно-Китайского моря через Корейский пролив [41]. В районе взаимодействия этих вод (обычно между 39 и 41° с.ш.) образуется устойчивый субарктический фронт, разделяющий море на две климатические зоны [41]. Кроме того, в Японском море идут интенсивные процессы образования мезомасштабных вихрей [9]. Благодаря этим географическим и океанографическим особенностям Японское море часто рассматривают как океан в миниатюре [41]. Представления, полученные о процессах в Японском море можно, с известной долей условности, переносить на Мировой океан в целом. Сходство на этом не заканчивается. Известно, что в настоящее время происходят глобальные изменения в состоянии вод Мирового океана – уменьшение содержания кислорода [38] и величины рН [23]. Подобные процессы наблюдаются и для открытой части Японского моря [15, 42]. Наблюдаемые изменения, с общепринятой точки зрения, обусловлены стагнацией вод и ростом содержания углекислого газа в атмосфере. Дополнительно, эти изменения могут усиливаться ростом первичной продукции (P; используемые в статье обозначения и аббревиатура даны в таблице 1), обусловленной эвтрофикацией вод Японского моря. Такое предположение было сделано в работе [13].
Таблица 1.
Используемы в статье символы и аббревиатура
| Символ и аббревиатура | Расшифровка символа | Единицы измерения |
|---|---|---|
| Р | Первичная продукция | г С/м2 день |
| Рship | Первичная продукция на основе судовых измерений (модель POIM) | г С/м2 день |
| Рsat | Первичная продукция на основе спутниковых наблюдений (модель VGPM) | г С/м2 день |
| P b | Удельная максимальная скорость фотосинтеза по судовым данным (ассимиляционное число) | мг С/мг Chl ч |
| $P_{{{\text{Sat}}}}^{b}$ | Ассимиляционное число по спутниковым данным, как функция температуры воды | мг С/мг Chl ч |
| Chl | Концентрация хлорофилла-а | мг/м3 |
| Chlsat | Концентрация хлорофилла-а по спутниковым данным | мг/м3 |
| Chleu | Содержание хлорофилла-а в эвфотическом слое по судовым данным | мг/м2 |
| ChleuSat | Содержание хлорофилла-а в эвфотическом слое по спутниковым данным | мг/м2 |
| Chlopt | Концентрация хлорофилла-а в первом оптическом слое по судовым данным | мг/м3 |
| БВ | Биогенные вещества (P, N, Si) | мкМ |
| ФАР | Фотосинтетически активная радиация | (моль квантов/м2 день) |
| Io | Интенсивность света на поверхности моря | моль квантов/м2 час |
| Ic | Интенсивность света, соответствующая компенсационной глубине | (моль квантов/м2 час) моль квантов/м2 день |
| Ik | Интенсивность света, при которой световая кривая переходит к насыщению, равная ≈10% от падающей на поверхность воды ФАР | моль квантов/м2 час |
| Td | Долгота дня | час |
| kd490 | Коэффициент диффузного ослабления света в воде для длины волны 490 нм | м–1 |
| Zc | Компенсационная глубина | м |
| ZeuSat | Глубина эвфотического слоя, оцененная из спутниковых данных | м |
| Zmld | Глубина перемешивания верхнего слоя моря | м |
| Zcr | Критическая глубина, меньше которой скорость продукции в столбе воды превышает скорость потери биомассы | м |
| Zopt | Глубина первого оптического слоя | м |
| ТПО | Температура поверхности океана | градус Цельсия |
| PODAAC | Physical Oceanography Distributed Archive Center | |
| DIM | Depth-Integrated Models | |
| POIM | Pacific Oceanological Institute Model | |
| VGPM | Vertically Generalized Production Model |
Исследования P в северной части Тихого океана, в том числе и в Японском море, были начаты Институтом океанологии им. П.П. Ширшова в начале 60-х годов 20-го века на НИС “Витязь” методом радиоуглеродной метки С14. В этих экспедициях было установлено, что P в субарктической климатической зоне Японского моря превышала 500 мг С/м2 день, а в субтропической части находилась в пределах 250–500 мг С/м2 день [7, 8]. Обзор последних работ по исследованию P и продукционных характеристик вод Японского моря дан в [23, 27, 31]. Из этих обзоров следует, что большинство исследований выполнено на основе спутниковых наблюдений цвета океана со сканера SeaWiFS [14, 19–21] и с использованием модифицированного варианта модели VGPM (Vertically Generalized Production Model) [20]. Благодаря спутниковым наблюдениям ясно зафиксировано “цветение” фитопланктона для весеннего и осеннего сезонов, немного отличающееся по времени для разных климатических зон Японского моря [22, 29].
Ранее нами были проведены измерения содержания хлорофилла-а (Chl), биогенных веществ (БВ) и величины P радиоуглеродным методом в столбе воды в северной части Японского моря по результатам пяти экспедиций для весеннего и осеннего сезонов [3–5, 10]. Целью настоящей работы является оценка P в северной части Японского моря для всех сезонов с использованием двух интегрированных по глубине моделей восстановления P, которые относятся к типу DIM (Depth-Integrated Models) [20]. Данный тип моделей предполагает использование интегрированных от поверхности до глубины эвфотического слоя функций всех входящих в них переменных: часто это концентрация Chl и фотосинтетически активная радиация (ФАР). Немаловажным параметром таких моделей является также и удельная максимальная скорость фотосинтеза, которая обычно берется для слоя оптимальных для фотосинтеза условий. Величины P были рассчитаны по данным судовых наблюдений Тихоокеанского океанологического института им. В.И. Ильичёва (ТОИ ДВО РАН), выполненных в различные сезоны года, и спутниковой информации. Сравнение результатов позволило установить возможные причины ошибок в оценках P Японского моря и рассмотреть особенности сезонных изменений Chl, P и определяющих их абиотических факторов среды в северной части моря.
МАТЕРИАЛ И МЕТОДЫ
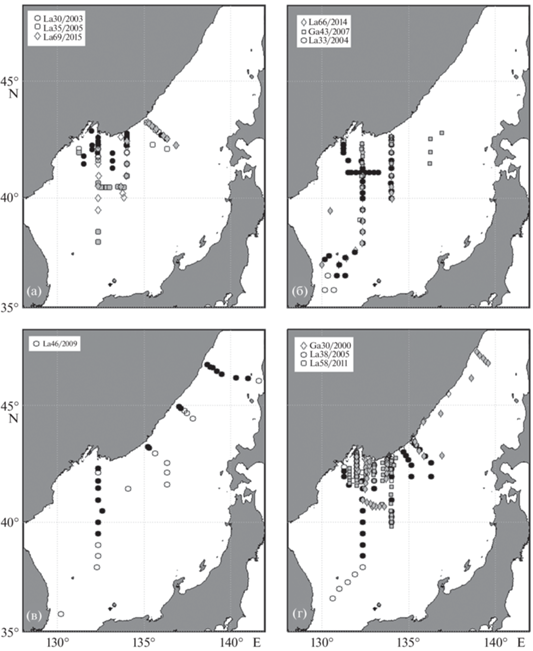
В работе проанализированы данные десяти экспедиций, выполненных в северной части Японского моря в районе 36°–47° с.ш. и 130°–141° в.д., включающей прибрежные воды России, в 2000–2015 гг. в периоды с февраля по ноябрь, охватывающие практически все четыре сезона (рис. 1). Экспедиции выполнялись на НИС “Академик М.А. Лаврентьев” (La) и “Профессор Гагаринский” (Ga). Для анализа использованы судовые данные: результаты CTД-зондирований, измерения концентрации БВ и Chl и спутниковые данные из баз Climate Change Initiative Ocean Colour, версия 3.1 и NASA Ocean Color Web [21].
Рис. 1.
Схемы расположения станций десяти экспедиций ТОИ ДВО РАН, выполненных в северо-западной части Японского моря в 2000–2015 гг. в период с февраля по ноябрь: (а) зима–начало весны, (б) весна, (в) лето и (г) осень. Значками круг, квадрат и ромб показаны станции различных экспедиций. Заполненные серым и черным значки означают наличие спутниковых данных на момент проведения судовых измерений.
Для оценки дневной P в слое фотосинтеза использованы две модели типа DIM: модель, разработанная в ТОИ ДВО РАН, POIM (Pacific Oceanological Institute Model) [3, 6] и модель VGPM (Vertically Generalized Production Model) [19, 20]. Для модели POIM использованы следующие судовые данные: температура воды по данным СТД-зондирований, концентрации БВ (NO3 + NO2 и PO4), полученные в соответствии с методикой [25] и концентрация Chl, измеренная спектрофотометрическим методом по методике [28]. Для оценки Р с применением модели VGPM в работе использованы следующие спутниковые данные: цвет океана (Chl и коэффициент диффузного ослабления света в воде для длины волны 490 нм, kd490) из базы CCI Ocean Colour, версия 3.1; дневная поверхностная ФАР из базы данных Ocean Productivity (NASA’s Ocean Colour Web); температура поверхности океана (ТПО) из базы данных PODAAC (Physical Oceanography Distributed Archive Center) [21, 37].
Модель POIM. В основе расчета Р лежит разработанная нами ранее модель непрямоугольной гиперболы для фотосинтеза, описывающей зависимость фотосинтеза от интенсивности света, полученная с учетом механизма фотосинтеза [3, 6, 44]:
(1)
$P = {{P}^{m}}\frac{{1 + {I \mathord{\left/ {\vphantom {I {{{I}_{k}}}}} \right. \kern-0em} {{{I}_{k}}}}}}{{2\gamma }}\left\{ {1 - \sqrt {1 - \frac{{4\gamma {I \mathord{\left/ {\vphantom {I {{{I}_{k}}}}} \right. \kern-0em} {{{I}_{k}}}}}}{{{{{(1 + {I \mathord{\left/ {\vphantom {I {{{I}_{k}}}}} \right. \kern-0em} {{{I}_{k}}}})}}^{2}}}}} } \right\}.$Результат численного интегрирования непрямоугольной гиперболы не зависит от толщины эвфотического слоя и абсолютного значения интенсивности света и равен 0.66. Таким образом, оценка величины дневной P в эвфотическом слое по судовым данным проводилась по следующему соотношению [3]:
Здесь P b – удельная максимальная скорость фотосинтеза или ассимиляционное число в подповерхностном слое, мг С/мг Chl ч; Chleu – содержание Chl в слое фотосинтеза, мг/м2, Td – долгота дня, час. На каждой станции суммированием концентрации Chl по вертикали от 0 до Zc определялось Chleu и P, рассчитанная по соотношению (2). Соотношение (2) соответствует предложенному ранее уравнению для расчета P в столбе воды (уравнение (12) в [20]). Необходимо отметить, что величина фактора F, используемая в данном уравнении, в нашем случае коэффициент 0.66 в уравнении (2), в моделях DIM весьма слабо зависит от дневной дозы ФАР. Поэтому зависимость P от ФАР учитывается длительностью светового дня.
Ассимиляционное число. К сожалению, мы не имели возможности определять P b во всех рейсах. Вместе тем, в осеннем рейсе Ga30 (12.10–03.11.2000) [3] нами были определены P b на всей акватории полигона методом 14С на борту судна: P b изменялось от 1.4 до 3.3 мг С/мг Chl ч и в среднем составляло 2.46 ± 0.87 мг С/мг Chl ч. В весеннем рейсе La33 (07–18.05.2004) P b были определены методом 14С на борту судна, их величина изменялась от 1.7 до 9.2 мг С/мг Chl ч и в среднем составляла 4.46 ± 0.95 мг С/мг Chl ч [5]. Кроме этого, в Амурском заливе и заливе Восток (северо-запад Японского моря) P b рассчитывали из P, определенной по изменению содержания кислорода в склянках. В августе 2008 в Амурском заливе, величина P b = 4.3 мг C/мг Chl ч [11], а в заливе Восток, в марте 2016 было получено значение P b = = 4.68 мг С/мг Chl ч [12]. По данным [7, 8] величина P b для Тихого океана при концентрации нитратов 0.3–2.0 мкМ составляла от 4.0 до 5.5 мг С/мг Chl ч, при меньшем содержании нитратов (<2 мкМ) – 2.0–3.5 мг С/мг Chl ч. По данным других авторов значение этой величины находится в пределах 1.8–6.3 мг С/мг Chl ч [43]. Таким образом, для весенних и летних месяцев мы брали для расчетов P P b = 4.5 мг С/мг Chl ч, а для осенних месяцев – 2.5 мг С/мг Chl ч.
Долгота дня и дневная доза ФАР. Известно, что величина P определяется, в первую очередь, падающим на растения светом. При расчете P используются разные световые характеристики: долгота дня, интенсивность света, дневная доза [24], эти три параметра могут быть рассчитаны на любой день года с помощью основной астрономической формулы [1, 30]:
(3)
${\text{sin(}}h{\text{)}} = {\text{cos}}(\varphi ){\text{cos}}(\delta ){\text{cos}}(\tau ) + {\text{sin}}(\varphi ){\text{sin}}(\delta ).$Здесь: h – высота солнца; φ – географическая широта; δ = 23°cos(2π(N + 9)/365) – склонение (N – порядковый номер дня года, начиная с 1 января); τ = 2πt/24 – часовой угол (t – время, час). Долгота дня определяется временем от восхода солнца до его захода. Величина ФАР на поверхности моря определяется высотой солнца h над горизонтом: I = I0sin(h), где I0 ≈ 6 моль квантов/м2 час – солнечная постоянная, соответствующая величине ФАР для безоблачного неба, когда солнце находится в зените, h = 90° [1, 24, 30]. Интегрируя уравнение (3) по времени в течение дня и умножая результат на I0, рассчитывали падающую на поверхность моря исследуемой акватории дневную дозу ФАР для соответствующего дня года (рис. 2а). Соотношение (3) позволяет также определить максимальную интенсивность света, когда солнце находится на максимальной высоте в любой день года. Долгота дня Td определялась нами с использованием формулы, приведенной в [35]. Для этого брались данные широты и дня года.
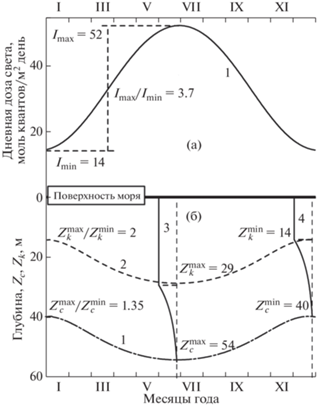
Рис. 2.
(а) – дневная доза ФАР (моль квант/м2 день), падающей на поверхность моря исследуемой акватории в различные сезоны (1) и соотношение доз лето/зима, (б) – компенсационная глубина, Zc (граница проникновения минимальной дозы света Ic ≈ ≈ 0.4 моль квантов/м2 день): (1) глубина насыщения фотосинтеза, Zk ≈ (Ik ≈ 4 моль квантов/м2 день), (2) их соотношение лето/зима; (3) кривая фотосинтеза летом и (4) зимой и их параметры (цифры), Японское море, 42° c.ш.
Глубина эвфотической зоны. В ряде случаев глубина эвфотического слоя измерялась традиционным методом – с помощью диска Секки. Однако этот показатель не всегда доступен измерениям. Например, измерения не могут быть проведены ночью, во время утренних и вечерних сумерек, а также в моменты сильного волнения, дрейфа судна, облачности т.д. По мнению ряда авторов, этот показатель часто не соответствует своему главному назначению – не характеризует глубину эвфотического слоя, т.к. не соответствует Zc [16, 24, 33]. Так, рост и фотосинтез фитопланктона зависит от абсолютного уровня радиации, а не от его процента. Исследователи давно предлагают вместо Zc использовать компенсационную дневную дозу света Ic в моль квантов/м2 день. Например, в зимние пасмурные дни в умеренных широтах может быть такая интенсивность света на поверхности моря, что в толще воды нет чистого фотосинтеза, хотя глубина 1% остается [16]. С физиологической точки зрения Ic более подходит для характеристики объекта, чем Zc. В литературе приводятся данные разных авторов о величине компенсационной дневной дозы света от ∼0.07 до ∼0.8 моль квантов/м2 день [16, 17, 24, 33]. Мы взяли среднюю величину 0.4 моль квантов/м2 день. В нашем случае глубина эвфотического слоя (продукционного слоя) Zс оценивалась для каждой станции двумя методами: (1) на основе данных о вертикальных профилях концентрации Chl и концентраций главных БВ; (2) на основе данных о компенсационной дневной дозе солнечного света Ic ≈0.4 моль квантов/м2 день [24].
Первый метод постулирует, что нижняя граница глубоководного максимума Chl соответствует нижней границе эвфотического слоя, т.е. компенсационной глубине Zc. Одновременно на этой границе наблюдается резкое повышение содержания нитратов (формирование “нитроклина”) [26, 33, 36].
Второй метод оценки глубины эвфотического слоя предполагает существование компенсационной дневной дозы солнечного света Iс, при которой продукция фитопланктона равна расходам на дыхание. Величина Zc оценивается по соотношению Zc = ln(Io/Ic)/kd, где kd – коэффициент диффузного ослабления света. В предыдущих работах установлено, что в районе исследований морская вода имеет достаточно высокую прозрачность, граница эвфотической зоны Zc составляла от 48 до 54 м, что соответствует коэффициенту диффузного ослабления света равному kd ≈ ≈ 0.095–0.085 м–1 [3–5]. По величине параметра kd акватория относится к типу II океанических вод [2]. При расчете глубины эвфотического слоя мы использовали величину kd, равную 0.09 м–1. Известно, что в районе исследований Td изменяется от 9 час в декабре до 15.2 час в июне, а дневная доза света – это 3.7 раза, от дневной минимальной дозы, что составляет примерно 14 моль квантов/м2 день в декабре до дневной максимальной дозы, что составляет примерно 52 моль квантов/м2 день в июне (рис. 2).
Таким, образом, величина Zc в течение года изменяется от $Z_{c}^{{{\text{min}}}}$ = ln(14/0.4)/0.09 = 39.5 м до $Z_{c}^{{\max }}$ = ln(52/0.4)/0.09 = 54.5 м (рис. 2б, кривая 1). Насыщение фотосинтеза происходит при интенсивности света Ik, которая, как правило, на порядок больше (10% от падающей радиации), чем Ic, и равна Ik ≈ 10 × Ic ≈ 4 моль квантов/м2 день (рис. 2б, кривая 2) [3, 6, 20]. Минимальная доза света в месте исследований $I_{D}^{{{\text{min}}}}$ = 14 моль квантов/м2 день в три раза превышает дозу насыщения фотосинтеза фитопланктона Ik ≈ 4.0 моль квантов/м2 день. Таким образом, уровень интенсивности света вполне достаточен для поддержания фотосинтеза фитопланктона в течение всего года.
Известно, что интенсивность света I в среде изменяется по закону Бугера–Ламберта–Бера. На рис. 3 показаны кривые фотосинтеза в зависимости от дневной дозы света в воде (в логарифмических координатах) для декабря (минимальная) и июня (максимальная). Для этих световых кривых продолжительность плато по глубине соизмерима с нелинейной частью, а глубины, соответствующие Ic равны 40 и 54 м в декабре и в июне, соответственно, т.е. отличаются в 1.35 раза, против 3.7 раза в воздухе (рис. 2 и 3).
Рис. 3.
Световая кривая фотосинтеза зимой (1) и летом (2) в логарифмических координатах (при равномерном распределении Chl): Ik = 4 – доза перехода к насыщению; $I_{D}^{{\min }}$ =14 и $I_{D}^{{\max }}$ = 52 – минимальная (зимой) и максимальная (летом) дневная световая доза, моль квантов/м2 день; ID – дневная световая доза; Z – глубина, м (Японское море, 42° c.ш.).

Модель VGPM. Дневную P в эвфотическом слое рассчитывали с помощью модели VGPM [19, 20]:
(4)
${{P}_{{{\text{sat}}}}} = 0.66125{{P}^{b}}Ch{{l}_{{{\text{sat}}}}}\frac{{{{I}_{0}}}}{{{{I}_{0}} + 4.1}}{{Z}_{c}}{{T}_{d}},$Массивы данных Chlsat и k490 представляют собой объединенную информацию с четырех спектрорадиометров: SeaWiFS, MODIS Aqua, MERIS и VIIRS с разрешением 4 × 4 км и дискретностью 8 суток, т.е. представляют собой среднее значение за этот промежуток времени. Из-за повышенной облачности в указанные периоды экспедиций, использование данных цвета океана с дискретностью 1 сутки не представлялось возможным, поскольку доступной для анализа информации было недостаточно. Данные ФАР со спектрорадиометра MODIS Aqua также имеют пространственное разрешение 4 × 4 км, а временнóе – 1 сутки. Изначально, данные ТПО с временны́м разрешением 1 сутки имели пространственное разрешение 0.01 × 0.01 градуса, но для соответствия другим спутниковым продуктам, используемым для восстановления P, были проинтерполированы на соответствующую сетку. Все данные брались так, чтобы станция попадала в область 4 × 4 км.
Считается, что Chlsat представляет собой среднюю концентрацию Chl в слое до первой оптической глубины, т.е. в слое из которого выходит около 90% процентов отраженной из воды солнечной радиации. Поэтому для сравнения спутниковых и судовых оценок поверхностной концентрации Chl (табл. 2, Chlsat и Chlopt соответственно) мы использовали именно этот слой, глубина которого определялась по формуле: Zopt= = 1/kd, где kd также использовался для всего спектра ФАР. Средние оценки Zopt для всех экспедиций не превысили 10 м.
Таблица 2.
Осредненные показатели параметров, анализируемых в работе, для всех экспедиций*)
| Экспедиция/ год | Сроки экспедиции | Долгота дня, час | Дневная
доза света, моль квантов/ м2 день |
Zc, м (1)//(2) | Chleu, мг/м2 | Chlopt, мг/м3 |
Chlsat, мг/м3 |
$P_{{{\text{Sat}}}}^{b}$, мг С/мг Chl ч |
Pship, г С/м2 день |
Psat, г С/м2 день |
|---|---|---|---|---|---|---|---|---|---|---|
| La30/2003 | 26.02–03.03 | 10.7 | 25.2 | 63//45 | 24 | 0.55 | 0.53 | 1.57 | 0.72 | 0.20 |
| La35/2005 | 12–25.03 | 11.7 | 32 | 60//48 | 28.7 | 0.34 | 0.36 | 2.48 | 0.84 | 0.40 |
| La69/2015 | 11.04–05.05 | 13.4 | 42.5 | 60//50 | 47.5 | 1.3 | 0.97 | 2.65 | 1.88 | 0.81 |
| La66/2014 | 21–29/04 | 13.6 | 44.5 | 60//52 | 41.5 | 0.75 | 0.56 | 3.67 | 1.67 | 0.60 |
| Ga43/2007 | 10–17.05 | 14.4 | 48.4 | 53//55 | 35 | 0.62 | 0.54 | 3.43 | 1.5 | 0.77 |
| La33/2004 | 07–18.05 | 14.4 | 48.4 | 48//54 | 37 | 0.63 | 1.09 | 4.34 | 1.58 | 1.22 |
| La46/2009 | 09–19.07 | 14.8 | 51 | 55//54 | 45 | 0.38 | 0.32 | 5.63 | 1.98 | 0.77 |
| La38/2005 | 15.10–01.11 | 10.2 | 20.5 | 57//44 | 19 | 0.34 | 0.47 | 5.69 | 0.52 | 0.75 |
| Ga30/2000 | 12.10–03.11 | 10.2 | 20.3 | 57//48 | 19.5 | 0.44 | 0.74 | 4.73 | 0.51 | 0.45 |
| La58/2011 | 27.10–08.11 | 9.8 | 18.3 | 55//43 | 47 | 0.67 | 0.46 | 4.63 | 0.87 | 0.58 |
РЕЗУЛЬТАТЫ
Начало зимне-весеннего “цветения” (La30, La35, La69). Съемки La30 (25.02–09.03.2003), La35 (16–24.03.2005) и La69 (11.04–05.05.2015) были проведены в зимнее и весеннее время, их результаты характеризуются всеми признаками переходного периода от зимней конвекции к началу весеннего “цветения” фитопланктона. Как видно из рис. 4, вовремя съемки La30 температура воды была низкой (<0.5°C) и практически постоянной во всем слое 0–600 м (рис. 4а). Это является результатом зимнего конвективного перемешивания, которое в Японском море достигает значительных глубин [13, 43]. Профили Chl, соединений азота (NO2 + + NO3) и фосфора (PO4) указывают на то, что продукция эвфотического слоя доминирует над деструкцией органического вещества – в верхнем слое концентрация Chl повышена, а концентрация БВ понижена из-за их ассимиляции (рис. 4б–4г).
Рис. 4.
Вертикальные профили температуры, T (а, д, и), хлорофилла-а, Chl (б, е, к), соединений азота, (NO2 + NO3) (в, ж, л), и фосфатов, PO4 (г, з, м) в рейсах La30 (а–г), La35 (д–з) и La69 (и–м) на разрезе меридиана 132.33° в.д.: Zc – компенсационная глубина; Zcr – критическая глубина (числа на профилях – номера станций).
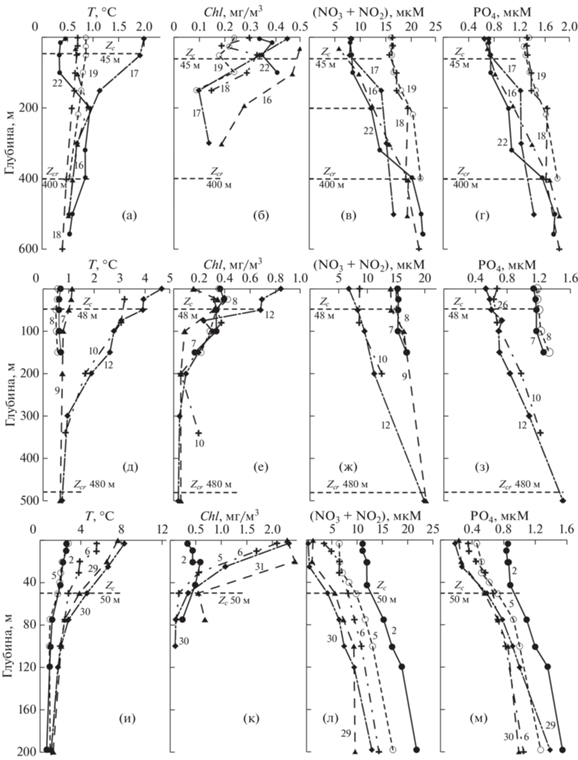
Повышенное содержание Chl (интегрированное по глубине) наблюдается не только в эвфотическом слое – 24 мг/м2 (табл. 1), но и на больших глубинах, при этом во всем слое до 300 м по нашим расчетам суммарное содержание Chl в 2–4 раза больше – до 70 мг/м2, чем в эвфотическом слое. Этот результат дополнительно свидетельствует об интенсивном вертикальном перемешивании вод, в результате которого клетки фитопланктона увлекаются конвекцией на глубины, существенно превышающие глубину эвфотического слоя.
По профилю Chl и БВ (первый метод) невозможно однозначно установить Zc. Она была оценена с помощью диска Секки и равнялась 45–47 м. Второй метод (Ic ≈ 0.4 моль/м2 день) дает близкую величину – Zc ≈ 45–48 м (рис. 3). В рейсе La30 величина Chleu составила 24 ± 11 мг/м2 (таблица 2), на всех станциях наблюдались высокие концентрации нитратов 5–15 мкМ (рис. 4в), а Pship в среднем была 0.72 ± 0.18 г С/м2 день.
Съемка, проведенная в более поздний (на 2 недели) период в рейсе La35 (16–24.03.2005), показала схожий результат: на станциях севернее 41° с.ш. температура, а также нитраты и фосфаты имели похожий вертикальный профиль, а повышенные значения Chl наблюдались до глубины 150–200 м (рис. 4д–4з, ст. 7–8). На станциях к югу от субарктического фронта (южнее 41° с.ш.) на верхних горизонтах температура воды была на 3–5°С выше, чем на глубинах (рис. 4д). Следовательно, в этот период, несмотря на интенсивное конвективное перемешивание, на южных станциях сохранялась вертикальная стратификация, обусловленная адвекцией вод из более южных областей и началом радиационного прогрева.
Как и в случае La30, профили Chl и БВ для La35 не позволяют однозначно оценить глубину Zc (оценка 1), поэтому для этого использовались измерения диска Секки, которые дают значениях в пределах 44–46 м. Второй метод дает схожую величину 45–48 м (рис. 4д–4з, табл. 1). Величина Chleu для этой экспедиции составила 29 ± 7 мг/м2, а Pship – 0.84 ± 0.21 г С/м2 день (табл. 2).
Первые станции разреза 1 (ст. 1–6) рейса La69 (11.04–05.05.2015) были выполнены 12–13 апреля, через примерно 3 недели после завершения съемки La35 (рис. 4и–4м). На ст. 2 и 5 температура воды на поверхности моря уже превышала температуру воды на глубине 50–100 м почти на 2°С; профили нитратов и фосфатов также уже отклонялись от вертикального. На южных стациях этого разреза (ст. 30–31), которые выполнялись в конце экспедиции (начало мая), температура воды поверхностного слоя была уже на 7–8°С выше в сравнении с глубинными горизонтами (рис. 4и). Концентрация Chl увеличивалась от 0.5 на севере (ст. 2) до 2 мг/м3 на юге (ст. 31, рис. 4к). Концентрации соединений азота и фосфора были выше там, где концентрация Chl была меньше (ст. 2) и ниже там, где концентрация Chl больше (рис. 4л–4м). На этом разрезе наблюдается переход от зимней конвекции (ст. 2) к началу весеннего “цветения” фитопланктона (ст. 5, 6) и далее к летней стратификации (ст. 29–30) (рис. 4и–4м).
Во время рейса La69 (11.04–05.05.2015) начинается бурное развитие фитопланктона, концентрация Chl возрастает выше 2 мг/м3, интеграционная величина Chleu достигает 47.5 ± 17 мг/м2, а Pship – 1.88 ± 0.45 г/м2 день, т.е. это период начала весеннего “цветения” фитопланктона.
Весна, формирование летней стратификации (La66, Ga43, La33). Весной, в апреле и мае месяце были проведены три съемки: La66 (14–29.04.2014), Ga43 (11–17.05.2007) и La33 (07–18.05.2004). В этот период в районе устанавливалась летняя стратификация (рис. 5).
Рис. 5.
Вертикальные профили температуры, T (а, д, и), хлорофилла-а, Chl (б, е, к), соединений азота, NO2 + NO3 (в, ж, л), и фосфатов, PO4 (г, з, м) в рейсах Ga43 (а–г), La66 (д–з) и La33 (и–м) на разрезе меридиана 132.33° в.д.: Zc – компенсационная глубина (числа на профилях – номера станций).
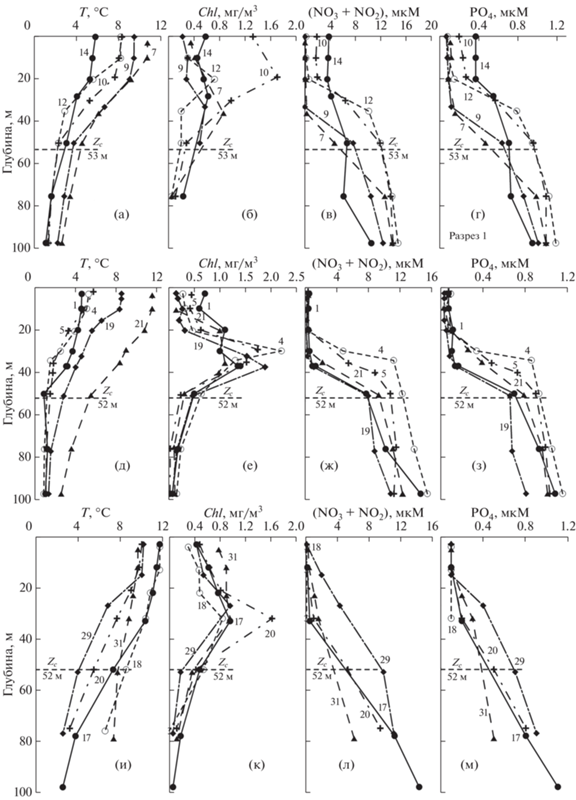
Температура на поверхности моря была на 4–12°С выше в сравнении с глубинными горизонтами, вода прогревалась до 50 м (рис. 5а, 5д, 5и). Формы вертикальных профилей Chl отличаются от тех, которые были получены в рейсах зимне-весеннего периода (La30, La35, La69). Содержание Chl в поверхностном слое уменьшается, более высокие концентрации наблюдаются в слое 20–40 м, где они достигают 2 мг/м3 (рис. 5б, 5е, 5к). Во всех съемках отмечается ассимиляция азота и фосфора до глубин около 20 м, а в некоторых случаях – до 40 м (рис. 5в–5г, 5ж–5з, 5л–5м). При этом соединения азота усваиваются фитопланктоном с большей скоростью, чем фосфора.
В апреле–мае увеличилась долгота дня и дневная доза света: Td ≈ 13.5–14.3 часов, ID ≈ 44–47 моль квантов/м2 день. В то же время сохранялось достаточно интенсивное снабжение эвфотического слоя БВ. Эти факторы создавали благоприятные условия для роста фитопланктона: среднее значение Chleu составляло для съемки La66 41.5 ± 16.6 мг/м2, а для Ga43 и La33 – 35 ± 10 и 37 ± 10.5 мг/м2, соответственно. Во время съемки La33 величина P b в среднем составляла 4.34 мг С/мг Chl ч. Расчеты показали, что для этой съемки величина Рship при учете P b, измеренного для станций по всему полигону, составила 1.87 ± ± 0.9 г С/м2 день. Для этого же рейса при среднем значении P b, величина Рship была 1.58 ± 0.45 г С/м2 день, т.е. примерно на 15% меньше. Используя среднее значение P b, полученное в экспедиции La33, мы оценили Рship для съемок, выполненных в других рейсах: La66 – 1.67 ± 0.63 и Ga43 – 1.5 ± ± 0.5 г С/м2 день, соответственно.
Сопоставление профилей Chl и P для рейса La33 (в котором величина P b определялась с помощью 14С на борту судна) показывает, что максимум Р расположен на той же глубине, что и максимум Chl [5]. Таким образом, максимальная продукция соответствует глубине, где синтезируется максимальное количество Chl.
Выше указывалось, что в этих съемках основная масса Chl наблюдается в слое 20–40 м, а в первом оптическом слое по нашим данным концентрация Chl в 2–5 раза меньше, чем на глубине. Спутник регистрирует именно эту меньшую концентрацию, поэтому толщина ZeuSat также меньше, чем Zс и ChleuSat примерно в 2 раза меньше, чем Chleu. Температура в это время выше (8–12°C), величина P b тоже выше – $P_{{{\text{Sat}}}}^{b}$ ≈ 3.7–4.4 мг С/мг Chl ч (против P b ≈4.5 мг С/мг Chl ч). В результате PSat ≈ ≈ 0.5Pship. В это время первый и второй пути оценок Zc различались незначительно и составляли 52–60 м (рис. 5, табл. 2).
Летне-осенний период (La46, La38, Ga30 и La58). Летом и осенью были проведены четыре съемки: La46 (09–19.07.2009), La38 (15.10–01.11.2005), Ga30 (12.10–03.11.2000) и La58 (27.10–08.11.2011). Их результаты отличаются от весенних более высокими градиентами температуры в слое скачка БВ (сравните рис. 6а–6з и рис. 5д–5м). Летняя съемка La46 отличается высокой концентрацией Chl, его интегральная величина Chleu составляла 45 ± 10 мг/м2. Здесь видно, что в первом оптическом слое содержание Chlopt ≈ 0.38 мг/м3 в 3–4 раза меньше, чем в максимуме слоя фотосинтеза на глубине 20–40 м (рис. 5б; табл. 2). В период этой съемки наблюдается интенсивное снабжение БВ эвфотического слоя (рис. 6в–6г), которое обеспечивает развитие фитопланктона и высокую Pship – 1.98 ± 0.4 г С/м2 день (табл. 2).
Рис. 6.
Вертикальные профили температуры, T (а, д, и), хлорофилла-а, Chl (б, е, к), соединений азота, (NO2 + NO3) (в, ж, л), и фосфатов, PO4 (г, з, м), в рейсах La46 (а–г), La38 (д–з) и La58 (и–м) на разрезе меридиана 132.33° в.д.: Zc – компенсационная глубина (числа на профилях – номера станций).
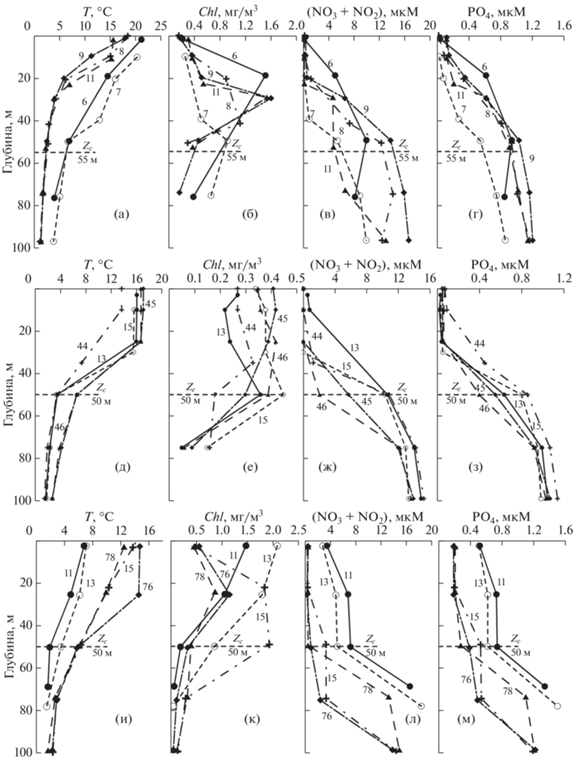
Осенняя съемка La38 (конец октября) проводилась накануне разрушения летней стратификации. Сохраняющийся высокий температурный градиент препятствует поступлению БВ в эвфотический слой, вследствие чего наблюдаются низкие концентрации Chl (до ≈0.5 мг/м3) и, соответственно, низкая величина Chleu = 19 ± 9 мг/м2 (рис. 6д–з). Такие же параметры наблюдались и при съемке Ga30, выполненной нами осенью 2000 г. [3]. Эти измерения показали величину P b = = 2.46 ± 0.87 мг С/мг Chl ч. Вместе с тем, ее спутниковая оценка была в вдвое выше $P_{{{\text{sat}}}}^{b}$ = 5.7 ± ± 0.87 мг С/мг Chl ч. Из-за того, что P b ≈ 0.5$P_{{{\text{sat}}}}^{b}$, интегральная продукция по судовым оценкам оказалась в 2 раза ниже, чем по спутниковым: Pship = 0.4 и Psat = 0.75 г С/м2 день [7] (табл. 2; рис. 7).
Рис. 7.
Сезонная изменчивость оценок первичной продукции по судовым (1) и спутниковым (2) данным, P (г С/м2 день), полученных в экспедициях в разные сезоны и годы (табл. 1–2) и дневной дозы солнечного света (3) (моль квантов/м2 день).

Более поздняя осенняя съемка La58 (27.10–08.11) проводилась в период разрушения летней стратификации (уменьшение температурного градиента), что приводило к более интенсивному снабжению БВ эвфотического слоя и, вследствие этого, более высоким концентрациям Chl, превышающими 2.0 мг/м3 (рис. 6и–6м). Во время рейса La58 была минимальная долгота дня (9.7 час) и дневная доза солнечной радиации (18.3 моль квантов/м2 день), несмотря на это в эвфотическом слое синтезировалось много Chl (47 ± 16 мг/м2) и отмечалась высокая PShip (0.87 ± 0.27 г С/м2 день). Величины PShip и PSat в данном случае отличались незначительно (табл. 2; рис. 7).
На рис. 7 графически представлена временнáя изменчивость P по судовым и спутниковым данным. Как указано выше, в марте (экспедиции La30 и La35) даже в условиях зимней конвекции в эвфотическом слое наблюдалось достаточно высокое значение Chleu и, так как PShip восстанавливалась через Chleu, то ее величины были также относительно высокими – 0.7–0.8 г С/м2 день (рис. 7, табл. 1). При этом PSat была вдвое меньше. Через месяц, в апреле (экспедиции La69 и La66) Р резко увеличивается: PShip – до 1.7–1.9 г С/м2 день, PSat – до 0.6–0.8 г С/м2×день. В мае (экспедиции Ga43 и La33) эти показатели незначительно уменьшаются по судовым оценкам, однако, увеличиваются по спутниковым. В июле (экспедиция La46) PShip достигает своих максимальных значений, превышая весенние: при Chleu, равном 45 мг/м2, PShip составила примерно 2.0 г С/м2 день. Однако по спутниковым оценкам наблюдается обратная ситуация – максимальные величины PSat, приуроченные к весеннему “цветению” фитопланктона, наблюдаются во второй половине мая (экспедиция La33), после чего наступает снижение первичного продуцирования и развития фитопланктона.
К сожалению, в августе и сентябре съемки не проводились. Осенью (экспедиции La38 и Ga30) PShip перед разрушением летней стратификацией из-за падения дневной дозы света характеризуется низкими величинами – около 0.4 г С/м2 день для обеих экспедиций, в то время как PSat близка к апрельским величинам. С разрушением летней стратификации и подтоком БВ в эвфотический слой PShip резко возрастает (экспедиция La58), а PSat, наоборот, падает, что связано с невозможностью спутникового спектрорадиометра регистрировать сигнал из слоя, расположенного ниже слоя первой оптической глубины, где в это время года сосредоточена основная масса продуцирующего фитопланктона.
На основании приведенных данных (табл. 2, рис. 7, 8) нами была оценена годовая P, которая составила 400 г C/м2 год для PShip и 225 г C/м2 год для PSat. При этом в течение года Chleu изменялась немногим более чем в 2 раза, от 20 до 47 мг/м2.
Рис. 8.
Диаграмма рассеяния судовых и спутниковых оценок первичной продукции (г С/м2 день) для всех исследуемых станций за период 2000–2015 гг. Сплошная линия — линия соответствия, пунктирная линия — линия линейной регрессии.
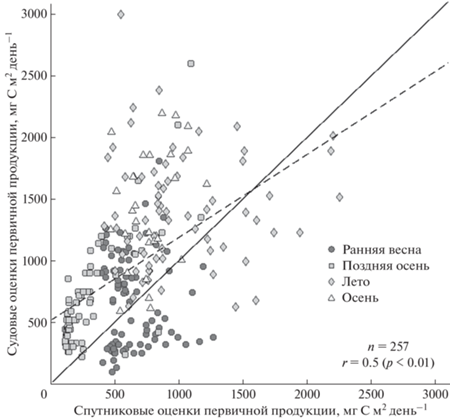
На рис. 8 представлена диаграмма рассеяния для судовых и спутниковых оценок P для всех исследуемых в работе станций. Корреляция между ними небольшая и составила 0.5 при высоком уровне значимости мене 0.01. Отклонение линии линейной регрессии от линии соответствия показывает, что спутниковые оценки могут быть как занижены, так и завышены во все рассматриваемые сезоны в зависимости от особенностей акватории. Однако на большом количестве станций наблюдались все же более низкие значения спутниковых оценок P по сравнению с судовыми, что также видно из средних величин P по экспедициям в отдельности (рис. 7, табл. 2).
ОБСУЖДЕНИЕ
На рис. 4 представлена полная картина перехода от режима зимней конвекции к режиму летней стратификации. Из наших результатов следует (табл. 2), что весенняя “вспышка” (La69 – Chleu = = 47.5 мг/м2, PShip = 1.88 г C/м2 день) характеризуется бурным развитием фитопланктона и, соответственно, высокой продуктивностью. Этому способствует интенсивное снабжение эвфотического слоя БВ и высокая дневная доза света ID ≈ ≈ 44 моль квантов/м2 день. Таким образом, с прекращением зимней конвекции и началом прогрева верхнего слоя моря наступает весеннее “цветение” фитопланктона. Это явление широко обсуждается в литературе [15, 18, 25, 33, 41, 45 ] . Для объяснения “цветения” фитопланктона Свердрупом была предложена концепция критической глубины, Zcr, т.е. глубины, меньше которой интегральная дневная продукция столба воды превышает интегральную величину затрат на дыхание [41]. Согласно этой концепции, весеннее “цветение” фитопланктона может начаться только тогда, когда глубина перемешивания верхнего слоя моря (Zmld) становится меньше Zcr, которая, как правило, в 510 раз больше Zc [18, 24, 29, 39]. Свердруп предложил уравнение связи между критической глубиной, интенсивностью ФАР на поверхности моря (Io) и интенсивностью ФАР на компенсационной глубине (Ic):
(5)
$\frac{{{{Z}_{{{\text{cr}}}}}}}{{1 - {{e}^{{ - {{k}_{d}}{{Z}_{{{\text{cr}}}}}}}}}} = \frac{1}{{{{k}_{d}}}}\frac{{{{I}_{0}}}}{{{{I}_{с}}}}.$Поскольку член ${{e}^{{ - {{k}_{d}}{{Z}_{{{\text{cr}}}}}}}}$ ⪡ 1, то соотношение (5) упрощается [29]:
Тогда, учитывая, что интенсивность света на поверхности моря в период выполнения работ La30 была 25 моль квантов/м2 день (рис. 2а) и, принимая kd ≈ 0.1 м–1, Ic = 0.4 моль квантов/м2 день, из соотношения (6) получим Zcr ≈ 625 м. Эта величина согласуется с тем фактом, что в этом районе в зимнее время на отдельных станциях вертикальное перемешивание достигает 1700 м [40, 41], что превышает значение, рассчитанное по уравнению (6). В целом для Японского моря максимальная глубина перемешанного слоя отмечается в феврале и по среднемноголетним данным находится в пределах 100–200 м [31]. Однако для субарктической области, где конвекция достигает значительных глубин, она может намного превышать эти значения [40, 41].Реальная ситуация в период интенсивного вертикального перемешивания оказывается более сложной. Как следует из рис. 4, концентрация Chl в верхнем слое повышена, а концентрация БВ понижена (рис. 4б–4г), что указывает на наличие продукции. В результате глубокого конвективного перемешивания повышенное содержание Chl наблюдается не только в эвфотическом слое, но и на нижележащих горизонтах до 150–300 м. При этом во всем слое интегральное содержание Chl в 2–4 раза больше, чем в эвфотическом (рис. 4). К похожему результату пришел автор работы [18] на примере Северной Атлантики, сделав вывод, что чистая продукция в полном столбе воды больше до начала весеннего “цветения”, а сам процесс роста фитопланктона начинается в середине зимы. Он предлагает отказаться от гипотезы Свердрупа. Очевидно, что аналогичный эффект наблюдается и в Японском море, но в этом направлении необходимы дальнейшие исследования.
После “цветения” в весенний период (апрель–май) в районе можно отметить следующие изменения: устанавливается летняя стратификация (рис. 5), температура на поверхности моря на 4–12°С превышает значения на подповерхностых горизонтах, верхний слой вод перемешивается и прогревается до 50 м, концентрация Chl в поверхностном слое уменьшается, его основная масса наблюдается в слое 20–40 м, отмечается ассимиляция азота и фосфора до глубин более 20 м, дневная доза света увеличивается до ID ≈ 45–47 моль квантов/м2 день. Эти факторы также создают благоприятные условия для роста фитопланктона. Вместе с тем, по мере установления летней стратификации, наблюдается снижение Chleu от 47.5 ± 17 мг/м2 (La69) до 41.5 ± 16.6 мг/м2 (La66) и до 35 ± 10 мг/м2 (Ga43) и величина PShip, соответственно, уменьшается от 1.88 ± 0.45 г/м2 день до 1.67 ± 0.9 и до 1.5 ± 0.5 гС/м2 день. Таким образом, в течение месяца продукционные параметры, Chleu и PShip, уменьшились примерно на 25%.
В этих съемках основная масса Chl наблюдается в слое 20–40 м, тогда как в первом оптическом слое концентрация Chl в 2–5 раза меньше. Спутниковый радиометр регистрирует именно эту меньшую концентрацию. Поэтому по спутниковым данным толщина эвфотического слоя примерно в два раза меньше, чем по судовым (ZeuSat ≈ ≈ 0.5ZeuShip), а поскольку ассимиляционные числа равны ($P_{{{\text{Sat}}}}^{b}$ ≈ $P_{{{\text{Ship}}}}^{b}$), то в результате величина PSat, также заметно ниже судовой (PSat ≈ 0.5PShip).
Однако, летняя съемка (La46, июль), как и весенние, отличается также высокой величиной Chleu = 45 ± 10 мг/м2 и PShip ≈1.98 ± 0.4 г С/м2 день. По нашим данным не наблюдалось летнего падения P (рис. 7, табл. 2), как это следует из работ [29, 43].
В начале осеннего периода (La38, конец октября) еще сохраняется летняя стратификация, но уменьшение долготы дня и световой дозы приводит к резкому снижению Chleu до 19 ± 9 мг/м2 и PShip до 0.52 ± 0.15 г С/м2 день.
В более поздний осенний период (La58, 27.10–08.11), когда происходит разрушение летней стратификации, приводящее к более интенсивному снабжению БВ эвфотического слоя и, вследствие этого, более высоким величинам Chleu = = 47 ± 16 мг/м2 и PShip ≈ 0.87 ± 0.27 г С/м2 день. Что свидетельствует об осеннем “цветении”, описанном во многих работах [24, 29, 43].
Для понимания причины расхождения судовых и спутниковых оценок Р, нами были проведены расчеты среднего содержания Chl в первом оптическом слое (Chlopt) и его сравнение с Chlsat. Результаты, представленные в таблице 2, показывают разумное согласие между двумя измерениями, за исключением рейсов La69 и La33. Причины расхождения для этих двух случаев могут быть разные. Наблюдения в рейсе La69 пришлись на период “цветения” фитопланктона. Как известно, в этот период концентрация Chl в эвфотическом слое чрезвычайно изменчива по времени и пространству, и в этом случае возникает повышенная неопределенность в оценке содержания Chl в первом оптическом слое. Во время экспедиции La33 наблюдался широкий по глубине максимум концентрации Chl (рис. 5к) и, по-видимому, спутниковые измерения неадекватно оценивали такой тип вертикального распределения Chl, давая почти вдвое завышенные результаты.
Сопоставление годовой кривой снабжения светом поверхности моря и Chl показывает, что дневная доза солнечного света является важным внешним фактором создания первичной продукции в северной части Японского моря (рис. 1, 7). Несмотря на то, что исследования проводились в течение 15 лет, общая тенденция сохранялась. Исключением является осенний период, когда разрушается летняя стратификация и содержание Chl возрастает примерно вдвое. По данным [37], PSat на севере Японского моря, составляет около 170 гС/м2 год, в весенний период спутник регистрирует небольшую положительную P ≈ 0.2 гС/м2 день. Как правило, спутник регистрирует практически вдвое меньшую величину P, чем судовые измерения – PShip/PSat = 400/225 ≈ 1.8 (рис. 7, 8). Можно выделить три причины занижения PSat. Во-первых, занижение величины P b в холодное время года примерно 2 раза. Во-вторых, во время летней стратификации основная масса Chl находится в слое 20–40 м, которую спутниковый радиометр не регистрирует, поскольку это глубже слоя первой оптической глубины [4, 5]. В-третьих, спутниковые оценки занижают толщину эвфотического слоя. Совокупность этих факторов, по нашему мнению, и является причиной этого различия.
ЗАКЛЮЧЕНИЕ
Анализ данных десяти экспедиций, выполненных в северной части Японского моря в различные сезоны, и массивов спутниковых данных позволил рассмотреть особенности изменения P в северной части моря в течение практически всего года. Судовые измерения показали, что процесс первичного продуцирования на этой акватории не прекращается во все рассматриваемые сезоны. Достаточно высокое содержание Chl и высокий уровень P отмечается даже в условиях зимней конвекции, что связано с большой глубиной вертикального перемешивания в Японском море. В весенний период максимальные величины Chl и P наблюдаются в поверхностном слое. Начиная с мая, после установления летней стратификации основная масса Chl и P сосредоточены в слое 20–40 м. Осенью, в середине октября начинается разрушение летней стратификации, в начале ноября понижается температурный градиент и увеличивается снабжение БВ эвфотического слоя, вследствие чего содержание Chl возрастает. При этом нами не зарегистрировано падения P в летний сезон, снижение продукции отмечается в периоды после весеннего “цветения” и перед разрушением стратификации в начале осени. Величина суммарной P во всем слое фотосинтеза для всего периода исследований по судовым оценкам оказалась примерно в два раза выше спутниковых оценок – 400 и 224 г С/м2 год, соответственно. Причиной этого различия может служить совокупность факторов – заниженные в холодное время года спутниковые оценки удельной оптимальной скорости фотосинтеза, которая выступает множителем в моделях P, невозможность регистрации со спутника Chl в слое оптимальных для фотосинтеза условий во время летней стратификации, который располагается ниже слоя первой оптической глубины, доступной спектрорадиометру, и заниженные спутниковые оценки толщины эвфотического слоя.
Источники финансирования. Работа выполнена при финансовой поддержке программы фундаментальных научных исследований государственных академий наук на 2021–2023 гг. (темы 0211-2021-0008 “Исследование основных процессов, определяющих состояние и изменчивость океанологических характеристик окраинных морей Азии и прилегающих районов Тихого и Индийского океанов” и 0211-2021-0014 “Эколого-биогеохимические процессы в морских экосистемах: роль природных и антропогенных факторов”).
Список литературы
Бартенева О.Д., Полякова Е.А., Русин Н.П. Режим естественной освещенности на территории СССР // Л.: Гидрометеоиздат, 1971. 240 с.
Ерлов Н.Г. Оптика моря. Л.: Гидрометеоиздат, 1980. 248 с.
Звалинский В.И., Лобанов В.Б., Захарков С.П., Тищенко П.Я. Хлорофилл, замедленная флуоресценция и первичная продукция в северо-западной части Японского моря осенью 2000 г. // Океанология. 2006. Т. 46. № 1. С. 27–37.
Звалинский В.И., Лобанова П.В., Тищенко П.Я., Лобанов В.Б. Оценка первичной продукции северо-восточной части Японского моря на основе судовых и спутниковых наблюдений в осенний период // Известия ТИНРО. 2018. Т. 195. С. 184–200.
Звалинский В.И., Лобанова П.В., Тищенко П.Я., Лобанов В.Б. Оценка первичной продукции северо-западной части Японского моря на основе судовых и спутниковых наблюдений // Океанология. 2019. Т. 59. №1. С. 45–55. https://doi.org/10.31857/S 0030-15745945-55
Звалинский В.И., Тищенко П.Я. Моделирование фотосинтеза и роста морского фитопланктона // Океанология. 2016. Т. 56. № 4. С. 1–15.
Кобленц-Мишке О.И. Величина первичной продукции Тихого океана // Океанология. 1965. Т. 5. № 2. С. 325–337.
Кобленц-Мишке О.И., Ведерников В.И. Первичная продукция // Биология океана. М.: Наука, 1977. С. 183–209.
Лобанов В.Б., Пономарев В.И., Салюк А.Н. и др. Структура и динамика синоптических вихрей северной части Японского моря // Дальневосточные моря России. Океанологические исследования. Наука: Москва, 2007. Т. 1. С. 450–473.
Лобанова П.В., Звалинский В.И., Тищенко П.Я. Первичная продукция фитопланктона и концентрация хлорофилла-а в западной части Японского моря по спутниковым и натурным данным // Современные проблемы дистанционного зондирования земли из космоса. 2017. Т. 14. Вып. 2. С. 135–147.
Тищенко П.П., Звалинский В.И., Тищенко П.Я., Семкин П.Ю. Первичная продукция Амурского залива (Японское море) в летний сезон 2008 года // Биология моря. 2017. Т. 43. Вып. 3. С. 195–202.
Тищенко П.П., Тищенко П.Я., Еловская О.А. и др. Условия формирования первичной продукции фитопланктона в заливе Восток (Японское море) весной 2016 г. // Известия ТИНРО, 2019. Т. 198. С. 164–185.
Тищенко П.Я., Талли Л.Д., Недашковский А.П. и др. Временнáя изменчивость гидрохимических свойств Японского моря // Океанология, 2002. Т. 42. № 6. С. 838–847.
Шамбарова Ю.В., Степочкин И.Е., Захарков С.П. Исследование изменчивости первичной продукции в Японском море по спутниковым данным на основе ЭОФ-анализа // Современные проблемы дистанционного зондирования Земли из космоса. 2015. Т. 12. № 1. С. 80–92.
Alin S., Evans W., Gao Z. et al. Ocean Acidification and Deoxygenation in the North Pacific Ocean. PICES Special Publication 5 / Eds. Christian J.R., Ono T. // Sidney, B.C., Canada, The North Pacific Marine Science Organization, c/o Institute of Ocean Sciences, 2019. 116 p.
Banse K. Should we continue to use the 1% light depth convention for estimating depth of phytoplankton for another 70 years? // Buttetin Limnol. Oceanogr. 2004. V. 13. № 3. P. 49–52.
Banse K., Postel J.R. On using pigment-normalized? Light-saturated carbon uptake with satellite-derived pigment for estimating column photosynthesis // Global Biogeochem. Cycles. 2003. V. 17. № 3. P. 1079. 2003.https://doi.org/10.1029/2002GB002021
Behrenfeld M.J. Abandoning Sverdrup’s Critical Depth Hypothesis on phytoplankton blooms // Ecology. 2010. V. 91. № 4. P. 977–989.
Behrenfeld M.J., Falkowski P.G. Photosynthetic rates derived from satellite based chlorophyll concentration // Limnology and Oceanography. 1997a. V. 42. № 1. P. 1–20.
Behrenfeld M.J., Falkowski P.G. A consumer’s guide to phytoplankton primary productivity models // Limnology and Oceanography. 1997b. V. 42. № 7. P. 1479–1491.
Climate Change Initiative (CCI) Ocean Colour Web. URL: https://www.oceancolour.org/.
Choi J.K., Noh J.H., Orlova T. et al. Phytoplankton and Primary Production // Oceanography of the East Sea (Japan Sea) / Springer Publishing. Switzerland, 2016. Ch. 10. P. 217–245. https://doi.org/10.1007/978-3-319-22720-7_10
Dore J.E., Lukas R., Sadler D.W. et al. Physical and biogeochemical modulation of ocean acidification in the central North Pacific // Proceedings National Academy Sciences, 2009. V. 106. № 30. P. 12235–12240.
EEOS 630 Biol. Ocean. Processes. Sverdrup’s critical depth concept and the vernal phytoplankton bloom. Chapter 11. Revised: 10/28/08. Ed. by E.D. Gallagher. 2010. P. 1–40.
Hansen H.P., Koroleff F. Determination of nutrients. Methods of Seawater Analysis (Eds. K. Grasshoff, K. Kremling, M. Ehrhardt), 3rd Edition, Weinheim, New York, Chiester, Brisban, Singapore, Toronto: Willey-VCH. 1999. P. 159–251.
Hickman A.E., Moore C.M., Sharples J. et al. Primary production and nitrate uptake within the seasonal thermocline of a stratified shelf sea // Mar. Ecol. Prog. Ser. 2012. V. 463. P. 30–57.
Ishizaka J., Yamada K. Phytoplankton and Primary Production in the Japan Sea // Remote Sensing of the Asian Seas / V. Barale and M. Gade (eds.). Springer International Publishing AG, part of Springer Nature. 2019. P. 177–189. https://doi.org/10.1007/978-3-319-94067-0_9.
Jeffrey S.W., Humphrey G.F. New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton // Biochem Physiol Pflanz. 1975. V. 167. P. 191–194.
Kim S.W., Saitoh, S., Ishizaka J. et al. Temporal and spatial variability of phytoplankton pigment concentration in the Japan Sea derived from CZCS images // J. of Ocean. 2000. V. 56. № 5. P. 527–538.
Kirk J.T. The nature and measurement of the light environment in the ocean // Primary production and biogeochemical cycles in the sea. New York: Plenum Press, 1992. P. 9–29.
Lim S.-H., Jang C.J., Oh I.S., Park J.-J. Climatology of the mixed layer depth in the East/Japan Sea // Journal of Marine Systems. 2012. V. 96–97. P. 1–14.
Lobanova P., Tilstone G.H., Bashmachnikov I., Brotas V. Accuracy assessment of primary production models with and without photoinhibition using Ocean-Colour Climate Change Initiative data in the north east Atlantic ocean // Remote Sensing. 2018. V. 10. № 7. 1116. https://doi.org/10.3390/rs10071116
Marra J. The compensation irradiance for phytoplankton in nature // Geophys. Res. Lett.2003. V. 31. L06305. https://doi.org/10.1029/2003GL018881
Morel A., Huot Y., Gentili B. et al. Examining the consistency of products derived from various ocean color sensors in open ocean (Case1) waters in the perspective of a multi-sensor approach // Remote Sensing of Environment. 2007. https://doi.org/10.1016/j.rse.2007.03.012
Ocean Productivity Home Page URL: http://www.science.oregonstate.edu/ocean.productivity/index.php
Painter S.C., Sanders R., Poulton A.J. et al. Nitrate uptake at photic zone depths is not important for export in subtropical ocean // Global Biogeochem Cycles. 2007. 21:GB4005. https://doi.org/1029/2006GB002807
PODAAC (Physical Oceanography Distributed Archive Center) URL: https://podaac.jpl.nasa.gov/.
Schmodtko S., Stramma L., Visbeck M. Decline in global oceanic oxygen content during the past five decades // Nature. 2017. V. 542. P. 335–339. https://doi.org/1038/nature21399
Sverdrup H.U. On conditions for the vernal blooming of phytoplankton // J. Conseil perm. int. Explor. Mer. 1953. V. 18. P. 287–295.
Talley L.D., Lobanov V., Ponomarev V. et al. Deep convection and brine rejection in the Japan Sea// Geophys. Res. Lett. 2003. V. 30. № 4. P. 1159.
Talley L., Min D.-H., Lobanov V. et al. Japan/East Sea Water Masses and their Relation to the Sea’s Circulation // Oceanography. 2006. V. 19. P. 32–49.
Tishchenko P., Lobanov V., Kaplunenko D. et al. Acidification and deoxygenation of the northwestern Japan/East Sea // J. Mar. Sci. Eng. 2021. V. 9. P. 953. https://doi.org/10.3390/ jmse9090953
Yamada K., Ishizaka J., Nagata H. Spatial and temporal variability of satellite estimated primary production in the Japan Sea from 1998 to 2002 // J. Oceanogr. 2005. V. 61. P. 857–869.
Zvalinsky V. I. A new approach to the modeling of marine ecosystems // PICES Scientific Report. 2001. № 17. P. 43–59.
Дополнительные материалы отсутствуют.