

Океанология, 2022, T. 62, № 6, стр. 965-978
Значение находки колониальных радиолярий в верхнеюрских отложениях арктической зоны Западной Сибири (полуостров Ямал)
В. С. Вишневская 1, 2, *, Э. О. Амон 2, Ю. А. Гатовский 3
1 Геологический институт РАН
Москва, Россия
2 Палеонтологический институт им. А.А. Борисяка РАН
Москва, Россия
3 Московский государственный университет имени М.В. Ломоносова
Москва, Россия
* E-mail: valentina.vishnaa@mail.ru
Поступила в редакцию 19.11.2021
После доработки 14.04.2022
Принята к публикации 28.04.2022
- EDN: VCHZQM
- DOI: 10.31857/S0030157422050203
Аннотация
Рассмотрено теоретическое и практическое значение пионерной находки колониальных радиолярий Siphonosphaera yamalica Vishnevskaya et Amon из семейства Collosphaeridae в ранневолжском радиоляриевом комплексе из баженовского горизонта Южно-Тамбейской скважины полуострова Ямал (верхняя юра, титон, Арктическая зона Западной Сибири). Время возникновения рода Siphonosphaera соотнесено с титоном (поздняя юра), приведены последовательности этапов появления новых представителей рода. Биономические характеристики Siphonosphaera позволяют сделать вывод о том, что акватория района Ямала представляла собой пелагиаль открытого океана, с нормальной соленостью и теплыми поверхностными водами.
В составе ямальской ассоциации радиолярий определены виды, входящие в зональный комплекс Parvicingula antoshkinae – P. blowi современной радиоляриевой шкалы юры Западной Сибири [4, 6, 7]. Зона Parvicingula antoshkinae – P. blowi датируется пределах нижнего – низов среднего подъяруса волжского региояруса, что соответствует нижнему титону Международной стратиграфической шкалы (145.0–149.24 Ma) [41].
Среди скелетов радиолярий ямальской ассоциации найдены мельчайшие сферические формы, одна из которых идентифицирована как представитель колониальных радиолярий отряда Collodaria Haeckel, 1881. Эта форма относится к роду Siphonosphaera Müller, 1858 из семейства Collosphaeridae Müller, 1858, и является новым видом Siphonosphaera yamalica Vishnevskaya et Amon, 2021 (рис. 1) [8, 9].
Рис. 1.
Местоположение комплекса радиолярий на полуострове Ямал и микрофотография на сканирующем электронном микроскопе Siphonosphaera yamalica Vishnevskaya et Amon; экз. ГИН № 170/1-TRI-90, баженовская свита, Южно-Тамбейская площадь, п-ов Ямал; нижний подъярус – низы среднего подъяруса волжского региояруса (нижний титон) [8]. Длина масштабной линейки 5 мкм. На обзорной контурной карте показаны места находок ископаемых сифоносфер на территории России: 1 – п-ов Ямал (титон); 2 – Восточная Камчатка (верхи среднего–низы верхнего миоцена); 3 – Курильская гряда (средний миоцен–плиоцен).
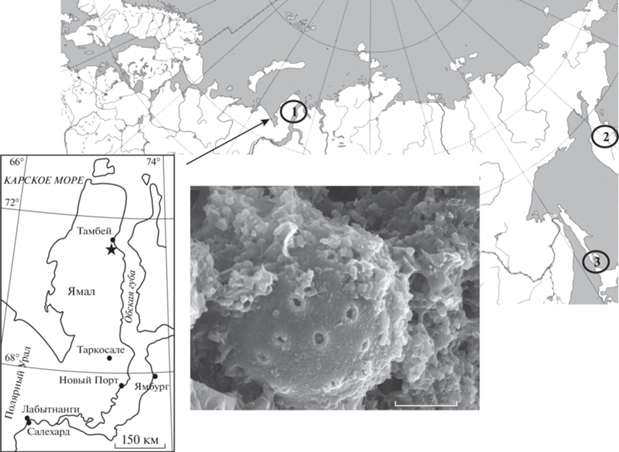
Данная находка ископаемых колониальных радиолярий рода Siphonosphaera является первой на территории Западной Сибири, и, в целом, в континентальной части России. Ранее значительно более молодые неогеновые представители рода Siphonosphaera были обнаружены только на Камчатке и в Дальневосточном регионе. Они установлены в комплексах радиолярий из пород верхов среднего – низов верхнего миоцена о-ва Карагинский [3] и в верхнем миоцене Кроноцкого района Восточной Камчатки [14]; а также в среднем–верхнем миоцене и в верхнем миоцене–плиоцене о-ва Кунашир Курильский гряды [15] (рис. 1).
Дополнительно заметим, что в ямальской ассоциации были также впервые встречены своеобразные радиолярии-сатурналиды из сем. Saturnalidae Deflandre 1953, которые ранее не отмечались ни в Западной Сибири, ни в Печорском бассейне, ни в Поволжье, ни в Московском бассейне [5].
Находка коллосферид на Ямале имеет немаловажное научное значение, поскольку, с одной стороны, проливает дополнительный свет на палеонтологическую историю данной группы радиолярий, способствуя уточнению ее филогении и эволюции, а с другой – позволяет делать некоторые палеогеографические заключения, основываясь на палеоэкологии группы. Радиолярии, являясь породообразующим элементом баженовской свиты, могут служить дополнительным, а иногда и ведущим инструментом в понимании условий ее формирования и реконструкции обстановок осадконакопления. Последнее имеет практическое значение в оценке перспектив нефтегазоносности исследуемой территории.
МАТЕРИАЛЫ И МЕТОДЫ
При изучении образцов пород из керна поисковой скважины 170 (координаты 71°30′ с.ш., 72°00′ в.д.), пробуренной на Южно-Тамбейской разведочной площади на полуострове Ямал (рис. 1), в кремнисто-карбонатных отложениях низов баженовской свиты в интервале глубин 3352.4–3355.90 м были выявлены скелеты радиолярий [6–8]. Кремнистые известняки были отобраны на радиоляриевый анализ в МГУ Ю.А. Гатовским и подвергнуты химической дезинтеграции c помощью уксусной и фтористоводородной кислот, благодаря чему удалось впервые выделить и получить представительный радиоляриевый комплекс [4–9]. Радиолярии в пробах многочисленные, сохранность их скелетов колеблется от удовлетворительной до хорошей и превосходной. Фотографирование радиолярий производилось на сканирующем электронном микроскопе VEGA2 TESCAN в Палеонтологическом институте им. А.А. Борисяка РАН (ПИН РАН) под руководством Е.А. Жегалло. Закрепление экземпляров радиолярий на металлических цилиндрических столиках (столбиках) осуществлялось с помощью заранее нанесенной двухсторонней углеродной ленты. Всего изучено 2 столика: один с извлеченными из породы скелетами радиолярий (157 экз.), другой с кусочками породы (13 шт.), вмещающей радиолярий, в том числе Siphonosphaera yamalica Vishnevskaya et Amon, 2021 в обр. TR-1-88/90 с глубины 3353.73 м.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Филогения сифоносфер. Колониальные радиолярии из отряда Collodaria изучались и изучаются уже более полутора веков, начиная с середины XIX-го столетия, и, в основном, на находках из планктона и поверхностного слоя осадков [17]. По поводу геологической истории колониальных радиолярий в литературе нет единого мнения, но во многих авторитетных источниках признается, что колониальные формы – это, в основном, современные (от голоцена до ныне) организмы [23 и др.]. Однако изредка их остатки находили в более древних, чем голоцен, отложениях.
Четвертичные коллодарии установлены в плейстоценовых осадках Пацифики и Карибского бассейна [47, 61, 68, 69 и др.].
Неогеновые колониальные формы описаны из нескольких регионов мира. Среднемиоценовые (серравальский ярус) сифоносферы Siphonosphaera brachysiphonia Dumitrică известны из формации “радиоляриевых сланцев” Румынии в предгорьях Южных Карпат [37, 38]. В донных осадках Тихого океана к западу от Маршалловых островов представители колониальных родов Siphonosphaera, Collosphaera, Polysolenia, Solenosphaera, Tribonosphaera встречены на нескольких интервалах от миоцена до плиоцена [68]. На востоке России миоценовые сифоносферы обнаружены на Камчатке, о. Карагинском [3, 14] и на Курилах [15]. В Южном полушарии коллодарии были найдены в миоценовых отложениях моря Уэдделла и на подводном плато Кергелен [20, 21, 50].
В олигоцене Центральной Пацифики установлена Siphonosphaera patinaria Haeckel [46], а в Атлантике в Карибском бассейне найдены плохой сохранности редкие коллосфериды олигоценового возраста [71]. Палеогеновые позднеэоценовые коллосфериды хорошей сохранности Acrosphaera sp. A были отмечены на подводном плато Кергелен в Южном полушарии [40]; Collosphaera poleogenica Vituchin описана из верхнего эоцена о-ва Карагинский, Восточная Камчатка [3].
Более древние коллосфериды, предположительно верхнего мела обнаружены на северо-западе Чешской Республики [62]. Наконец, самые древние находки предположительно колониальных радиолярий были сделаны в нижнетриасовых и пермских отложениях тропических районов океана Панталасса (Южный Китай и Япония) [45, 72].
В последние два десятилетия палеонтологи все чаще обращаются к результатам исследований в области молекулярной филогении (SSU rRNA Molecular Phylogeny) в попытках выяснить историю эволюционного развития радиолярий [25, 28, 48, 70, 74–76 и др.]. В частности установлено, что насселлярии являются сестринской группой (sister-group) по отношению к колониальным радиоляриям и образуют с ними единый клад [48, 74, 75]. Насселлярии и коллодарии дивергировали от основного ствола радиолярий в промежутке между юрой и эоценом, т.е. в этом интервале могли появиться пра-коллодарии [48, 53]. Настоящая колониальность появляется с олигоцена, точнее с границы эоцена/олигоцена, когда при переходе от эвтрофного океана эоцена к олиготрофному олигоцена, радиолярии выработали механизм колониальности в результате конкуренции с диатомеями за кремний dSi [51, 53].
Полагают, что коллодарии ответвились от насселлярий в среднем эоцене, причем первоначально это были одиночные формы [25, 44, 53]. Хронология событий, восстановленная на основе расчетов молекулярной дивергенции, указывает на то, что бесскелетные (naked) линии коллодарий впервые появились около 45.6 млн лет назад (эоцен), что совпало с диверсификацией диатомовых водорослей в пелагиали океанов. Колониальные коллодарии появились после формирования циркуляции Мирового океана, подобной современной, и развития олиготрофных условий в экваториальной части Тихого океана около 33.4 млн лет назад (рюпель, олигоцен) [22, 44]. Еще одним подтверждением этому являются результаты молекулярного анализа, подтвердившие, что ответвление Siphonosphaera внутри Collosphaeridae предшествовало разделению на Collosphaera и Acrosphaera [22].
Факт находки S. yamalica в нижневолжских (нижнетитонских) отложениях укладывается в названный интервал юры–эоцена. Скорее всего, эти первые сифоносферы были одиночными и еще не могли образовывать колонии. Данные физиологической и электронной микроскопии показали, что трубчатые выступы, которые присутствуют у представителей рода Siphonosphaera, могут быть результатом силицификации на краях пор оболочки [22], благодаря чему эта группа колониальных радиолярий смогла сохраниться в ископаемом состоянии. Мы предполагаем, что появление Siphonosphaera в верхней юре, было связано, по-видимому, с началом формирования современной системы океанов, в частности, с возникновением Пацифики [18].
Таким образом, не исключено, что коллодарии возникли в поздней юре, затем в нижнем и среднем мелу они не встречены или исчезли из геологической летописи и вновь появились, возможно, сначала в позднем мелу [62], во время становления Северной Атлантики, а затем палеогене (табл. 1). Их приход на арену эволюции в эоцене, олигоцене, миоцене, плиоцене и плейстоцене связан с активизацией формообразующих океаны процессов. В голоцене представлено уже все многообразие колониальных форм, но нельзя исключить, что в позднем мезозое и кайнозое происходили процессы скрытой эволюции, от которой не осталось материальных следов.
Таблица 1.
Состав рода Siphonosphaera
| Таксон | Литературный источник | Возраст |
|---|---|---|
| Siphonosphaera compacta (Brandt, 1905) sensu Strelkow et Reschetnjak, 1971 | [31], s. 339, taf. 9, fig. 10
. [17], с. 355–356, табл. 9 , фиг. 73 . |
Современный |
| Siphonosphaera crystalloides Hilmers, 1906 sensu Popofsky, 1917 | [64], s. 267, textfig. 31. | Современный |
| Siphonosphaera donghaiense Chen, 1987 | [34], p. 223, pl. 1, fig. 3 . | Современный |
| Siphonosphaeramacropora Strelkow et Reschetnjak, 1971 | [17], с. 357–358, рис. 29 . | Современный |
| Siphonosphaera magnisphaera Takahashi, 1982 | [59], p. 19, pl. 2, fig. 1. [79], p. 59, pl. 4, figs. 1, 3 . [61], pl. 10, figs. 5, 10 , pl. 11, figs. 7, 8 . [58], figs. 4 A, B . |
Современный |
| Siphonosphaera martensi Brandt, 1905 | [31], s. 339, taf. 9, fig. 13
. [17], с. 356, рис. 28 . [59], p. 19, pl. 43, fig. 3 . [79], p. 59, pl. 4, figs 4–5, 7–8 . [35], pl. 10, figs. 3, 4 . |
Современный |
| Siphonosphaera paradoxa Hilmers, 1906 sensu Popofsky, 1917 | [64], s. 266, textfig. 30. | Современный |
| Siphonosphaera pericyclis Chen, 1987 | [34], p. 222–223, pl. 1, figs. 1, 2. | Современный |
| Siphonosphaera polypora Chen, 1987 | [34], p. 222, pl. 1, figs. 5, 6 . | Современный |
| Siphonosphaera polysiphonia Haeckel, 1887 | [67], p. 89, pl. 1, fig. 7
. [79], p. 60. [58], figs. 4 C, D . |
Современный |
| Siphonosphaera schlotti Hilmers, 1906 sensu Popofsky, 1917 | [64], s. 267, textfig. 32. | Современный |
| Siphonosphaera socialis mazosphaeroides Strelkow et Reschetnjak, 1971 | [17], с. 354, рис. 27, B–D . | Современный |
| Siphonosphaera socialis tubililoba Strelkow et Reschetnjak, 1971 | [17], с. 355, рис. 27 , Е. | Современный |
| Siphonosphaera sp. A sensu Takahashi, 1991 | [79], p. 60, pl. 4, fig. 2. | Современный |
| Siphonosphaera sp. B sensu Takahashi, 1991 | [79], p. 60, pl. 4, fig. 6 . | Современный |
| Siphonosphaera sp. sensu Strelkow et Reschetnjak, 1971 | [17], с. 358, табл. 7 , фиг. 45 . | Современный |
| Siphonosphaera tenera Brandt, 1885 | [30], s. 266, taf. 7, fig. 48
. [31], s. 339, taf. 9, fig. 13 . [17], с. 356–357, рис. 67–71 . [63], p. 81. |
Современный |
| Siphonosphaera tizardi Fowler, 1898 | [39], p. 1032, pl. LXVI, fig. 1. | Современный |
| Siphonosphaera tubulosa (Müller) sensu Strelkow et Reschetnjak, 1971 | [17], с. 350–352, табл. 9
, фиг. 61–66
. [61], pl. 9, figs. 2, 5–7 . [35], pl. 10, figs. 5, 6 . |
Современный |
| Siphonosphaera? paraphoros Matsuzaki et Suzuki, 2014 | [55], p. 206, pl. 1, figs. 1–4 . | Плейстоцен средний– современный |
| Siphonosphaera hyalina Caulet, 1986 | [33], p. 849, pl. 2, fig. 1. | Поздний неоген–раннечетвертичный; ранний плиоцен–ранний плейстоцен |
| Siphonosphaera vesuvius Lazarus, 1992 | [50], p. 794–705, pl. 2, figs. 1–8 . | Неоген, поздний миоцен, тортонский ярус |
| Siphonosphaera cyathina Haeckel, 1887 | [42], p. 105, pl. 6, fig. 10
. [17], с. 352–353, рис. 26 . [15], с. 96. |
Средний миоцен–плиоцен–Современный |
| Siphonosphaera brachysiphonia Dumitrică, 1978 | [38], p. 35–37, figs. 1a–h. | Неоген, средний миоцен, серравальский ярус |
| Siphonosphaera minima Carnevale, 1908 | [32], p. 10, pl. 1, fig. 21 . | Неоген, средний миоцен, лангийский ярус |
| Siphonosphaera minima laevigata Principi, 1909 | [66], p. 21, pl. 1, fig. 3 . | Неоген, средний миоцен, лангийский ярус |
| Siphonosphaera preveriana Carnevale, 1908 | [32], p. 10, pl. 1, fig. 13 . | Неоген, средний миоцен, лангийский ярус |
| Siphonosphaera rariporata Carnevale, 1908 | [32], p. 10, pl. 1, fig. 17 . | Неоген, средний миоцен, лангийский ярус |
| Siphonosphaera squinaboli Carnevale, 1908 | [32], p. 10, pl. 1, fig. 14 . | Неоген, средний миоцен, лангийский ярус |
| Siphonosphaera socialis Haeckel, 1887 | [17], с. 353–354, табл. 8
, фиг. 60
,
табл. 9
, фиг. 72
, рис. 27
. [59], p. 19, pl. 2, fig. 2–4 , 8 . [61], pl. 10, figs. 1–6 , pl. 11, figs. 1–4 , 6–9 . [65], p. 5. [58], figs. 4 J–F . |
Неоген–современный, миоцен, голоцен |
| Siphonosphaeramembranella Reshetnyak et Runeva, 1978 | [14], с. 99–100, табл. 1, рис. 4 . | Средний – поздний миоцен |
| Siphonosphaera patinaria Haeckel, 1887 | [42], p. 105, pl. 6, figs. 7, 8
. [46], p. 9. |
Палеоген–современный. Олигоцен, неоген, плейстоцен, аквитанский, хаттский ярусы |
| Siphonosphaera yamalica Vishnevskaya et Amon, 2021 | [8], с. 6, табл. I , фиг. 1, 1а. | Поздняя юра, титонский ярус |
В составе рода Siphonosphaera насчитывается, по нашему мнению, 33 таксона; эти систематические единицы разного ранга – видовые, подвидовые, не определенные до вида и приведенные в открытой номенклатуре – зафиксированы в литературных источниках издания XIX–XXI веков (табл. 1). Валидность таксонов подтверждена по [29, 52, 75, 76]. Также следует согласиться с результатами таксономической ревизии рода Siphonosphaera, приведенной в [60] где устранена часть синонимов, описанных ранее в составе рода, например, в статьях [12, 43]. В частности, вид Siphonosphaera monotubulosa Hilmers, 1906 следует рассматривать в составе рода Choenicosphaera; аналогично, ряд вновь описанных, а также известных видов, отнесенных к роду Siphonosphaera в трактовке [12], реально принадлежит родам Collosphaera, Disolenia, Trisolenia, Polysolenia. Но в целом, как это не раз подчеркивалось в литературе [29 и др.], необходима полноценная ревизия систематики сифоносфер, которая, несомненно, будет выполнена специалистами в будущем.
Большинство перечисленных в табл. 1 таксонов является современными формами, а 14 зафиксированы в доголоценовых отложениях, что составляет более трети от общего числа – 42.4%.
В порядке от более молодых к более древним ископаемые виды представлены:
Siphonosphaera? paraphoros Matsuzaki et Suzuki, 2014 (рис. 2, 6). Средний плейстоцен – современный, интервал существования ~0–750 тыс. лет назад. Регион обитания Северное полушарие, Северная Пацифика у берегов Японии, широтная полоса вдоль ~41° с.ш. [55].
Рис. 2.
Этапы развития (I–III) сифоносфер. Позднеюрский этап: (1) – Siphonosphaera yamalica, поздняя юра, нижневолжская эпоха, Палеоарктика, п-ов Ямал ([8], табл. I, фиг. 1, 1). Неоген-палеогеновый этап: (2) – Siphonosphaera patinaria, поздний олигоцен – ранний неоген, Центральная Пацифика ([42], pl. 6, fig. 8 ); (3) – Siphonosphaera brachysiphonia, средний миоцен, Неотетис, Южные Карпаты ([38], fig. 1a); (4) – Siphonosphaera vesuvius, поздний миоцен, южная часть Индийского океана, подводное плато Кергелен ([50], pl. 2, fig. 1); (5) – Siphonosphaera hyalina, ранний плиоцен–ранний плейстоцен, Юго-Западная Пацифика, подводное плато Фиджи ([33], pl. 2, fig. 1). Современный этап: (6) – Siphonosphaera? paraphoros, средний плейстоцен – современный, Северная Пацифика у северо-восточных берегов Японии ([55], pl. 1, fig. 1); (7) – Siphonosphaera martensi, современный, Центральная тропическая Пацифика ([79], pl. 4, fig. 8 ); (8, 9) – Siphonosphaera socialis, современный, Северо-Западная Пацифика ([59], pl. 2, fig. 8 ); (10) – Siphonosphaera magnisphaera, современный, Северо-Западная Пацифика ([59], pl. 2, fig. 1).

Siphonosphaera hyalina Caulet, 1986 (рис. 2, 5). Поздний неоген – раннечетвертичный, ранний плиоцен – ранний плейстоцен, занклская – гелазская эпохи, интервал существования ~5.3–1.8 млн лет назад. В неогене регион обитания Южное полушарие, ЮЗ Пацифика, западная окраина подводного плато Фиджи к востоку от северной Австралии, полоса широт ~16°–20° ю.ш. [33]. В раннем плейстоцене S. aff. hyalina отмечен в Северном полушарии, Северная Пацифика, широтная полоса вдоль ~38° с.ш. [56].
Siphonosphaera vesuvius Lazarus, 1992 (рис. 2, 4). Неоген, поздний миоцен, тортонская эпоха, интервал существования ~11.6–7.2 млн лет назад. Регион обитания Южное полушарие, южная часть Индийского океана, подводное плато Кергелен, полоса широт ~55°–58° ю.ш. [50].
Siphonosphaera cyathina Haeckel, 1887. Миоцен – современный. В качестве современного вида характеризуется чертами космополитизма с тяготением к тропической зоне Мирового океана [17]). Неогеновый представитель вида найден в верхнем миоцене – плиоцене (11.6–2.6 млн лет назад) о-ва Кунашир Большой Курильской гряды [14] в полосе широт вдоль ~44° с.ш.
Siphonosphaera brachysiphonia Dumitrică, 1978 (рис. 2, 3). Неоген, средний миоцен, раннесерравальская эпоха, интервал существования ~13.8–12.8 млн лет назад. Регион обитания Северное полушарие, Неотетис, Центральная Румыния, Южные Карпаты, полоса широт ~44°–45° с.ш. [37, 38].
Группа видов Siphonosphaera rariporata Carnevale, 1908, S. minima Carnevale, 1908, S. squinaboli Carnevale, 1908, S. preveriana Carnevale, 1908 и S. minima laevigata Principi, 1909. Неоген, средний миоцен, лангийская эпоха, интервал существования ~15.9–13.8 млн лет назад. Регион обитания Северное полушарие, Неотетис, Италия, широта ~42° с.ш. [32, 66].
Siphonosphaera membranella Reshetnyak et Runeva, 1978. Неоген, средний миоцен, интервал существования ~15.9–11.6 млн лет назад. Регион обитания Северное полушарие, Восточная Камчатка, полоса широт ~54°–59° с.ш. [3, 14].
Siphonosphaera socialis Haeckel, 1887. Неоген – современный, миоцен и голоцен. В качестве современного вида характеризуется чертами космополитизма с тяготением к тропической зоне Мирового океана [17], отмечен, в частности, в восточной части Индийского океана [58, 61] и в море Банда на западе Тихого океана [61]), полоса широт от 8° ю.ш. до 10° с.ш. Голоценовый S. socialis отмечен в Северной Пацифике в полосе широт ~38°–40° с.ш. [56]. Неогеновый (миоценовый без подразделения на ярусы, ~23.0 млн лет назад) представитель этого вида был обнаружен в донных осадках к северу от побережья Гавайев в центральной части Северной Пацифики, полоса широт ~44°–45° с.ш. [65].
Siphonosphaera patinaria Haeckel, 1887 (рис. 2, 2). Палеоген–неоген, поздний олигоцен–ранний неоген, хаттская–аквитанская эпохи, интервал существования ~26–21 млн лет назад. Регион обитания Северное полушарие, Центральная Пацифика, полоса широт ~7°–8° с.ш. [46].
Siphonosphaera yamalica Vishnevskaya et Amon, 2021 (рис. 2, 1). Поздняя юра, нижневолжская (нижний титон) эпоха, интервал существования ~149–147 млн лет назад. Регион обитания Северное полушарие, Палеоарктика, п-ов Ямал, полоса широт ~70°–72° с.ш. [8, 9].
График распределения таксонов Siphonosphaera во времени наглядно демонстрирует, что в развитии рода имелись три неравновеликих этапа: позднеюрский (рис. 2, I), неоген-палеогеновый (рис. 2, II) и современный (рис. 2, III). Современный этап характеризуется максимальным числом видов сифоносфер, а два более древних – менее обильны и представлены единичными, редко, небольшими группами видов; возможно, это связано с тем, что находки в древних отложениях просто могли не сохраняться.
При этом неоген – палеогеновые (олигоценовые) виды возникали по отдельности с относительно небольшими по длительности перерывами (промежутками) от ~2 до ~6 млн лет, и от современной палеогеновую эпоху отделяет около 25 млн лет. Максимум обилия неогеновых форм приходился на миоцен, когда они могли появляться группами. Для позднеюрского этапа известна лишь наша единственная находка, указывающая на изолированное внезапное импульсное возникновение и исчезновение одной формы.
Анализируя сравнительную морфологию и преемственность форм можно проследить линию последовательной трансформации видов: позднеюрский Siphonosphaera yamalica (рис. 2, 1) → → средненеогеновый S. brachysiphonia (рис. 2, 3) → → современный S. martensi (рис. 2, 7). Их объединяет сходная конструкция скелета: небольшое число трубочек, их невеликие диаметр и длина, а также среднее число регулярных небольших пор. От других современных и неогеновых видов этого рода отличия конструкции более существенные, например, у современной Siphonosphaera cyathina Haeckel присутствует большое число трубочек (до 9–10 на половину диаметра), а у S. hyalina Caulet развиты всего две длинные трубки, расположенные полярно.
Значение находки Siphonosphaera для палеогеографии. Основные черты биологии колониальных коллосферид, к которым принадлежит рассматриваемый вид Siphonosphaera yamalica, позволяют сделать некоторые вероятные заключения о палеобстановках водного бассейна, располагавшегося на месте находки данного вида на Ямале на территории северо-западной арктической зоны Западной Сибири.
Напомним, что, в целом, коллодарии обладают уникальным набором своеобразных морфологических и экологических свойств, отличающих их от остальных Radiolaria: в этом отряде объединены формы, ведущие колониальный образ жизни, как с минеральным кремнистым скелетом, так и лишенные его. Молекулярно-филогенетический анализ показал, что среди коллодарий выделяются три монофилетических систер-группы: а) скелетные Collosphaeridae; б) лишенные скелета, но имеющие спикулы Sphaerozoidae и в) бесскелетные Collophidiidae [25, 53, 74, 75].
Колонии представляют собой собрания одиночных особей-индивидов (капсул) какого-либо вида, соединенных ризоподиальной сетью и погруженных в общую желатиновую (студенистую) вакуолистую массу (матрикс), образующую оболочку. Колонии могут иметь сферическую, сфероидальную, овальную или цилиндроидную тубообразную форму и достигать размеров от нескольких миллиметров до одного и более метра (до 3–5 м) [17, 23, 24, 28, 77, 78]. После отмирания колонии распадаются на погибшие индивиды и опускаются на дно; индивиды растворяются морской водой во время этого транспорта, а также иловыми водами в тафоценозе, а часть скелетных остатков сохраняется в осадке и фоссилизируется в ориктоценоз.
Современные колонии радиолярий обитают в поверхностных водах океанов, тяготея к низкоширотной тропической зоне центральных зон океанов. Они плавают (парят) в воде благодаря особым свойствам студенистой массы и ее вакуолям, обитая, как правило, в самых поверхностных слоях (0–10 м), и изредка встречаясь на глубине 30–50 м [17]. В состоянии стресса число и размеры вакуолей сокращаются, вплоть до их исчезновения, индивиды собираются в комки, в результате чего колония тонет, опускается вниз. Стрессовые состояния вызываются резкими колебаниями (скачками) температуры, чередованием потеплений и похолоданий, ветром и сильным волнением, выпадением осадков и др. В таких случаях колонии опускаются до глубин 50–200 м и вновь поднимаются к поверхности с наступлением благоприятных условий [19].
Пищу для индивидов в колониях составляют диатомеи, перидинеи, инфузории, мелкие рачки, личинки Decapoda и Echinodermata, которые приклеиваются к поверхности колонии, затем втягиваются внутрь и в них проникают ризоподии индивидов [17]. Сами колонии являются пищей для копепод, амфипод, сальп и некоторых личинок беспозвоночных (например, моллюсков, омаров) [28, 53, 75].
Колониальные радиолярии – это типичные тепловодные обитатели открытого океана, они преимущественно встречаются в тропических и субтропических олиготрофных зонах центральных круговоротов открытых океанов, и, в меньшей степени, в прибрежных акваториях [77], в частности, в Аденском заливе [19], Филиппинском и Восточно-Китайском морях [54]. Часто они встречаются массово, и в отдельных случаях их общее количество просто колоссально: численность может достигать 16–20 тыс. колоний в 1 м3 [19]. Космополитные коллодарии Siphonosphaera polysiphonia и Sphaerozoum punctatum, являющиеся видами-доминантами в экваториальных водах восточной части Индийского океана в интервале глубин 0–200 м, имели обилие 661 инд./м3 и 5500 инд./м3 соответственно [57, 58]; а в фотическом поверхностном слое в низкоширотной зоне Центральной Пацифики плотность их популяций колебалась в пределах от 30 до 20 000 колоний в 1 м3 [36].
В геологическом прошлом некоторые сифоносферы обитали в средних и более высоких широтах, но также в тепловодных обстановках. Находка Siphonosphaera cyathina в осадках о. Кунашир [15] позволила сделать вывод о наличие поверхностных тепловодных масс в бассейне миоцен-плиоценового времени, находившегося в районе Большой Курильской гряды. Аналогично, обитание S. membranella в среднем миоцене в акватории еще более северного района Восточной Камчатки [3, 14], а присутствие S. vesuvius в позднем миоцене акватории района подводного плато Кергелен в Южном полушарии [50] связывалось с теплыми водами.
Со временем, один из представителей рода Siphonosphaera – Siphonosphaera cyathina – приспособился к существованию в весьма суровых условиях. Так, группа колониальных радиолярий была установлена в донных осадках и в живом виде в районах высоких и экстремально высоких широт в Центральном Арктическом бассейне и в окраинном Норвежском море [11, 26, 27]. В Центральной Арктике в полосе широт 80°–82° с.ш. севернее Шпицбергена обнаружено планктонными ловами значительное число видов живых радиолярий-полицистин (145 видов), среди которых найдены колониальные: Acrosphaera flammabunda Popofsky, *Collosphaera huxleyi Haeckel, *Collosphaera macropora Popofsky, *Collosphaera tuberosa Haeckel, Otosphaera auriculata Haeckel, Siphonosphaera cyathina Haeckel, *Sphaerozoum punctatum Müller, *Collosphaera sp. (звездочкой отмечены виды, живые особи которых несли в себе водорослевые симбионты) [27]. Эти коллодарии были обнаружены ловами в слое 0–120 м [27].
Многообразие видов радиолярий-полицистин в высокой Арктике представлено преимущественно теплолюбивыми тропическими и субтропическими формами (98 из 145 выявленных видов). Заметим при этом, что температура воды в Арктике на широте 80° с.ш. на глубине 100 м колеблется в пределах от –2 до +4°С. Проникновение в Арктику и существование подобной “экзотичной” радиоляриевой фауны в мало благоприятных условиях объясняется вторжением (incursions) теплых водных масс из тропической и субтропической Атлантики в течение голоцена [11, 26, 27]. Видам была свойственна высокая морфологическая пластичность, что, в сочетании с симбиозом с микроводорослями, позволило адаптироваться к суровым условиям.
А в холодноводной Субарктической Пацифике в современных и четвертичных препаратах постоянно (единичными или несколькими экземплярами) встречаются Polysolenia arktios Nigrini и Polysolenia pseudoarktios Caulet [56].
Можно высказать предположение, что обитание колониальных радиолярий в районе Ямала в поздней юре также, вероятно, могло быть вызвано инвазией теплого течения из Тетической области.
Для коллодарий характерно присутствие фотосинтезирующих эндосимбионтов-микроводорослей с хроматофорами желтого цвета, которых иногда называют “зооксантеллами” [17], и которые либо рассеяны в студенистой массе, либо окружают индивиды плотным слоем. В колониях Collosphaeridae и Collozoidae может находиться 26 × 106 клеток эндосимбионтов [23]. По материалам исследований планктона Аденского залива [19], содержание хлорофилла-а в водорослевых симбионтах колониальных радиолярий почти в три раза больше, чем в свободно живущем фитопланктоне. В низкоширотной зоне Центральной Пацифики в слое воды верхних 150 м первичная продукция, производимая симбионтами-фотосинтетиками колониальных радиолярий, колебалась в пределах от 6.5 до 40.7 мг С/м2 (в среднем 23.6 мг С/м2) в день [36].
Колониальные радиолярии предоставляют симбионтам благоприятные условия существования, и подобное свойство колоний получило специальное название – приобретенная фототрофия (acquired phototrophy). Она встречается у радиолярий, фораминифер, инфузорий и динофлагеллят, но наиболее экологически значима среди первых трех. Приобретенная фототрофия фораминифер и радиолярий имеет решающее значение для биогеохимических циклов (углерод, стронций, карбонаты, силикаты) в субтропических и тропических зонах океанов [73].
Среди эндосимбионтов коллодарий часто упоминаются динофлагелляты, относящиеся к роду Scrippsiella [77]; симбиоз проявляется в том, что динофиты обретают защиту в колонии, а сами они служат источником питания для индивидов колонии. Колониальные коллодарии развили специфические приспособления, которые позволяют им выживать и процветать в олиготрофной среде именно благодаря эндосимбионтам.
Эти биономические характеристики позволяют сделать некоторые палеогеографические выводы и заключения, основываясь на принципе актуализма.
Прежде всего следует сказать, что район обитания S. yamalica в баженовском бассейне по-видимому относился к пелагической зоне открытого океана или находился на внешней периферии крупного прибрежно-океанического бассейна. Это следует из характеристик биогеографии современных коллосферид [17, 53].
В связи с тем, что коллодарии были встречены только в открытых зонах океанов, и никогда во внутренних морях, то важным условием их существования является нормальная океаническая соленость, следовательно, таковая была характерна и для рассматриваемого района. Это совпадает с оценкой солености вод баженовского моря, данной по другим источникам: по абсолютному показателю она была близка к современной океанической – в прибереговых акваториях 31.5‰, в открытой пелагиали составляла 34.0‰ [10]. Заметим попутно, что для Siphonosphaera polysiphonia, признанного вида-доминанта в акваториях востока Индийского океана, установлена значительная положительная корреляция к солености и температуре верхнего водного слоя (0–150 м), иными словами, снижение этих параметров вело к снижению плотности популяции [53].
О глубинах этой акватории обитания, опираясь на биномические характеристики S. yamalica, сказать затруднительно – это, в равной степени, могли быть как сравнительно значительные глубины в несколько сот метров [2, 10], так и относительное мелководье/холмистое подводное плато [16]. В связи с тем, что колониальные формы в своем повседневном образе жизни тяготеют к самому верхнему горизонту водной толщи (поверхностные воды, 0–10 м), то они не могут служить строгим индикатором истинной глубины палеобассейна, которая могла измеряться десятками, сотнями и тысячами метров.
Коллодарии в периоды резких скачков и перепадов температуры, а также во время волнений и штормов, опускаются до глубин 30–50 м и глубже до 200 м, за пределы базиса волн, но это мало что говорит о глубине района обитания на Ямале. Возможно, что такие глубины 30–200 м были здесь представлены, возможно, были и более глубокие.
Современные коллодарии в своем географическом распространении тяготеют к тепловодным условиям низких широт. В частности, температура воды в Аденском заливе, в котором было встречено колоссальное количество колониальных радиолярий, варьируется от +15° до +28°C в зависимости от муссонных ветров и сезона [19]. Следовательно, находка S. yamalica позволяет предположить, что условия обитания этой формы на северо-западе Западносибирского баженовского морского бассейна были столь же теплыми (или близко к тому). Это не противоречит оценкам температуры вод, предложенным по другим данным — среднегодовая температура вод в эпипелагиали в раннебаженовское (средневолжское) время вблизи северо-западного берега составляла +15–18°С, вблизи северо-восточного +13–14°С [10].
Выше говорилось о том, что динофитовые играют значительную роль в биологии коллодарий, выступая как эндосимбионты и объекты питания. В палеонтологической летописи ископаемые остатки динофлагеллят известны со среднего триаса, а в баженовском горизонте найдены как органикостенные, так и известковистые цисты динофлагеллят [13, 80]. Этот фактор – наличие динофитовых в планктоне пелагиали баженовского бассейна – возможно сыграл важную и существенную роль в появлении колониальных радиолярий в поздней юре Западной Сибири.
Колониальные коллодарии, населяя воды океанов, играли и играют громадную роль в первичной продукции углерода. Они развили специфические приспособления, которые позволяют им выживать и процветать благодаря микроводорослевым эндосимбионтам. Производимая колониями органика в виде желатинового матрикса, компонентов клеток индивидов колоний, а также эндосимбионтов и паразитов могла служить источником органического вещества в нафтидогенезе и генерации нефти в Западной Сибири. С некромассой радиолярий, включая сюда колониальных, на дно бассейна поступало значительное количество органического вещества. Следовательно, радиолярий можно рассматривать как группу планктонных микрорганизмов, способную при определенных условиях служить важным источником органического вещества, которое могло участвовать в образовании нефти и газа [1, 49].
ЗАКЛЮЧЕНИЕ
Первая находка представителя колониальных радиолярий Siphonosphaera yamalica из семейства Collosphaeridae в верхнеюрских (титонских) отложениях на п-ове Ямал имеет важное научное теоретическое и практическое значение.
Эта находка S. yamalica в верхнеюрских отложениях северо-запада Западной Сибири расширяет знание палеонтологии и географии распространения ископаемых колониальных форм. До настоящего времени сведения о колониальных радиоляриях ограничивались в основном данными о современных живых, пойманных ловами, или погибших индивидах, собранных в донных осадках. Очень редки находки ископаемых форм в кайнозойских отложениях Пацифики, Атлантики и Индийского океана – в эоцене, плиоцене и плейстоцене. Единичны данные о более древних, докайнозойских, предположительно колониальных радиоляриях, которые были найденных в верхней перми (лопингий) Южного Китая. Время возникновения рода Siphonosphaera соотнесено с поздневолжским временем (титон, поздняя юра).
Таким образом, впервые колониальные радиолярии установлены в юре (около 147–149 млн лет – начало волжского века, Ямал, Арктическая Сибирь). Ранее самыми древними считались позднеэоценовые – около 26 млн лет назад. Следовательно, становление рода, вероятно, произошло более чем на 120 млн лет раньше, чем предполагалось.
Кроме того, находка S. yamalica вносит немалый новый вклад в познание особенностей эволюционного развития данной группы радиолярий. Теперь можно с большей уверенностью говорить о докайнозойском юрском времени возникновения Collodaria, как это предполагалось согласно данным молекулярной филогении.
Биономические характеристики коллодарий в целом, и рода Siphonosphaera в частности, позволяют сделать вывод о том, что акватория района Ямала в ранневолжское время представляла собой пелагиаль открытого океана, с нормальной соленостью и тепловодными (от +15°С и выше) условиями.
Колониальные радиолярии могли при определенных условиях служить важным источником органического вещества в нафтидогенезе.
Благодарности. Авторы выражают признательность А.С. Алексееву (МГУ) и Д.Н. Засько (ИО РАН, Россия) за ценные советы и замечания, Г.А. Калмыкову (МГУ) за помощь в подборе каменных материалов, Е.А. Жегалло (ПИН РАН) за работу на сканирующем электронном микроскопе, искреннюю благодарность рецензенту А.Г. Матулю за критический разбор и конструктивное обсуждение статьи. Работа выполнена по гостемам ГИН РАН и ПИН РАН, а также частичной поддержке Программы Президиума РАН “Эволюция органического мира и планетарных процессов”.
Список литературы
Амон Э.О. Радиолярии в экосистеме баженовского моря (поздняя юра–начало раннего мела, Западная Сибирь) // Литология и геология горючих ископаемых. Вып. V (21). Екатеринбург: Изд-во УГГУ, 2011. С. 145–169.
Бочкарев В.С., Огнев Д.А., Черданцев С.Г. Палеобатиметрия эпиконтинентальных морей на примере мела Западной Сибири // Меловая система России и ближнего зарубежья: проблемы стратиграфии и палеогеографии / Ред. Барабошкин Е.Ю. и др. Новосибирск: Изд-во СО РАН, 2008. С. 44–46.
Витухин Д.И. Расчленение кайнозоя Дальнего Востока России по радиоляриям. М.: Наука, 1993. 105 с.
Вишневская В.С., Гатовский Ю.А. Первая находка позднеюрских радиолярий на Ямале (Арктическая Сибирь) // Проблемы региональной геологии Северной Евразии. М.: МГРИ-РГГРУ, 2020. С. 17–20.
Вишневская В.С., Гатовский Ю.А. Позднеюрские Saturnalidae (Radiolaria) Ямала (Арктическая Сибирь) // Юрская система России: проблемы стратиграфии и палеогеографии / Ред. Захаров В.А. и др. Сыктывкар: ИГ Коми НЦ УрО РАН, 2020. С. 46–48.
Вишневская В.С., Гатовский Ю.А. Верхнеюрские радиолярии Арктической периферии Западной Сибири (п-ов Ямал) // Теоретические и прикладные аспекты палеонтологии / Ред. Розанов А.Ю. и др. СПб.: ВСЕГЕИ, 2021. С. 17–18.
Вишневская В.С., Амон Э.О., Гатовский Ю.А. Радиоляриевая биостратиграфия баженовского горизонта (верхняя юра–нижний мел) Западной Сибири // Стратиграфия. Геол. корреляция, 2020. Т. 47. № 8. С. 1–20. https://doi.org/10.31857/S0869592X20060101
Вишневская В.С., Амон Э.О., Гатовский Ю.А., Жегалло Е.А. Первая находка колониальных радиолярий в юрских отложениях (Арктическая зона Сибири) // Палеонтол. журн. 2021 . № 4. С. 3–9. https://doi.org/10.31857/S0031031X21040152
Вишневская В.С., Амон Э.О., Гатовский Ю.А., Жегалло Е.А. Колониальные радиолярии Siphonosphaera в юрских отложениях севера Западной Сибири // ПАЛЕОСТРАТ-2021: годичное собрание (научная конференция) секции палеонтологии МОИП и Московского отделения Палеонтологического общества при РАН. Москва, 25–26 января 2021 г. Тезисы докладов. М.: ПИН РАН, 2021. С. 16.
Захаров В.А. Условия формирования волжско-берриасской высокоуглеродистой баженовской свиты Западной Сибири по данным палеоэкологии // Эволюция биосферы и биоразнообразия / Ред. Рожнов С.В. М.: Т-во научных изданий КМК, 2006. С. 552–568.
Кругликова С.Б. Радиолярии Polycystina из донных отложений Мирового океана как биоиндикаторы изменений окружающей среды. М.: ГЕОС, 2013. 231 с.
Меншуткин В.В., Петрушевская М.Г. Классификация радиолярий семейства Collosphaeridae фенетическими методами // Исследования фауны морей. 1989. Т. 41(49). С. 61–99.
Панченко И.В., Балушкина Н.С., Барабошкин Е.Ю. и др. Комплексы палеобиоты в абалакско-баженовских отложениях центральной части Западной Сибири // Нефтегазовая геол. Теория и практика. 2015. Т. 10. № 2. С. 1–29. http://www.ngtp.ru/rub/2/24_2015.pdf.
Решетняк В.В., Рунева Н.П. Колониальные радиолярии сем. Collosphaeridae позднемиоценовых отложений Камчатки // Фауна и систематика одноклеточных животных / Ред. Крылов М.В. Л.: ЗИН АН СССР, 1978. С. 96–100.
Рунева Н.П., Ушко К.А. Неогеновые радиолярии Большой Курильской гряды и их стратиграфическое значение // Микрофауна нефтегазоносных районов СССР / Ред. Любимова П.С., Мятлюк Е.В. Л.: ВНИГРИ, 1984. С. 94–101.
Стафеев А.Н., Ступакова А.В., Суслова А.А. и др. Баженовский горизонт Сибири (титон–нижний берриас): тектонические и гидродинамические условия осадконакопления // Георесурсы. 2019. Т. 21. № 2. С. 117–128. https://doi.org/10.18599/grs.2019
Стрелков А.А., Решетняк В.В. Колониальные радиолярии Spumellaria Мирового океана // Исслед. фауны морей. 1971. № 9(17). С. 295–373.
Фролов В.Т., Фролова Т.И. Происхождение Тихого океана. 2-е изд., доп. М.: МАКС Пресс,2011. 52 с.
Хмелева Н.Н. Роль радиолярий при оценке первичной продукции в Красном море и Аденском заливе // Докл. АНСССР. 1967. Т. 173. № 6. С. 1430–1433.
Abelmann A. Oligocene to Middle Miocene radiolarian stratigraphy of southern high latitudes from Leg 113, Sites 689 and 690, Maud Rise // Proc. ODP. Sci. Results. V. 113 / Eds. Barker F.P. et al. College Station, TX, 1990. P. 676–708.
Abelmann A. Early to Middle Miocene radiolarian stratigraphy of the Kerguelen Plateau, Leg 120 // Proc. ODP. Sci. Results. V. 120 / Eds. Wise S.W. et al. College Station, TX, 1992. P. 757–784.
Amaral Zettler L., Anderson O.R., Caron D.A. Towards a molecular phylogeny of colonial spumellarian Radiolaria // Mar. Micropaleontol. 1999. V. 36. P. 67–79.
Anderson O.R. Radiolaria. New York: Springer, 1983. 355 p.
Anderson O.R., Nigrini C., Boltovskoy D. et al. Class Polycystina // The Second Illustrated Guide to the Protozoa / Eds. Lee J.J. et al. Society of Protozoologists, Lawrenec, KS. 2002. P. 994–1022.
Biard T., Pill L., Decelle J. et al. Toward an integrative morpho-molecular classification of the Collodaria (Polycystinea, Radiolaria) // Protist. 2015. V. 166. Iss. 3. P. 374–388. https://doi.org/10.1016/j.protis.2015.05.002
Bjørklund K.R., Kruglikova S.B. Polycystine radiolarians in surface sediments in the Arctic Ocean basins and marginal seas // Mar. Micropaleontol. 2003. V. 49. P. 231–273.
Bjørklund K.R., Kruglikova S.B., Anderson O.R. Modern incursions of tropical Radiolaria into the Arctic Ocean // J. Micropaleontol. 2012. V. 31. P. 139–158.
Boltovskoy D., Anderson O.R., Correa N.M. Radiolaria and Phaeodaria // Handbook of the Protists / Eds. Archibald J. et al. Springer, Cham. 2017. P. 731–763.
Boltovskoy D., Kling S.A., Takahashi K., Bjørklund K. World Atlas of Distribution of Recent Polycystina (Radiolaria) // Palaeontologia Electronica. 2010. V. 13. Issue 3; 18A. 230 p. http://palaeo-electronica.org/ 2010_3/215/index.html.
Brandt K. Die koloniebilden Radiolarien (Sphaerozoeen) des Golfes von Neapel // Fauna und Flora des Golfes von Neapel. 1885. V. 13. S. 1–276.
Brandt K. Zur Systematik der koloniebildenden Radiolarien // Zool. Jahrb. Suppl. 1905. V. 8. S. 31–352.
Carnevale P. Radiolaire e silicoflagellati di Bergonzano (Reggio Emilia) // Memorie del Reale Instituto Veneto di Scienze, Lettereed Arti. 1908. V. 28(3). P. 1–46.
Caulet J.-P. Radiolarians from the Southwest Pacific // Init. Reports DSDP. V. 90 / Eds. Kennett J.P. et al. Washington, D.C., 1986. P. 835–861.
Chen W. Some new species of Radiolaria from surface sediments of the East China sea and the South China sea // Chin. J. Oceanol. Limnol. 1987. V. 5. № 3. P. 222–228.
Chen M., Zhang Q., Zhang L., Liu L. Radiolaria in the sediments from the northwest Pacific and its marginal seas. Beijing: Science Press, 2017. 268 p.
Dennett M.R., Caron D.A., Michaels A.F. et al. Video plankton recorder reveals high abundances of colonial Radiolaria in surface waters of the central North Pacific // J. Plankton Res. 2002. V. 24. № 8. P. 797–805.
Dumitrică P. Badenian Radiolaria from Central Paratethys // Chronostratigraphie und Neostratotypen. Miozän der Zentralen Paratethys. Bd. 6, Miozän M4 Badenien / Ed. Brestenská E. Bratislava: VEDA, 1978. P. 231–261.
Dumitrică P. On the status of the collosphaerid radiolarian species Siphonosphaera brachysiphonia Dumitrică, 1978 and Siphonosphaera arkys Su, 1982 // Acta Palaeontol. Romaniae. 2019. V. 15. № 1. P. 35–38.
Fowler G.H. Contributions to our knowledge of the plankton of the Faroe Channel. No. VII. A. General data of the stations. B. The Protozoa. C. The Medusae // Proceedings of the General Meetings for Scientific Business of the Zoological Society of London. 1898. P. 1016–1032.
Goll R.M., Caulet J.P. Sur la présence d’un Collosphaeridae dans les sédiments Eocene supérieur du Plateau de Kerguelen-Heard // Comptes rendus de l’Académie des Sciences (Paris). 1985. Série II 301. P. 1375–1380.
Gradstein F.M., Ogg J.G., Schmitz M.D., Ogg G.M. Geologic Time Scale 2020. V. 2. Elsevier BV. 2020. 1357 p.
Haeckel E. Report on the Radiolaria collected by H.M.S. Challenger during the years 1873–1876 // Rep. Sci. Results Voy. H.M.S. Challenger, 1873–1876. Zool. 1887. V. 18. P. 1–1803.
Hilmers C. Zur Kenntnis der Collosphaeriden. Inaugural-Dissertation zur Erlandung der Dorkorwürde der hohen philosophischen Fakultät der Königlichen Christian-Albrecht-Universität in Kiel. Druck von C. Schaidt, Kiel, 1906. P. 1– 93.
Ishitani Y., Ujiié Y., de Vargas C. et al. Phylogenetic Relationships and Evolutionary Patterns of the Order Collodaria (Radiolaria) // PLoS ONE. 2012. V. 7(5): e35775. https://doi.org/10.1371/journal.pone.0035775
Ito T., Zhang L., Zhang M.-H. et al. Guiuva sashidai n. gen. n. sp., a probable colonial Radiolaria from the Lopingian (Upper Permian) in South China // Palaeoworld. 2017. V. 26. P. 631–637. https://doi.org/10.1016/j.palwor.2017.04.001
Kamikuri S., Nishi H., Moore T.C. et al. Data Report: Late Oligocene and Early Miocene radiolarians, Sites 1218 and 1219, Central Pacific // Proc. ODP. Sci. Results. V. 199 / Eds. Wilson P.A. et al. College Station, TX, 2005. P. 1–13. https://doi.org/10.2973/odp.proc.sr.199.212.2005.
Knoll A.H., Johnson D.A. Late Pleistocene evolution of the collosphaerid radiolarian Buccinosphaera invaginata Haeckel // Micropaleontol. 1975. V. 21. P. 60–68. https://doi.org/10.2307/1485155
Kunitomo Y., Sarashina I., Iijima M. et al. Molecular phylogeny of acantharian and polycystine radiolarians based on ribosomal DNA sequences, and some comparisons with data from the fossil record // Europ. Journ. Protistology. 2006. V. 42. P. 143–153. https://doi.org/10.1016/j.ejop.2006.04.001
Lampitt R. S., Salter,I., Johns D. Radiolaria: Major exporters of organic carbon to the deep ocean // Global Biogeochemical Cycles. 2009. V. 23, GB1010. P. 1–9. https://doi.org/10.1029/2008GB003221
Lazarus D. Antarctic Neogene radiolarians from the Kerguelen Plateau, Legs 119 and 120 // Proc. ODP. Sci. Results. V. 120 / Eds. Wise S.W. et al. College Station, TX, 1992. P. 785–809.
Lazarus D., Kotrc B., Wulf G., Schmidt D.N. Radiolarians decreased silicification as an evolutionary response to reduced Cenozoic ocean silica availability // PNAS. 2009. V. 106. P. 9333–9338.
Lazarus D., Suzuki N., Caulet J.-P. An evaluated list of Cenozoic-Recent radiolarian species names (Polycystinea), based on those used in the DSDP, ODP and IODP deep-sea drilling programs // Zootaxa. 2015. V. 3999(3). P. 301–333. https://doi.org/10.11646/zootaxa.3999.3.1
Lazarus D., Suzuki N., Ishitani Y., Takahashi K. Paleobiology of the Polycystine Radiolaria. Wiley Blackwell, 2021. 966 p.
Liu L., Zhang Q., Chen M.H. et al. Radiolarian biogeography in surface sediments of the Northwest Pacific marginal seas // Science China Earth Sciences. 2017. V. 60. P. 517–530. https://doi.org/10.1007/s11430-016-5179-4
Matsuzaki K.M., Suzuki N., Nishi H. et al. Middle to Late Pleistocene radiolarian biostratigraphy in the water-mixed region of the Kuroshio and Oyashio currents, northeastern margin of Japan (JAMSTEC Hole 902-C9001C) // J. Micropalaeontol. 2014. V. 230. № 2. P. 205–222. https://doi.org/10.1144/jmpaleo2013-012
Matul A., Abelmann A., Gersonde R. New data on the distribution and microphotographs of radiolarians in the bottom surface sediments of the North Pacific and Bering Sea obtained within the KALMAR II and INOPEX projects // Data in Brief. 2019. V. 26, 104448. P. 1–3. https://doi.org/10.1016/j.dib.2019.104448
Munir S., Rogers J., Zhang X. et al. The horizontal distribution of planktonic siliceous Radiolarian community in the Eastern Indian Ocean // Water. 2020. V. 12. P. 1–29. https://doi.org/10.3390/w12123502
Munir S., Sun J., Morton S.L. The First Record and Classification of Planktonic Radiolarian (Phylum Retaria) and Phaeodarian (Phylum Cercozoa) in the Eastern Indian Ocean // Biology. 2021. V. 10, 202. P. 1–41. https://doi.org/10.3390/biolog_y10030202.
Nishimura A., Yamauchi M. Radiolarians from the Nankai Trough in the northwest Pacific // News of Osaka Micropaleontologists. 1984. Spec. V. 6. P. 1–148.
O’Dogherty L., Caulet J.-P., Dumitrica P., Suzuki N. Catalogue of Cenozoic radiolarian genera (Class Polycystinea) // Catalog of Cenozoic radiolarians. Ed. O’Dogherty L. Geodiversitas. 2021. V. 43 (21). P 709–1185. https://doi.org/10.5252/geodiversitas2021v43a21. http://geodiversitas.com/43/21.
Paverd P.J. van de. Recent Polycystine Radiolaria from the Snellius-II Expedition. Ph.D. thesis. Center for Marine Earth Science (the Netherlands) and Paleontological Museum in Oslo (Norway), 1995. 351 p.
Perner J. O radiolariích z čzeského útvaru křídového // Věstník Královské České Spolecnosti Nauk. Třida mathematichko-přirodovĕdecká. Ročnik 1891. Praha, 1891. S. 255–269.
Petrushevskaya M.G., Swanberg N.R. Variability in skeletal morphology of colonial radiolaria (Actinopoda: Polycystinea: Collosphaeridae) // Micropaleontol. 1990. V. 36. № 1. P. 65–85.
Popofsky A. Die Collosphaeriden der Deutschen Südpolar-Expedition 1901–1903. Mit Nachtragzu den Spumellarien und Nassellarien // Deutsche Südpolar-Expedition 1901–1903. 1917. Zoologie II 16(3). S. 235– 278.
Popova-Goll I.M., Goll R.M. Cenozoic Radiolaria biostratigraphy of Hole 1223A in the North Pacific: ODP Leg 200 // Proc. ODP. Sci. Results. 200 / Eds. Kasahara J. et al. 2006. P. 1–24. www-odp.tamu.edu/publications/200_SR/007/007.htm.
Principi P. Cotributo allo studio dei Radiolari Miocenici Italiani // Bollettino della Società Geologica Italiana. 1909. V. 28. P. 1–22.
Renz G.W. The distribution and ecology of Radiolaria in the Central Pacific plankton and surface sediments // Bull. Scripps Inst. Oceanogr. 1976. V. 22. P. 1–267.
Riedel W.R., Sanfilippo A. Cenozoic Radiolaria from the western tropical Pacific, Leg 7 // Init. Reports DSDP. V. 7 / Eds. Winterer E.L., Riedel R.D. Washington, D.C., 1971. P. 1529–1672.
Riedel W., Westberg M.J. Neogene radiolarians from the eastern tropical Pacific and Caribbean, Deep Sea Drilling Project Leg 68 // Init. Reports DSDP, V. 68 / Eds. Amidei R., M. Lee. Washington, D.C., 1983. P. 289–300.
Sandin M.M., Biard T., Romac S. et al. A morpho-molecular perspective on the diversity and evolution of Spumellaria (Radiolaria) // Protist. 2021. V. 172 125806. P. 1–24. https://doi.org/10.1016/j.protis.2021.125806
Sanfilippo A., Riedel W.R. Cenozoic Radiolaria (exclusive of theoperids, artostrobiids and amphipyndacids) from the Gulf of Mexico, Deep Sea Drilling Project Leg 10 // Init. Reports DSDP. V. 10 / Eds. Worzel J.L. al. Washington, D.C., 1973. P. 475–611.
Sano H., Kuwahara K., Yao A., Agematsu S. Panthalassan seamount-associated Permian-Triassic boundary siliceous rocks, Mino terrane, central Japan // Paleontol. Research. 2010. V. 14. № 4. P. 293–314.
Stocker D.K., Johnson M.D., de Vargas C., Not F. Acquired phototrophy in aquatic protists // Aquatic Microbial Ecology. Inter Research. 2009. V. 57(3,3). P. 279–310.
Suzuki N., Aita Y. Radiolaria: achievements and unresolved issues: taxonomy and cytology // Plankton Benthos Res. 2011. V. 6. № 2. P. 69–91. https://doi.org/10.3800/pbr.6.69
Suzuki N., Not F. Biology and Ecology of Radiolaria // Marine Protists / Eds. Ohtsuka S. et al. Springer Japan, 2015. P. 179–222. https://doi.org/10.1007/978-4-431-55130-0
Suzuki N., O’Dogherty L., Caulet J.-P., Dumitrica P. A new integrated morpho- and molecular systematic classification of Cenozoic radiolarians (Class Polycystinea) – suprageneric taxonomy // Catalog of Cenozoic radiolarians / Ed. O’Dogherty L. Geodiversitas. 2021. V. 43. № 15. P. 405–573. https://doi.org/10.5252/geodiversitas2021v43a15
Swanberg N.R. The trophic role of colonial Radiolaria in oligotrophic oceanic environments // Limnol. Oceanogr. 1983. V. 28. № 4. P. 655–666.
Swanberg N., Anderson O.R. Collozoum caudatum sp. nov.: A giant colonial radiolarian from equatorial and Gulf Stream waters // Deep-Sea Research. 1981. V. 28A. P. 1033–1047.
Takahashi K. Radiolaria: flux, ecology, and taxonomy in the Pacific and Atlantic // Ocean Biocoenosis, Series No. 3. / Ed. Honjo S. Woods Hole Oceanogr. Inst. Press, 1991. 303 p.
Vishnevskaya V.S., Ovechkina M.N., Ustinova M.A. Biostratigraphy and Paleogeography of the Bazhenovo Formation (Upper Jurassic and Lower Cretaceous) Based on Radiolarians, Nannoplankton and Calcareous Dinocysts // Paleontol. J. 2019. V. 53. № 9. P. 916–921. https://doi.org/10.1134/s003103011909017x
Дополнительные материалы отсутствуют.