

Океанология, 2022, T. 62, № 6, стр. 950-964
Распределение склерактиниевых кораллов, населяющих глубины свыше 1000 м в Тихом океане
Н. Б. Келлер 1, *, Н. С. Оськина 1, **, Д. М. Ольшанецкий 2, ***, Ю. А. Зарайская 2, ****
1 Институт океанологии им. П.П. Ширшова РАН
Москва, Россия
2 Геологический Институт РАН
Москва, Россия
* E-mail: keller@ocean.ru
** E-mail: nsoskina@mail.ru
*** E-mail: mitia@list.ru
**** E-mail: yulya.zarayskaya@gmail.com
Поступила в редакцию 12.02.2019
После доработки 19.02.2020
Принята к публикации 25.02.2022
- EDN: PNGFRE
- DOI: 10.31857/S0030157422060053
Аннотация
Анализ собственных и литературных данных по видовому разнообразию склерактиниевых кораллов Тихого океана позволил сравнить их распространение в свете прежде не учитывавнегося фактора – наличия глобального океанического конвейера поверхностных и глубинных течений, являющегося естественным барьером для распространения личинок полипов. Показано, что батиальные кораллы предпочитают западные акватории Тихого океана восточным и что в их распределении существует четкая меридиональная асимметрия. На абиссальных глубинах такая закономерность не наблюдается. Так же показано, что сообщества кораллов, населяющие глубины 1000–2000 м, древнее, чем абиссальная фауна склерактиний, что служит дополнительным аргументом в пользу ранее предложенной гипотезы опроисхождении абиссальных видов Fungiacyatus marenzelleri и Leptopenus discus от бесскелетных предков.
ВВЕДЕНИЕ
Изучение глубоководных склерактиниевых кораллов в океане началось еще в XVIII веке. Библиография по разным аспектам их исследований включает сотни публикаций, в которых описываются отдельные виды кораллов из разных регионов Мирового океана, в основном, из области шельфа и верхней батиали. К настоящему времени известны более 1000 видов этих организмов. Полная ревизия глубоководных склерактиний на видовом уровне была проведена трудами двух ученых – Стефана Кернса [10–18] и Хельмута Зибровиуса [29]. Однако эти исследователи, сделавшие упор на описание и ревизию видов, не создали единых карт географического распространения этих животных по всему Мировому океану по разным диапазонам глубин.
Изучение мелководных одиночных кораллов проще, чем глубоководных, которое затруднено из-за редкости их поселений на океанических просторах. Однако за более чем 200 лет многими экспедициями, среди которых существенная роль принадлежит российским, был собран представительный материал по склерактиниям. На его основе мы провели пространственное исследование, чтобы выявить особенности их распространения по регионам и глубинам. Кроме того, нас интересовало распределение поселений кораллов относительно расположения петель глобального океанского конвейера. Существование конвейера, объединяющего несколько поверхностных и глубинных течений, было доказано относительно недавно, и его роль в развитии глубинной океанской биоты пока только изучается [5, 7, 8].
МАТЕРИАЛЫ И МЕТОДЫ
В основу нашей работы был положен как материал, собранный в экспедициях Института океанологии РАН и обработанный Н.Б Келлер [1–4], так и постанционные списки видов, приведенные в многочисленных зарубежных статьях и монографиях [10–24, 29]. Таксономическое положение видов определялось согласно ревизованной Зибровиусом и Кернсом систематике склерактиний [10–17, 26, 28, 29]. При этом мы использовали сведения о находках видов, не рассматривая число экземпляров, поскольку они приводятся не в каждой работе и могут вызвать неправильную интерпретацию.
В результате были составлены списки видов, встреченных в выбранных нами определенных интервалах глубин, в каждом из которых присутствует десятки видов примерно с 400 станций (табл. 1).
Таблица 1.
Число видов и родов склерактиниевых кораллов по интервалам глубин (первые число – наши данные, вторые числа курсивом – литературные данные)
| Глубина, м | Число станций | Число видов | Число родов | |||
|---|---|---|---|---|---|---|
| 1000–2000 | 27 | 298 | 29 | 80 | 16 | 37 |
| 2000–3000 | 5 | 20 | 5 | 16 | 4 | 10 |
| 3000–4000 | 6 | 11 | 5 | 8 | 4 | 3 |
| 4000–5000 | 27 | 31 | 5 | 3 | 3 | 3 |
РЕЗУЛЬТАТЫ
По данным наших сборов была составлена карта станций (рис. 1), на которых были обнаружены склерактинии, обитающие в Тихом океане в диапазоне глубин от 1000 до 2000 м (табл. 2). Оказалось, что 20 таких станций сконцентрированы в западной части Пацифики и только 2 встречены в ее восточной части у побережья северной Америки. Две петли глубоководного конвейера охарактеризованы находками кораллов, тогда как две других петли конвейера, расположенные в более высоких широтах южной Пацифики, практически лишены таких находок.
Рис. 1.
Распространение кораллов в диапазоне глубин 1000–2000 м по нашим данным с номерами станций, линиями показана схема глобального океанского конвейера согласно картам Кошлякова М.Н. (светлыми линиями обозначена поверхностная компонента циркуляции, темными – глубинная) [8].
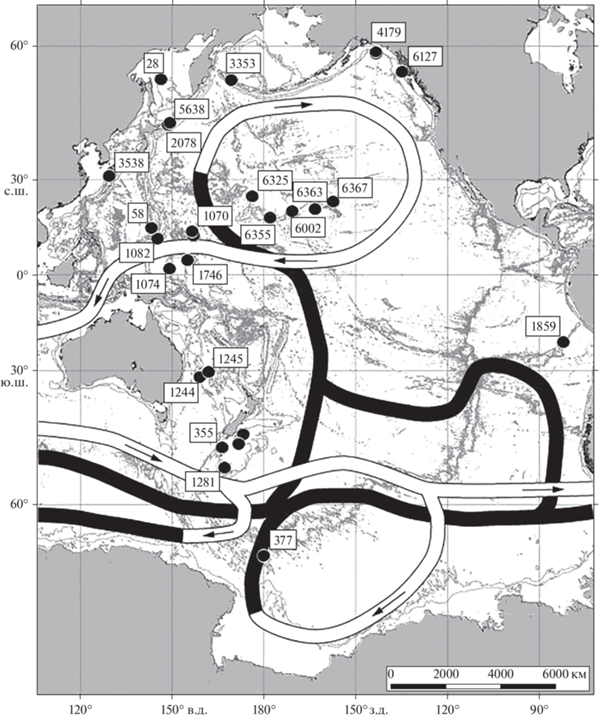
Таблица 2.
Виды склерактиний, поднятых судами Института океанологии c глубины 1000–2000 м
| №№ станций | Глубина, м | Географические координаты |
Название вида |
|---|---|---|---|
| 1244 | 1640 | 31°42′ S 159°00′ E | Fungiacyathus.margaretae Cairns, Deltocyathus vaughani Yabe et Eguichi |
| 1245 | 1210 | 30°25′ S 181°48′ E | Stephanocyathus coronatus (Pourtales), Deltocyathus vaughani Yabe et Eguichi, Conotrochus brunneus Moseley, Fungiacyatus marenzelleri Vaughan |
| 1275 | 1620 | 46°16′ S 173°10′ E | Flabellum impensum Squires |
| 1276 | 1400 | 48°25′ S 171°42′ E | Flabellum knoxi Ralph. et Squires |
| 1281 | 1026 | 53°22′ S 167°07′ E | Fl. knoxi Ralph. et Squires |
| 1746 | 1350 | 4°54′ N 154°59′ E | Madrepora oculata Linnaeus |
| 2078 | 1700 | 44°09′ N 148°38′ E | Javania borealis Cairns, Crispatotrochu rubescens Moseley, Caryophyllia. alaskensis Vaughan |
| 3353 | 1680 | 53°53′ N 169°15′ E | Javania cailleti (Duchassaing et Michelotti) Caryophyllia alaskensis Vaughan |
| 3358 | 1977 | 31°02′ N 129°20′ E | Fungiacyathus. stephanus (Alcock), Letepsammia formosissima Moseleyi, Anthemiphyllia dentata (Alcock) |
| 4179 | 1258 | 12°40′ N 124°59′ E | Caryophyllia. alaskensis Vaughan |
| 5638 | 1845 | 44°36′ N 149°07′ E | Caryophyllia. ambrosia Alcock |
| 6002-12 | 1940 | 20°41′ N 170°52′ W | Deltocyathus parvulus Keller |
| 6127 | 1340 | 55°22′ N 134°50′ W | Caryophyllia ambrosia Alcock |
| 6325 | 1950 | 25°08′ N 176°10′ E | Deltocyathus. magnificus Moseley Deltocyathus murrayi Gardiner et Waugh |
| 6355 | 1630 | 18°32′ N 177°59′ W | Flabellum trapezoidum Keller |
| 6363 | 1350 | 21°10′ N 163°16′ W | Flabellum marcus Keller |
| 6367 | 1420 | 23°32′ N 157°23′ W | Caryophyllya pacifica Keller, Vaughanella.oreophila Keller |
| 355 | 1200 | 49°10′ S 166°23′ E | Flabellumapertum Moseley |
| 377 | 900 | 67°21′ S 179°53′ E | Gardineria antarctica Gardiner |
| 1047 | 1490 | 12°54′ N 156°59′ E | Flabellum. marcus Keller |
| 1054 | 1424 | 12°52′ N 156°54′ E | Flabellum marcus Keller Fungiacyathus pliciseptus Keller |
| 1070 | 1860 | 14°09′ N 156°31′ E | Peponocyatus australiensis (Duncan) |
| 1074 | 1930 | 02°12′ N 148°03′ E | Enalopsammia rostrata (Pourtales) |
| 1082 | 1150 | 12°00′ N 145°05′ E | Madrepora oculata Linnaeus |
| Ly28 | 1642 | 54°00′ N 146°17′ E | Caryophyllyajaponica Marenzeller |
| 58 | 970 | 15°23′ N 143°07′ E | Madrepora oculata Linnaeus |
| 1859 | 1058 | 21°40′ S 82°01′ W | Desmophyllum dianthus (Esper) |
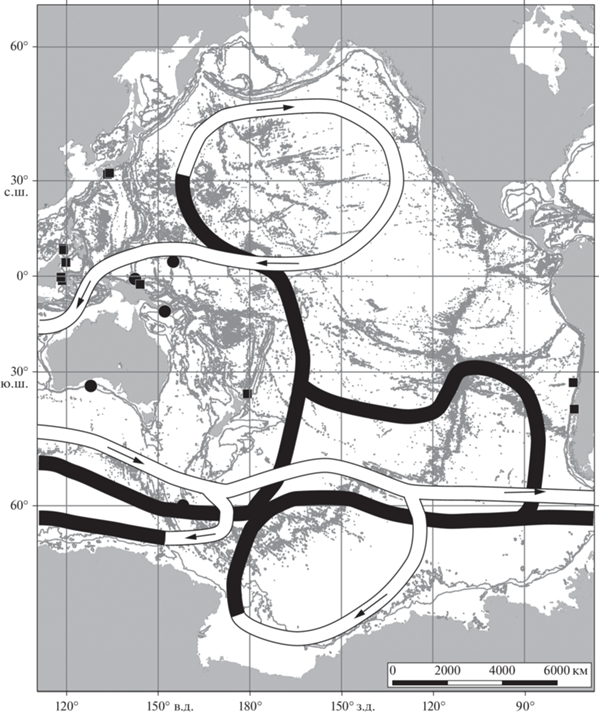
По литературным свидетельствам в этом диапазоне глубин обнаружено еще 80 видов, поднятых с 298 станций (табл. 1). Вместе с нашими находками мы получили аналогичную, но более четкую и выразительную картину приуроченности кораллов к западному сектору Тихого океана. Наибольшее число видов этих животных обитает между 30° с.ш. и 50° ю.ш. в пределах центральной части петли конвейера, образованной поверхностной и придонной его частями и в азиатском регионе. В верхней северной петле конвейера встречено всего 6 станций с видами кораллов. В южной части океана внутри двух петель конвейера, верхняя из которых образована его глубинной частью, находки склерактиний отсутствуют (рис. 2). Отмеченная здесь приуроченность глубоководных кораллов к западной части океана были описаны нами ранее [3, 4] и для мелководных кораллов. Был сделан вывод, что в западной части Тихого океана тропического и субтропического поясов зафиксировано наибольшее число видов и экземпляров мелководных склерактиний, особенно в районе Индо-Малайского архипелага и у Филиппин.
Рис. 2.
Распространение кораллов в диапазоне глубин 1000–2000 м. Кружком обозначены станции нашей коллекции, квадратами – станции из литературных источников, линиями – схема глобального океанского конвейера согласно картам Кошлякова М.Н. (светлыми линиями обозначена поверхностная компонента циркуляции, темными – глубинная) [8].

Подобная закономерность прослеживается и на глубине 2000–3000 м (за исключением двух станций в районе Перуано-Чилийского желоба). С 18 станций поднято 16 видов кораллов, два из которых принадлежат роду Fungiacyathus, два роду Stephanocyathus, пять роду Caryophyllia, по одному родам Crispatotrochus, Paracyathus, Мadrepora, Deltocyathus и Enallopsammia, два роду Flabellum (табл. 3). В западных частях океана сосредоточена основная масса кораллов (рис. 3).
Таблица 3.
Виды склерактиний, обнаруженные на глубинах 2000–3000 м по нашим и литературным данным
| Глубина, м | Географические координаты | Название вида |
|---|---|---|
| 3000 | 40°30′ N 125°45′ W | Fungiacyathus marenzelleri Vaughan |
| 2542 | 13°29′ S 147°12′ E | Fungiacyathus stephanus (Alcock) |
| 2000 | 11°46′ S 178°27′ W | Stephanocyathus nobilis (Moseley) |
| 2500 | 09°13′ N 118°51′ E | Stephanocyathus imperialis Cairns |
| 2450 | 29°42′ S 159°48′ E | Caryophyllia scobinosa Alcock |
| 2309 | 1°55′ S 127°42′ E | Caryophyllia diomedeae Marenzeller |
| 2603 | 31°55′ N 133°23′ E | Caryophyllia cornulum (Cairns) |
| 2350 | 0°13′ S 118°12′ E | Caryophyllia cornulum (Cairns) |
| – | 32°14′ N 134° E | |
| 2430 | 04°49′ S 154°54′ E | Caryophyllia ambrosia Alcock |
| 2970 | 00°56′ S 142°25′ E | |
| 2515 | 36°00′ S 179°12′ E | Crispatotrochus curvatus Cairns |
| 2000 | 37°29′ S 172°00′ E | Paracyathus conceptus Gardiner |
| 2161 | 1°19′ S 118°40′ E | Мadrepora oculata Linnaeus |
| 2310 | 21°10′ N 163°13′ E | Deltocyathus lens Alcock |
| 2570 | 4°38′ N 119°49′ E | Flabellum conicus Moseley |
| 2755 | 33°56′ S 127°55′ E | Flabellum impensum Squires |
| 2022 | 9°13′ N 118°51′ E | Enallopsammia rostrata (Pourtales) |
Рис. 3.
Распространение кораллов в диапазоне глубин 2000–3000 м. Кружком обозначены станции нашей коллекции, квадратами – станции из литературных источников, линиями – схема глобального океанского конвейера согласно картам Кошлякова М.Н. (светлыми линиями обозначена поверхностная компонента циркуляции, темными – глубинная) [8].
В глубинном интервале 3000–4000 м, т е в переходной к абиссали зоне (табл. 4), было обнаружено 11 станций (табл. 1), с которых поднято всего 8 видов склерактиний, три из которых принадлежит роду Fungiacyathus, два – роду Leptopenus, два – роду Deltocyathus и один – роду Flabellum. Вне петли конвейера в северной Пацифике единичные находки приурочены к азиатскому побережью и к западному побережью северной Америки. Отмечены немногочисленные станции с кораллами вдоль побережья Южной Америки. Внутри большей петли конвейера найдены всего 2 станции (рис. 4, 5).
Таблица 4.
Виды склерактиний, обнаруженные на глубинах 3000–4000 м по нашим и литературным данным
| Глубина, м | Географические координаты | Название вида |
|---|---|---|
| 3676 | 31°19′ N 119°39′ W | Fungiacyatus marenzelleri Vaughan |
| 3700 | 32°25′ N 120°40′ W | Fungiacyatus marenzelleri Vaughan |
| 3880 | 34°24′ N 128°12′ E | Fungiacyatus marenzelleri Vaughan |
| 3000 | 40°30′ N 125°45′ W | Fungiacyatus marenzelleri Vaughan |
| 3820 | 80°07′ S 104°10′ W | Fungiacyatus marenzelleri Vaughan |
| 11°30′ S 152°10′ E | Fungiacyatus marenzelleri Vaughan | |
| 3743 | 14°11′ S 140°05′ E | Fungiacyatus marenzelleri Vaughan |
| 44°24′ N 149°10′ E | Fungiacyathus paliferus (Alcock) | |
| 3840 | 7°10′ S 85°50′ W | Fungiacyathus pseudostephana Keller |
| 3949 | 33°31′ S 74°43′ W | Leptopenus discus Moseley |
| 3743 | 46°22′ N 153°03′ E | Leptopenus solidus Keller |
| 3070 | 11°30′ S 152°10′ E | Deltocyathus magnificus Moseley |
| 4000 | 28°53′ N 137°21′ E | Deltocyathus. parvulus Keller |
| 3949 | 124° S 80°47′ W | Deltocyathus parvulus Keller |
| 3980 | 12°54′ N 160°44′ E | Deltocyathus parvulus Keller |
| 3743 | 46°22′ N 153°03′ E | Flabellum marcus Keller |
Рис. 4.
Распространение кораллов в диапазоне глубин 3000–4000 м по нашим данным с номерами станций, линиями показана схема глобального океанского конвейера согласно картам Кошлякова М.Н. (светлыми линиями обозначена поверхностная компонента циркуляции, темными – глубинная) [8].
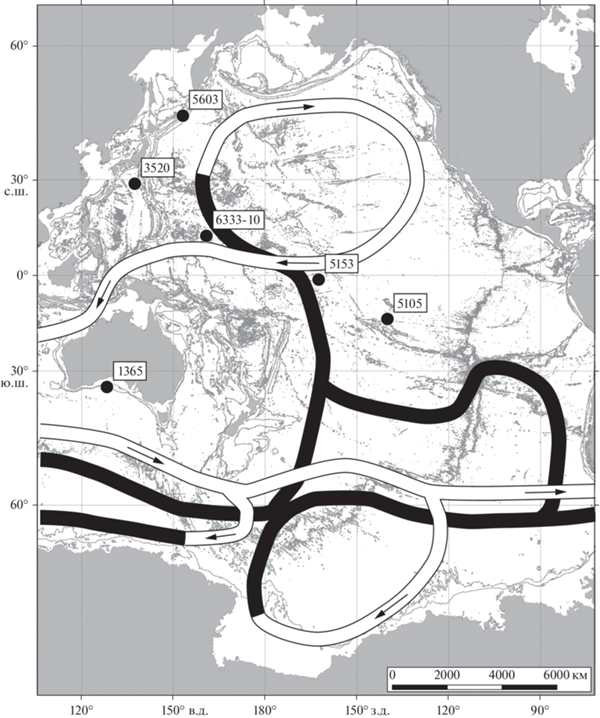
Рис. 5.
Распространение кораллов в диапазоне глубин 3000–4000 м. Кружком обозначены станции нашей коллекции, квадратами – станции из литературных источников, линиями – схема глобального океанского конвейера согласно картам Кошлякова М.Н. (светлыми линиями обозначена поверхностная компонента циркуляции, темными – глубинная) [8].

В нашей статье [7], посвященной абиссальным кораллам и их связи с океанским конвейером, сообщается, что на глубинах более 4000 м в Тихом океане живут виды только трех родов кораллов: Fungiacyathus marenzelleri (Vaughan, 1906), Leptopenus discus Moseley, 1881, Leptopenus. solidus keller, 1972 и Deltocyathus parvulus Keller, 1982 (табл. 5).
Таблица 5.
Виды склерактиний на глубине свыше 4000 м в Тихом океане поднятые судами института океанологии
| Распространение вида Fungiacyathus marenzelleri | |||||
|---|---|---|---|---|---|
| глубина, м | географические координаты | глубина, м | географические координаты | глубина, м | географические координаты |
| 5400 | 54°48′ S 159°10′ E | 4395 | 49°49′ S 148°24′ E | 5540 | 38°25′ S 132°28′ E |
| 5140 | 42°32′ N 150°41′ E | 5070 | 46°07′ N 155°16′ E | 5060 | 44°42′ N 153°49′ E |
| 5780 | 39°02′ N 151°50′ E | 6125 | 33°18′ N 149°45′ E | 5450 | 38°02′ N 146°33′ E |
| 6328 | 53°37′ S 159°40′ E | 4655 | 53°37′ S 159°40′ E | 4477 | 40°22′ N 135°49′ W |
| 4325 | 05°01′ S 139°50′ W | 4200 | 01°22′ S 162°31′ E | 5020 | 46°10′ N 153°07′ E |
| 5210 | 45°18′ N 153°07′ E | 5120 | 45°14′ N 155°15′ E | 5070 | 45°26′ N 154°12′ E |
| 5220 | 45°26′ N 154°12′ E | 6117 | 44°07′ N 149°34′ E | 4720 | 44°17′ N 149°33′ E |
| 5740 | 53°58′ N 157°37′ W | 4740 | 57°00′ N 148°18′ W | 4620 | 53°35′ N 163°23′ W |
| 4820 | 51°40′ N 163° W | 4919 | 35°33′ S 159°05′ E | 4961 | 36°55′ S 159°31′ E |
| Распространение вида Leptopenus discus | |||||
| 4655 | 46°56′ N 143°53′ W | 5000 | 52°13′ N 163°43′ W | 4820 | 51°40′ N 163° W |
| Распространение вида Deltocyathus parvulus | |||||
| 5080 | 25°27′ N 143°22′ E | 4600 | 5°00′ S 135°31′ E | 4700 | 21°01′ S 161°03′ W |
| 4680 | 20°06′ N 169°07′ W | 4900 | 10°58′ N 135°38′ E | ||
Наиболее многочисленные находки первых двух видов отмечены в Курило-Камчатском и Алеутском желобах и вдоль западного побережья северной Америки. Внутри двух замкнутых петель глобального конвейера эти виды не зафиксированы [7]. Третий вид не выходит за пределы Тихого океана [4]. Распространение видов F. marenzelleri и L. discus в Мировом океане показано на картах (рис. 6, 7).
Рис. 6.
Распространение кораллов в диапазоне глубин 4000–5000 и более метров по нашим данным с номерами станций, линиями показана схема глобального океанского конвейера согласно картам Кошлякова М.Н. (светлыми линиями обозначена поверхностная компонента циркуляции, темными – глубинная) [8].

Рис. 7.
Распространение Fungiacyathus marenzelleri (Vaughan, 1906) – обозначен кружком и Leptopenus discus (Moseley, 1881) – обозначен звездочкой [7] в диапазоне глубин 4000–5000 и более метров, линиями показана схема глобального океанского конвейера согласно картам Кошлякова М.Н. (светлыми линиями обозначена поверхностная компонента циркуляции, темными – глубинная) [8].

Итак, большая часть видов приходится на глубины 1000–2000 м, существенно меньше видов на отрезках 2000–3000 м и 3000–4000 м (табл. 3, 4). Второй максимум численного, но не видового обилия приходится на глубину от 4000 до 5000 метров, где обитают лишь 3 вида трех родов кораллов (рис. 6, 7). Судя по карте глубинной циркуляции, на этом батиметрическом отрезке глубинная часть океанского конвейера не является препятствием для распространения этих абиссальных видов кораллов, которые встречаются по всей территории морского дна, не отдавая явного предпочтения западной или восточной акваториями Тихого океана.
На всех прочих глубинах мы имеем дело с четко выраженной меридиональной асимметрией в распространении кораллов в Тихом океане.
ОБСУЖДЕНИЕ
В одной из предыдущих статей [5] мы предположили, что глобальный океанский конвейер влияет на распространение глубоководных кораллов двумя способами. Глубоководная его часть может служить либо барьером для расселения придонной группы их личинок, либо может разносить те личинки, которые способны подниматься над местом нереста вдоль траектории движения конвейера [5, 7]. Судя по распространению как всей верхнебатиальной фауны кораллов, так и отдельных видов в Тихом океане, глубинная часть конвейера служила непреодолимым барьером для ее проникновения из западной в восточную Пацифику (рис. 1, 2, 4). Любопытно, что та петля, которая ограничена только глубоководными частями конвейера, характеризуется полным отсутствием кораллов на всех глубинах, кроме абиссальных (рис. 1–6). Кораллы, распространенные в северной, прилегающей к азиатскому материку части, возможно использовали поверхностную часть конвейера для проникновения к берегам Аляски (рис. 1). Но южнее 60° c.ш. они не смогли распространиться: может быть, не успели по времени, если предположить их молодой возраст, или им помешал калифорнийский апвеллинг. Единичные находки близ побережья южной Америки могут быть связаны с проникновением кораллов из Атлантики через существовавший еще в раннем плиоцене Панамский канал [19].
Исходной для ныне живущих кораллов была мелководная фауна океана Тетис. С распадом Тетиса и закрытием в середине миоцена прохода между Индийским океаном и Средиземным морем началось распространение видов в прилегающие акватории. При этом в Тихом океане преобладающая часть склерактиниевых кораллов приходится на западную Пацифику.
Меридиональная асимметрия в распределении многих групп донных беспозвоночных в низких широтах – широко распространенное явление, отражающее меридиональную продукционную асимметрию. Она выражается в возрастании на западе океанов роли таких таксонов батиальных беспозвоночных, как стебельчатые лилии, брахиоподы, и глубоководные кораллы и связывается с системой разнонаправленных круговоротов поверхностных вод у восточных и западных побережий одного и того же океана, как предполагается, возникающих в результате вращения Земли и действия сил Кориолиса [4]. Однако барьер в виде подводной части глобального конвейера в абиссали не работает, так как мощное течение по всей видимости не доходит до самого дна океана и не является препятствием в расселении личинок двух самых глубоководных видов.
При историческом анализе условий формирования фауны обычно оценивался уровень генетических связей между отдельными географическими комплексами таксонов низких рангов, а не собственно возраст фауны и отдельных составляющих ее зональных или биогеографических комплексов. Важнее получить хотя бы спорные, но подкрепленные данными по истории формирования Океана представления об абсолютном возрасте той или иной фауны. Мы попытались выяснить возраст видов кораллов, населяющих разные выделенные нами батиметрические горизонты, что может пролить дополнительный свет на историю развития Океана.
Не строящие рифов глубоководные кораллы, лишенные зооксантелл, обитающие в широком диапазоне глубин, от литорали до абиссали, появляются лишь в средней юре. К концу мела они уже приобрели современный облик. Возраст появления большинства наиболее распространенных в океане родов склерактиний варьирует от средней юры до плейстоцена, но большая их часть появилась в палеогене [25, 27] (табл. 6).
Таблица 6.
Возраст родов, к которым принадлежат исследованные виды
| № | Название рода | Глубина, м | Возраст |
|---|---|---|---|
| 1 | Fungiacyathus | 69–6328 | Эоцен – ныне |
| 2 | Leptopenus | 682– 5000 | Современный |
| 3 | Stephanophyllia | 15–1137 | Эоцен, миоцен – ныне |
| 4 | Madrepora | 15–2700 | Эоцен, миоцен – ныне |
| 5 | Caryophyllia | 0–3200 | Верхняя юра – ныне |
| 6 | Trochocyathus | 32–1675 | Средняя юра – ныне |
| 7 | Stephanocyathus | 141–2210 | Палеоген: (палеоцен) – ныне |
| 8 | Deltocyathus | 13–5080 | Палеоген (эоцен) – ныне |
| 9 | Lophelia | 60–2170 | Палеоген, неоген (олигоцен?, миоцен) – ныне |
| 10 | Conotrochus | 110–1089 | Палеоген (эоцен) – ныне |
| 11 | Crispatotrochus | 82–2329 | Средний мел – палеоген (эоцен) – ныне |
| 12 | Desmophyllum | 35–2460 | Средний мел – ныне |
| 13 | Solenosmilia | 220–2165 | Современный |
| 14 | Paracyathus | 0–1472 | Палеоген (эоцен) – ныне |
| 15 | Peponocyathus | 10–1097 | Палеоген (олигоцен) – ныне |
| 16 | Flabellum | 22–2260 | Верхний мел – ныне |
| 17 | Vaughanella | 100–2000 | Современный |
| 18 | Javania | 32–2165 | Палеоген (эоцен) – ныне |
| 19 | Gardineria | 2–728 | Современный |
| 20 | Enalopsammia | 110–2165 | Неоген (миоцен) – ныне |
Однако возраст появления большей части видов, населяющих современный океан, остается неясным. Это происходит потому, что палеонтология имеет дело исключительно с мелководными склерактиниями, обитающими на глубинах существенно менее 1000 м. Древних одиночных полипов в континентальных разрезах находят в осадках глубин, лежащих не намного глубже, чем современные рифовые отложения [9]. Глубоководные же отложения с кораллами на суше практически не встречаются. При бурении океанского дна в глубоководных зонах океанов склерактинии, как правило, не обнаруживаются, поэтому определить возраст появления видов глубоководных кораллов обычными стратиграфическими методами практически невозможно. Для оценки возникновения ряда видов нами сделана первая попытка подойти к этой проблеме не традиционным биостратиграфическим методом, а палеоэкологическим путем. Для этого использовались данные о температуре вод океана в геологическом прошлом и установленные нами температурные диапазоны 45 видов кораллов, большинство которых принадлежит таким древнейшим родам как Caryophyllia (c верхней юры) и Flabellum (с верхнего мела), а также возникшего в палеогене рода Javania и современного рода Gardineria на фоне разнообразных видов с космополитическим распространением (табл. 6) [6]. Этот метод мы применили здесь в надежде выяснить возраст наиболее распространенных из имеющихся в нашей коллекции тихоокеанских кораллов. Виды были распределены по четырем градациям: наиболее холодноводная (А), умеренно холодноводная (Б), умеренная (В) и тепловодная (Г). По нашим расчетам виды градации А возникли не ранее позднего эоцена – начала олигоцена. Виды градации Б образовались в середине – конце эоцена. Виды градации В произошли в палеоцене и ранее. Время возникновения видов градации Г – низы палеоцена, возможно мел [6].
Если наши выводы о возрасте верны, оказывается, что наиболее древними являются виды, обнаруженные нами на отрезке глубин 1000–2000 м. Как было показано ранее [6], некоторые из этих видов могли появиться и в течение эоцена и даже в палеоцене, то есть 45–60 миллионов лет назад и даже раньше. Холодноводные же виды с узкими температурными границами, показывающие максимум обилия экземпляров на глубине от 4000 до 5000 метров, являются более молодыми и могли возникнуть не ранее начала олигоцена, то есть в пределах 30–31 миллиона лет назад или даже в миоцене, 11–12 млн лет назад [6]. По-видимому, это связано с другим исходным фактором происхождения фауны, не имеющим отношения к Тетису. Этим подтверждается наше первоначальное предположение об исключительной молодости глубоководной фауны [4].
Итак, на глубинах более 4000 м в Тихом океане живут только 3 вида трех родов кораллов. Это Fungiacyathus marenzelleri (Vaughan, 1906), Leptopenus discus Moseley, 1881 и Deltocyathus parvulus N. Keller, 1982 (табл. 5).
Наши исследования последних лет, касающиеся адаптивной морфологии абиссальных склерактиний показали, что эти виды представляют собой как бы живую модель того, каким мог бы быть скелет коралла на ранних стадиях его формирования. Особенно ярко это выражено у видов рода Leptopenus: тонкий и ажурный скелет является как бы корочкой, выделенной подошвой актинии, а центральная колонна состоит из тонких игольчатых шиповидных элементов. Эти обстоятельства в сочетании с результатами независимых экспериментов Саммарко и Ричмонда [4] привели нас к выводу, что предковыми формами склерактиний вероятнее всего были бесскелетные организмы, относящиеся либо к самостоятельной филогенетической линии, либо, что более вероятно, к линии, происшедшей в результате утраты скелета одной из групп ругоз – их палеозойских аналогов. Потеря или частичная утрата скелета в экстремальных ситуациях, неоднократно случавшихся в постпалеозойской истории Земли, могла происходить у кораллов неоднократно [4]. Неблагоприятные обстоятельства они вероятнее всего переживали в рефугиях батиали. При изменении экологических условий на благоприятные склерактинии могли подняться на верхние участки континентального склона, что сопровождалось вспышками видообразования.
Исходя из видового и количественного обилия коралловой фауны многие исследователи считали Индо-Малайский архипелаг основным центром происхождения не только мелководной, но также и глубоководной фауны. В частности, на примере кораллов, населяющих эту акваторию, Кун и Сквайр разработали схему исторического развития семейства Micrabaciidae. Они считают, что постепенное проникновение этого семейства в глубины началось в палеоцене в Индо-Малайском районе Тихого океана и сопровождалось редукцией скелета и неотеническим развитием его глубоководных видов. Кун приводит цепь генетически близких родов этого семейства, последовательно завоевывающих все большие глубины: Discopsammia – Stephanopsammia – Stephanophyllia – Letepsammia – Leptopenus [4]. Все современные роды, кроме Letepsammia и Leptopenus обитают на глубинах не более 800 м. Если это так, то в филогенетической ветви, выстроенной Куном, с той же степенью вероятности можно было бы вести линию семейства в обратном порядке, начиная от рода Leptopenus, если допустить, что этот примитивный род является родоначальником семейства Micrabaciidae, пережившим в убежищах на глубинах океана неблагоприятные периоды. Это один из примеров, показывающих, что рассуждения о предковых формах современных глубоководных кораллов в большинстве случаев имеют чисто умозрительный характер. Поэтому в отношении центров формирования глубоководной фауны мы склонны присоединиться к мнению, что их вообще могло не существовать, и процесс завоевания больших глубин океана мог начаться в любой момент его постмезозойской истории, так как каждому виду свойственно стремление расширить свой ареал.
Еще одним подтверждением молодости абиссальной фауны коралловых полипов служит удивительное совпадение особенностей распределения мелководных и батиальных видов. Конечно, все изложенное не может рассматриваться как прямое доказательство молодости абиссальных видов кораллов. Как мы уже писали, современные палеонтологические свидетельства не могут ни опровергнуть, ни подтвердить высказанные нами предположения. В многочисленных публикациях, посвященным находкам ископаемых кораллов, обычно лишь отмечается принадлежность кораллов к тем или иным семействам ныне процветающих полипов и их плиоцен–плейстоценовое происхождение.
Итак, два из населяющих абиссаль видов (F. marenzelleri и L. discus), по нашему мнению, молодые, возникшие в глубинных слоях океана от бесскелетных предков, и в дальнейшем распространившиеся в более высокие горизонты. Третий вид, Deltocyathus parvulus, произошел от мелководных предков и спустился в абиссаль в результате перехода к неотеническому развитию. Об этом подробно рассказано в нашей предыдущей публикации [4]. Таким образом, если наши определения возраста видов склерактиний верны, приходится признать, что существующая в настоящее время фауна абиссальных коралловых полипов, представляет собой молодое, плейстоцен – голоценовое образование. Все это еще раз подтверждает правомочность нашего предположения о молодости глубоководной фауны.
ВЫВОДЫ
Анализ карт, построенных на основании авторских и литературных данных о сообществах склерактиний в Тихом океане, показал, что, судя по распространению батиальной фауны кораллов в Тихом океане, глубинная часть конвейера служила непреодолимым барьером для ее проникновения из западной в восточную Пацифику. При этом кораллы четко предпочитают западное полушарие восточному, что свидетельствует о существовании меридиональной асимметрия в их распределении и связано с влиянием глобального океанского конвейера и с историей развития океана. Однако в абиссали распределение кораллов иное и не подчиняется такому правилу.
Предположительный возраст видов, населяющих выделенные батиметрические горизонты, заставляет признать, что существующая в настоящее время фауна абиссальных коралловых полипов представляет собой молодое, плейстоцен – голоценовое образование.
Это рассматривается в качестве дополнительного аргумента в пользу ранее предложенной конструкции происхождения глубоководных склерактиний от мягкотелых предков.
Источники финансирования. Исследование выполнено в рамках государственного задания: тема № 0128-2021-0008.
Список литературы
Келлер Н.Б. Одиночные мадрепоровые кораллы (Madreporaria). В кн.: Бентос подводной горы Маркус-Неккер и смежных районов Тихого океана. М.: ИО АН СССР, 1981. С. 28–39.
Келлер Н.Б. Склерактиниевые беззооксантеллятные кораллы подводных гор Наска и Сала и Гомес. В кн.: Бентос высокоширотных районов. М.: ВНИРО, 1998. С. 89–93.
Келлер Н.Б. Пространственное распределение видов мадрепоровых кораллов, не имеющих зооксантелл (Cnidaria, Anthozoa) // Океанология. 1998. Т. 38. № 2. С. 227–232.
Келлер Н.Б. Глубоководные склерактиниевые кораллы. М: Крассанд, 2012. 384 с.
Келлер Н.Б., Оськина Н.С., Савилова Т.А. Распространение глубоководных склерактиниевых кораллов в Атлантическом океане // Океанология. 2017. Т 57. № 2. С. 331–338.
Келлер Н.Б., Оськина Н.С., Николаев С.Д. Новый подход к определению возраста глубоководных видов склерактиний на основании температурных диапазонов их обитания // Доклады РАН. 2009. Т. 425. № 2. С. 218–222.
Келлер Н.Б., Оськина Н.С., Савилова Т.А. Существут ли связь абиссальных склерактиниевых коралов с океанским конвейером // Океанология. 2017. Т. 57. № 4. С. 611–617.
Кошляков М.Н., Тараканов Р.Ю. Перенос воды через субантарктический фронт и глобальный океанский конвейер // Океанология. 2011. Т. 51. № 5. С. 743–767.
Alloiteau J. Contribution a la systematique des Madreporaires fossils. Paris: Centre national de la recherche scientifique, 1957. 462 p.
Cairns S.D. New records of ahermatypic corals (Scleractinia) from the Hawaiian and Line Islands // Occas. Papers of the Bernice Pauahi Bishop Mus. 1984. V. 25 (10). 30 p.
Cairns S.D. A revision of the ahermatypic Scleractinia of the Philippine Islands and adjacent waters. Part 1: Fungiacyathidae, Micrabaciidae, Turbinoliinae, Guyniidae, and Flabellida // Smithsonian Contrib. Zool. 1989. № 486. 136 p.
Cairns S.D. Scleractinia of the temperate North Pacific // Smithsonian Contrib. Zool. 1994. V. 557. 150 p.
Cairns S.D. The marine fauna of New Zealand: Scleractinia (Cnidaria: Anthozoa) // New Zealand Oceanograph. Inst. Mem. 1995. V. 103. 144 p.
Cairns S.D. Cnidaria Anthozoa: Deep-water azooxanthellate Scleractinia from Vanuatu, Wallis and Futuna Islands // A.Crosnier (ed.). Resultats des Camp. Musorstom. V. 20. Mem. Mus. Histoire Naturelle. 1999. № 180. P. 31–167.
Cairns S.D. The Azooxanthellate Scleractinia (Coelenterata Anthozoa) of Australia // Records of the Australian Museum. 2004. V. 56. P. 259–329.
Cairns S.D., Parker S.A. Review of the recent Scleractinia of South Australia, Victoria, and Tasmania // Records of the south Australian Museum. Monograph Series 3. 1992. 82 p.
Cairns S.D., Haussermannt V, Forssterra G. A review of the Scleractinia (Cnidaria: Anthozoa) of Chile, with the description of two new species // Zootaxa. 2005. V. 1018. P. 15–46.
Cairns S.D., Zibrowius H. Cnidaria Anthozoa: Azooxanthellate Scleractinia from the Philippine and Indonesian Regions. In: A. Crosnier & P.Bouchet (eds.). / Result. des Camp. Musorstom. V.16. Mem. Mus. Nat. Hist. Nat. 1997. V. 172. 214 p.
Coates A.G., Stallard R.F. How old is the Isthmus ofPanama? // Bull. Mar. Sci. 2013. V. 89. P. 801–813. https://doi.org/10.5343/bms.2012.1076
Gardiner J.S., Waugh P. The flabellid and turbinolid corals // Sci. Rep. John Murray Expedition (1933–1934). 1938. V. 5. № 7. P. 167–202.
Gardiner J.S., Waugh P. Madreporaria excluding Flabellidae and Turbinolidae // Sci. Rep. John Murray Expedition (1933–1934). 1939. V. 6(5). P. 225–242.
Marenzeller E von. Reports on the dredging operations off the west coast of Central America to the Galapagos, to the west coast of Mexico and in the Gulf of California, in charge of Alexander Agassiz, carried on by the U.S. Fish Commission steamer “Albatross”, during 1891, Lieut. Commander Z. L. Tanner, U.S.N., Commanding. XXXIII. Stein- und Hydro-korallen // Bulletin of the Museum of Comparative Zoology at Harvard College. 1904. V. 43 (2)/ P. 75–87.
Moseley H.N. On the deep-sea Madreporaria // Voyage H. M. S. “Challenger” Repts. Scient. Results. Zool. London. 1881. V. 2. Part. 7. P. 127–208.
Vaughan T.W. Reports on the scientific results of the expedition to the eastern tropical Pacific by the U.S. Fish commission steamer Albatross from October, 1904 to march, 1905. Part 6: Madreporaria // Bull. of the Museum of Comparative zoology. 1906. V. 50. № 3. P. 61–72.
Vaughan T.W. Recent Madreporaria of the Hawaiian Islands and Laysan // Bull. U.S. Nat. Mus. 1907. № 59. 427 p.
Vaughan T.W., Wells J.W. Revision of the suborders, families and genera of the Scleractinia // Geol. Soc. Amer. Spec. Pap. 1943. V. 44. 363 p.
Wells J.W. Notes on Indo-Pacific Scleractinian corals. Part. 10. Late Pleistocene ahermatypic corals from Vanuatu // Pacific Science. 1984. V. 38. № 3. P. 205–219.
Wells J.W. Scleractinia // In Treatise on invertebrate paleontology. 1956. Ed. R. C. Moore. F. Coelenterata. Univ. Kansas Press. P. 328–444.
Zibrowius H. Les scléractiniaires du miocène au pléistocene de Sicile et de Calabre de Giuseppe Seguenza (1864, 1880) (Cnidaria, Anthozoa) // Academia Peloritana dei Pericolanti Classe 1 di Scienze Fis. Mat., Nat. 1991. V. LXV11 (1989). Supplemento N 1 (1). 135 p.
Дополнительные материалы отсутствуют.