

Онтогенез, 2023, T. 54, № 2, стр. 172-175
Развитие идеи Н.К. Кольцова о генетической организации междисков политенных хромосом Drosophila melanogaster
И. Ф. Жимулев a, *, Т. Ю. Ватолина a, В. Г. Левицкий b, Т. Д. Колесникова a, А. В. Цуканов b
a Институт молекулярной и клеточной биологии СО РАН
630090 Новосибирск, просп. ак. Лаврентьева, 8/2, Россия
b Институт цитологии и генетики СО РАН
630090 Новосибирск, просп. ак. Лаврентьева, 10, Россия
* E-mail: zhimulev@mcb.nsc.ru
Поступила в редакцию 27.12.2022
После доработки 24.03.2023
Принята к публикации 04.04.2023
- EDN: XFKXAP
- DOI: 10.31857/S0475145023020064
Аннотация
В работе впервые исследовали организацию промоторов генов развития и генов, необходимых для общеклеточных функций – “домашнего хозяйства” клетки в полном геноме Drosophila melanogaster. С помощью биоинформатических методов показано, что гены, промоторы которых расположены в междисках политенных хромосом, обогащены функциями, связанными с общеклеточными процессами, тогда как остальная часть генов (примерно половина генома Drosophila) связана с узкоспециализированными процессами, происходящими в ходе развития. В промоторной зоне генов “домашнего хозяйства” обнаружено четыре специфичных мотива, которые могут присутствовать у разных генов индивидуально или в различных комбинациях. Существенная часть междисковых промоторов не содержит выявленных мотивов. Анализ, проведенный с помощью Gene Ontology, показал, что для отдельных групп междисковых генов, содержащих в промоторах один мотив или их комбинации, характерно выполнение определенных функций.
ВВЕДЕНИЕ
Вскоре после открытия политенных хромосом в 1934 г. началась бурное обсуждение их структуры и генетической организации, и Н.К. Кольцов (Koltzoff, 1934) сделал ряд предположений, которые привлекли внимание исследователей. Он первым высказал гипотезу о том, что хромосомы этого типа состоят из многих нитей, т.е. они многонитчатые (или политенные), кроме того, он предположил, что гены находятся в междисках политенных хромосом. После этого проблемы генетической организации интерфазных хромосом оказались в центре внимания многих генетиков на долгие годы. Высказывали многочисленные гипотезы, из которых хотелось бы упомянуть три: (Crick, 1971; Paul, 1972; Zhimulev, Belyaeva, 1975). В первых двух авторы предположили, что каждый ген занимает диск и соседний с ним междиск, авторы третьей предположили, что гены домашнего хозяйства и гены развития занимают соответственно междиски и разные типы дисков.
Ранее нами был предложен метод одновременной локализации дисков и междисков на физической карте ДНК и на цитологической карте, что позволило совместить расположение генов разных типов на хромосомах и карте ДНК. На следующем этапе, используя различия в спектрах белков и модификаций гистонов в участках локализации генов разных типов, разработали математическую модель организации хроматина (4НММ) (Zhimulev et al., 2014). С помощью этой модели весь хроматин в интерфазных ядрах клеток дрозофилы был разделен на четыре состояния (названы, как принято по названию оптического цвета – аквамарин, лазурит малахит, рубин). Каждый цвет соответствует степени деконденсации хроматина и открытости его для транскрипции. Самый деконденсированный – аквамарин, самый конденсированный – рубин, соответствуют междискам и черным дискам соответственно, лазурит – это серые разрыхленные диски (Zykova et al., 2018). Благодаря этой модели все гены генома дрозофилы были распределены в две группы – 6562 гена домашнего хозяйства (в литературе также используется термин “гены с повсеместной активностью”), промоторы которых расположены в участках хроматина типа аквамарин (междиски политенных интерфазных хромосом), а тело гена (экзоны и интроны) – в хроматине лазурит (в серых дисках политенных хромосом). Оставшиеся 5664 гена развития расположены в остальной части генома.
В области исследований организации промоторов у генов высших Metazoa достигнуты огромные успехи. Сейчас хорошо известно, что в окрестностях точки инициации транскрипции – Inr, ниже и выше на 100–200 пар нуклеотидов, располагаются специфические мотивы (короткие специфические для различных функции последовательности нуклеотидов), совершенно необходимые для инициации и продолжения транскрипции (Lenhard et al., 2012; Danino et al., 2015; Anderson, Sandelin, 2020; Ramalingam et al., 2021). Тем не менее в понимании структуры промоторов все-таки довольно много неизученного. Во-первых, до сих пор нет исследований полногеномной организации промоторов у генов развития и “домашнего хозяйства”, поскольку до сих пор не было метода, позволяющего разделить эти группы генов, во-вторых, нет четкого полногеномного разделения на гены “домашнего хозяйства” и гены развития, несмотря на то, что сведения об этом уже накапливаются и показано, что промоторы этих групп генов могут различаться. В данной работе мы попытались найти наиболее важные закономерности в организации мотивов в промоторах генов “домашнего хозяйства” и генах развития в полном геноме дрозофилы.
МАТЕРИАЛЫ И МЕТОДЫ
Мы проанализировали 13 574 белок-кодирующих гена Drosophila melanogaster из базы данных FlyBase (R 5.57). В анализ были взяты 5'-области генов от –300 до +200 п.н. относительно сайтов начала транскрипции и полностью картированные в доменах состояний хроматина аквамарин, лазурит, малахит и рубин, как описано ранее (Levitsky et al., 2020). Из анализа были удалены все околопромоторные участки, которые имели кодирующие последовательностям в пределах анализируемой области. Таким образом, набор околопромоторных участков соответствовал 6562, 874, 1628 и 3162 генам аквамарин, лазурит, малахит и рубин соответственно. Мы применили инструмент STREME для de novo поиска мотивов в околопромоторных участков для каждого состояния хроматина в трех тестах, относительно либо других состояний хроматина, либо набора последовательностей, содержащего случайные геномные локусы. Мы применили поправку Бонферрони к обогащенным мотивам: значение p/N < 0.05, где p означает статистическую значимость мотива, а N обозначает количество мотивов, найденных STREME (https:// meme-suite.org/meme/doc/streme.html). Использовали инструмент TOMTOM (https://meme-suite.org/ meme/tools/tomtom) для сравнения обогащенных мотивов с мотивами для известных факторов транскрипции из баз данных CISBP (http:// cisbp.ccbr.utoronto.ca/) и JASPAR (https://jaspar.uio.no/).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
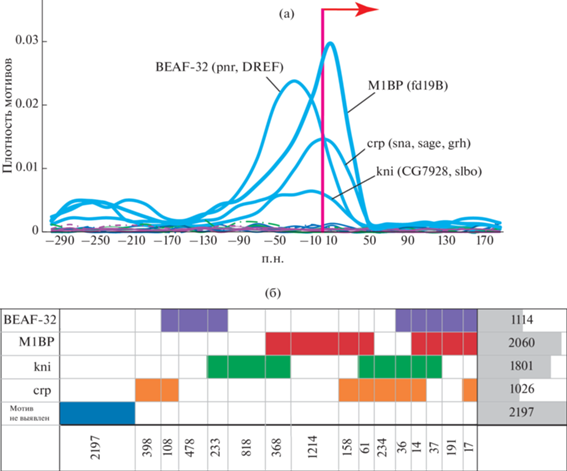
Воспользовавшись данными модели 4НММ, мы отобрали гены “домашнего хозяйства” и гены развития в полном геноме дрозофилы и затем обработали последовательности нуклеотидов в промоторах обеих групп генов с помощью инструмента STREME (Sensitive, Thorough, Rapid, Enriched Motif Elicitation). Эта программа позволяет выявлять de novo обогащение мотивами в определенных районах генома. Изучены околопромоторные участки (от –300 до +200 п.н. вокруг точки инициации транскрипции) у 6562 генов из аквамарин хроматина при сравнении их с последовательностями генов контрастного состояния – рубин (3162 генов), а также против генов полного генома дрозофилы и против суммы генов в трех состояниях, исключая аквамарин (5664 гена). Наши результаты показали, что можно выделить достаточно много мотивов, по которым различаются промоторы генов, локализованных в разных состояниях хроматина. Поэтому для ужесточения степени достоверности оценок из этих значений отобрали только те, которые удовлетворяют следующим дополнительным критериям: максимально низкое значение E-value, максимальное обогащение по сравнению с фоном, наличие сведений в других базах данных, а также воспроизводимость в разных вариантах сравнений. Оказалось, что этим критериям удовлетворяют только четыре мотива: M1BP (fd19B), BEAF-32 (pnr, DREF), crp (sna, sage, grh), kni (CG7928, slbo) (рис. 1а). Локализация этих мотивов в остальных хроматинах, помимо аквамарина, показана на уровне оси абсцисс (рис. 1а).
Рис. 1.
Характеристики промоторных участков 6562 генов домашнего хозяйства дрозофилы. (а) Суммарная локализация мотивов относительно точки инициации транскрипции (красная стрелка). По оси абсцисс показано расстояние до точки инициации транскрипции, по оси ординат частота локализации мотива в данной координате околопромоторного участка. Приведены наиболее употребительные названия мотива, а в скобках указаны названия, которые встречаются в других статьях/базах данных. Изученные мотивы присутствуют только в промоторных участках хроматина аквамарин (голубая линия), все остальные промоторные области не содержат этих мотивов (зеленая, синяя, фиолетовая линии на уровне нулевого значения по оси абсцисс). (б) Числа генов домашнего хозяйства, имеющих только один мотив (вертикальная колонка цифр справа) или комбинации мотивов (горизонтальная последовательность цифр внизу). Прямоугольниками разных цветов обозначено наличие изучаемого мотива (отсутствие цвета в клетке обозначает отсутствие мотива в данном наборе генов) генов).
Аналогичная работа была проведена для сравнений генов в максимально контрастном состоянии рубин (данные не приводятся). Выявлен один наиболее устойчивый мотив – ТАТА-бокс.
Оказалось, что среди промоторов генов домашнего хозяйства у дрозофилы возможны разнообразные комбинации мотивов. Кроме того, 33% генов этого типа вообще не имеют ни одного из этих мотивов (рис. 1б). Аналогичная работа была проведена для анализа промоторов генов развития, имеющих только очень специфический мотив – ТАТА-бокс (данные не приводятся). Мотивы Mes2, GAGA, ttk обладают меньшей специфичностью. Также обнаружено, что около 50% генов развития не имеют ни ТАТА, ни каких-либо других из найденных мотивов. В аналогичном сравнении локализации мотивов у генов развития максимальная специфика обнаружена только для ТАТА-бокса в положении от –75 до –25 п.н. от точки инициации транскрипции. Анализ, проведенный с помощью Gene Ontology, показал, что для отдельных групп междисковых генов, содержащих в промоторах один мотив или их комбинации, характерно выполнение определенных функций.
Таким образом впервые описаны специфические мотивы в промоторах генов двух разных типов на уровне всего генома и показано, что наборы мотивов у генов “домашнего хозяйства” и генов развития совершенно разные, а наиболее известные мотивы не являются универсальными.
Список литературы
Andersson R., Sandelin A. Determinants of enhancer and promoter activities of regulatory elements // Nat. Rev. Genet. 2020. V. 21. P. 71–87.
Crick F. General model for the chromosomes of higher organisms // Nature. 1971. V. 234. P. 25–27.
Danino Y.M., Even D., Ideses D. et al. The core promoter: At the heart of gene expression // Biochim. Biophys. Acta. 2015. V. 1849(8). P. 1116–1131.
Koltzoff N.K. The structure of the chromosomes in the salivary glands of Drosophila // Science. 1934. V. 60. P. 312–313.
Lenhard B., Sandelin A., Carninci P. Metazoan promoters: emerging characteristics and insights into transcriptional regulation // Nat. Rev. Genet. 2012. V. 13(4). P. 233–245.
Levitsky V.G., Zykova T.Yu., Moshkin Y.M., Zhimulev I.F. Nucleosome positioning around transcription start site correlates with gene expression only for active chromatin state in Drosophila interphase chromosomes // International J. Molecular Sciences. 2020. V. 21(23). P. 9282.
Paul J. General theory of chromosomes structures and gene activation in eukaryotes // Nature. 1972. V. 238. P. 444–446.
Ramalingam V., Natarajan M., Johnston J. et al. TATA and paused promoters active in differentiated tissues have distinct expression characteristics // Mol. Syst. Biol. 2021. V. 17. P. e9866.
Zhimulev I.F., Belyaeva E.S. Proposals to the problem of structural and functional organization of polytene chromosomes // Theor. Appl. Genet. 1975. V. 45(8). P. 335–340.
Zhimulev I.F., Zykova T.Yu., Goncharov F.P. et al. Genetic organization of polytene chromosome bands and interbands in Drosophila melanogaster // PLoS One. 2014. V. 9(7). P. e101631.
Zykova T.Yu., Levitsky V.G., Belyaeva E.S. et al. Polytene chromosomes – a portrait of functional organization of the Drosophila genome // Curr. Genomics. 2018. V. 19(3). P. 179–191.
Дополнительные материалы отсутствуют.