

Палеонтологический журнал, 2019, № 1, стр. 56-60
Новый вид рода Homocloeus Jordan (Coleoptera: Anthribidae) из миоценового мексиканского янтаря
А. А. Легалов a, b, *, В. Ю. Назаренко c, **, Е. Э. Перковский c, ***
a Институт систематики и экологии животных СО РАН
Новосибирск, Россия
b Алтайский государственный университет
Барнаул, Россия
c Институт зоологии НАН Украины
Киев, Украина
* E-mail: fossilweevils@gmail.com
** E-mail: nazarenko@izan.kiev.ua
*** E-mail:
Поступила в редакцию 19.05.2017
Принята к публикации 07.07.2017
Аннотация
Из мексиканского янтаря описан новый вид Homocloeus popovi sp. nov., сходный с H. puncticollis и отличающийся головотрубкой без срединного киля, сильнее изогнутым к основанию надкрылий поперечным килем, уплощенными промежутками надкрылий и 5-м промежутком с бугорком перед скатом надкрылий. Это первая находка представителя рода Homocloeus в ископаемом состоянии.
Ложнослоники – семейство долгоносикообразных жуков, представленное в современной фауне более чем 3800 видами (Rheinheimer, 2004). Первые находки вымерших подсемейств семейства Anthribidae сделаны в средней-верхней юре Каратау (Легалов, 2011, 2013). Самый древний представитель подсемейства Anthribinae описан из отложений середины мела Хетаны в России (Жерихин, 1993; Головнева, 2013), а Choraginae – из берриаса–баррема Монтсека в Испании (Soriano et al., 2006). В эоценовых и миоценовых янтарях ложнослоники хорошо представлены (Грачев, Перковский, 2008; Legalov, 2015; Poinar, Legalov, 2017; Legalov et al., 2018).
Фауна долгоносикообразных жуков миоценовых янтарей Нового Света выявлена довольно подробно. При этом основное число видов описано из доминиканского янтаря (Legalov, 2015; Poinar et al., 2016; Poinar, Legalov, 2017). Мексиканский янтарь добывался в значительно меньшем объеме и слабее исследован; из него известны представитель рода Eugnamptus Schoenherr, 1839 из семейства Rhynchitidae и пять видов семейства Curculionidae из родов Semnorhynchus Faust, 1896, Neoulosomus O’Brien et Wibmer, 1982, Caulophilus Wollaston, 1854, Geratozygops Davis et Engel, 2006 и Anthonomus Germar, 1817 (Zimmermann, 1971; Poinar, Brown, 2007; Poinar, Legalov, 2014a, b; Legalov, Poinar, 2015; Barrios-Izás, Coty, 2016). Ложнослоники описаны также из мексиканского янтаря (Poinar, Legalov, 2016) – это вид рода Euparius Schoenherr, 1823 трибы Cratoparini и пять видов рода Piesocorynus Dejean, 1834 трибы Piesocorynini подсемейства Anthribinae. В представленной работе описывается новый вид рода Homocloeus Jordan, 1904, ранее не отмеченного в палеонтологической летописи.
Экземпляр происходит из янтароносных слоев на севере нагорья Чьяпас в районе Симоховеля в штате Чьяпас (юго-восток Мексики). Карты янтароносной области представлены у Г. Пойнара (Poinar, 1992) и М. Солорзано-Крамер (Solórzano Kraemer, 2010). Янтарь, вероятно, продуцировался Hymenaea mexicana из семейства бобовых (Poinar, Brown, 2002). Мексиканский янтарь ассоциируется со свитами Ла Кинта, Балумтун Сандстон и Мазантик Шайл и датируется нижним-средним миоценом с возрастом от 15 до 20 млн лет (Serrano-Sanchez et al., 2015).
Голотип хранится в коллекции Национального музея Шотландии (NMS) в Эдинбурге.
СЕМЕЙСТВО ANTHRIBIDAE BILLBERG, 1820
ПОДСЕМЕЙСТВО ANTHRIBINAE BILLBERG, 1820
Триба Zygaenodini Lacordaire, 1866
Род Homocloeus Jordan, 1904
Homocloeus popovi Legalov, Nazarenko et Perkovsky, sp. nov.
Табл. VIII, фиг. 1 (см. вклейку)
Anthribidae: Ross et al., 2016, с. 50, рис. 14 .
Название вида в память Ю.А. Попова, известного российского палеоэнтомолога.
Голотип – самка, NMS G.2004.6.8.1; мексиканский янтарь; нижний-средний миоцен.
Описание (рис. 1). Тело черное, покрытое полуприлегающими светлыми густыми волосками. Рисунок дорсальной поверхности тела пятнистый, неясный, имеются продольные боковые темные полосы на диске переднеспинки и темные пятна на середине диска; более темное основание надкрылий, за ним в передней трети нерезкая светлая поперечная перевязь, за которой приблизительно в середине и у вершины надкрылий преобладают темные пятна, между ними в задней трети и у вершины светлые пятна образуют малозаметные поперечные перевязи. Волоски преимущественно суженные к вершине или параллельносторонние, в местах сочленений сегментов и их склеритов и, частично, на надкрыльях они светлее и заметно расширены. Поперечное расстояние между соседними волосками обычно в 1–3 раза превышает их ширину, в продольном направлении волоски могут перекрываться, вследствие чего образуют на кутикуле спутанные ряды. В местах соединений сегментов, их склеритов и т.п. волоски часто сгущены и образуют более или менее выраженные щеточки или бахромки. Головотрубка короткая, немного длиннее головы, без килей, с крупными мандибулами. Длина головотрубки составляет 0.8 ширины на вершине и основании, 0.9 ширины на середине и 0.5 длины переднеспинки. Мандибулы крупные, с тупым зубцом на первой трети внутреннего края. Челюстные щупики длинные, 4-члениковые. Верхняя губа свободная, почти квадратная. Глаза округлые, без выемки на переднем крае, мелкофасеточные, не сближенные на основании лба. В поперечном ряду около 35 фасеток. Длина глаза в 1.1–1.3 раза больше ширины. Лоб широкий, уплощенный, его ширина составляет 0.8 ширины головотрубки у основания. Виски слабо удлиненные, их длина составляет 0.7 длины глаза. Темя мелко и густо пунктированное, слабовыпуклое. Усиковые бороздки ямковидные. Усики не коленчатые, длинные, достигают базальной трети переднеспинки, прикреплены возле середины головотрубки, латерально. Стволик овальный. Членики жгутика усиков конические. Булава хорошо выраженная, трехчлениковая, не компактная, уплощенная. Вершина ее последнего членика асимметричная, хорошо обособленная. Длина 1‑го членика усиков в 1.8 раза больше его ширины. Длина 2-го членика в 3.8 раза больше его ширины, в 1.7 раза больше длины и ширина составляет 0.8 ширины 1‑го членика. Длина 3-го членика в 4.0 раза больше его ширины, в 1.1 раза больше длины и ширина равна ширине 2-го членика. Длина 4-го членика в 4.4 раза больше его ширины, равна длине и ширина составляет 0.9 ширины 3-го членика. Длина 5-го членика в 2.4 раза больше его ширины, составляет 0.6 длины и в 1.1 раза больше ширины 4-го членика. Длина 7-го членика в 2.0 раза больше его ширины. Длина 8-го членика в 2.2 раза больше его ширины, в 1.1 раза больше длины и его ширина равна ширине 7‑го членика. Длина 9‑го членика в 1.5 раза больше его ширины, в 2.3 раза больше длины и его ширина в 3.3 раза больше ширины 8-го членика. Длина 10‑го членика составляет 0.6 его ширины, составляет 0.5 длины и 1.1 ширины 9-го членика. Длина 11‑го членика в 1.3 раза больше его ширины, в 1.7 раза больше длины 10-го членика и составляет 0.9 его ширины. Переднеспинка почти колоколовидная, без бугорков. Длина переднеспинки в 2.1 раза больше длины головотрубки. Диск переднеспинки густо точечный, промежутки пунктировки узкие, в 2–5 раз меньше диаметра точек, образуют спутанную крупноячеистую скульптуру. Суббазальный киль немного удален от основания переднеспинки, слабо изогнутый к основанию надкрылий, по бокам переходит в латеральный. Расстояние между суббазальным килем и основанием переднеспинки составляет 0.04 расстояния между ним и вершиной переднеспинки. Латеральный киль явственный, расположен в базальной и серединной частях переднегруди. В апикальной трети переднегруди он становится морщинковидным, исчезая в ее передней четверти. Надкрылья слабовыпуклые, с довольно широкими почти плоскими промежутками и слабыми бороздками, без крупных бугорков на промежутках. Длина надкрылий в 1.8, брошка в 2.1 раза больше длины переднеспинки. Вершины надкрылий закруглены раздельно. 5-й промежуток с небольшим бугорком перед скатом надкрылий. Переднегрудь пунктированная. Длина прекоксальной части переднегруди равна длине передних тазиковых впадин; посткоксальной – 0.5 длины передних тазиковых впадин. Передние тазиковые впадины округленные. Средние тазиковые впадины разделенные. Заднегрудь довольно длинная, ее длина в 1.8 раза больше длины задних тазиковых впадин. Метэпистерны широкие, их длина в 2.4 раза больше ширины на середине. Вентриты выпуклые. Длина 1-го вентрита составляет 0.7 длины заднего тазика. Длина 2-го вентрита в 1.2 раза больше длины 1-го вентрита. Длина 3-го вентрита равна длине 2-го вентрита. Длина 4-го вентрита немного больше длины 3-го вентрита. Длина 5-го вентрита в 2.0 раза больше длины 4-го вентрита. Пигидий большой, слабовыпуклый. На его основании заметно слабое треугольное вдавление, ограниченное килями. Анальный клапан полуоткрыт. Вершины кокситов и стилей зазубрены, отогнуты наружу, вместе с вершиной яйцеклада частично выступают за пределы брюшка. Ноги длинные. Бедра утолщенные и немного уплощенные. Длина передних бедер в 2.5 раза больше ширины. Длина задних бедер в 2.1 раза больше ширины. Голени почти прямые, едва расширенные к вершине, без мукро и шпор. Вершины голеней с венчиком шипиков, среди которых с внутренней стороны левой средней и правой задней голеней заметны по 2 более крупных шипа. Длина передних голеней в 5 раз больше ширины. Длина задних голеней в 4.3 раза больше ширины. Лапки длинные, 1-й членик удлиненный. 2-й членик конический, угловидно-вырезанный на вершине и охватывающий 3-й членик. 3-й членик двухлопастный. 5-й членик удлиненный. Коготки с зубцами, они примерно вдвое короче коготков. Длина 2-го членика передних лапок составляет 0.7 ширины на вершине, его ширина в 1.6 раза больше ширины 1-го членика. Длина 3‑го членика передних лапок составляет 0.9 ширины на вершине, его длина составляет 0.8 длины и его ширина составляет 0.6 ширины 2-го членика. Длина 5-го членика передних лапок в 2.2 раза больше ширины на вершине, его длина в 2.2 раза больше длины и его ширина составляет 0.9 ширины 3-го членика. Длина 1-го членика средних лапок в 2.2 раза больше ширины на вершине. Длина 2-го членика средних лапок составляет 0.9 ширины на вершине, его длина составляет 0.5 длины и его ширина в 1.4 раза больше ширины 1-го членика. Длина 3-го членика средних лапок составляет 0.8 ширины на вершине, его длина составляет 0.7 длины и его ширина составляет 0.7 ширины 2‑го членика. Длина 5-го членика средних лапок в 2.6 раза больше ширины на вершине, его длина в 2.3 раза больше длины и его ширина составляет 0.7 ширины 3-го членика. Длина 1-го членика задних лапок в 3.5 раза больше ширины на вершине. Длина 2-го членика задних лапок в 2.0 раза больше ширины на вершине, его длина составляет 0.6 длины и его ширина в 1.1 раза больше ширины 1-го членика. Длина 3-го членика задних лапок почти равна ширине на вершине, его длина составляет 0.3 длины и его ширина составляет 0.6 ширины 2-го членика. Длина 5-го членика задних лапок в 3.4 раза больше ширины на вершине, его длина в 3.0 раза больше длины и его ширина составляет 0.9 ширины 3-го членика.
Рис. 1.
Homocloeus popovi sp. nov., голотип NMS G.2004.6.8.1: а – голова, дорсо-латерально; б – переднеспинка, латерально; в – голова, латерально; г – общий вид, латерально. Длина масштабного отрезка на фиг. а–в соответствует 0.5 мм, на фиг. г – 1 мм.
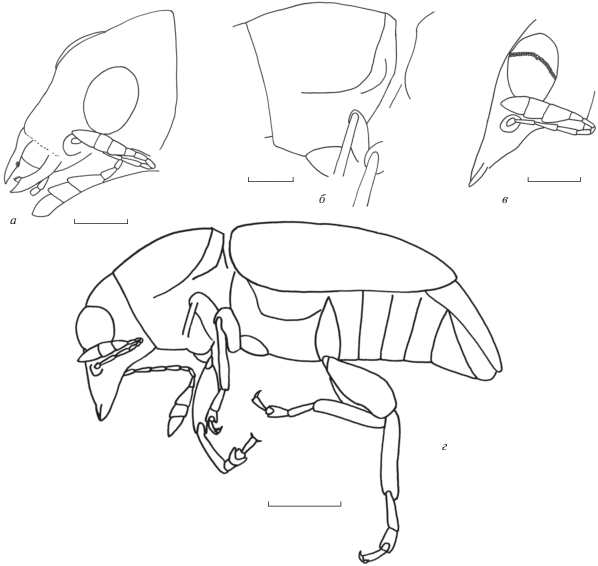
Размеры в мм: длина тела без головотрубки – 5.9; длина головотрубки – 0.7. Длина левого глаза 1.2, ширина 1.0. Длина волосков дорсальной поверхности тела около 0.024–0.03, ширина – 0.002–0.004.
Сравнение. Новый вид сходен с H. puncticollis Jordan, 1906 из Мексики из-за переднеспинки и надкрыльев без бугорков, и отличается головотрубкой без серединного киля, сильнее изогнутым к основанию надкрылий поперечным килем, уплощенными промежутками надкрылий и 5‑м промежутком с бугорком перед скатом надкрылий.
Замечания. Короткая головотрубка с крупными мандибулами, суббазальный киль на переднеспинке, свободная верхняя губа, явственный пигидий и не коленчатые усики указывают на принадлежность нового вида к семейству Anthribidae. Латерально прикрепленные усики и 2-й членик лапок, угловидно вырезанный на вершине и охватывающий 3-й членик, позволяют поместить этот вид в подсемейство Anthribinae. Новый вид относится к трибе Zygaenodini, поскольку характеризуется суббазальным килем на переднеспинке, отодвинутым от ее основания, ямковидными усиковыми бороздками, 2-м члеников усиков значительно длиннее, чем 1-й членик, головотрубкой с длиной меньше, чем ее ширина и мелкофасеточными глазами. Округлые глаза, не сближенные на основании лба, указывают на принадлежность к роду Homocloeus.
Род Homocloeus наиболее разнообразен на Больших Антильских островах, представлен в современной фауне 15 описанными видами, распространенными в равнинных тропических лесах от Центральной Мексики до севера Бразилии (Пара, Эспириту-Санту). Из Мексики известен единственный вид, описанный из Туспана (Jordan, 1906), в то время как для о-ва Гаити указано 13 неописанных видов (Valentine, 2003). Отсутствие сведений об экологии этого рода не позволяет объяснить его отсутствие в доминиканском янтаре. Однако следует отметить, что мексиканский янтарный лес характеризуется как низинный лес вблизи мангровых зарослей, а доминиканский янтарный лес – как низинный, более удаленный от моря тропический лес (Solórzano Kraemer et al., 2015).
Материал. Голотип.
Авторы выражают благодарность Эндрю Россу (National Museums Scotland, Edinburgh, UK) за предоставление материала для исследования и Д.Е. Щербакову (ПИН) за ценные замечания.
Список литературы
Головнева Л.Б. Новые данные о позднемеловых флорах Ульинского прогиба (Западное Приохотье) // Палеоботаника. 2013. Т. 4. С. 148–167.
Грачев В.Г., Перковский Е.Э. Новый вид рода Glaesotropis (Insecta: Coleoptera: Anthribidae) из ровенского янтаря // Палеонтол. журн. 2008. № 1. С. 62–64.
Жерихин В.В. Подотряд Polyphaga // Мезозойские насекомые и остракоды Азии. М.: Наука, 1993. С. 20–37 (Тр. Палеонтол. ин-та РАН. Т. 252).
Легалов А.А. Первая находка жуков-ложнослоников (Coleoptera: Anthribidae) в юре Казахстана // Палеонтол. журн. 2011. № 6. С. 34–37.
Легалов А.А. Обзор семейства Anthribidae (Coleoptera) из юры Каратау. Подсемейство Protoscelinae. Род Protoscelis Medvedev, 1968 // Палеонтол. журн. 2013. № 3. С. 57–65.
Barrios-Izás M., Coty D. A new fossil species of Caulophilus Wollaston, 1854 (Coleoptera: Curculionidae: Cossoninae) from Mexican amber // Col. Bull. 2016. V. 70. № 1. P. 177–179.
Jordan K. Insecta. Coleoptera. Rhynchophora. Anthribidae // Biologia Centrali-Americana. 1906. V. 4. Pt 6. P. 299–314.
Legalov A.A. Fossil Mesozoic and Cenozoic weevils (Coleoptera, Obrienioidea, Curculionoidea) // Paleontol. J. 2015. V. 49. № 13. P. 1442–1513.
Legalov A.A., Poinar G.O.Jr. A new species of the genus Anthonomus Germar, 1817 (Coleoptera: Curculionidae) in Mexican amber // Paleontol. J. 2016. V. 50. № 9. P. 986–990.
Legalov A.A., Nazarenko V.Y., Perkovsky E.E. A new genus of fungus weevils (Coleoptera: Anthribidae) in Rovno amber // Fossil Record. 2018. V. 21. P. 207–212.
Poinar G.O.Jr. Life in amber. Stanford, CA: Stanford Univ. Press, 1992. 368 p.
Poinar G.O. Jr., Brown A.E. Hymenaea mexicana sp. nov. (Leguminosae: Caesalpinioideae) from Mexican amber indicates Old World connections // Bot. J. Linn. Soc. 2002. V. 139. P. 125–132.
Poinar G.O. Jr., Brown A.E. Eugnamptus proterus sp. n. (Coleoptera: Curculionoidea: Rhynchitidae), a tooth-nosed snout beetle in Mexican amber // Proc. Entomol. Soc. Wash. 2007. V. 109. № 4. P. 880–885.
Poinar G.O.Jr., Brown A.E., Legalov A.A. First record of the genus Pseudopilolabus Legalov, 2003 (Coleoptera: Attelabidae) in Dominican amber // Fossil Record. 2016. V. 19. P. 11–16.
Poinar G.O.Jr., Legalov A.A. New Cryptorhynchinae (Coleoptera: Curculionidae) in Dominican amber // Hist. Biol. 2014a. V. 26. № 4. P. 502–534.
Poinar G.O.Jr., Legalov A.A. New species of the subfamily Conoderinae (Coleoptera: Curculionidae) in Dominican amber // Hist. Biol. 2014b. V. 26. № 5. P. 556–562.
Poinar G.Jr., Legalov A.A. New Anthribidae (Coleoptera: Curculionoidea) in Dominican and Mexican ambers // Palaeontol. Electron. 2016. V. 19. Iss. 2. № 24A. P. 1–38.
Poinar G.Jr., Legalov A.A. Five new species from the subfamily Entiminae (Coleoptera: Curculionidae) in Dominican amber // Palaeontol. Electron. 2017. V. 20. Iss. 2. № 23A. P. 1–13.
Rheinheimer J. Illustrierter Katalog und Bibliographie der Anthribidae der Welt (Insecta: Coleoptera) // Mitt. Entomol. Ver. Stuttgart. 2004. Bd 39. H. 1–2. 288 s.
Ross A.J., Mellish C.J.T., Crighton B., York P.V. A catalogue of the collections of Mexican amber at the Natural History Museum, London and National Museums Scotland, Edinburgh, UK // Bol. Soc. Geol. Mexicana. 2016. V. 68. № 1. P. 45–55.
Serrano-Sanchez M. de L., Hegna T.A., Schaaf P. et al. The aquatic and semiaquatic biota in Miocene amber from the Campo La Granja mine (Chiapas, Mexico): Paleoenvironmental implications // J. South Amer. Earth Sci. 2015. V. 62. P. 243–256.
Solórzano Kraemer M.M. Mexican amber // Biodiversity of fossils in amber from the major world deposits / Ed. Penney D. Manchester: Siri Sci. Press, 2010. P. 42–56.
Solórzano Kraemer M.M., Kraemer A.S., Stebner F. et al. Entrapment bias of arthropods in Miocene amber revealed by trapping experiments in a tropical forest in Chiapas, Mexico // PLoS ONE. 2015. V. 10. № 3. e0118820. doi 10.1371/journal.pone.0118820
Soriano C., Gratshev V.G., Delclos X. New Early Cretaceous weevils (Insecta, Coleoptera, Curculionoidea) from El Montsec, Spain // Cret. Res. 2006. V. 27. P. 555–564.
Valentine B.D. A catalogue of West Indies Anthribidae (Coleoptera) // Insecta Mundi. 2003. V. 17. P. 49–67.
Zimmermann E.C. Mexican Miocene amber weevils (Insecta: Coleoptera: Curculionidae) // Univ. Calif. Publ. Entomol. 1971. V. 63. № 2. P. 103–106.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал