

Палеонтологический журнал, 2019, № 5, стр. 90-99
Морская черепаха (Сheloniidae) из среднего эоцена Северного Кавказа
Е. А. Звонок a, *, Е. В. Сыромятникова b, c, И. Г. Данилов c, А. Ф. Банников b
a Луганский национальный университет имени Тараса Шевченко
Луганск, Украина
b Палеонтологический институт им. А. А. Борисяка РАН
Москва, Россия
c Зоологический институт РАН
Санкт-Петербург, Россия
* E-mail: aban@paleo.ru
Поступила в редакцию 18.12.2018
После доработки 15.02.2019
Принята к публикации 21.02.2019
Аннотация
В статье описан неполный скелет ювенильной особи морской черепахи (Cheloniidae) из терминального бартона (конец среднего эоцена) местонахождения Горный Луч (Краснодарский край, Россия), определенный как Eochelone sp. Новая находка расширяет представления о географическом распространении черепах рода Eochelone Dollo, 1903, которые ранее были известны из лютета Бельгии и Украины, бартона Англии и приабона Испании в Европе. Кроме того, описанный экземпляр имеет общие признаки с “Allopleuron” qazaqstanense Karl et al., 2012 из бартона и, возможно, лютета Казахстана.
Ископаемые остатки эоценовых морских черепах (Chelonioidea: Cheloniidae) Восточного Тетиса известны преимущественно по разрозненным костям, реже – по небольшим ассоциациям и блокам костей скелета (см. Данилов и др., 2017). Сопоставимо полных по числу костей экземпляров эоценовых хелониид Восточного Тетиса описано всего три: 1) череп Puppigerus nessovi Averianov, 2005 (местонахождение Иково, Луганская обл., Украина; нижний лютет; Звонок, 2013); 2) большая часть скелета (голотип) “Allopleuron” qazaqstanense Karl et al., 2012 (неизвестное местонахождение, Казахстан, лютет?; Karl et al., 2012; Звонок и др., 2015); 3) часть скелета Argillochelys antiqua (Konig, 1825) (местонахождение Вышгород, Киевская обл., Украина; киевская свита, верхний лютет – нижний бартон; Averianov, 2002; Zvonok, Danilov, 2017). Кроме того, известен панцирь с рядом других элементов скелета пока не описанного экземпляра, предварительно отнесенного к Argillochelys sp. из бартона Казахстана (Звонок и др., 2011; Данилов и др., 2017).
В 2001 г. одним из авторов (А.Ф.Б.) в верхней части кумского горизонта (терминальный бартон) на левом берегу р. Пшеха, в 1 км выше хутора Горный Луч (Апшеронский р-н, Краснодарский край, Россия; рис. 1) был найден частично сочлененный скелет ювенильной особи морской черепахи, который из перечисленных экземпляров по полноте, вероятно, уступает только голотипу “Allopleuron” qazaqstanense. Настоящая статья посвящена описанию этого скелета, который определяется как Eochelone sp.
Помимо черепах, фауна местонахождения Горный Луч включает в себя богатый комплекс морских рыб: 49 таксонов видового ранга костистых рыб и 11 – хрящевых рыб, а также стрижеобразную птицу Argornis caucasicus Karhu, 1999 (Karhu, 1999; Банников, 2010, 2017, 2018; Малышкина и др., 2013).
Изученный экземпляр морской черепахи хранится в Палеонтологическом ин-те им. А.А. Борисяка РАН (ПИН), Москва. Другие материалы, упоминаемые в статье, хранятся в следующих учреждениях: MGSB – Геологический музей Барселонской семинарии, Барселона; ZIN PH – Зоологический ин-т РАН, палеогерпетологическая коллекция, С.-Петербург; IRSNB – Бельгийский королевский музей естественных наук, Брюссель; МРИМ – Мангистауский региональный исторический музей, Актау; NHM – Музей естественной истории, Лондон; RM – Музей Ринополис, Ганна; NMNHU-P – Национальный научно-природоведческий музей НАН Украины, отдел палеозоологии, Киев.
Для сравнения использовались данные по следующим эоценовым морским черепахам: “Allopleuron” qazaqstanense Karl et al., 2012 (Karl et al., 2012; Звонок и др., 2015; фотографии голотипа RM, без номера); “Argillochelys” africana Tong et Hirayama, 2008 (Tong, Hirayama, 2008); A. antiqua (Owen, 1849) (Owen, Bell, 1849; Zvonok, Danilov, 2017; фотографии экз. IRSNB, № 1653 и NHM, № 38952); A. cuneiceps (Owen, 1849) (Owen, Bell, 1849; фотографии голотипа NHM, № 41636 и экз. NHM, № 38949); Argillochelys sp. (Звонок, Данилов, 2019); Eochelone athersuchi Moody, 1980 (Moody, 1980); E. brabantica Dollo, 1903 (Casier, 1968; Lapparent de Broin et al., 2018); E. monstigris Grant-Mackie et al., 2011 (Grant-Mackie et al., 2011); E. voltregana Lapparent de Broin et al., 2018 (Lapparent de Broin et al., 2018); Erquelinnesia gosseleti (Dollo, 1886) (Zangerl, 1971); Euclastes sp. (Weems, Brown, 2017); Osonachelus decorata Lapparent de Broin et al., 2014 (Lapparent de Broin et al., 2014); Puppigerus camperi (Gray, 1831) (Moody, 1974; Tong et al., 2012) и Puppigerus nessovi Averianov, 2005 (Аверьянов, 2005; Звонок, 2013; Звонок и др., 2013).
В статье применена анатомическая терминология Р. Цангерла и др. (Zangerl et al., 1988; для щитков черепа), Ю. Гэффни (Gaffney, 1979; для костей черепа), И.Г. Данилова с соавт. (2017; для элементов панциря).
Авторы благодарны Г.-В. Карлу (Геттингенский ун-т, Германия) за предоставление фотографий голотипа “Allopleuron” qazaqstanense, Р. Хираяме (Ун-т Васеда, Токио) за фотографии экземпляров Argillochelys spp. и А.О. Аверьянову за ценные замечания к рукописи.
ОПИСАНИЕ МАТЕРИАЛА
Материал по Eochelone sp. (экз. ПИН, № 5136/1) представлен скелетом ювенильной особи морской черепахи, сохранившимся в виде разрозненных костей, костей в сочленении или их отпечатков в блоке породы. При разделении этого блока на две части наблюдаются две поверхности, на каждой из которых видны складывающиеся части или отпечатки одних и тех же костей (рис. 2, 3).
Рис. 2.
Остатки Eochelone sp. (экз. ПИН, № 5136/1) из среднего эоцена местонахождения Горный Луч (Краснодарский край; Россия) на поверхности первой части блока породы: а – внешний вид, б – схематический рисунок. Обозначения: CE – I центральный щиток; сo – костальные пластинки; сr – блок костей крыши черепа; ep – эпипластрон; hu – плечевая кость; hyo – гиопластрон; hyp – гипопластрон; k – гребни гио- и гипопластрона; mx – верхнечелюстная кость; ne – невральные пластинки; nu – нухальная пластинка; PC – прецентральный щиток; pe – периферальные пластинки; sc – лопатка; su – I супрапигальная пластинка; ve – шейный позвонок; xi – ксифипластрон. Сплошной линией обозначены края костей, штриховой линией – борозды роговых щитков, серым цветом – поверхность костей, штрихом – сломы костей, без заливки – отпечатки костей.
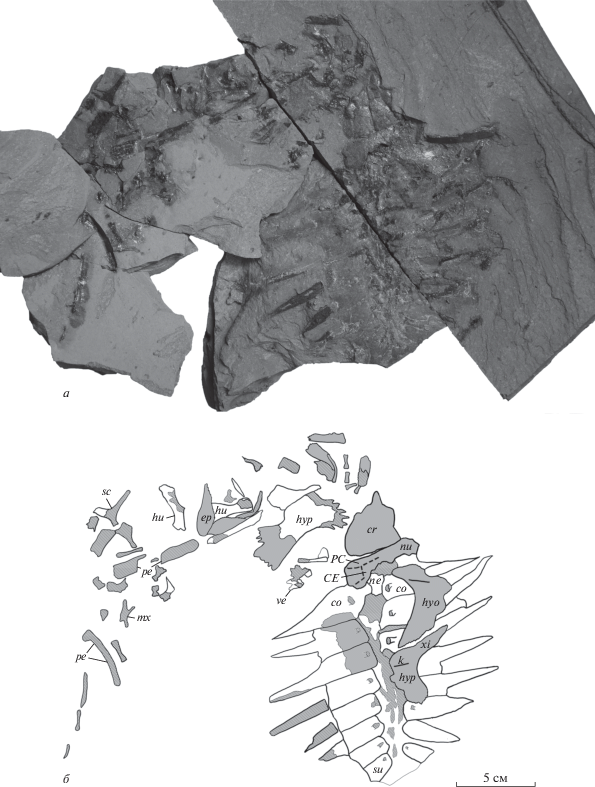
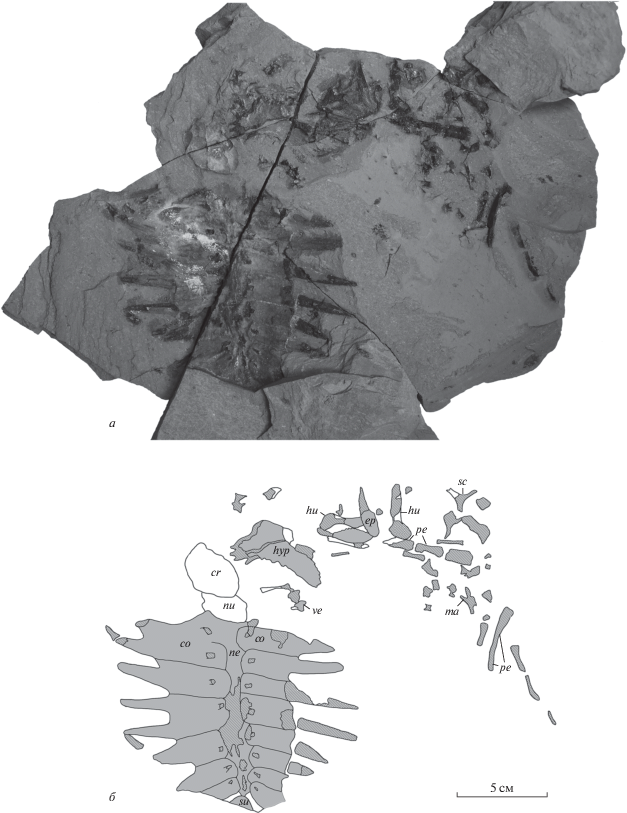
Рис. 3.
Остатки Eochelone sp. (экз. ПИН, № 5136/1) из среднего эоцена местонахождения Горный Луч (Краснодарский край; Россия) на поверхности второй части блока породы: а – внешний вид, б – схематический рисунок. Обозначения см. в подписях к рис. 2.
На поверхности первой части блока (рис. 2) видна наружная поверхность крыши черепа и расположенный позади нее карапакс, представленный фрагментом нухальной пластинки (виден снаружи) и другими элементами в виде отпечатков и поврежденных костей (полный ряд невральных пластинок, передняя часть I супрапигальной пластинки и восемь пар костальных пластинок). В области отпечатка карапакса в неестественном положении вентральной поверхностью вверх лежат правые гиопластрон и сочлененные гипо- и ксифипластрон. Слева и частично спереди от описанной части скелета располагается беспорядочное скопление других костей, часть из которых повреждена или представлена отпечатками. В стороне от основной массы костей также сохранился частично разрушенный симфиз зубных костей, который был распилен для изучения деталей строения альвеолярной поверхности.
На поверхности второй части блока (рис. 3) видны отпечатки крыши черепа и нухальной пластинки, частично поврежденная вентральная поверхность карапакса и такое же, как и на первом блоке, скопление костей.
Наружная поверхность костей крыши черепа и панциря имеет орнаментацию в виде тонких гребней, известную также у протостегид рода Rhinochelys Seeley, 1869 и некоторых меловых и палеогеновых хелониид (Lapparent de Broin et al., 2014, 2018). Глубокой орнаментации в виде сети борозд, как у Osonachelus decorata, не наблюдается.
На поверхности крыши черепа видны очертания следующих роговых щитков: правого и частично левого предлобных, лобного, лобно-теменного, обоих теменных, межтеменного, обоих надглазничных, височных и левого бокового височного (рис. 4). Предлобных щитков одна пара, они занимают антеролатеральную часть лобных и всю поверхность предлобных костей. Лобный щиток занимает медиальные части лобных костей и небольшую (антеромедиальную) часть теменных. Лобно-теменной щиток относительно крупный, цельный (непарный), расположен в медиальной части теменных костей, постепенно сужаясь к их заднему краю, подобно “Allopleuron” qazaqstanense, Argillochelys antiqua и Eochelone brabantica. Дополнительный щиток, вклинивающийся между парой лобно-теменных щитков сзади у “Argillochelys” africana и Erquelinnesia gosseleti, у экз. ПИН, № 5136/1 отсутствует. Теменных щитков, расположенных в задней части теменных костей, три (включая межтеменной), как у большинства других эоценовых хелониид, за исключением Argillochelys cuneiceps и Osonachelus decorata, имеющих один теменной щиток. Надглазничные щитки, расположенные в постеролатеральных частях лобных, антеролатеральных частях теменных и передних частях заглазничных костей, крупные. Височные щитки, видимые в постеролатеральной части теменных и постеромедиальной части левой заглазничной костей, и левый боковой височный щиток, часть которого видна в латеральной части левой заглазничной кости, крупные и не разделены.
Рис. 4.
Дорсальная поверхность крыши черепа Eochelone sp. (экз. ПИН, № 5136/1) из среднего эоцена местонахождения Горный Луч (Краснодарский край; Россия): а – внешний вид, б – схематический рисунок. Обозначения: FP – лобно-теменной щиток; FR – лобный щиток; fr – лобная кость; IP – межтеменной щиток; LT – боковой височный щиток; PA – теменной щиток; pa – теменная кость; PF – предлобный щиток; pf – предлобная кость; po – заглазничная кость; SO – надглазничный щиток; TE – височный щиток.

Предлобная кость (сохранилась только правая; рис. 4) узкая, сочленяется с лобной костью сзади (остальные контакты не видны), формирует дорсальные края наружного носового отверстия и глазницы.
Лобные кости (имеются обе; рис. 4) поперечно вытянутые, их медиальные края, контактирующие друг с другом, длиннее латеральных, так как спереди лобные кости вклиниваются между предлобными. Контакт с предлобной костью вогнутый. Антеролатерально (слева сильнее, чем справа) лобные кости выходят в края глазниц (в отличие от “Argillochelys” africana, у которых лобные кости не выходят в края глазниц). Постеролатерально, вогнутым краем лобная кость контактирует с заглазничной костью, а сзади имеет прямой или слегка вогнутый контакт с теменной костью.
Теменные кости (имеются обе; рис. 4) без глубоких вырезок в постеролатеральных частях для чешуйчатых костей (в отличие от Puppigerus camperi), контактируют по средней линии друг с другом, сочленяются с лобными костями спереди и с заглазничными костями латерально; остальные контакты не видны.
Заглазничные кости (левая – целая, правая в виде двух фрагментов; рис. 4) формируют задние края глазниц, сочленяются с лобными костями антеромедиально и с теменными костями медиально; остальные контакты не видны.
Верхнечелюстная кость (сохранилась только левая) расщеплена продольно на две части – медиальную и латеральную (рис. 2, 3, 5, а). Судя по вертикальному переднему краю предлобного отростка, наружное носовое отверстие было ориентировано почти вертикально. Лабиальный гребень, видимо, был высоким. Видны сочленовные поверхности с предлобной и предчелюстной костями в дорсальной и передней частях предлобного отростка соответственно.
Рис. 5.
Отдельные элементы скелета Eochelone spp.: а – медиальная сторона левой верхнечелюстной кости Eochelone sp. (экз. ПИН, № 5136/1); б–г – эпипластроны: б – E. voltregana (экз. MGSB, № M-4; Lapparent de Broin et al., 2018, рис. 6, c1), в – Eochelone sp. (экз. ПИН, № 5136/1), г – E. brabantica (экз. IRSNB, № R61; Lapparent de Broin et al., 2018, рис. 9 , c2); д–ж – гиопластроны: д – E. voltregana (экз. MGSB, № 74.642. G.L.V., T-46; Lapparent de Broin et al., 2018, рис. 6, c1), е – Eochelone sp. (экз. ПИН, № 5136/1), ж – E. brabantica (экз. IRSNB, № R61; Lapparent de Broin et al., 2018, рис. 9 , c2); з – лопатка Eochelone sp. (экз. ПИН, № 5136/1); и, к – плечевые кости Eochelone sp. (экз. ПИН, № 5136/1): и – вентральная сторона левой плечевой кости, к – дорсальная сторона правой плечевой кости. Обозначения: en – эктэпикондилярная вырезка; gl – гленоидная шейка; lp – латеральный отросток; lr – лабиальный гребень; mp – медиальный отросток; pfs – шов с предлобной костью; pms – шов с предчелюстной костью; pp – предлобный отросток; pt – ямка для крепления сухожилий mm. latissimus dorsi и teres major. На рис. 5, а сплошной линией обозначен край задней части лабиального гребня верхнечелюстной кости, слабо видимый на фотографии, штриховой линией – предполагаемое положение края передней части лабиального гребня верхнечелюстной кости; на рис. 5, д–ж сплошной линией обозначен край центральной фонтанели.
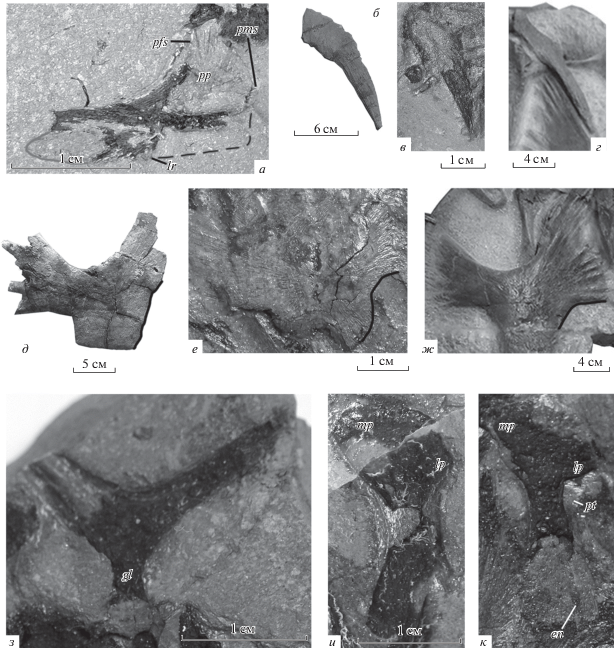
Зубные кости (сохранился симфиз зубных костей, левая кость сильно повреждена) имеют длинные ветви (рис. 6). Симфиз короткий, лабиальные гребни высокие, в отличие от Puppigerus spp., Erquelinnesia gosseleti и Euclastes sp., у которых лабиальных гребней нет. Альвеолярная поверхность около симфиза в поперечном сечении слабовогнутая, без лингвального гребня, как у “Allopleuron” qazaqstanense и Eochelone brabantica, и в отличие от Argillochelys spp., имеющих развитые лингвальные гребни.
Рис. 6.
Поперечное сечение зубной кости Eochelone sp. (экз. ПИН, № 5136/1) из среднего эоцена местонахождения Горный Луч (Краснодарский край; Россия). Дорсальный край лабиального гребня слегка обломан. Штриховой линией обозначены реконструированные части зубной кости, скрытые в горной породе.

Нухальная пластинка представлена передней и левой частями (рис. 2). Передний край пластинки почти прямой. Наличие/отсутствие постнухальных фонтанелей установить невозможно из-за поврежденности задней части кости. На наружной поверхности пластинки видны три борозды, передняя из которых интерпретируется как задний край прецентрального щитка, средняя – как латеральный край I центрального щитка и задняя – как передний край I плеврального щитка или часть латерального края I центрального щитка. Прецентральный щиток почти такой же ширины, как нухальная пластинка. I центральный щиток узкий, по крайней мере спереди.
Невральные пластинки представлены полным (непрерывным) рядом, однако их точное число неясно (рис. 2, 3). Невральные пластинки, расположенные между III–VI костальными пластинками, имеют шестиугольную форму с короткими передне-боковыми сторонами. Невральная пластинка, расположенная между VI и VII костальными пластинками, шестиугольная с примерно равными передне- и задне-боковыми сторонами. Форма других невральных пластинок отчетливо не видна.
Супрапигальные пластинки представлены только передней частью I супрапигальной пластинки, имевшей, по-видимому, трапециевидную форму (рис. 3).
Костальные пластинки представлены полным комплектом – восемью парами (рис. 2, 3). Все костальные пластинки, кроме правых VII и VIII и левой VIII, у которых латеральные части не сохранились, имеют длинные свободные ребра, которые у некоторых пластинок (II–IV) по длине почти равны длине остальной части (тела) пластинки.
Периферальные пластинки сохранились в числе не менее шести элементов (рис. 2, 3). Пластинки узкие латеромедиально, что, по-видимому, связано с ювенильным возрастом особи.
Эпипластроны представлены одним (правым или левым) элементом (рис. 2, 3, 5, в). Гулярный выступ эпипластрона широкий и округленный, как у Argillochelys antiqua и Eochelone voltregana.
Гиопластроны представлены правым элементом, который виден с вентральной стороны, что определяется по наличию продольного киля (рис. 2, 5, е). Вырезки от латеральных и центральной фонтанелей большие. При реконструкции медиальные края гио- и гипопластронов формировали вытянутую в переднем направлении центральную фонтанель, которая была длиннее латеральных. Данная фонтанель была шире, чем контакты гио- и гипопластрона или латеральные фонтанели по отдельности (рис. 7).
Рис. 7.
Реконструкция пластрона Eochelone sp. (экз. ПИН, № 5136/1) из среднего эоцена местонахождения Горный Луч (Краснодарский край; Россия). Обозначения: CE – центральная фонтанель; en – энтопластрон; epi – эпипластрон; hyo – гиопластрон; hyp – гипопластрон; LA – латеральная фонтанель; xi – ксифипластрон. Штриховой линией обозначена предполагаемая форма энтопластрона.
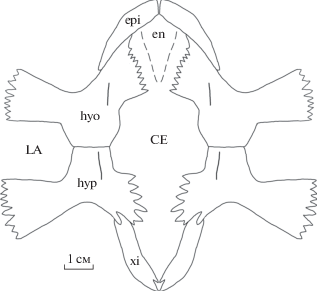
Гипопластроны представлены правым элементом, находящимся в сочленении с ксифипластроном и видимым с вентральной стороны, и изолированным левым элементом, видимым с дорсальной стороны (рис. 2, 3). На вентральной поверхности правого гипопластрона располагается продольный киль. На левом гипопластроне хорошо видны мелкие и густо посаженные медиальные и часть латеральных отростков.
Ксифипластроны представлены правым элементом, находящимся в сочленении с гипопластроном (рис. 2). Ксифипластрон узкий, в отличие от широких ксифипластронов Puppigerus camperi.
Позвонки представлены одним неопределенным шейным позвонком, у которого частично сохранились невральная дуга и постзигапофизы (рис. 2, 3). Длина невральной дуги больше, чем ширина. Постзигапофизы широко расставлены.
Лопатка (правая или левая; рис. 2, 3, 5, з) без дистальной части акромиального отростка. Гленоидная шейка имеется. Угол между скапулярным и акромиальными отростками составляет около 110°, что меньше, чем у Osonachelus decorata.
Плечевые кости представлены обоими элементами: на одном блоке породы видна вентральная сторона левой и дорсальная сторона правой плечевых костей, а на другом – отпечатки обеих костей (рис. 2, 3, 5, и, к). У левой кости видна часть поверхности диафиза, у правой кости видна поверхность дистальной части и области, примыкающей к латеральному отростку. Остальная часть поверхности костей разрушена. Медиальный отросток низкий. Латеральный отросток округлый, незначительно удален от головки. Диафиз шире, чем у других описанных экземпляров Eochelone. Ямка для крепления сухожилий mm. latissimus dorsi и teres major расположена близко к заднему краю. В дистальной части кости на дорсальной стороне видна эктэпикондилярная вырезка.
Помимо описанных костей, у данного экземпляра наблюдаются и другие элементы, в том числе не менее 16 костей, по-видимому, конечностей и их поясов (рис. 2, 3), однако их точное анатомическое положение невозможно определить из-за плохой сохранности.
ОБСУЖДЕНИЕ
Экз. ПИН, № 5136/1 относится к Chelonioidea на основании слабого развития окостенений костальных пластинок, узкого ксифипластрона и наличия гленоидной шейки лопатки (Данилов и др., 2017). Экз. ПИН, № 5136/1 относится к Cheloniidae на основании того, что прецентральный щиток занимает более половины ширины нухальной пластинки (Hirayama, 1998; Данилов и др., 2017).
Экз. ПИН, № 5136/1 относится к роду Eochelone на основании сочетания следующих признаков: лобные кости участвуют в формировании краев глазниц (контакт предлобной и заглазничной костей отсутствует), нижнечелюстной симфиз короткий, без лингвальных гребней, и лобно-теменной щиток сильно вытянут в заднем направлении (Casier, 1968; Данилов и др., 2017; Zvonok, Danilov, 2017).
Экз. ПИН, № 5136/1 отличается от других эоценовых хелониид следующими признаками: от “Argillochelys” africana – участием лобной кости в формировании края глазницы и отсутствием дополнительного лобно-теменного щитка; от Argillochelys spp. – отсутствием лингвального гребня на зубной кости; от Osonachelus decorata – наличием трех теменных щитков и меньшим углом между отростками лопатки; от Erquelinnesia gosseleti – отсутствием дополнительного лобно-теменного щитка и наличием лабиального гребня зубной кости; от Euclastes sp. из приабона США – наличием лабиального гребня зубной кости; от Puppigerus spp. – отсутствием вырезки в постеролатеральной части теменной кости, наличием лабиального гребня зубной кости и узким ксифипластроном (Zangerl, 1971; Moody, 1974; Tong et al., 2012; Звонок и др., 2013; Lapparent de Broin et al., 2014; Weems, Brown, 2017; Zvonok, Danilov, 2017).
Экз. ПИН, № 5136/1 имеет общий признак с Eochelone voltregana (он же отличительный от E. brabantica) – эпипластрон с широким и округленным гулярным выступом (рис. 5, б–г). Форма центральной фонтанели не пентагональная, однако сходна с таковой голотипа E. voltregana тем, что вытянута в переднем направлении по сравнению с латеральными фонтанелями (см. рис. 5, д–ж, 7). Другие признаки из диагноза E. voltregana (форма карапакса, наличие киля на карапаксе, форма и расположение щитков карапакса, контакт ребер с периферальными пластинками и форма ребер) у экз. ПИН, № 5136/1 неизвестны. Еще три признака могут быть ювенильными: это мелкие и густо посаженные отростки гио- и гипопластронов, широкая центральная фонтанель (состояния этих признаков отличаются от Eochelone voltregana и свойственны E. brabantica) и широкие плечевые кости (состояние признака совпадает с таковым у E. voltregana; рис. 2, 5, и–к, 7; Lapparent de Broin et al., 2018). На ювенильный возраст экз. ПИН, № 5136/1 указывают также его мелкий размер и слабо развитый (округленный) медиальный отросток плечевой кости (см. рис. 5, и, к; Zug et al., 1986). Перечисленные данные позволяют утверждать ювенильный возраст экз. ПИН, № 5136/1. Они не позволяют отнести данный экземпляр к E. voltregana или E. brabantica из-за того, что черты этих двух видов смешаны. Поэтому мы определяем экз. ПИН, № 5136/1 как Eochelone sp.
Полноценное сравнение экз. ПИН, № 5136/1 с Eochelone athersuchi и E. monstigris невозможно из-за того, что общие важные для диагностики детали морфологии не сохранились или не изображены в публикациях (Moody, 1980; Grant-Mackie et al., 2011).
Экз. ПИН, № 5136/1 обнаруживает сходство с “Allopleuron” qazaqstanense (экз. ZIN PH №№ 2/177, 8/177) в вытянутой в заднем направлении форме лобно-теменного щитка и отсутствии развитых лингвальных гребней в области симфиза зубных костей. Этот вид имеет слабое окостенение панциря, характерное для Eochelone, но строение верхнечелюстной кости экз. ZIN PH, № 6/177 указывает на то, что вторичное костное небо у него было развито, как минимум умеренно, а лингвальные гребни были высокими сзади, хотя и выклинивались спереди (Karl et al., 2012; Zvonok et al., 2015). Наличие умеренно развитого костного неба (которое достигает от 1/2 до 2/3 расстояния между передним краем черепа и fossa temporalis inferior) и лингвальных гребней не свойственны Eochelone и являются признаками рода Argillochelys (Zvonok, Danilov, 2017). Экз. ПИН, № 5136/1 не может быть отнесен к “Allopleuron” qazaqstanense, так как последний вид требует переописания и уточнения родовой принадлежности, а общие с ним признаки, прослеживаемые у экз. ПИН, № 5136/1 (вытянутый в заднем направлении лобно-теменной щиток и отсутствие лингвального гребня у симфиза зубных костей) известны также у типового вида Eochelone – E. brabantica (Casier, 1968).
Экз. ПИН, № 5136/1 расширяет представления о стратиграфическом и географическом распространении рода Eochelone в Европе. Прежде этот род был известен из лютета Бельгии и Украины, бартона Англии и приабона Испании, а теперь определен из бартона Северного Кавказа, что согласуется с тем, что в эоцене Южная Испания и Северный Кавказ были частью единого морского бассейна – океана Тетис (Данилов и др., 2017; Lapparent de Broin et al., 2018; Meulenkamp et al., 2000; рис. 8).
Рис. 8.
Палеогеографическая карта океана Тетис (без восточной оконечности) и прилегающих акваторий в среднем и позднем эоцене (по: Meulenkamp et al., 2000, с изменениями) с обозначением местонахождений черепах рода Eochelone: а – брюссельские пески, Бельгия; б – бассейн Гэмпшир, Англия; в – формация Вик-Манльеу, Испания; г – местонахождение Иково, Украина; д – местонахождение Горный Луч, Россия.
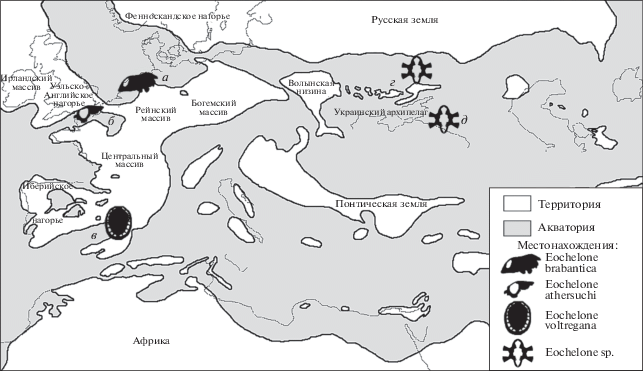
Актуальным остается вопрос о родовой принадлежности описанного из бартона Казахстана “Allopleuron” qazaqstanense, признаки которого имеются у экз. ПИН, № 5136/1. Изучение дополнительных материалов по этому виду, обладающему признаками родов Argillochelys и Eochelone, может прояснить его таксономическую принадлежность и уточнить связи между различными представителями указанных родов.
Список литературы
Аверьянов А.О. Новая морская черепаха (Testudines, Cheloniidae) из среднего эоцена Узбекистана // Палеонтол. журн. 2005. № 6. С. 73–76.
Банников А.Ф. Ископаемые позвоночные России и сопредельных стран. Ископаемые колючеперые рыбы (Teleostei, Acanthopterygii). М.: ГЕОС, 2010. LXI+244 с.
Банников А.Ф. Фауна рыб из бартона (средний эоцен) Северного Кавказа (р. Пшеха) // Интегративная палеонтология: перспективы развития для геологических целей. Матер. LXIII сессии Палеонтол. об-ва при РАН. СПб., 2017. С. 168-170.
Банников А.Ф. Фауна костистых рыб из бартона (верхи среднего эоцена) Северного Кавказа (р. Пшеха) // Тр. Палеонтол. об-ва. Том I / Ред. Рожнов С.В. М.: ПИН РАН, 2018. С. 7–14.
Данилов И.Г., Сыромятникова Е.В., Суханов В.Б. Подкласс Testudinata // Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Часть 4 / Ред. Лопатин А.В., Зеленков Н.В. М.: ГЕОС, 2017. С. 27–395.
Звонок Е.А. Строение черепа Puppigerus nessovi Averianov, 2005 (Reptilia, Testudines, Cheloniidae sensu lato) из среднеэоценовых отложений Украины // Геологічн. журн. 2013. № 1. С. 57–67.
Звонок Е.А., Данилов И.Г. Палеогеновые черепахи Крыма // Палеонтол. журн. 2019. № 1. С. 62–73.
Звонок Е.А., Данилов И.Г., Сыромятникова Е.В. и др. Предварительные результаты изучения черепах из палеогена Мангышлака (Казахстан) // Современная палеонтология: классические и новейшие методы. Тез. докл. VIII Всеросс. науч. школы молодых ученых-палеонтологов. М.: ПИН РАН, 2011. С. 20–21.
Звонок Е.А., Данилов И.Г., Сыромятникова Е.В. и др. Остатки морских черепах из местонахождения Иково (Луганская область, Украина; средний эоцен) // Палеонтол. журн. 2013. № 6. С. 47–58.
Звонок Е.А., Удовиченко Н.И., Братишко А.В. Новые данные о морфологии и систематическом положении морской черепахи Allopleuron qazaqstanense Karl et al. из среднего эоцена Казахстана // Палеонтол. журн. 2015. № 2. С. 66–77.
Малышкина Т.П., Гонсалес-Барба Х., Банников А.Ф. Находки зубов эласмобранхий в бартоне Северного Кавказа (Россия) и Крыма (Украина) // Палеонтол. журн. 2013. № 1. С. 76–80.
Averianov A.O. Review of Mesozoic and Cenozoic sea turtles from the former USSR // Russ. J. Herpetol. 2002. V. 9. № 2. P. 137–154.
Casier E. Le squelette céphalique de Eochelone brabantica L. Dollo, du Bruxellien (Lutétien inférieur) de Belgique, et sa comparaison avec celui de Chelone mydas Linné // Bull. Inst. Roy. Sci. Natur. Belg. 1968. № 44. P. 1–22.
Gaffney E.S. Comparative cranial morphology of recent and fossil turtles // Bull. Amer. Mus. Natur. Hist. 1979. V. 164. P. 65–376.
Grant-Mackie J.A., Hill J., Gill B.J. Two Eocene chelonioid turtles from Northland, New Zealand // New Zeal. J. Geol. Geophys. 2011. № 54. P. 181–194.
Hirayama R. Oldest known sea turtle // Nature. 1998. № 392. P. 705–708.
Karhu A. A new genus and species of the family Jungornithidae (Apodiformes) from the Late Eocene of the Northern Caucasus, with comments on the ancestry of hummingbirds // Smithson. Contrib. Paleobiol. 1999. № 89. P. 207–216.
Karl H.-V., Groning E., Brauckmann C. New materials of the giant sea turtle Allopleuron (Testudines: Chelonioidea) from the marine Late Cretaceous of Central Europe and the Palaeogene of Kazakhstan // Studia Geol. Salmanticensia. 2012. Vol. espec. № 9. P. 153–173.
Lapparent de Broin F. de, Murelaga X., Farrés F. et al. An exceptional cheloniid turtle, Osonachelus decorata nov. gen., nov. sp., from the Eocene (Bartonian) of Catalonia (Spain) // Geobios. 2014. V. 47. № 3. P. 111–132.
Lapparent de Broin F. de, Murelaga X., Pérez-García A. et al. The turtles from the upper Eocene, Osona County (Ebro Basin, Catalonia, Spain): new material and its faunistic and environmental context // Fossil Rec. 2018. № 21. P. 237–284.
Meulenkamp J.E., Sissingh W., Calvo P. et al. Late Lutetian (44–41 Ma) // Atlas Peri-Tethys, Palaeogeographical maps / Eds. Dercourt J., Gaetani M., Vrielynck B. et al. P.: CCGM/CGMW, 2000. P. 163–170.
Moody R.T.J. The taxonomy and morphology of Puppigerus camperi (Gray), an Eocene sea turtle from northern Europe // Bull. Brit. Mus. (Natur. Hist.) Geol. 1974. V. 25. P. 153–186.
Moody R.T.J. Notes on some European Palaeogene turtles // Tert. Res. 1980. V. 2. № 4. P. 161–168.
Owen R., Bell A. Reptilia of the Tertiary beds. Part 1. Chelonia // Monography of the Fossil Reptilia of the London Clay. L.: Palaeontogr. Soc., 1849. P. 1–76.
Tong H., Hirayama R. A new species of Argillochelys (Testudines: Cryptodira: Cheloniidae) from the Ouled Abdoun phosphate basin, Morocco // Bull. Soc. Géol. Fr. 2008. V. 179. № 6. P. 623–630.
Tong H., Hirayama R., Tabouelle J. Puppigerus camperi (Testudines: Cryptodira: Cheloniidae) from the Ypresian (Early Eocene) of Ouled Abdoun basin, Morocco // Bull. Soc. Géol. Fr. 2012. V. 183. № 6. P. 635–640.
Weems R., Brown K.M. More-complete remains of Procolpochelys charlestonensis (Oligocene, South Carolina), an occurrence of Euclastes (upper Eocene, South Carolina), and their bearing on Cenozoic pancheloniid sea turtle distribution and phylogeny // J. Paleontol. 2017. V. 91. № 6. P. 1228–1243.
Zangerl R. Two toxochelyid sea turtles from the Landenian sands of Erquelinnes (Hainaut) of Belgium // Mém. Inst. Roy. Sci. Natur. Belg. 1971. № 169. P. 1–32.
Zangerl R., Hendrickson L.P., Hendrickson J.R. A redescription of the Australian flatback sea turtle Natator depressus // Bishop Mus. Bull. Zool. 1988. № 1. P. 1–69.
Zvonok E.A., Danilov I.G. A revision of fossil turtles from the Kiev Clays (Ukraine, Middle Eocene) with comments on the history of the collection of fossil vertebrates of A.S. Rogovich // Proc. Zool. Inst. RAS. 2017. V. 321. № 4. P. 485–516.
Zug G.R., Wynn A.H., Ruckdeschel C. Age determination of loggerhead sea turtles, Caretta caretta, by incremental growth marks in the skeleton. Washington: Smithson. Inst. Press, 1986. 34 p.
Дополнительные материалы отсутствуют.
Инструменты
Палеонтологический журнал